Эукариогенез
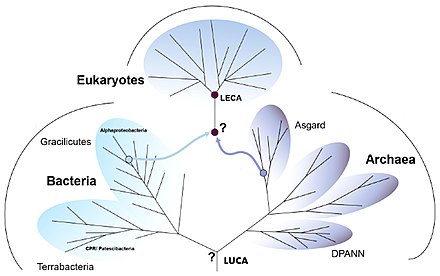
Эукариогенез , процесс, в результате которого возникли эукариотическая клетка и родословная, является важной вехой в эволюции жизни, поскольку эукариоты включают в себя все сложные клетки и почти все многоклеточные организмы. Широко распространено мнение, что этот процесс включал симбиогенез , в котором археон и бактерия объединились, чтобы создать первого общего предка эукариот ( FECA ). Эта клетка имела новый уровень сложности и возможностей, с ядром, по крайней мере, одной центриолью и ресничкой , факультативно аэробными митохондриями , полом ( мейоз и сингамия ), спящей кистой с клеточной стенкой из хитина и/или целлюлозы и пероксисомами . Он превратился в популяцию одноклеточных организмов, в которую входил последний общий предок эукариот ( LECA ), приобретая по пути способности, хотя последовательность этапов оспаривается и, возможно, началась не с симбиогенеза. В свою очередь, LECA породила коронную группу эукариотов , содержащую предков животных , грибов , растений и разнообразных одноклеточных организмов.
Контекст
[ редактировать ]Жизнь возникла на Земле, когда она достаточно остыла, чтобы образовались океаны. Последним универсальным общим предком (LUCA) был организм , имевший рибосомы и генетический код ; он жил около 4 миллиардов лет назад. Это дало начало двум основным ветвям прокариотической жизни: бактериям и археям . Среди этих мелкоклеточных, быстро делящихся предков возникли эукариоты с гораздо более крупными клетками, ядрами и своеобразной биохимией. [1] [2] Эукариоты образуют домен , который содержит все сложные клетки и большинство типов многоклеточных организмов , включая животных , растения и грибы . [3] [4]
Симбиогенез
[ редактировать ]
Согласно теории симбиогенеза (также известной как эндосимбиотическая теория ), отстаиваемой Линн Маргулис , член археи получил в качестве компонента бактериальную клетку. Архейная клетка входила в группу Асгарда . Бактерия была одной из альфапротеобактерий , которая обладала способностью использовать кислород при дыхании. Это позволило ему – и включавшим его архейным клеткам – выжить в присутствии кислорода, который был ядовитым для других организмов, приспособленных к восстановительным условиям. эукариотической клетки Эндосимбиотические бактерии стали митохондриями , обеспечивающими большую часть энергии клетки. [1] [5] Линн Маргулис и коллеги предположили, что клетка также приобрела бактерию Spirochaete в качестве симбионта, обеспечивающего клеточный скелет из микротрубочек и способность двигаться, в том числе способность объединять хромосомы в два набора во время митоза , деления клетки. [6] Совсем недавно архей Асгарда был идентифицирован как принадлежащий к Heimdallarchaeota . [7]
Последний общий предок эукариот (LECA)
[ редактировать ]Последний общий предок эукариот (LECA) — гипотетический последний общий предок всех ныне живущих эукариот, живший около 2 миллиардов лет назад. [3] [4] и, скорее всего, представлял собой биологическую популяцию . [8] Считается, что это был протист с ядром, по крайней мере, одной центриолью и ресничкой , факультативно аэробными митохондриями, полом ( мейоз и сингамия ), спящей кистой с клеточной стенкой из хитина и/или целлюлозы и пероксисомами . [9] [10]
Было высказано предположение, что LECA питается путем фагоцитоза , поглощая другие организмы. [9] [10] Однако в 2022 году Нико Бремер и его коллеги подтвердили, что LECA имеет митохондрии, и заявили, что у него несколько ядер, но оспаривали его фаготрофность. Это означало бы, что способность, обнаруженная у многих эукариот, поглощать материалы, развилась позже, а не была приобретена сначала и затем использована для поглощения альфа-протеобактерий, которые стали митохондриями. [11]
