Оплодотворение

Оплодотворение или оплодотворение (см. различия в написании ), также известное как генеративное оплодотворение , сингамия и оплодотворение , [1] Это слияние гамет , в результате которого образуется зигота и начинается ее развитие в новый индивидуальный организм или потомство. [2] Хотя такие процессы, как оплодотворение или опыление , которые происходят до слияния гамет, иногда неофициально называют оплодотворением. [3] это технически отдельные процессы. Цикл оплодотворения и развития новых особей называется половым размножением . Во время двойного оплодотворения у покрытосеменных гаплоидная , мужская гамета соединяется с двумя гаплоидными полярными ядрами образуя триплоидное первичное ядро эндосперма в процессе вегетативного оплодотворения.
История
[ редактировать ]В древности Аристотель задумал образование новых особей путем слияния мужской и женской жидкостей, при этом форма и функция возникают постепенно, в порядке, названном им эпигенетическим . [4]
В 1784 г. Спалланцани установил необходимость взаимодействия яйцеклетки самки и сперматозоида самца для образования зиготы у лягушек. [5] В 1827 году Карл Эрнст фон Бэр наблюдал яйцо терианского млекопитающего. впервые [4] Оскар Гертвиг (1876) в Германии описал слияние ядер сперматозоидов и яйцеклеток морского ежа . [5]
Эволюция
[ редактировать ]Эволюция оплодотворения связана с возникновением мейоза , так как оба они являются частью полового размножения , зародившегося у эукариот . Одна из гипотез утверждает, что мейоз возник в результате митоза. [6]
Внесение удобрений в растения
[ редактировать ]
Гаметами, участвующими в оплодотворении растений, являются сперматозоид (мужская) и яйцеклетка (женская). Различные группы растений имеют разные методы, с помощью которых гаметы, образующиеся из мужских и женских гаметофитов, собираются вместе и оплодотворяются. У мохообразных и птеридофитных наземных растений оплодотворение спермия и яйцеклетки происходит внутри архегония . У семенных растений мужской гаметофит формируется внутри пыльцевого зерна. После опыления пыльцевое называемую зерно прорастает , пыльцевая трубка разрастается и проникает в семязачаток через крошечную пору, микропиле . Сперма переносится из пыльцы через пыльцевую трубку в семязачаток, где яйцеклетка оплодотворяется. У цветковых растений из пыльцевой трубки высвобождаются два спермия, и происходит второе оплодотворение с участием второго спермия и центральной клетки семяпочки, которая является второй женской гаметой. [7]
Рост пыльцевой трубки
[ редактировать ]В отличие от подвижных сперматозоидов животных, сперма большинства семенных растений неподвижна и доставляется с помощью пыльцевой трубки к семязачатку, где высвобождается сперма. [8] Пыльцевая трубка проникает через рыльце пестика и проходит через внеклеточный матрикс столбика, прежде чем достичь завязи. Затем возле цветоложа она прорывает семязачаток через микропиле (отверстие в стенке семязачатка) и пыльцевая трубка «прорывается» в зародышевый мешок, высвобождая сперму. [9] Считалось, что рост пыльцевой трубки зависит от химических сигналов, исходящих от пестика, однако эти механизмы были плохо изучены до 1995 года. Работа, проведенная на растениях табака, выявила семейство гликопротеинов , называемых TTS-белками, которые усиливают рост пыльцевых трубок. [9] Пыльцевые трубки росли как в среде для проращивания пыльцы, не содержащей сахара, так и в среде с очищенными белками TTS. Однако в среде TTS пробирки росли в 3 раза быстрее, чем в среде без сахара. [9] Белки TTS также были помещены в различные места полуопыляемых in vivo пестиков, и было замечено, что пыльцевые трубки сразу же тянулись к белкам. Трансгенные растения, лишенные способности производить белки TTS, имели более медленный рост пыльцевых трубок и снижали плодовитость. [9]
Разрыв пыльцевой трубки
[ редактировать ]Было показано , что разрыв пыльцевой трубки с высвобождением сперматозоидов у Arabidopsis зависит от сигнала женского гаметофита. Специфические белки, называемые протеинкиназами FER, присутствующие в яйцеклетке, контролируют выработку высокореактивных производных кислорода, называемых активными формами кислорода (АФК). было показано, С помощью GFP что уровни АФК самые высокие на стадиях цветения, когда семяпочка наиболее восприимчива к пыльцевым трубкам, и самые низкие во время развития и после оплодотворения. [8] Высокие количества АФК активируют каналы ионов кальция в пыльцевой трубке, заставляя эти каналы поглощать ионы кальция в больших количествах. Повышенное поглощение кальция приводит к разрыву пыльцевой трубки и выбросу спермы в семязачаток. [8] Анализы кормления пестиков, в которых растениям давали дифенил иодоний хлорид (DPI), подавляли концентрацию АФК у Arabidopsis , что, в свою очередь, предотвращало разрыв пыльцевой трубки. [8]
Цветущие растения
[ редактировать ]После оплодотворения завязь начинает набухать и превращаться в плод . [10] У многосемянных плодов для слияния с каждой семяпочкой необходимо несколько зерен пыльцы. Рост пыльцевой трубки контролируется вегетативной (или трубчатой) цитоплазмой. Гидролитические ферменты секретируются пыльцевой трубкой, которая переваривает женскую ткань по мере того, как трубка растет вниз по рыльцу и столбику; переваренная ткань используется в качестве источника питательных веществ для пыльцевой трубки по мере ее роста. Во время роста пыльцевой трубки по направлению к яичнику генеративное ядро делится, образуя два отдельных ядра спермия (гаплоидное число хромосом). [11] – Таким образом, растущая пыльцевая трубка содержит три отдельных ядра, два спермия и одну трубку. [12] Сперматозоиды взаимосвязаны и диморфны, крупный у ряда растений также связан с ядром трубки, а соединенный между собой спермий и ядро трубки образуют «мужскую зародышевую единицу». [13]
