Криптофиции
Эта статья нуждается в дополнительных цитатах для проверки . ( август 2010 г. ) |
| Криптофиты | |
|---|---|
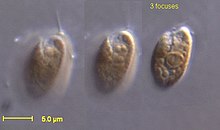 | |
| Родомонас Салина | |
| Научная классификация ( неопределенное место среди эукариот ) | |
| Домен: | |
| (без рейтинга): | |
| Суперкласс: | |
| Сорт: | Криптофиции |
| Заказы | |
| Синонимы | |
| |
Cryptophyceae — класс водорослей . [1] большинство из которых имеют пластиды . Известно около 230 видов. [2] они распространены в пресной воде, а также встречаются в морских и солоноватых средах обитания. Каждая клетка имеет размер около 10–50 мкм , уплощенную форму, с передней бороздкой или карманом. По краю кармана обычно имеются два слегка неравных жгутика .
Некоторые проявляют миксотрофию . [3]
Характеристики
[ редактировать ]
Криптофиты отличаются наличием характерных экструсом, называемых эжектосомами или эжектомосомами, которые состоят из двух соединенных спиральных лент, удерживаемых под натяжением. [4] Если клетки раздражаются механическим, химическим или световым стрессом, они разряжаются, перемещая клетку зигзагообразным курсом в сторону от возмущения. С карманом связаны крупные эектосомы, видимые под световым микроскопом; более мелкие встречаются под перипластом , окружением клеток, специфичных для криптофитов. [5] [6]
За исключением Chilomonas , у которого есть лейкопласты , криптофиты имеют один или два хлоропласта. Они содержат хлорофиллы а и с вместе с фикобилипротеинами и другими пигментами и различаются по цвету (от коричневого, красного до синевато-зеленого). Каждая окружена четырьмя мембранами, а находится редуцированное клеточное ядро, называемое нуклеоморфом между двумя средними . Это указывает на то, что пластида произошла от эукариотического симбионта, который, как показали генетические исследования, представлял собой красную водоросль . [7] Однако пластиды сильно отличаются от пластид красных водорослей: фикобилипротеины присутствуют, но только в просвете тилакоида и присутствуют только в виде фикоэритрина или фикоцианина . В случае «Родомонады» кристаллическая структура была определена до 1,63 Å; [8] и было показано, что альфа-субъединица не имеет никакого отношения ни к одному другому известному фикобилипротеину.
Некоторые криптофиты, такие как Cryptomonas , могут образовывать пальмеллоидные стадии, но легко покидают окружающую слизь и снова становятся свободноживущими жгутиконосцами. Некоторые виды Cryptomonas также могут образовывать неподвижные микробные цисты – стадии покоя с жесткими клеточными стенками, позволяющие выжить в неблагоприятных условиях. Жгутики криптофитов прикреплены параллельно друг другу и покрыты двудольными волосками, называемыми мастигонемами , которые образуются внутри эндоплазматической сети и транспортируются на поверхность клетки. Небольшие чешуйки также могут присутствовать на жгутиках и теле клетки. Митохондрии имеют плоские кристы , митоз открытый; половом размножении Сообщалось также о .
Группа разработала целый ряд светопоглощающих пигментов, называемых фикобилинами, которые способны поглощать длины волн, недоступные другим растениям или водорослям, что позволяет им жить в самых разных экологических нишах. [9] Способность, возникающая в результате эволюции уникального светособирающего антенного комплекса, полученного из двух реликтовых частей фикобилисомы красных водорослей , которая была полностью демонтирована в ходе эндосимбиотического процесса. [10]
Хотя криптофиты обычно считаются бесполыми, половое размножение действительно имеет место; обнаружены как гаплоидные, так и диплоидные формы. Два вида Teleaulax amphioxeia и Plagioselmis prolonga теперь считаются одним и тем же видом, где T. amphioxeia представляет собой диплоидную форму, а P. prolonga - гаплоидную форму. Диплоидная форма наиболее распространена, когда в воде больше питательных веществ. Две гаплоидные клетки часто сливаются, образуя диплоидную клетку, смешивая свои гены. [11]
Классификация
[ редактировать ]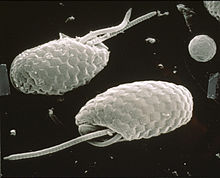

Первое упоминание о криптофитах, по-видимому, было сделано Кристианом Готфридом Эренбергом в 1831 году. [12] во время изучения Инфузории . Позднее ботаники отнесли их к отдельной группе водорослей , классу Cryptophyceae или отделу Cryptophyta, а зоологи отнесли к жгутиковым простейшим отряда Cryptomonadina. В некоторых классификациях криптомонады считались близкими родственниками динофлагеллят из -за их (казалось бы) схожей пигментации и группировались как Pyrrophyta . Имеются значительные доказательства того, что хлоропласты криптофитов тесно связаны с хлоропластами гетероконтов и гаптофитов , и эти три группы иногда объединяют в Chromista . Однако предположение о том, что сами организмы тесно связаны, не очень убедительно и они могли приобрести пластиды независимо. В настоящее время считается, что они являются членами клады Diaphoretickes и образуют вместе с Haptophyta группу Hacrobia . Парфри и др. и Бурки и др. поместил Cryptophyceae как сестринскую кладу зеленых водорослей . [13] [14]
Одна из предложенных группировок выглядит следующим образом: (1) Cryptomonas , (2) Chroomonas / Komma и Hemiselmis , (3) Rhodomonas / Rhinomonas / Storeatula , (4) Guillardia / Hanusia , (5) Geminigera / Plagioselmis / Teleaulax , (6) Proteomonas sulcata , (7) Falcomonas daucoides . [15]
- Класс Cryptophyceae Fritsch 1937 [Cryptomonadea Stein 1878, исправление. Шенихен 1925 ]
- Род Wysotzkia Lemmermann 1899 г.
