Раскопать
| Раскопать Временной диапазон: | |
|---|---|
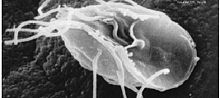 | |
| Лямблии лямблии , паразит-выпускник | |
| Научная классификация (устарело как парафилетический) | |
| Домен: | Эукариоты |
| (без рейтинга): | Раскопать ( Кавальер-Смит ), 2002 г. |
| Фила и классы | |
см. текст | |

Excavata — обширная и разнообразная, но парафилетическая группа одноклеточных эукариот . [1] [2] Группа была впервые предложена Симпсоном и Паттерсоном в 1999 году. [3] [4] а название латинизировано и присвоено звание Томасом Кавалье-Смитом в 2002 году. Оно содержит множество свободноживущих и симбиотических простейших, а также включает некоторых важных паразитов человека, таких как лямблии и трихомонады . [5] Раньше считалось, что раскопки включены в ныне устаревшее королевство Протиста . [6] Их отличали от других линий на основе электронно-микроскопической информации о том, как устроены клетки (они имеют отличительную ультраструктурную идентичность ). [4] Они считаются базальной линией жгутиковых . [7]
На основе филогеномного анализа было показано, что эта группа включает три широко разделенные группы эукариот: дискобиды, метамонады и малавимонады. [8] [9] [10] [11] Ниже представлен современный взгляд на состав раскопок, свидетельствующий о том, что группа является парафилетической. За исключением некоторых Euglenozoa , все они не фотосинтезируют .
Характеристики
[ редактировать ]Большинство раскопок представляют собой одноклеточные гетеротрофные жгутиконосцы. Лишь некоторые эвгленозои являются фотосинтезирующими. У некоторых (особенно анаэробных кишечных паразитов) количество митохондрий значительно уменьшено. [5] В некоторых раскопках отсутствуют «классические» митохондрии , и они называются «амитохондриатами», хотя большинство из них сохраняют митохондриальную органеллу в сильно модифицированной форме (например, гидрогеносому или митосому ). У тех, у кого есть митохондрии, митохондриальные кристы могут быть трубчатыми, дискоидными или, в некоторых случаях, пластинчатыми. Большинство раскопок имеют два, четыре и более жгутиков . [4] Многие из них имеют заметную вентральную питающую бороздку с характерной ультраструктурой , поддерживаемой микротрубочками — «выкопанный» вид этой бороздки дал организмам свое название. [3] [6] Однако различные группы, у которых отсутствуют эти признаки, считаются производными на основе генетических данных (в первую очередь филогенетических деревьев молекулярных последовательностей). [6]
Слизевики Acrasidae — единственные находки, демонстрирующие ограниченную многоклеточность. Как и другие клеточные слизевики , они большую часть жизни живут в виде отдельных клеток, но иногда собираются в более крупные скопления.
Предлагаемая группа
[ редактировать ]Отношения с раскопками всегда были неопределенными, что позволяет предположить, что они не являются монофилетической группой. [12] Филогенетический анализ часто не относит малавимонады к одной ветви с другими Excavata. [13]
Считалось, что раскопки включают в себя несколько групп:
| Королевство/Супертип | Включенные таксоны | Репрезентативные роды (примеры) | Описание |
|---|---|---|---|
| Дискоба или JEH или Эозоа | Цукубеа | Цукубамонас | |
| эвгленозоа | Эвглена , Трипаносома | Многие важные паразиты, одна большая группа с пластидами (хлоропластами). | |
| Гетеролобозея (Percolozoa) | Неглерия , Акрасис | Большинство чередуют жгутиковые и амебоидные формы. | |
| Якобинец | Якоба , Реклиномонас | Свободноживущие, иногда лорчатые жгутиконосцы, с очень богатыми генами митохондриальными геномами. | |
| Метамонада или POD | Преаксостила | Оксимонады , Тримастикс | Амитохондриальные жгутиковые, либо свободноживущие ( Trimastix , Paratrimastix ), либо живущие в задних кишках насекомых. |
| блудливый | Лямблии , Карпедиемонас | Амитохондрии, преимущественно симбиоты и паразиты животных. | |
| Парабазалия | Трихомонада | Амитохондриальные жгутиковые, обычно кишечные комменсалы насекомых. Некоторые патогены человека. | |
| Анаэрамеба | Анаэрамёба была ленивой | Анаэробные протисты с гидрогеносомами вместо митохондрий. | |
| Неолука | Малавимонадида | Малавимонас |
Дискоба или клада JEH