Двойное оплодотворение — это процесс у покрытосеменных (цветковых растений), при котором два спермия из каждой пыльцевой трубки оплодотворяют две клетки женского гаметофита (иногда называемого зародышевым мешком), который находится внутри семязачатка. После того, как пыльцевая трубка попадает в гаметофит, ядро пыльцевой трубки распадается и высвобождаются два спермия; один из двух сперматозоидов оплодотворяет яйцеклетку (в нижней части гаметофита возле микропиле), образуя диплоидную (2n) зиготу . Это момент, когда действительно происходит оплодотворение; опыление и оплодотворение — два отдельных процесса. Ядро другого спермия сливается с двумя гаплоидными полярными ядрами (содержащимися в центральной клетке) в центре гаметофита. Полученная клетка является триплоидной (3n). Эта триплоидная клетка делится митозом и образует эндосперм , питательными веществами богатую ткань внутри семени . [7] Два материнских ядра центральных клеток (полярные ядра), входящие в состав эндосперма, возникают в результате митоза из единственного продукта мейоза, который также дал начало яйцеклетке. Следовательно, материнский вклад в генетическую конституцию триплоидного эндосперма вдвое превышает вклад эмбриона.
Один примитивный вид цветкового растения, Nuphar polysepala , имеет диплоидный эндосперм, образующийся в результате слияния спермия с одним, а не с двумя материнскими ядрами. Считается, что на ранних этапах развития линий покрытосеменных в этом способе размножения произошла дупликация, в результате которой образовались семиклеточные / восьмиядерные женские гаметофиты и триплоидные эндоспермы с соотношением материнского и отцовского генома 2: 1. [14]
У многих растений развитие мякоти плода пропорционально проценту оплодотворенных семязачатков. Например, у арбуза необходимо доставить около тысячи зерен пыльцы и равномерно распределить их по трем лепесткам рыльца, чтобы образовался плод нормального размера и формы. [ нужна ссылка ]
Самоопыление и ауткроссинг
[ редактировать ]Ауткроссинг , или перекрестное оплодотворение, и самооплодотворение представляют собой разные стратегии с разными выгодами и затратами. По оценкам, 48,7% видов растений являются либо раздельнополыми, либо самонесовместимыми облигатными ауткроссами. [15] Также подсчитано, что около 42% цветковых растений в природе демонстрируют смешанную систему спаривания. [16]
В наиболее распространенной системе смешанного скрещивания отдельные растения дают цветы одного типа, а плоды могут содержать самооплодотворенные, ауткроссированные или смесь типов потомства. Переход от перекрестного оплодотворения к самоопылению является наиболее распространенным эволюционным переходом у растений и неоднократно происходил во многих независимых линиях. [17] Около 10-15% цветковых растений преимущественно самоопыляются. [17]
В условиях, когда опылители или партнеры редки, самооплодотворение обеспечивает репродуктивную гарантию . [17] Таким образом, самооплодотворение может привести к улучшению способности к колонизации. У некоторых видов самооплодотворение сохраняется на протяжении многих поколений. Capsella Rubella — это самооплодотворяющийся вид, который стал самосовместимым 50–100 000 лет назад. [18] Arabidopsis thaliana — преимущественно самоопыляющееся растение, уровень ауткроссинга в дикой природе составляет менее 0,3%; [19] исследование показало, что самооплодотворение у A. thaliana развилось примерно миллион лет назад или больше . [20] У давно укоренившихся самооплодотворяющихся растений маскирование вредных мутаций и возникновение генетической изменчивости происходят нечасто и, следовательно, вряд ли принесут достаточную пользу в течение многих поколений для поддержания мейотического аппарата. Следовательно, можно было бы ожидать, что самооплодотворение в природе будет заменено амейотической бесполой формой размножения, которая будет менее затратной. Однако фактическое сохранение мейоза и самооплодотворения как формы размножения у давно существующих самооплодотворяющихся растений может быть связано с непосредственной выгодой от эффективного рекомбинационного восстановления повреждений ДНК во время образования зародышевых клеток, обеспечиваемых мейозом в каждом поколении. [ нужна ссылка ]
Оплодотворение у животных
[ редактировать ]Механика оплодотворения тщательно изучалась на морских ежах и мышах. Это исследование затрагивает вопрос о том, как сперматозоид и соответствующая яйцеклетка находят друг друга, а также вопрос о том, как только один сперматозоид попадает в яйцеклетку и доставляет ее содержимое. Оплодотворение состоит из трех этапов, обеспечивающих видоспецифичность:
- Хемотаксис
- Активация спермы/акросомальная реакция
- Адгезия спермы/яйцеклетки
Внутренний и внешний
[ редактировать ]Решение о том, использует ли животное (точнее позвоночное) внутреннее или внешнее оплодотворение , часто зависит от метода рождения. Яйцекладущие животные, откладывающие яйца с толстой кальциевой скорлупой, например куры , или с толстой кожистой скорлупой, обычно размножаются посредством внутреннего оплодотворения, так что сперма оплодотворяет яйцеклетку, не проходя через толстый защитный третичный слой яйцеклетки. Яйцеживородящие и живородящие животные также используют внутреннее оплодотворение. Хотя некоторые организмы размножаются посредством амплексуса , они все же могут использовать внутреннее оплодотворение, как у некоторых саламандр. Преимущества внутреннего оплодотворения включают минимальные потери гамет, большую вероятность оплодотворения отдельных яиц, более длительный период защиты яиц и избирательное оплодотворение. Многие самки обладают способностью хранить сперму в течение длительного периода времени и могут оплодотворять яйцеклетки по собственному желанию. [ нужна ссылка ]
Напротив, яйцекладущие животные, производящие яйца с тонкой третичной оболочкой или вообще без оболочек, используют методы наружного оплодотворения. Таких животных точнее можно назвать яйцекладущими. [21] Внешнее оплодотворение выгодно тем, что оно сводит к минимуму контакт (что снижает риск передачи заболеваний) и увеличивает генетическую изменчивость.