- Род Ургорри Лаза-Мартинес 2012 г.
- Отряд Tetragonidiales Kristiansen 1992 г.
- Семейство Tetragonidiaceae Bourelly ex Silva1980.
- Род Bjornbergiella Bicudo 1966.
- Род Tetragonidium Pascher 1914.
- Семейство Tetragonidiaceae Bourelly ex Silva1980.
- Орден Pyrenomonadales Новарино и Лукас 1993 г.
- Семейство Baffinellaceae Daugbjerg & Norlin 2018 [16]
- Род Baffinella Norlin & Daugbjerg 2018.
- Семейство Chroomonadaceae Clay, Cugrens & Lee, 1999 г.
- Пол? Смитимастик Скворцов 1969 [ Смитиэлла Скворцов 1968 номин. Нелегально. ]
- Род Chroomonas Hansgirg 1885 г.
- Род Falcomonas Hill 1991 г.
- Род Hemiselmis Parke, 1949 г.
- Род Комма Хилл 1991 г.
- Род Нодеана Скворцов 1968.
- Род Planonephros Christensen 1978.
- Род Protochrysis Pascher 1911.
- Семейство Geminigeraceae Clay, Cugrens & Lee, 1999 г.
- Род Geminigera Hill 1991 г.
- Род Guillardia Hill & Wetherbee, 1990 г.
- Род Phia Özdikmen 2009 [ Hanusia Deane et al. 1998, а не Криппс 1989 ]
- Род Plagioselmis Butcher 1967 из Новарино, Lucas & Morrall 1994.
- Род Teleaulax Hill 1991.
- Семейство Pyrenomonadaceae Новарино и Лукас, 1993 г.
- Род Proteomonas Hill и Wetherbee, 1986 г.
- Род Rhinomonas Hill и Wetherbee, 1988 г.
- Род Rhodomonas Karsten 1898 [ Pyrenomonas Santore 1984 ]
- Род Storeatula Hill 1991.
- Семейство Baffinellaceae Daugbjerg & Norlin 2018 [16]
- Заказать Cryptomonadales Pascher 1913 г.
- Семья? Буцлиелловые Скворцов 1968
- Род Бучлиелла Скворцов 1968.
- Род Скворцовиелла Бурелли 1970.
- Семья ? Циатомонады Прингсхайм 1944 г.
- Род Cyathomonas de Fromentel 1874 г.
- Семья? Hilleaceae Пашер 1967
- Род Calkinsiella Скворцов 1969.
- Род Hillea Schiller 1925 г.
- Семья ? Pleuromastigaceae Bourrelly ex Silva 1980
- Пол? Опистостигма Шерффель 1911 г.
- Род Pleuromastix Scherffel 1912, не Namyslowski 1913.
- Род Xanthodiscus Schewiakoff 1892 г.
- Семейство Cryptomonadaceae Ehrenberg 1831 [Campylomonadaceae Clay, Kugrens & Lee 1999 ; Cryptochrysidaceae Pascher 1931 ]
- Пол? Чиломонас Эренберг 1831 г.
- Род? Протокриптохрис Скворцов 1969
- Род Cryptella Pascher 1929.
- Род Криптохлорис Шиллер 1925.
- Род Cryptochrysis Pascher 1911.
- Род Cryptomonas Ehrenberg 1832 [ Campylomonas Hill 1991 ]
- Род Cyanomastix Lackey 1936.
- Род Isoselmis Butcher 1967.
- Род Киселевия Скворцов 1969.
- Род Мейериелла Скворцов 1968 г.
- Род Olivamonas Скворцов 1969.
- Род Protocryptomonas Скворцов 1969 ex Bicudo 1989.
- Семья? Буцлиелловые Скворцов 1968
Ссылки
[ редактировать ]- ^ Хан Х, Арчибальд Дж. М. (май 2008 г.). «Латеральный перенос интронов в пластидном геноме криптофитов» . Нуклеиновые кислоты Рез . 36 (9): 3043–53. дои : 10.1093/нар/gkn095 . ПМК 2396441 . ПМИД 18397952 .
- ^ Cryptophyceae - :: База водорослей
- ^ «Криптофиты – криптомонады» . Архивировано из оригинала 10 июня 2011 г. Проверено 2 июня 2009 г.