LECA описывается как имеющая «впечатляющую клеточную сложность». [12] Его камера была разделена на отсеки. [12] По-видимому, он унаследовал набор белков эндосомального сортировочного комплекса , которые позволяют ремоделировать мембраны, включая отщипывание везикул с образованием эндосом . [13] Его аппараты для транскрипции ДНК в РНК , а затем для перевода РНК в белки были разделены, что позволило осуществлять обширный процессинг РНК и усложнить экспрессию генов. [14] У него были механизмы для перестановки генетического материала и, возможно, для управления собственной способностью к развитию . Все это дало LECA «убедительную группу избирательных преимуществ». [12]
Эукариотический секс
[ редактировать ]Секс у эукариот — сложный процесс, состоящий из мейоза и оплодотворения , который может быть связан с размножением . [15] Часто и Роджер [16] на основе филогенетического анализа предположил, что факультативный пол, вероятно, присутствовал у общего предка всех эукариот. На ранних этапах эволюции эукариот, около 2 миллиардов лет назад, организмы нуждались в решении основной проблемы, заключающейся в том, что окислительный метаболизм высвобождает активные формы кислорода , которые повреждают генетический материал, ДНК . [15] Эукариотический секс обеспечивает процесс гомологичной рекомбинации во время мейоза, позволяющий использовать информационную избыточность для восстановления таких повреждений ДНК . [15]
Сценарии
[ редактировать ]Биологи предложили несколько сценариев создания эукариотов. Хотя существует широкое согласие в том, что LECA должно было иметь ядро, митохондрии и внутренние мембраны, порядок их приобретения оспаривается. В синтрофной модели первый общий предок эукариот (FECA, около 2,2 млрд лет назад ) приобрел митохондрии, затем мембраны, а затем ядро. В фаготрофной модели у него появилось ядро, затем мембраны, затем митохондрии. В ходе более сложного процесса он в короткие сроки получил все три, а затем и другие возможности. Были предложены и другие модели. Что бы ни случилось, должно было быть создано множество линий, но LECA либо вытеснила других линий, либо объединилась с другими линиями, чтобы сформировать единую точку происхождения эукариотов. [12] Ник Лейн и Уильям Мартин утверждали, что митохондрии были на первом месте, поскольку энергия была ограничивающим фактором размера прокариотической клетки. [17] Фаготрофная модель предполагает способность поглощать пищу, позволяя клетке поглощать аэробную бактерию, ставшую митохондрией. [12]
Юджин Кунин и другие, отмечая, что археи имеют много общих черт с эукариотами, утверждают, что рудиментарные черты эукариот, такие как выстланные мембранами компартменты, были приобретены до того, как эндосимбиоз добавил митохондрии к ранней эукариотической клетке, в то время как клеточная стенка была утрачена. Точно так же приобретение митохондрий не следует рассматривать как завершение процесса, поскольку после или во время эндосимбиотического обмена необходимо было создать новые сложные семейства генов. Таким образом, от FECA к LECA, мы можем думать об организмах, которые можно рассматривать как протоэукариоты. В конце процесса LECA уже представлял собой сложный организм с наличием семейств белков, участвующих в клеточной компартментализации. [18] [19]
Диверсификация: коронные эукариоты
[ редактировать ]эукариотов В свою очередь, LECA дала начало коронной группе , в которую вошли предки животных , грибов , растений и разнообразных одноклеточных организмов с новыми возможностями и сложностью эукариотической клетки. [20] [21] Одиночные клетки без клеточных стенок хрупкие и имеют низкую вероятность окаменения . Если они окаменели, у них мало особенностей, позволяющих четко отличить их от прокариот: размер, морфологическая сложность и (в конечном итоге) многоклеточность . Ранние окаменелости эукариот, относящиеся к позднему палеопротерозою , включают микроокаменелости акритархов с относительно прочными богато украшенными углеродистыми пузырьками Tappania от 1,63 млрд лет назад и Shuiyousphaeridium от 1,8 млрд лет назад. [21]
Ссылки
[ редактировать ]- ^ Jump up to: а б с МакГрат, Кейси (31 мая 2022 г.). «Основной момент: раскрытие истоков LUCA и LECA на Древе Жизни» . Геномная биология и эволюция . 14 (6): evac072. дои : 10.1093/gbe/evac072 . ПМЦ 9168435 .