Морские ежи
[ редактировать ]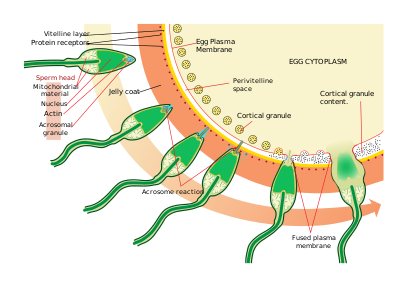
Сперматозоиды находят яйцеклетку посредством хемотаксиса — типа взаимодействия лиганда и рецептора. Resact — это пептид из 14 аминокислот, выделенный из желеобразной оболочки A. punctulata , который привлекает миграцию сперматозоидов.
Найдя яйцеклетку, сперма проникает в желеобразную оболочку посредством процесса, называемого активацией спермы. При другом взаимодействии лиганд/рецептор олигосахаридный компонент яйцеклетки связывается и активирует рецептор сперматозоида и вызывает акросомальную реакцию . Акросомальные пузырьки сперматозоида сливаются с плазматической мембраной и высвобождаются. В этом процессе молекулы, связанные с мембраной акросомального пузырька, такие как биндин, выходят на поверхность сперматозоида. Это содержимое переваривает желеобразную оболочку и, в конечном итоге, желточную оболочку. Помимо высвобождения акросомальных пузырьков, происходит взрывная полимеризация актина с образованием тонкого шипа в головке спермия, называемого акросомальным отростком .
Сперма связывается с яйцеклеткой посредством другой лигандной реакции между рецепторами на желточной мембране . Белок поверхности сперматозоида бидин связывается с рецептором на желточной мембране, идентифицируемым как EBR1.
Слияние плазматических мембран сперматозоида и яйцеклетки, вероятно, опосредовано биндином. В месте контакта слияние вызывает образование конуса оплодотворения .
Млекопитающие
[ редактировать ]Млекопитающие оплодотворяются изнутри посредством совокупления . [22] После эякуляции у мужчины многие сперматозоиды перемещаются в верхнюю часть влагалища (за счет сокращений влагалища) через шейку матки и по всей длине матки , чтобы встретиться с яйцеклеткой. у самки обычно В случаях, когда происходит оплодотворение, овуляция длится от нескольких часов до совокупления до нескольких дней после него; поэтому у большинства млекопитающих эякуляция чаще предшествует овуляции, чем наоборот. [ нужна ссылка ]
Когда сперматозоиды попадают в переднюю часть влагалища, они не способны к оплодотворению (т. е. являются недееспособными). [ нужны разъяснения ] и характеризуются медленными линейными двигательными моделями. Эта подвижность в сочетании с мышечными сокращениями обеспечивает транспорт спермы к матке и яйцеводам . [23] В микроокружении женских репродуктивных путей существует градиент pH, так что pH возле отверстия влагалища ниже (приблизительно 5), чем в яйцеводах (приблизительно 8). [24] Специфический для сперматозоидов pH-чувствительный белок-переносчик кальция, называемый CatSper, увеличивает проницаемость сперматозоидов для кальция по мере его дальнейшего продвижения в репродуктивный тракт. Внутриклеточный приток кальция способствует капацитации и гиперактивации сперматозоидов, вызывая более сильную и быструю нелинейную подвижность по мере приближения сперматозоидов к ооциту. сперматозоид Капацитированный и яйцеклетка встречаются и взаимодействуют в ампуле фаллопиевой трубы . Реотаксис, термотаксис и хемотаксис — известные механизмы, которые направляют сперму к яйцеклетке на заключительной стадии миграции сперматозоидов. [25] Сперматозоиды реагируют (см. Термотаксис спермы ) на температурный градиент ~ 2 ° C между яйцеводом и ампулой. [26] и хемотаксические градиенты прогестерона были подтверждены как сигнал, исходящий от клеток кумулюса офоруса, окружающих ооциты кролика и человека. [27] Капацитированные и гиперактивированные сперматозоиды реагируют на эти градиенты, изменяя свое поведение и перемещаясь в сторону комплекса кумулюс-ооцит. Другие хемотаксические сигналы, такие как формил Met-Leu-Phe (fMLF), также могут направлять сперматозоиды. [28]
Зона пеллюцида , толстый слой внеклеточного матрикса, который окружает яйцеклетку и аналогичен роли желточной оболочки морских ежей, связывает сперму. В отличие от морских ежей, сперма связывается с яйцеклеткой до акросомальной реакции. ZP3 , гликопротеин в зоне пеллюцида, отвечает за адгезию яйцеклеток/сперматозоидов у человека. Рецепторная галактозилтрансфераза (GalT) связывается с остатками N-ацетилглюкозамина на ZP3 и важна для связывания со сперматозоидами и активации акросомной реакции. ZP3 достаточен, хотя и не нужен для связывания сперматозоидов с яйцеклетками. Существуют два дополнительных рецептора сперматозоидов: белок массой 250 кДа, который связывается с белком, секретируемым яйцеводом, и SED1, который независимо связывается с оболочкой. Считается, что после акросомной реакции сперматозоиды остаются связанными с прозрачной оболочкой через открытые рецепторы ZP2. Эти рецепторы неизвестны у мышей, но были идентифицированы у морских свинок. [ нужна ссылка ]
