Мохообразные


Мохообразные ( / ˈ b r aɪ . ə ˌ f aɪ t s / ) [1] — группа наземных растений , иногда рассматриваемая как таксономическое подразделение , которое включает три группы несосудистых наземных растений (эмбриофитов): печеночники , роголистники и мхи . [2] В строгом смысле отдел Bryophyta состоит только из мхов. Мохообразные обычно ограничены в размерах и предпочитают влажную среду обитания, хотя некоторые виды могут выжить и в более засушливых условиях. [3] Мохообразные включают около 20 000 видов растений. [4] [5] Мохообразные образуют закрытые репродуктивные структуры (гаметангии и спорангии), но не образуют ни цветов , ни семян . Они размножаются половым путем спорами и бесполым путем фрагментации или образования гемм . [6] Хотя в последние годы мохообразные считались парафилетической группой, почти все самые последние филогенетические данные подтверждают монофилию этой группы, первоначально классифицированную Вильгельмом Шимпером в 1879 году. [7] Термин мохообразный происходит от древнегреческого βρύον ( brúon ) «древесный мох, печеночник» и φυτόν ( phutón ) «растение».
Функции
[ редактировать ]Отличительными особенностями мохообразных являются:
- В их жизненном цикле преобладает многоклеточного гаплоидного гаметофита. стадия
- Их спорофиты диплоидны . и неразветвлены
- У них нет настоящей сосудистой ткани , содержащей лигнин (хотя у некоторых есть специализированные ткани для транспорта воды). [8]
среда обитания
[ редактировать ]Мохообразные существуют в самых разных средах обитания. Их можно найти растущими в диапазоне температур (холодная Арктика и жаркие пустыни), возвышенностях (от уровня моря до альпийских гор) и влажности (от засушливых пустынь до влажных тропических лесов). Мохообразные могут расти там, где васкуляризированные растения не могут, потому что они не зависят от корней для поглощения питательных веществ из почвы . Мохообразные могут выжить на камнях и голой почве. [9]
Жизненный цикл
[ редактировать ]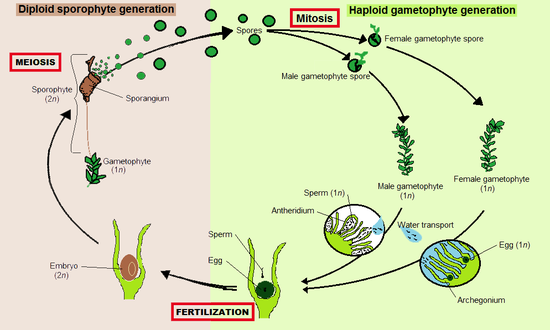
Как и все наземные растения (эмбриофиты), мохообразные имеют жизненный цикл со сменой поколений . [10] В каждом цикле гаплоидный гаметофит, каждая клетка которого содержит фиксированное число непарных хромосом , чередуется с диплоидным спорофитом, клетки которого содержат два набора парных хромосом. Гаметофиты производят гаплоидные сперматозоиды и яйцеклетки, которые сливаются с образованием диплоидных зигот, которые превращаются в спорофиты. производят гаплоидные споры Спорофиты путем мейоза , которые превращаются в гаметофиты.
Мохообразные являются доминирующими гаметофитами, [11] это означает, что более заметным и долгоживущим растением является гаплоидный гаметофит. Диплоидные спорофиты появляются лишь изредка и остаются прикрепленными к гаметофиту и питаются от него. [12] У мохообразных спорофиты всегда неразветвленные и образуют один спорангий (спорообразующую капсулу), но каждый гаметофит может дать начало сразу нескольким спорофитам.
Печеночники, мхи и роголистники большую часть жизни проводят в виде гаметофитов. Гаметангии (органы, производящие гаметы), архегонии и антеридии образуются на гаметофитах, иногда на кончиках побегов, в пазухах листьев или скрыты под слоевищами. Некоторые мохообразные, такие как печеночник Marchantia , создают сложные структуры, несущие гаметангии, называемые гаметангиофорами. Сперматозоиды имеют жгутики и должны плыть от антеридиев, из которых они производятся, к архегониям, которые могут находиться на другом растении. Членистоногие могут способствовать передаче спермы. [13]
Оплодотворенные яйца становятся зиготами, из которых внутри архегониев развиваются зародыши спорофитов. Зрелые спорофиты остаются прикрепленными к гаметофиту. Они состоят из ножки, называемой щетинкой, и одного спорангия или капсулы. Внутри спорангия в результате мейоза образуются гаплоидные споры . Они распространяются, чаще всего ветром, и, если они приземляются в подходящей среде, могут развиться в новый гаметофит. Таким образом, мохообразные распространяются путем сочетания плавающих сперматозоидов и спор, подобно ликофитам , папоротникам и другим криптогамам .