- ^ Вайс, Мэдлин С.; Соуза, Флорида; Мрнявац, Н.; и др. (2016). «Физиология и среда обитания последнего всеобщего общего предка» (PDF) . Природная микробиология . 1 (9): 16116. doi : 10.1038/nmicrobiol.2016.116 . ПМИД 27562259 . S2CID 2997255 .
- ^ Jump up to: а б Габальдон, Т. (октябрь 2021 г.). «Происхождение и ранняя эволюция эукариотической клетки». Ежегодный обзор микробиологии . 75 (1): 631–647. doi : 10.1146/annurev-micro-090817-062213 . ПМИД 34343017 . S2CID 236916203 .
- ^ Jump up to: а б Вёзе, ЧР ; Кандлер, Отто ; Уилис, Марк Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Jump up to: а б Латорре, А.; Дурбан, А; Мойя, А.; Перето, Дж. (2011). «Роль симбиоза в эволюции эукариот» . В Гарго, Мюриэль; Лопес-Гарсиа, «Очищение»; Мартин Х. (ред.). Происхождение и эволюция жизни: астробиологическая перспектива . Кембридж: Издательство Кембриджского университета. стр. 326–339. ISBN 978-0-521-76131-4 . Архивировано из оригинала 24 марта 2019 года . Проверено 27 августа 2017 г.
- ^ Маргулис, Линн ; Чепмен, Майкл; Герреро, Рикардо; Холл, Джон (29 августа 2006 г.). «Последний общий предок эукариот (LECA): приобретение подвижности цитоскелета у аэротолерантных спирохет в протерозойском эоне» . Труды Национальной академии наук . 103 (35): 13080–13085. Бибкод : 2006PNAS..10313080M . дои : 10.1073/pnas.0604985103 . ПМЦ 1559756 . ПМИД 16938841 .
- ^ Уильямс, Том А.; Кокс, Саймон Дж.; Фостер, Питер Г.; Сёллёси, Гергеи Дж.; Эмбли, Т. Мартин (9 декабря 2019 г.). «Филогеномика обеспечивает надежную поддержку двухдоменного древа жизни» . Экология и эволюция природы . 4 (1): 138–147. Бибкод : 2019NatEE...4..138W . дои : 10.1038/s41559-019-1040-x . ПМК 6942926 . ПМИД 31819234 .
- ^ О'Мэлли, Морин А.; Леже, Мишель М.; Уайдман, Джереми Г.; Руис-Трилло, Иньяки (18 февраля 2019 г.). «Представления о последнем общем предке эукариот». Экология и эволюция природы . 3 (3): 338–344. Бибкод : 2019NatEE...3..338O . дои : 10.1038/s41559-019-0796-3 . hdl : 10261/201794 . ПМИД 30778187 . S2CID 256718457 .
- ^ Jump up to: а б Леандер, Б.С. (май 2020 г.). «Хищные протисты» . Современная биология . 30 (10): 510–516 р. дои : 10.1016/j.cub.2020.03.052 . ПМИД 32428491 . S2CID 218710816 .
- ^ Jump up to: а б Штрассерт, Юрген Ф.Х.; Ирисарри, Икер; Уильямс, Том А.; Бурки, Фабьен (25 марта 2021 г.). «Молекулярная временная шкала эволюции эукариот с последствиями для происхождения пластид, полученных из красных водорослей» . Природные коммуникации . 12 (1): 1879. Бибкод : 2021NatCo..12.1879S . дои : 10.1038/s41467-021-22044-z . ПМЦ 7994803 . ПМИД 33767194 .
