Двойное оплодотворение

Двойное оплодотворение или двойное оплодотворение (см. различия в написании ) — сложный оплодотворения механизм покрытосеменных растений . Этот процесс включает слияние женского гаметофита или мегагаметофита , также называемого эмбриональным мешком , с двумя мужскими гаметами (сперматозоидами). Оно начинается, когда пыльцевое зерно прилипает к стигматической поверхности плодолистика — женской репродуктивной структуры цветков покрытосеменных. Пыльцевое зерно начинает прорастать (если только у этого конкретного вида не возникает и не активируется тип самонесовместимости , действующий на рыльце пестика), образуя пыльцевую трубку , которая проникает через столбик и распространяется вниз по направлению к завязи , следуя выделяемым химическим сигналам. за яйцом. Кончик пыльцевой трубки затем попадает в яичник, проникая через отверстие микропиле в семязачатке , и выпускает два спермия в зародышевый мешок (мегагаметофит).
Зрелый зародышевый мешок неоплодотворенной семяпочки семиклеточный и восьмиядерный. Он устроен в форме 3+1+3 (сверху вниз), т.е. 3 антиподальные клетки, 1 центральная клетка (двуядерная), 2 синергиды и 1 яйцеклетка. Один спермий оплодотворяет яйцеклетку, а другой спермий сливается с двумя полярными ядрами большой центральной клетки мегагаметофита. Гаплоидный сперматозоид и гаплоидная яйцеклетка сливаются , образуя диплоидную зиготу, этот процесс называется сингамией , в то время как другой сперматозоид и диплоидная центральная клетка сливаются, образуя триплоидную первичную клетку эндосперма (тройное слияние). Некоторые растения могут образовывать полиплоидные ядра. Большая клетка гаметофита затем разовьется в эндосперм — богатую питательными веществами ткань, которая питает развивающийся эмбрион. Завязь , окружающая семяпочки, развивается в плод, который защищает семена и может рассеивать их. [ 1 ]
Два материнских ядра центральных клеток (полярные ядра), входящие в состав эндосперма, возникают в результате митоза из того же единственного продукта мейоза , который дал начало яйцеклетке. Материнский вклад в генетическую конституцию триплоидного эндосперма вдвое превышает вклад сперматозоида.
В исследовании растения Arabidopsis thaliana , проведенном в 2008 году, впервые с использованием in vivo визуализации была задокументирована миграция мужских ядер внутри женской гаметы при слиянии с женскими ядрами. Также были определены некоторые гены, участвующие в процессах миграции и слияния. [ 2 ]
Сообщалось о доказательствах двойного оплодотворения у Gnetales , которые являются нецветковыми семенными растениями. [ 3 ]
Краткая история
[ редактировать ]Двойное оплодотворение было открыто более века назад Сергеем Наващиным в Киеве . [ 4 ] и Леон Гиньяр во Франции . Каждый сделал открытие независимо от другого. [ 5 ] Lilium martagon и Fritillaria tenella использовались в первых наблюдениях двойного оплодотворения, которые были сделаны с помощью классического светового микроскопа . Из-за ограничений светового микроскопа оставалось много вопросов, касающихся процесса двойного оплодотворения, оставшихся без ответа. Однако с развитием электронного микроскопа на многие вопросы были получены ответы. В частности, наблюдения группы У. Йенсена показали, что мужские гаметы не имеют клеточных стенок и что плазматическая мембрана гамет близка к плазматической мембране клетки, окружающей их внутри пыльцевого зерна. [ 6 ]
Двойное оплодотворение у голосеменных растений