[ редактировать ]Euglenozoa и Heterolobosea (Percolozoa) или Eozoa (по названию Кавальера-Смита). [14] ), по-видимому, являются особенно близкими родственниками и объединены наличием дискоидных крист внутри митохондрий (Superphylum Discicristata ). Близкие отношения были показаны между Discicristata и Jakobida . [15] последние имели трубчатые кристы, как и большинство других протистов, и поэтому были объединены под названием таксона Discoba , которое было предложено для этой, очевидно, монофилетической группы. [1]
Метамонады
[ редактировать ]Метамонады необычны тем, что не имеют классических митохондрий — вместо этого у них есть гидрогеносомы , митосомы или неохарактеризованные органеллы. Сообщается, что оксимонада Monocercomonoides полностью утратила гомологичные органеллы. Существуют конкурирующие объяснения. [16] [17]
Малавимонады
[ редактировать ]были малавимонады Было предложено, чтобы членами Excavata из-за их типичной морфологии раскопок и филогенетического родства с другими группами раскопок в некоторых молекулярных филогениях. Однако их положение среди эукариот остается неясным. [2]
Анциромонады
[ редактировать ]Анциромонады представляют собой небольшие свободноживущие клетки с узкой продольной бороздкой на одной стороне клетки. Борозда анциромонад не используется для «суспензионного питания», в отличие от «типичных раскопок» (например, малавимонады, якобиды, тримастиксы , карпедиемоны , киперферлии и др.). Вместо этого анциромонады захватывают прокариотов, прикрепленных к поверхностям. Филогенетическое расположение анциромонад плохо изучено (на 2020 год), однако некоторые филогенетические анализы относят их к близким родственникам малавимонад. [9]
Эволюция
[ редактировать ]Происхождение эукариотов
[ редактировать ]Традиционное объяснение происхождения эукариот состоит в том, что хеймдалархеи или другие археи приобрели альфапротеобактерию. [18] как эндосимбионт , и что он стал митохондрией , органеллой, обеспечивающей окислительное дыхание эукариотической клетки. [19]
Вместо этого Цезарь аль Джвари и Сандра Балдауф утверждают, что эукариоты, возможно, начались с эндосимбиоза Deltaproteobacterium или Gammaproteobacterium , что объясняет необъяснимое иначе присутствие анаэробных бактериальных ферментов в Metamonada. Сестра Preaxostyla в составе Метамонады представляет собой остальную часть эукариотов, которые приобрели альфапротеобактерию. По их сценарию, гидрогеносома и митосома , которые традиционно считаются «органеллами, происходящими из митохондрий», предшествуют митохондриям и вместо этого происходят от более ранней симбиотической бактерии. [17]
Филогения
[ редактировать ]В 2023 году, используя молекулярно-филогенетический анализ 186 таксонов, Аль Джвари и Балдауф предложили филогенетическое древо, в котором метамонада Parabasalia является базальными эукариотами. Дискоба и остальные эукариоты, по-видимому, возникли как сестринский таксон Preaxostyla, включив одну альфапротеобактерию в виде митохондрий в результате эндосимбиоза. Таким образом, Fornicata более тесно связаны, например, с животными, чем с Parabasalia. Остальные эукариоты появились в Раскопках как сестры Дискобы; поскольку они находятся в одной кладе, но кладистически еще не считаются частью Excavata, Excavata в этом анализе являются в высшей степени парафилетическими. [17]
| "Экскавата" | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Anaeramoeba . связана с Parabasalia, но может оказаться более базальной, поскольку корень дерева часто трудно определить [20]
См. также
[ редактировать ]Галерея
[ редактировать ]
 Trypanosoma brucei (Euglenozoa: Kinetoplastida )
Trypanosoma brucei (Euglenozoa: Kinetoplastida ) Бодо сп. (Эвгленозоа: кинетопластида)
Бодо сп. (Эвгленозоа: кинетопластида) Перколомонас сп. (Перколозоа)
Перколомонас сп. (Перколозоа) Стефанопогон сп. (Перколозоа)
Стефанопогон сп. (Перколозоа)
 Акразис розовый (Percolozoa: Heterolobosea)
Акразис розовый (Percolozoa: Heterolobosea) Якобиды ( Jacobida )
Якобиды ( Jacobida )

 Лямблии сп. (Метамонада: Fornicata: Diplomonadida )
Лямблии сп. (Метамонада: Fornicata: Diplomonadida )











Ссылки
[ редактировать ]- ^ Перейти обратно: а б Хампл, Владимир; Обнимаю, Лора; Ли, Джессика В.; и др. (2009). «Филогеномный анализ подтверждает монофилию Excavata и разрешает отношения между «супергруппами» эукариот » . ПНАС . 106 (10): 3859–3864. Бибкод : 2009PNAS..106.3859H . дои : 10.1073/pnas.0807880106 . ПМК 2656170 . ПМИД 19237557 .