- ^ Грэм, Ле; Грэм, Дж. М.; Уилкокс, Л.В. (2009). Водоросли (2-е изд.). Сан-Франциско, Калифорния: Бенджамин Каммингс (Пирсон). ISBN 9780321559654 .
- ^ Морролл, С.; Гринвуд, AD (1980). «Сравнение периодических подструктур трихоцист Cryptophyceae и Prasinophyceae». БиоСистемы . 12 (1–2): 71–83. Бибкод : 1980BiSys..12...71M . дои : 10.1016/0303-2647(80)90039-8 . ПМИД 6155157 .
- ^ Грим, Дж. Н.; Стэхелин, Луизиана (1984). «Эжектомомы жгутиковых Chilomonas paramecium - визуализация с помощью методов замораживания-перелома и выделения». Журнал протозоологии . 31 (2): 259–267. дои : 10.1111/j.1550-7408.1984.tb02957.x . ПМИД 6470985 .
- ^ Дуглас, С.; и др. (2002). «Сильно редуцированный геном порабощенного ядра водоросли» . Природа . 410 (6832): 1091–1096. Бибкод : 2001Natur.410.1091D . дои : 10.1038/35074092 . ПМИД 11323671 .
- ^ Уилк, К.; и др. (1999). «Эволюция светособирающего белка путем добавления новых субъединиц и перестановки консервативных элементов: кристаллическая структура криптофитного фикоэритрина с разрешением 1,63 Å» . ПНАС . 96 (16): 8901–8906. дои : 10.1073/pnas.96.16.8901 . ПМК 17705 . ПМИД 10430868 .
- ^ Каллиер, Вивиан (23 октября 2019 г.). «Этот тип водорослей поглощает больше света для фотосинтеза, чем другие растения» . Смитсоновский журнал .
- ^ Мичи, Кэтрин А.; Харроп, Стивен Дж.; Рэтбоун, Гарри В.; Уилк, Кристина Э.; Дэн, Чан Ин; Хоф-Эмден, Керстин; Хиллер, Роджер Г.; Грин, Беверли Р.; Курми, Пол М.Г. (март 2023 г.). «Молекулярные структуры раскрывают причину спектральных вариаций в белках светособирающих антенн криптофитов» . Белковая наука . 32 (3): e4586. дои : 10.1002/pro.4586 . ПМЦ 9951199 . ПМИД 36721353 .
- ^ Альтенбургер, А.; Блоссом, ОН; Гарсия-Куэтос, Л.; Якобсен, Х.Х.; Карстенсен, Дж.; Лундхольм, Н.; Хансен, П.Дж.; Моэструп, О.; Харагути, Л. (2020). «Диморфизм криптофитов - случай Teleaulax amphioxeia / Plagioselmis prolonga и его экологические последствия» . Достижения науки . 6 (37). Бибкод : 2020SciA....6.1611A . дои : 10.1126/sciadv.abb1611 . ПМЦ 7486100 . ПМИД 32917704 .
- ^ Новарино, Г. (2012). «Таксономия криптомонад в 21 веке: первые 200 лет» . Психологические отчеты: современные достижения в таксономии водорослей и ее применениях: филогенетическая, экологическая и прикладная перспектива : 19–52 . Проверено 16 октября 2018 г.
- ^ Парфри, Лаура Вегенер ; Лар, Дэниел Дж.Г.; Нолл, Эндрю Х.; Кац, Лаура А. (16 августа 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–13629. Бибкод : 2011PNAS..10813624P . дои : 10.1073/pnas.1110633108 . ПМК 3158185 . ПМИД 21810989 .
- ^ Бурки, Фабьен; Каплан, Майя; Тихоненков Денис Владимирович; Златогурский, Василий; Минь, Буй Куанг; Радайкина Людмила Владимировна; Смирнов, Алексей; Мыльников Александр П.; Килинг, Патрик Дж. (27 января 2016 г.). «Распутывание ранней диверсификации эукариот: филогеномное исследование эволюционного происхождения Centrohelida, Haptophyta и Cryptista» . Учеб. Р. Сок. Б. 283 (1823): 20152802. doi : 10.1098/rspb.2015.2802 . ISSN 0962-8452 . ПМК 4795036 . ПМИД 26817772 .
- ^ «Криптомонады» . Проверено 24 июня 2009 г.
- ^ Даугбьерг, Нильс; Норлин, Андреас; Лавджой, Конни (25 июля 2018 г.). « Baffinella frigidus gen. et sp. nov. (Baffinellaceae fam. nov., Cryptophyceae) из Баффинового залива: морфология, пигментный профиль, филогения и реакция скорости роста на три абиотических фактора» (PDF) . Дж. Фикол . 54 (5): 665–680. дои : 10.1111/jpy.12766 . ISSN 1529-8817 . ПМИД 30043990 . S2CID 51718360 . Архивировано из оригинала (PDF) 12 октября 2018 г. Проверено 26 мая 2019 г.