[ редактировать ]Гораздо более элементарная форма двойного оплодотворения встречается при половом размножении отряда голосеменных растений, широко известного как Gnetales. [ 3 ] В частности, это событие было зарегистрировано как у Ephedra , так и у Gnetum , подмножества гнетофитов . [ 7 ] У Ephedra nevadensis в яйцеклетке откладывается один двуядерный сперматозоид. После первоначального оплодотворения второе ядро спермия направляется для оплодотворения дополнительного ядра яйцеклетки, обнаруженного в цитоплазме яйцеклетки. У большинства других семенных растений это второе «ядро вентрального канала» обычно оказывается функционально бесполезным. [ 8 ] У Gnetum gnemon в женской цитоплазме внутри женского гаметофита существуют многочисленные свободные ядра яйцеклеток . После проникновения зрелого женского гаметофита в пыльцевую трубку женская цитоплазма и свободные ядра перемещаются, окружая пыльцевую трубку. Из двуядерной спермы выделяются два ядра сперматозоидов, которые затем сливаются со свободными ядрами яйцеклеток, образуя две жизнеспособные зиготы, что является гомологичной характеристикой между семействами Ephedra и Gnetum . [ 9 ] В обеих семьях в результате второго оплодотворения образуется дополнительный диплоидный эмбрион . Этот нештатный эмбрион позже абортируется, что приводит к синтезу только одного зрелого эмбриона. [ 10 ] Дополнительный продукт оплодотворения эфедры не питает первичный зародыш, поскольку за обеспечение питательными веществами отвечает женский гаметофит. [ 9 ] Более примитивный процесс двойного оплодотворения у голосеменных приводит к образованию двух диплоидных ядер, заключенных в одной яйцеклетке. Это отличается от состояния покрытосеменных , которое приводит к разделению яйцеклетки и эндосперма . [ 11 ] Сравнительные молекулярные исследования генома G. gnemon показали, что гнетофиты более тесно связаны с хвойными, чем с покрытосеменными. [ 12 ] [ 13 ] [ 14 ] Отказ от антофитной гипотезы, согласно которой гнеталеи и покрытосеменные являются сестринскими таксонами, приводит к предположению, что процесс двойного оплодотворения является продуктом конвергентной эволюции и возник независимо у гнетофитов и покрытосеменных. [ 15 ]
Двойное оплодотворение экстракорпорально
[ редактировать ]Двойное оплодотворение in vitro часто используется для изучения молекулярных взаимодействий, а также других аспектов слияния гамет у цветковых растений. Одним из основных препятствий в развитии двойного оплодотворения in vitro между мужскими и женскими гаметами является удержание сперматозоида в пыльцевой трубке, а яйцеклетки - в зародышевом мешке. Контролируемое слияние яйцеклетки и сперматозоида уже было достигнуто с помощью растений мака . [ 16 ] Прорастание пыльцы , попадание пыльцевой трубки и процессы двойного оплодотворения протекают нормально. Фактически, этот метод уже использовался для получения семян различных цветковых растений и получил название «оплодотворение в пробирке». [ 17 ]
Связанные структуры и функции
[ редактировать ]Мегагаметофит
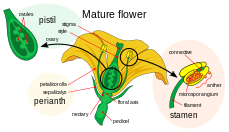
[ редактировать ]Женский гаметофит, мегагаметофит, участвующий в двойном оплодотворении у покрытосеменных гаплоидных растений , называется зародышевым мешком. Он развивается внутри семязачатка , окруженного завязью у основания плодолистика . Мегагаметофит окружают (один или) два покрова , образующие отверстие, называемое микропиле . Мегагаметофит, который обычно является гаплоидным , происходит из (обычно диплоидной ) материнской клетки мегаспоры , также называемой мегаспороцитом . Следующая последовательность событий варьируется в зависимости от конкретного вида, но у большинства видов происходят следующие события. Мегаспороцит подвергается мейозу, образуя четыре гаплоидные мегаспоры. Из четырех образовавшихся мегаспор выживает только одна. Эта мегаспора претерпевает три цикла митоза, в результате чего образуются семь клеток с восемью гаплоидными ядрами (центральная клетка имеет два ядра, называемых полярными ядрами). Нижний конец зародышевого мешка состоит из гаплоидной яйцеклетки, расположенной посередине двух других гаплоидных клеток, называемых синергиды . Синергиды участвуют в привлечении и направлении пыльцевой трубки к мегагаметофиту через микропиле. На верхнем конце мегагаметофита расположены три антиподальные клетки.