- ^ Бремер, Нико; Триа, Фернандо Д.К.; Скейо, Йосип; Гарг, Шрирам Г.; Мартин, Уильям Ф. (31 мая 2022 г.). «Реконструкция наследственного состояния прослеживает митохондрии, но не фагоцитоз до последнего общего предка эукариот» . Геномная биология и эволюция . 14 (6). дои : 10.1093/gbe/evac079 . ПМЦ 9185374 . ПМИД 35642316 .
- ^ Jump up to: а б с д и Куманду, В. Лила; Уикстед, Билл; Джинджер, Майкл Л.; ван дер Гизен, Марк; Дакс, Джоэл Б.; Филд, Марк К. (2013). «Молекулярная палеонтология и сложность последнего общего предка эукариот» . Критические обзоры по биохимии и молекулярной биологии . 48 (4): 373–396. дои : 10.3109/10409238.2013.821444 . ПМЦ 3791482 . ПМИД 23895660 .
- ^ Макарова Кира С.; Ютин, Наталья; Белл, Стивен Д.; Кунин, Евгений В. (6 сентября 2010 г.). «Эволюция разнообразных систем деления клеток и образования пузырьков у архей» . Обзоры природы Микробиология . 8 (10): 731–741. дои : 10.1038/nrmicro2406 . ПМК 3293450 . ПМИД 20818414 .
- ^ Мартин, Уильям; Кунин, Евгений В. (2006). «Интроны и происхождение компартментализации ядра и цитозоля». Природа . 440 (7080): 41–45. дои : 10.1038/nature04531 . ISSN 0028-0836 .
- ^ Jump up to: а б с Хорандл, Э.; Шпейер, Д. (7 февраля 2018 г.). «Как кислород породил эукариотический пол» . Труды Королевского общества B: Биологические науки . 285 (1872). Королевское общество. дои : 10.1098/rspb.2017.2706 . ПМК 5829205 . ПМИД 29436502 .
- ^ Дакс, Дж.; Роджер, Эй Джей (1999). «Первая половая линия и актуальность факультативного секса». Журнал молекулярной эволюции . 48 (6): 779–783. Бибкод : 1999JMolE..48..779D . дои : 10.1007/pl00013156 . ПМИД 10229582 . S2CID 9441768 .
- ^ Лейн, Ник ; Мартин, Уильям Ф. (2010). «Энергетика сложности генома». Природа . 467 (7318): 929–934. Бибкод : 2010Natur.467..929L . дои : 10.1038/nature09486 . ПМИД 20962839 . S2CID 17086117 .
- ^ Кунин, Евгений В. (март 2005 г.). «Невероятно расширяющийся предок эукариотов» . Клетка . 140 (5): 606–608. дои : 10.1016/j.cell.2010.02.022 . ПМК 3293451 . ПМИД 20211127 .
- ^ Мартин, Дж.; Эттема, TJG (февраль 2013 г.). «От археев к эукариотам: темные века эволюции эукариотической клетки» . Биохим Соц Транс . 41 (1): 451–7. дои : 10.1042/BST20120292 . ПМИД 23356327 .
- ^ Ван де Пер, Ив; Бальдауфрид, Сандра Л.; Дулиттл, У. Форд; Мейерид, Аксель (2000). «Обновленная и комплексная филогения рРНК (коронных) эукариот, основанная на калиброванных по скорости эволюционных расстояниях» . Журнал молекулярной эволюции . 51 (6): 565–576. Бибкод : 2000JMolE..51..565В . дои : 10.1007/s002390010120 . ПМИД 11116330 . S2CID 9400485 .
- ^ Jump up to: а б Баттерфилд, Нью-Джерси (2015). «Ранняя эволюция эукариот» . Палеонтология . 58 (1): 5–17. Бибкод : 2015Palgy..58....5B . дои : 10.1111/пала.12139 .
Внешние ссылки
[ редактировать ]- Влечение и секс среди наших микробных последних общих предков эукариот , The Atlantic , 11 ноября 2020 г.