У млекопитающих связывание сперматозоида с GalT инициирует акросомную реакцию . В результате этого процесса высвобождается гиалуронидаза , которая переваривает матрицу гиалуроновой кислоты в оболочках вокруг яйцеклетки. Кроме того, вблизи ооцита высвобождаются гепариноподобные гликозаминогликаны (ГАГ), которые способствуют акросомной реакции. [29] За этим следует слияние плазматических мембран спермия ооцита и сперматозоида, которое позволяет ядру , типичной центриоли и атипичной центриоли , прикрепленной к жгутику , но не к митохондриям , проникнуть в ооцит. [30] Белок CD9 , вероятно, опосредует это слияние у мышей (связывающий гомолог). Яйцеклетка « активируется » при слиянии с одним сперматозоидом и тем самым меняет свою клеточную мембрану, чтобы предотвратить слияние с другими сперматозоидами. цинка . Во время этой активации высвобождаются атомы [ нужна ссылка ]
Этот процесс в конечном итоге приводит к образованию диплоидной клетки, называемой зиготой . Зигота делится с образованием бластоцисты и, попадая в матку, имплантируется в эндометрий, начиная беременность . Имплантация эмбриона не в стенку матки приводит к внематочной беременности , которая может привести к гибели матери.
У таких животных, как кролики, половой акт вызывает овуляцию, стимулируя высвобождение гипофизарного гормона гонадотропина; этот выпуск значительно увеличивает вероятность беременности.
Люди
[ редактировать ]
Оплодотворение у человека — это союз человеческой яйцеклетки и сперматозоида , обычно происходящий в ампуле фаллопиевой трубы , в результате которого образуется одноклеточная зигота , первый этап жизни в развитии генетически уникального организма. [31] и инициирование эмбрионального развития . Ученые открыли динамику оплодотворения человека еще в девятнадцатом веке.
Термин «зачатие» обычно относится к «процессу зачатия, включающему оплодотворение или имплантацию, или и то, и другое». [32] Его использование делает его предметом семантических споров о начале беременности , обычно в контексте дебатов об абортах .При гаструляции , которая происходит примерно через 16 дней после оплодотворения, в имплантированной бластоцисте развиваются три зародышевых листка: энтодерма, эктодерма и мезодерма, а генетический код отца полностью участвует в развитии эмбриона; позднее спаривание невозможно. Кроме того, межвидовые гибриды выживают только до гаструляции и не могут дальше развиваться.Однако в некоторой литературе по биологии развития человека упоминается концепт , а в такой медицинской литературе упоминаются «продукты зачатия», такие как постимплантационный эмбрион и окружающие его мембраны. [33] Термин «концепция» обычно не используется в научной литературе из-за его вариативного определения и коннотации.
Насекомые
[ редактировать ]
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( февраль 2015 г. ) |
Насекомые разных групп, в том числе Odonata ( стрекозы и стрекозы ) и перепончатокрылые ( муравьи , пчелы и осы ), практикуют отложенное оплодотворение. Среди Odonata самки могут спариваться с несколькими самцами и хранить сперму до тех пор, пока не будут отложены яйца. Самец может парить над самкой во время откладки яиц (яйцекладки), чтобы не дать ей спариваться с другими самцами и заменить его сперму; в некоторых группах, таких как дартеры, самец продолжает хватать самку своими пряжками во время откладки яиц, при этом пара летает в тандеме. [34] Среди социальных перепончатокрылых пчелиные матки спариваются только во время брачных полетов, в течение короткого периода, продолжающегося несколько дней; королева может спариваться с восемью или более трутнями . Затем она хранит сперму до конца своей жизни, возможно, в течение пяти лет или более. [35] [36]
Оплодотворение у грибов
[ редактировать ]У многих грибов (кроме хитридов ), как и у некоторых простейших, оплодотворение представляет собой двухэтапный процесс. Сначала цитоплазмы двух гаметных клеток сливаются (так называемая плазмогамия ), образуя дикариотические или гетерокариотические клетки с несколькими ядрами. Эта клетка может затем делиться с образованием дикариотических или гетерокариотических гиф . Второй этап оплодотворения — кариогамия , слияние ядер с образованием диплоидной зиготы.