Микрогаметофит
[ редактировать ]Мужские гаметофиты или микрогаметофиты, участвующие в двойном оплодотворении, содержатся в пыльцевых зернах. Они развиваются внутри микроспорангиев или пыльцевых мешочков пыльников тычинок. Каждый микроспорангий содержит диплоидные материнские клетки микроспор или микроспороциты. Каждый микроспороцит подвергается мейозу, образуя четыре гаплоидные микроспоры, каждая из которых со временем может развиться в пыльцевое зерно. Микроспора подвергается митозу и цитокинезу , чтобы произвести две отдельные клетки: генеративную клетку и трубчатую клетку. Эти две клетки помимо споровой стенки составляют незрелое пыльцевое зерно. По мере созревания мужского гаметофита генеративная клетка переходит в трубчатую клетку, а генеративная клетка подвергается митозу, производя две сперматозоиды. Когда пыльцевое зерно созревает, пыльники раскрываются, высвобождая пыльцу. Пыльца переносится ветром или животными-опылителями на пестик другого цветка и откладывается на рыльце пестика. По мере прорастания пыльцевого зерна клетка трубки образует пыльцевую трубку, которая удлиняется и распространяется вниз по длинному стержню плодолистика к завязи, где ее сперматозоиды высвобождаются в мегагаметофите. Отсюда происходит двойное оплодотворение. [ 18 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Бергер, Ф. (январь 2008 г.). «Двойное оплодотворение: от мифов к реальности». Половое размножение растений . 21 (1): 3–5. дои : 10.1007/s00497-007-0066-4 . S2CID 8928640 .
- ^ Бергер, Ф.; Хамамура Ю., Ингуфф М. и Хигасияма Т. (август 2008 г.). «Двойное оплодотворение - пойманы с поличным». Тенденции в науке о растениях . 13 (8): 437–443. doi : 10.1016/j.tplants.2008.05.011 . ПМИД 18650119 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б В. Рагхаван (сентябрь 2003 г.). «Некоторые размышления о двойном оплодотворении, от его открытия до наших дней». Новый фитолог . 159 (3): 565–583. дои : 10.1046/j.1469-8137.2003.00846.x . ПМИД 33873607 .
- ^ Кордиум Э.Л. (2008). «[Двойное оплодотворение у цветковых растений: 1898-20&&&--». Цитол. Жене. (на русском языке). 42 (3): 12–26. ПМИД 18822860 .
- ^ Дженсен, Вашингтон (февраль 1998 г.). «Двойное оплодотворение: личный взгляд». Половое размножение растений . 11 (1): 1–5. дои : 10.1007/s004970050113 . S2CID 33416360 .
- ^ Дюма К. и Роговский П. (август 2008 г.). «Оплодотворение и раннее семяобразование» . Comptes Rendus Biologies . 331 (10): 715–725. дои : 10.1016/j.crvi.2008.07.013 . ПМИД 18926485 .
- ^ Кармайкл, Дж. С.; Фридман, МЫ (1 декабря 1995 г.). «Двойное оплодотворение у Gnetum gnemon: связь между клеточным циклом и половым размножением» . Растительная клетка . 7 (12): 1975–1988. дои : 10.1105/tpc.7.12.1975 . ISSN 1040-4651 . ПМК 161055 . ПМИД 12242365 .
- ^ Фридман, Уильям Э. (1990). «Половое размножение Ephedra nevadensis (Ephedraceae): дополнительные доказательства двойного оплодотворения нецветущих семенных растений». Американский журнал ботаники . 77 (12): 1582–1598. дои : 10.1002/j.1537-2197.1990.tb11399.x . JSTOR 2444491 .