В трех группах спорофит развивается по-разному. И у мхов, и у роголистников есть меристемная зона, где происходит деление клеток. У роголистников меристема начинается у основания, где заканчивается ножка, и деление клеток толкает тело спорофита вверх. У мхов меристема расположена между капсулой и верхушкой стебля (щетинкой) и производит клетки вниз, удлиняя стебель и поднимая капсулу. У печеночников меристема отсутствует, и удлинение спорофита происходит почти исключительно за счет разрастания клеток. [14]
Сексуальность
[ редактировать ]Расположение антеридиев и архегониев на отдельном растении-мохо обычно постоянно внутри вида, хотя у некоторых видов оно может зависеть от условий среды. Основное разделение происходит между видами, у которых антеридии и архегонии встречаются на одном и том же растении, и видами, у которых они встречаются на разных растениях. Термин однодомный может использоваться, когда антеридии и архегонии встречаются на одном и том же гаметофите , и термин двудомный , когда они встречаются на разных гаметофитах. [15]
У семенных растений « однодомные употребляют », когда цветки с пыльниками (микроспорангии) и цветки с семязачатками (мегаспорангии) встречаются на одном и том же спорофите , и « двудомные », где они встречаются на разных спорофитах. Эти термины иногда могут использоваться вместо «моно- и двудомных» для описания гаметофитов мохообразных. «Однодомный» и «однодомный» происходят от греческого слова «один дом», «раздельнополый» и «двудомный» от греческого слова «два дома». Использование «-ойной» терминологии относится к гаметофитной сексуальности мохообразных в отличие от спорофитной сексуальности семенных растений. [15]
Однодомные растения обязательно являются гермафродитами, то есть одно и то же растение производит гаметы обоих полов. [15] Точное расположение антеридиев и архегониев у однодомных растений различно. Они могут располагаться на разных побегах (автоические), на одном и том же побеге, но не вместе в общей структуре (паройные или паройные) или вместе в общем «соцветии» (синойные или синойные). [15] [16] Двудомные растения однополые , то есть отдельное растение имеет только один пол. [15] Все четыре типа (автойный, паройный, синойный и двудомный) встречаются у видов мхов рода Bryum . [16]
Классификация и филогения
[ редактировать ]

Традиционно все живые наземные растения без сосудистых тканей относили к одной таксономической группе, часто к отделу (или типу). Термин «Мохообразные» впервые был предложен Брауном в 1864 году. [17] Еще в 1879 году термин Bryophyta был использован немецким бриологом Вильгельмом Шимпером для описания группы, содержащей все три клады мохообразных (хотя в то время роголистники считались частью печеночников). [18] [7] Г.М. Смит поместил эту группу между Algae и Pteridophyta . [19] Хотя исследование 2005 года подтвердило эту традиционную монофилетическую точку зрения, [20] к 2010 году среди систематиков сложился широкий консенсус в отношении того, что мохообразные в целом не являются естественной группой (т. е. являются парафилетическими ). [21] [22] [23] Однако исследование 2014 года пришло к выводу, что эти предыдущие филогении, основанные на последовательностях нуклеиновых кислот, подвержены систематическим ошибкам в составе, и что, кроме того, филогении, основанные на аминокислотных последовательностях, позволяют предположить, что мохообразные в конце концов являются монофилетическими. [24] С тех пор, отчасти благодаря увеличению наборов геномных и транскриптомных данных, почти все филогенетические исследования, основанные на ядерных и хлоропластических последовательностях, пришли к выводу, что мохообразные образуют монофилетическую группу. [24] [25] [18] [26] [27] [28] [29] [30] [31] Тем не менее, филогении, основанные на митохондриальных последовательностях, не подтверждают монофилетическую точку зрения. [32]
Три клады мохообразных - это Marchantiophyta (печеночники), Bryophyta (мхи) и Anthocerotophyta (роголистники). [33] Однако было предложено перевести эти клады в классы Marchantiopsida, Bryopsida и Anthocerotopsida соответственно. [18] В настоящее время имеются убедительные доказательства того, что печеночники и мхи принадлежат к монофилетической кладе Setaphyta . [25] [32] [34]
Монофилетический взгляд
[ редактировать ]Предпочтительная модель, основанная на филогении аминокислот, указывает на мохообразных как на монофилетическую группу: [24]
| эмбриофиты |
| ||||||