У хитридиевых грибов оплодотворение происходит в один этап со слиянием гамет, как у животных и растений.
Оплодотворение у простейших
[ редактировать ]Оплодотворение у простейших
[ редактировать ]У простейших различают три типа процессов оплодотворения: [37]
Оплодотворение в водорослях
[ редактировать ]Водоросли, как и некоторые наземные растения, претерпевают смену поколений . Некоторые водоросли изоморфны, при этом и спорофит (2n), и гаметофит (n) морфологически одинаковы. Когда размножение водорослей описывается как оогамное, мужские и женские гаметы морфологически различаются: для женских гамет имеется большое неподвижное яйцо, а мужские гаметы одножгутиковые (подвижные). В процессе сингамии они образуют новую зиготу, снова регенерируя поколение спорофитов.
Оплодотворение и генетическая рекомбинация
[ редактировать ]Мейоз приводит к случайному разделению генов, вносимых каждым родителем. Каждый родительский организм обычно идентичен, за исключением части генов; поэтому каждая гамета генетически уникальна. При оплодотворении родительские хромосомы объединяются. У человека (2²²)² = 17,6x10. 12 с разными зиготы Для неполовых хромосом возможны хромосомами, даже если предположить отсутствие хромосомного кроссинговера . Если пересечение происходит один раз, то в среднем (4²²)² = 309x10 24 Для каждой пары возможны генетически разные зиготы, не учитывая, что события кроссинговера могут происходить в большинстве точек каждой хромосомы. Хромосомы X и Y не подвергаются событиям кроссинговера. [ нужна ссылка ] и поэтому исключаются из расчета. Митохондриальная ДНК наследуется только от материнского родителя.
Астра спермия и центросомы зиготы
[ редактировать ]Вскоре после слияния спермия с яйцеклеткой две центриоли эмбриона спермия образуют первую центросому и астральную микротрубочку . [40] Центриоль спермия, расположенная рядом с мужским пронуклеусом, рекрутирует белки перицентриолярного материала яйцеклетки , образуя первую центросому зиготы. [41] Эта центросома образует ядра микротрубочек в форме звезд, называемых астральными микротрубочками. Микротрубочки охватывают всю валиум яйцеклетки, позволяя пронуклеусу яйцеклетки использовать кабели, чтобы добраться до мужского пронуклеуса. Когда мужские и женские пронуклеусы приближаются друг к другу, одна центросома разделяется на две центросомы, расположенные в интерфазе между пронуклеусами. Затем центросома через астральные микротрубочки поляризует геном внутри пронуклеусов. [42]
Партеногенез
[ редактировать ]Организмы, которые обычно размножаются половым путем, также могут размножаться посредством партеногенеза , при котором неоплодотворенная женская гамета дает жизнеспособное потомство. Эти потомки могут быть клонами матери или в некоторых случаях генетически отличаться от нее, но унаследовать только часть ее ДНК. Партеногенез происходит у многих растений и животных, а у других может быть вызван химическим или электрическим стимулом яйцеклетки. В 2004 году японским исследователям под руководством Томохиро Коно удалось после 457 попыток объединить яйцеклетки двух мышей путем блокирования определенных белков, которые в норме предотвращали бы такую возможность; полученный эмбрион нормально развился в мышь. [43]
Аллогамия и автогамия
[ редактировать ]Аллогамия , также известная как перекрестное оплодотворение, относится к оплодотворению яйцеклетки одного человека мужской гаметой другого.
Автогамия, также известная как самооплодотворение, встречается у таких гермафродитных организмов, как растения и плоские черви; в нем две гаметы от одного индивидуального слияния.
Другие варианты двуполого размножения
[ редактировать ]Некоторые относительно необычные формы размножения: [44] [45]
Гиногенез : сперма стимулирует развитие яйцеклетки без оплодотворения или сингамии. Сперма может проникнуть в яйцеклетку.
Гибридогенез : один геном удаляется для образования гаплоидных яиц.
Мейоз собак : (иногда называемый «постоянной нечетной полиплоидией») один геном передается менделевским способом, другие передаются клонально.
Преимущества перекрестного оплодотворения
[ редактировать ]Обычно считается, что основным преимуществом перекрестного оплодотворения является предотвращение инбридинговой депрессии . Чарльз Дарвин в своей книге 1876 года «Эффекты перекрестного и самооплодотворения в растительном мире» (стр. 466–467) подытожил свои открытия следующим образом. [46]
«В настоящей книге было показано, что потомство от союза двух различных особей, особенно если их предки находились в совершенно разных условиях, имеет огромное преимущество в росте, весе, конституциональной силе и плодовитости по сравнению с самооплодотворенными особями. потомство от одного и того же родителя, и этого факта вполне достаточно, чтобы объяснить развитие половых элементов, то есть возникновение двух полов».