- ^ Перейти обратно: а б Симпсон, Аластер ГБ; Инагаки, Юдзи; Роджер, Эндрю Дж. (2006). «Комплексная мультигенная филогения раскопанных протистов раскрывает эволюционное положение «примитивных» эукариот» . Молекулярная биология и эволюция . 23 (3): 615–625. дои : 10.1093/molbev/msj068 . ПМИД 16308337 .
- ^ Перейти обратно: а б Симпсон, Аластер ГБ; Паттерсон, Дэвид Дж. (декабрь 1999 г.). «Ультраструктура Carpediemonas membranifera (Eukaryota) со ссылкой на« гипотезу раскопок » ». Европейский журнал протистологии . 35 (4): 353–370. дои : 10.1016/S0932-4739(99)80044-3 .
- ^ Перейти обратно: а б с Симпсон, Аластер ГБ (ноябрь 2003 г.). «Цитоскелетная организация, филогенетическое сходство и систематика спорного таксона Excavata (Eukaryota)» . Международный журнал систематической и эволюционной микробиологии . 53 (6): 1759–1777. дои : 10.1099/ijs.0.02578-0 . ПМИД 14657103 .
- ^ Перейти обратно: а б Докинз, Ричард ; Вонг, Ян (2016). Рассказ предка . Хоутон Миффлин Харкорт. ISBN 978-0544859937 .
- ^ Перейти обратно: а б с Кавальер-Смит, Томас (2002). «Фаготрофное происхождение эукариот и филогенетическая классификация простейших». Международный журнал систематической и эволюционной микробиологии . 52 (2): 297–354. дои : 10.1099/00207713-52-2-297 . ПМИД 11931142 .
- ^ Доусон, Скотт С.; Паредес, Александр Р. (2013). «Альтернативные цитоскелетные ландшафты: новизна цитоскелета и эволюция простейших с базальными выемками» . Современное мнение в области клеточной биологии . 25 (1): 134–141. дои : 10.1016/j.ceb.2012.11.005 . ПМЦ 4927265 . ПМИД 23312067 .
- ^ Бурки, Фабьен; Роджер, Эндрю Дж.; Браун, Мэтью В.; и др. (январь 2020 г.). «Новое дерево эукариотов» . Тенденции в экологии и эволюции . 35 (1): 43–55. дои : 10.1016/j.tree.2019.08.008 . ПМИД 31606140 . S2CID 204545629 .
- ^ Перейти обратно: а б Браун, Мэтью В.; Хейсс, Аарон А.; Камикава, Рёма; и др. (19 января 2018 г.). «Филогеномика помещает сиротские линии протистанов в новую супергруппу эукариот» . Геномная биология и эволюция . 10 (2): 427–433. дои : 10.1093/gbe/evy014 . ПМЦ 5793813 . ПМИД 29360967 .
- ^ Хейсс, Аарон А.; Колиско, Мартин; Экелунд, Флеминг; и др. (4 апреля 2018 г.). «Комбинированное морфологическое и филогеномическое повторное исследование малавимонад, важнейшего таксона для установления эволюционной истории эукариот» . Королевское общество открытой науки . 5 (4): 171707. Бибкод : 2018RSOS....571707H . дои : 10.1098/rsos.171707 . ПМЦ 5936906 . ПМИД 29765641 .