- ^ Jump up to: а б Кармайкл, Джеффри С.; Фридман, Уильям Э. (1996). «Двойное оплодотворение у Gnetum gnemon (Gnetaceae): его влияние на эволюцию полового размножения внутри Gnetales и Anthophyte Clade». Американский журнал ботаники . 83 (6): 767–780. дои : 10.1002/j.1537-2197.1996.tb12766.x . JSTOR 2445854 .
- ^ Фридман, МЫ (25 апреля 1995 г.). «Дупликация организмов, инклюзивная теория приспособленности и альтруизм: понимание эволюции эндосперма и репродуктивного синдрома покрытосеменных» . Труды Национальной академии наук . 92 (9): 3913–3917. Бибкод : 1995PNAS...92.3913F . дои : 10.1073/pnas.92.9.3913 . ISSN 0027-8424 . ПМК 42072 . ПМИД 11607532 .
- ^ Фридман, Уильям Э. (1994). «Эволюция эмбриогении семенных растений, происхождение развития и ранняя история эндосперма». Американский журнал ботаники . 81 (11): 1468–1486. дои : 10.1002/j.1537-2197.1994.tb15633.x . JSTOR 2445320 .
- ^ Боу, Л. Мишель; Пальто, Гвенаэль; де Памфилис, Клод В. (11 апреля 2000 г.). «Филогения семенных растений на основе всех трех геномных компартментов: современные голосеменные растения монофилетичны, а ближайшие родственники Gnetales - хвойные» . Труды Национальной академии наук . 97 (8): 4092–4097. Бибкод : 2000PNAS...97.4092B . дои : 10.1073/pnas.97.8.4092 . ISSN 0027-8424 . ЧВК 18159 . ПМИД 10760278 .
- ^ Зима, Кай-Уве; Беккер, Аннетт; Мюнстер, Томас; Ким, Ян Т.; Сэдлер, Хайнц; Тайссен, Гюнтер (22 июня 1999 г.). «Гены MADS-бокса показывают, что гнетофиты более тесно связаны с хвойными, чем с цветковыми растениями» . Труды Национальной академии наук . 96 (13): 7342–7347. Бибкод : 1999PNAS...96.7342W . дои : 10.1073/pnas.96.13.7342 . ISSN 0027-8424 . ПМК 22087 . ПМИД 10377416 .
- ^ Магаллон, С.; Сандерсон, MJ (1 декабря 2002 г.). «Отношения между семенными растениями, выведенные на основе высококонсервативных генов: сортировка противоречивых филогенетических сигналов среди древних линий» . Американский журнал ботаники . 89 (12): 1991–2006. дои : 10.3732/ajb.89.12.1991 . ISSN 1537-2197 . ПМИД 21665628 .
- ^ Чау, Шу-Мио; Паркинсон, Кристофер Л.; Ченг, Юйчан; Винсент, Томас М.; Палмер, Джеффри Д. (11 апреля 2000 г.). «Филогения семенных растений, сделанная на основе всех трех геномов растений: монофилия современных голосеменных растений и происхождение Gnetales от хвойных» . Труды Национальной академии наук . 97 (8): 4086–4091. Бибкод : 2000PNAS...97.4086C . дои : 10.1073/pnas.97.8.4086 . ISSN 0027-8424 . ЧВК 18157 . ПМИД 10760277 .
- ^ Зенктелер, М. (1990). «Экстракорпоральное оплодотворение и широкая гибридизация высших растений». Crit Rev Plant Sci . 9 (3): 267–279. дои : 10.1080/07352689009382290 .
- ^ Рагхаван, В. (2005). Двойное оплодотворение: развитие зародыша и эндосперма у цветковых растений (иллюстрированное ред.). Биркхойзер. стр. 17–19. ISBN 978-3-540-27791-0 .
- ^ Кэмпбелл Н.А.; Рис Дж. Б. (2005). Биология (7-е изд.). Сан-Франциско, Калифорния: Pearson Education, Inc., стр. 774–777. ISBN 978-0-8053-7171-0 .
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растений Группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Воспроизведение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||