Кроме того, некоторые считают, [47] что долгосрочным преимуществом ауткроссинга в природе является повышение генетической изменчивости, которая способствует адаптации или предотвращению вымирания (см. Генетическая изменчивость ).
См. также
[ редактировать ]- Слияние клеток
- Концептуальная шапка
- Устройство для зачатия
- Женская сперма
- Развитие плода
- Экстракорпоральное оплодотворение
- Кагуя (мышь)
- Партеногенез – тип размножения, не предполагающий оплодотворения.
- Опыление
- Преэмбрион
- Пронуклеус
- Суперфекундация
- Суперфетация
- Нарушение симметрии и кортикальная ротация
- Корковая реакция
- Полиспермия
Ссылки
[ редактировать ]- ^ «пропитка» . Оксфордский словарь для продвинутых учащихся .
- ^ Сиу, Карен К.; Серран, Витор Уго Б.; Зийят, Ахмед; Ли, Джеффри Э. (2021). «Клеточная биология оплодотворения: прикрепление и слияние гамет» . Журнал клеточной биологии . 220 (10). дои : 10.1083/jcb.202102146 . ПМЦ 8406655 . ПМИД 34459848 . Проверено 14 января 2023 г.
- ^ «Оплодотворение» . Мерриам-Вебстер . Проверено 10 июля 2018 г.
- ^ Перейти обратно: а б Майеншайн, Джейн (2017). «Первый век клеточной теории: от структурных единиц к сложным живым системам». Интегрированная история и философия науки . Ежегодник Института Венского кружка. Том. 20. стр. 43–54. дои : 10.1007/978-3-319-53258-5_4 . ISBN 978-3-319-53257-8 . S2CID 89731001 .
- ^ Перейти обратно: а б Биркхед, Тим Р.; Монтгомери, Роберт (2009). «Три столетия исследований спермы». Биология спермы . стр. 1–42. дои : 10.1016/B978-0-12-372568-4.00001-X . ISBN 978-0-12-372568-4 .
- ^ Уилкинс А.С., Холлидей Р. (январь 2009 г.). «Эволюция мейоза из митоза» . Генетика . 181 (1): 3–12. doi : 10.1534/genetics.108.099762 . ПМК 2621177 . ПМИД 19139151 .
- ^ Перейти обратно: а б Фор, Дж. Э. (1999). «Двойное оплодотворение цветковых растений: происхождение, механизмы и новая информация об экстракорпоральном оплодотворении». В Крести, М.; Кай, Г.; Москателли, А. (ред.). Оплодотворение у высших растений . Берлин, Гейдельберг.: Springer. стр. 79–89. ISBN 978-3-642-59969-9 .
- ^ Перейти обратно: а б с д Дуань, Цяохун; Кита, Дэниел; Джонсон, Эрик А; Аггарвал, Мини; Гейтс, Лаура; Ву, Хен-Мин; Чунг, Алиса Ю (2014). «Активные формы кислорода опосредуют разрыв пыльцевой трубки с высвобождением спермы для оплодотворения арабидопсиса» . Природные коммуникации . 5 : 3129. Бибкод : 2014NatCo...5.3129D . дои : 10.1038/ncomms4129 . ПМИД 24451849 .
- ^ Перейти обратно: а б с д Чунг, Элис Ю ; Ван, Хун; Ву, Хен-Мин (1995). «Цветочный передающий тканеспецифичный гликопротеин привлекает пыльцевые трубки и стимулирует их рост» . Клетка . 82 (3): 383–93. дои : 10.1016/0092-8674(95)90427-1 . ПМИД 7634328 .
- ^ Джонстон, Адам (2001). Биология: факты и практика для уровня A. Издательство Оксфордского университета. п. 95 . ISBN 0-19-914766-3 .
- ^ Справочник по растениеводству . Чичестер, Западный Суссекс, Англия: Джон Уайли. 2007. с. 466. ИСБН 978-0-470-05723-0 .
- ^ Кирк, Дэвид; Старр, Сеси (1975). Биология сегодня . Дель Мар, Калифорния: CRM. п. 93 . ISBN 978-0-394-31093-0 .
- ^ Рагхаван, Валаямгхат (2006). Двойное оплодотворение: развитие зародыша и эндосперма у цветкового растения . Берлин: Springer-Verlag. п. 12 . ISBN 978-3-540-27791-0 .
- ^ Фридман, Уильям Э; Уильямс, Джозеф Х (2003). «Модульность женского гаметофита покрытосеменных и ее влияние на раннюю эволюцию эндосперма у цветковых растений» . Эволюция . 57 (2): 216–30. дои : 10.1111/j.0014-3820.2003.tb00257.x . ПМИД 12683519 .
- ^ Игич Б., Кон младший (2006). «Распространение систем скрещивания растений: изучите предвзятость против обязательного скрещивания видов». Эволюция . 60 (5): 1098–103. дои : 10.1554/05-383.1 . ПМИД 16817548 . S2CID 40964 .
- ^ Гудвилли С., Калиш С., Экерт К.Г. (2005). «Эволюционная загадка смешанных систем спаривания у растений: возникновение, теоретические объяснения и эмпирические данные». Анну. Преподобный Экол. Эвол. Сист . 36 : 47–79. doi : 10.1146/annurev.ecolsys.36.091704.175539 . S2CID 3755371 .