- ^ Килинг, Патрик Дж.; Бурки, Фабьен (19 августа 2019 г.). «Прогресс к Древу эукариотов» . Современная биология . 29 (16): Р808–Р817. Бибкод : 2019CBio...29.R808K . дои : 10.1016/j.cub.2019.07.031 . ПМИД 31430481 .
- ^ Лаура Вегенер Парфри ; Эрика Барберо; Элиз Лассер; Мика Данторн; Дебашиш Бхаттачарья; Дэвид Дж. Паттерсон ; Лаура А. Кац (декабрь 2006 г.). «Оценка поддержки текущей классификации эукариотического разнообразия» . ПЛОС Генетика . 2 (12): е220. doi : 10.1371/JOURNAL.PGEN.0020220 . ISSN 1553-7390 . ПМЦ 1713255 . ПМИД 17194223 . Викиданные Q21090155 .
- ^ Тайс, Александр К.; Жихала, Дэвид; Панек, Томаш; и др. (2021). «PhyloFisher: Филогеномный пакет для разрешения эукариотических отношений» . ПЛОС Биология . 19 (8): e3001365. дои : 10.1371/journal.pbio.3001365 . ПМЦ 8345874 . ПМИД 34358228 .
- ^ Кавальер-Смит, Томас (23 декабря 2009 г.). «Царства Protozoa и Chromista и эозойный корень эукариотического дерева» . Письма по биологии . 6 (3). Королевское общество: 342–345. дои : 10.1098/rsbl.2009.0948 . ISSN 1744-9561 . ПМК 2880060 . ПМИД 20031978 .
- ^ Родригес-Эспелета, Найара; Бринкманн, Хеннер; Бургер, Гертрауд; и др. (2007). «К разгадке эукариотического дерева: филогенетическое положение якобидов и церкозойных» . Современная биология . 17 (16): 1420–1425. Бибкод : 2007CBio...17.1420R . дои : 10.1016/j.cub.2007.07.036 . ПМИД 17689961 .
- ^ Буи, Элизабет Т.; Брэдли, Питер Дж.; Джонсон, Патрисия Дж. (3 сентября 1996 г.). «Общее эволюционное происхождение митохондрий и гидрогеносом» . Труды Национальной академии наук . 93 (18): 9651–9656. Бибкод : 1996PNAS...93.9651B . дои : 10.1073/pnas.93.18.9651 . ISSN 0027-8424 . ПМЦ 38483 . ПМИД 8790385 .
- ^ Перейти обратно: а б с Аль Джвари, Цезарь; Балдауф, Сандра Л. (28 апреля 2023 г.). «Раскопанный корень древа жизни эукариот» . Достижения науки . 9 (17): eade4973. Бибкод : 2023SciA....9E4973A . дои : 10.1126/sciadv.ade4973 . ISSN 2375-2548 . ПМЦ 10146883 . ПМИД 37115919 .
- ^ Триа, ФДК; Брюкнер Дж.; Скейо, Дж.; Ксавье, Джей Си; Капуст, Н.; Кнопп, М.; и др. (7 мая 2021 г.). «Дупликация генов прослеживает митохондрии до возникновения сложности эукариот» . Геномная биология и эволюция . 13 (5). дои : 10.1093/gbe/evab055 . ПМЦ 8175051 . ПМИД 33739376 .
- ^ Перейти обратно: а б Эме, Лаура; Тамарит, Дэниел; Касерес, Ева Ф.; и др. (09.03.2023). «Вывод и реконструкция хеймдаллархейного происхождения эукариот» . Природа . 618 (7967): 992–999. bioRxiv 10.1101/2023.03.07.531504 . дои : 10.1038/s41586-023-06186-2 . ПМЦ 10307638 . ПМИД 37316666 .
- ^ Лестница, Кортни В.; Таборский, Петр; Саломаки, Эрик Д.; и др. (декабрь 2021 г.). «Анаэрмебы — это дивергентная линия эукариот, которая проливает свет на переход от анаэробных митохондрий к гидрогеносомам» . Современная биология . 31 (24): 5605–5612.e5. Бибкод : 2021CBio...31E5605S . дои : 10.1016/j.cub.2021.10.010 . ISSN 0960-9822 . ПМИД 34710348 . S2CID 240054026 .