- ^ Перейти обратно: а б с Райт, С.И.; Калиш, С; Слотт, Т (2013). «Эволюционные последствия самоопыления растений» . Труды Королевского общества B: Биологические науки . 280 (1760): 20130133. doi : 10.1098/rspb.2013.0133 . ПМЦ 3652455 . ПМИД 23595268 .
- ^ Брандвейн, Янив; Слотте, Таня; Хаззури, Халед М; Райт, Стивен I; Куп, Грэм (2013). «Геномная идентификация основополагающих гаплотипов раскрывает историю самоопыляющегося вида Capsella Rubella» . ПЛОС Генетика . 9 (9): e1003754. arXiv : 1307.4118 . Бибкод : 2013arXiv1307.4118B . дои : 10.1371/journal.pgen.1003754 . ПМЦ 3772084 . ПМИД 24068948 .
- ^ Эбботт, Р.Дж.; Гомес, МФ (1989). «Генетическая структура популяции и уровень ауткроссинга Arabidopsis thaliana (L.) Heynh» . Наследственность . 62 (3): 411–418. дои : 10.1038/hdy.1989.56 .
- ^ Тан С., Тумаджян С., Шерман-Бройлс С., Планнол В., Го Ю.Л., Ху Т.Т., Кларк Р.М., Насралла Дж.Б., Вайгель Д., Нордборг М. (2007). «Эволюция самоопыления Arabidopsis thaliana». Наука . 317 (5841): 1070–2. Бибкод : 2007Sci...317.1070T . дои : 10.1126/science.1143153 . ПМИД 17656687 . S2CID 45853624 .
- ^ Лоде, Тьерри (2001). Стратегии воспроизводства животных [ Стратегии воспроизводства в животном мире ] (на французском языке). Дюнод. ISBN 978-2-10-005739-9 . [ нужна страница ]
- ^ Нагиб, Марк (19 апреля 2020 г.). Достижения в изучении поведения . Академическая пресса. ISBN 978-0-12-820726-0 .
- ^ Суарес, СС; Пейси, А.А. (2006). «Транспорт спермы в женских репродуктивных путях» . Обновление репродукции человека . 12 (1): 23–37. дои : 10.1093/humupd/dmi047 . ПМИД 16272225 .
- ^ Нг, Ка Ин Бонни; Мингельс, Роэль; Морган, Хиуэл; Маклон, Ник; Чеонг, Ин (1 января 2018 г.). «Динамика кислорода, температуры и pH in vivo в женских репродуктивных путях и их значение для зачатия человека: систематический обзор» . Обновление репродукции человека . 24 (1): 15–34. дои : 10.1093/humupd/dmx028 . ПМИД 29077897 .
- ^ Ли, Шуай; Винутаянон, Випави (2016). «Яйцевод: роль в оплодотворении и раннем развитии эмбриона» . Журнал эндокринологии . 232 (1): Р1–Р26. дои : 10.1530/JOE-16-0302 . ПМИД 27875265 .
- ^ Бахат, Анат; Айзенбах, Майкл (2006). «Термотаксис спермы» . Молекулярная и клеточная эндокринология . 252 (1–2): 115–9. дои : 10.1016/j.mce.2006.03.027 . ПМИД 16672171 . S2CID 22094855 .
- ^ Тевес, Мария Э; Гвидобальди, Гектор А; Уньятес, Диего Р.; Санчес, Рауль; Миска, Вернер; Публиковер, Стивен Дж; Моралес Гарсия, Адуэн А; Джохалас, Лаура С. (2009). «Молекулярный механизм хемотаксиса спермы человека, опосредованный прогестероном» . ПЛОС ОДИН . 4 (12): е8211. Бибкод : 2009PLoSO...4.8211T . дои : 10.1371/journal.pone.0008211 . ПМК 2782141 . ПМИД 19997608 .
- ^ Гнесси Л., Фаббри А., Сильвестрони Л., Моретти С., Фрайоли Ф., Перт С.Б., Исидори А. (1986). «Доказательства наличия специфических рецепторов N-формиловых хемотаксических пептидов на сперматозоидах человека». J Clin Эндокринол Метаб . 63 (4): 841–846. doi : 10.1210/jcem-63-4-841 . ПМИД 3018025 .
- ^ Ли, Китай; Клейтон, МК; Бушмейер, С.М.; Во-первых, Нидерланды; Акс, РЛ (1 сентября 1986 г.). «Гликозаминогликаны в репродуктивных путях овцы и их влияние на акросомные реакции в сперматозоидах крупного рогатого скота in vitro». Журнал зоотехники . 63 (3): 861–867. дои : 10.2527/jas1986.633861x . ПМИД 3759713 . S2CID 853558 .
- ^ Фишман, Эмили Л; Джо, Кёнг; Нгуен, Куинь П.Х; Конг, Донг; Ройфман, Рэйчел; Чекич, Энтони Р.; Ханал, Сушил; Миллер, Энн Л; Симерли, Кальвин; Шаттен, Джеральд; Лонкарек, Ядранка; Меннелла, Вито; Авидор-Рейсс, Томер (2018). «Новая атипичная центриоль сперматозоида функционирует во время оплодотворения человека» . Природные коммуникации . 9 (1): 2210. Бибкод : 2018NatCo...9.2210F . дои : 10.1038/s41467-018-04678-8 . ПМЦ 5992222 . ПМИД 29880810 .
- ^ «Зигота | Определение, развитие, примеры и факты | Британника» . www.britanica.com . Проверено 14 января 2023 г.
- ^ «Концепция» . Мерриам-Вебстер . Проверено 27 января 2019 г.
- ^ Мур, КЛ; ТВМ Персо (2003). Развивающийся человек: клинически ориентированная эмбриология . Компания WB Saunders. ISBN 0-7216-6974-3 . [ нужна страница ]
- ^ Дейкстра, Клаас-Доуве Б. (2006). Полевое руководство по стрекозам Британии и Европы . Британское издательство дикой природы. стр. 8–9. ISBN 0-9531399-4-8 .
- ^ Вальдбауэр, Гилберт (1998). Книга ошибок птицелова . Издательство Гарвардского университета.
- ^ Сельское хозяйство и защита потребителей. «Пчеловодство в Африке: жизнь колонии и социальная организация» . ФАО.
- ^ Тарин, Хуан Дж.; Кано, Антонио (2000). Оплодотворение у простейших и многоклеточных животных: клеточные и молекулярные аспекты . Springer Science & Business Media. ISBN 978-3-540-67093-3 . [ нужна страница ]
- ^ Репродукция # Автогамия
- ^ «Автогамия | биология» .
- ^ Авидор-Рейсс, Томер; Мазур, Мэтью; Фишман, Эмили Л.; Синдхвани, Пунит (1 октября 2019 г.). «Роль центриолей сперматозоидов в воспроизводстве человека – известное и неизвестное» . Границы клеточной биологии и биологии развития . 7 : 188. дои : 10.3389/fcell.2019.00188 . ПМЦ 6781795 . ПМИД 31632960 .
- ^ Фишман, Эмили Л.; Джо, Кёнг; Нгуен, Куинь ПХ; Конг, Донг; Ройфман, Рэйчел; Чекич, Энтони Р.; Ханал, Сушил; Миллер, Энн Л.; Симерли, Кальвин; Шаттен, Джеральд; Лонкарек, Ядранка; Меннелла, Вито; Авидор-Рейсс, Томер (декабрь 2018 г.). «Новая атипичная центриоль сперматозоида функционирует во время оплодотворения человека» . Природные коммуникации . 9 (1): 2210. Бибкод : 2018NatCo...9.2210F . дои : 10.1038/s41467-018-04678-8 . ПМЦ 5992222 . ПМИД 29880810 .
- ^ Кавацца, Томмазо; Такеда, Юко; Полити, Антонио З.; Аушев, Магомет; Алдаг, Патрик; Бейкер, Клара; Чоудхари, Минакши; Буцявичюс, Йонас; Лукинавичюс, Гражвидас; Старейшина, Кей; Блейни, Мартин; Лукас-Хан, Андреа; Ниманн, Хайнер; Герберт, Мэри; Шу, Мелина (май 2021 г.). «Унификация родительского генома очень подвержена ошибкам у эмбрионов млекопитающих» . Клетка . 184 (11): 2860–2877.e22. дои : 10.1016/j.cell.2021.04.013 . ПМК 8162515 . ПМИД 33964210 .
- ^ Коно, Томохиро; Обата, Яёи; Ву, Кьонг; Нива, Кацутоши; Оно, Юкико; Ямамото, Юджи; Пак Ын Сон; Со, Чон Сон; Огава, Хидехико (2004). «Рождение партеногенетических мышей, способных развиваться до взрослой жизни». Природа . 428 (6985): 860–4. Бибкод : 2004Natur.428..860K . дои : 10.1038/nature02402 . ПМИД 15103378 . S2CID 4353479 .
- Биджал П. Триведи (21 апреля 2004 г.). «Конец самцам? Мыши созданы для размножения без спермы» . Нэшнл Географик . Архивировано из оригинала 23 апреля 2004 г.
- ^ Стенберг, П; Саура, А (2013). «Мейоз и его отклонения у полиплоидных животных». Цитогенетические и геномные исследования . 140 (2–4): 185–203. дои : 10.1159/000351731 . ПМИД 23796636 . S2CID 19955975 .
- ^ Сток, М; Устинова Дж.; Бетто-Коллиард, К; Шартл, М; Мориц, К; Перрин, Н. (2011). «Одновременная передача менделевского и клонального генома у полностью триплоидных позвоночных, размножающихся половым путем» . Труды Королевского общества B: Биологические науки . 279 (1732): 1293–1299. дои : 10.1098/rspb.2011.1738 . ПМЦ 3282369 . ПМИД 21993502 .
- ^ Дарвин, Чарльз (1876). Эффекты перекрестного и самооплодотворения в растительном царстве . Дж. Мюррей. стр. 466–467. OCLC 57556547 .
- ^ Отто, СП; Герштейн, AC (2006). «Зачем заниматься сексом? Популяционная генетика пола и рекомбинации». Труды Биохимического общества . 34 (4): 519–22. дои : 10.1042/BST0340519 . ПМИД 16856849 . S2CID 14542950 .
Внешние ссылки
[ редактировать ]- оплодотворения (зачатия) Видео