В соответствии с этой точкой зрения, по сравнению с другими живыми наземными растениями, во всех трех линиях отсутствует сосудистая ткань, содержащая лигнин, и разветвленные спорофиты, несущие множественные спорангии. Выдающееся положение гаметофита в жизненном цикле также является общей чертой трех линий мохообразных (все современные сосудистые растения имеют доминантные спорофиты). Однако если эта филогения верна, то сложный спорофит живых сосудистых растений мог развиться независимо от более простого неразветвленного спорофита, присутствующего у мохообразных. [24] Более того, эта точка зрения подразумевает, что устьица возникли только один раз в эволюции растений, а затем были потеряны в печеночниках. [25] [28]
Парафилетический взгляд
[ редактировать ]
| мохообразные |

В этом альтернативном взгляде группа Setaphyta сохраняется, но вместо этого роголистники являются сестрами сосудистых растений. [34] (Другая парафилетическая точка зрения предполагает, что роголистники сначала разветвляются.) [32]
Традиционная морфология
[ редактировать ]Традиционно при классификации по морфологическим признакам мохообразные отличались отсутствием сосудистой структуры. Однако это различие проблематично, во-первых, потому, что некоторые из самых ранних дивергентных (но теперь вымерших) небриофитов, таких как хорнеофиты , не имели настоящей сосудистой ткани, и, во-вторых, потому, что многие мхи имеют хорошо развитые водопроводящие сосуды. [35] [36] Более полезное различие может заключаться в строении их спорофитов . У мохообразных спорофит представляет собой простую неразветвленную структуру с единственным спорообразующим органом ( спорангием ), тогда как у всех других наземных растений — полиспорангиофитов — спорофит разветвлен и несет множество спорангиев. [37] [38] Контраст показан на кладограмме ниже: [39]
| Наземные растения |
| |||||||||||||||
Эволюция
[ редактировать ]Вероятно, произошло несколько различных событий террестриализации, в ходе которых первоначально водные организмы колонизировали сушу, как раз в пределах линии Viridiplantae . [40] Однако между 510–630 миллионами лет назад среди зеленых водорослей появились наземные растения . [41] Молекулярно-филогенетические исследования пришли к выводу, что мохообразные являются самыми ранними дивергентными линиями современных наземных растений. [42] [2] [43] [44] Они дают представление о миграции растений из водной среды на сушу. Ряд физических особенностей связывает мохообразных как с наземными, так и с водными растениями. [45]
Сходства с водорослями и сосудистыми растениями
[ редактировать ]Зеленые водоросли, мохообразные и сосудистые растения имеют хлорофилл a и b, а структуры хлоропластов схожи. [46] Подобно зеленым водорослям и наземным растениям, мохообразные также производят крахмал, хранящийся в пластидах, и содержат целлюлозу в своих стенках. [46] Определенные адаптации, наблюдаемые у мохообразных, позволили растениям колонизировать наземную среду Земли. Чтобы предотвратить высыхание тканей растения в наземной среде, может присутствовать восковая кутикула, покрывающая мягкие ткани растения и обеспечивающая защиту. У роголистников и мхов устьица обеспечивают газообмен между атмосферой и внутренним межклеточным пространством. Развитие гаметангиев обеспечило дополнительную защиту именно гамет, зиготы и развивающегося спорофита. [47] Мохообразные и сосудистые растения ( эмбриофиты ) также имеют эмбриональное развитие, чего не наблюдается у зеленых водорослей. [46] Хотя у мохообразных нет по-настоящему васкуляризированной ткани, у них есть органы, которые специализируются на транспортировке воды и выполнении других специфических функций, аналогичных, например, функциям листьев и стеблей сосудистых наземных растений. [46]
Мохообразные зависят от воды для размножения и выживания. Как и у папоротников и ликофитов , на поверхности растения необходим тонкий слой воды, чтобы обеспечить перемещение жгутиковых сперматозоидов между гаметофитами и оплодотворение яйцеклетки. [47]
Сравнительная морфология
[ редактировать ]Сводная морфологическая характеристика гаметофитов трех групп мохообразных:
| Печеночники | мхи | Роголистники | |
|---|---|---|---|
| Структура | Таллоидный или листоватый | Листовидный | Таллоид |
| Симметрия | Дорзивентральный или радиальный | Радиальный | Дорзивентральный |
| Ризоиды | Одноклеточный | Многоклеточный | Одноклеточный |
| Хлоропласты/клетка | Много | Много | Один |
| Протонемата | Уменьшенный | Подарок | Отсутствующий |
| Гаметангия ( антеридии и архегонии ) | Поверхностный | Поверхностный | Погруженный |
Сводная морфологическая характеристика спорофитов трех групп мохообразных:
| Печеночники | мхи | Роголистники | |
|---|---|---|---|
| устьица | Отсутствующий | Подарок | Подарок |
| Структура | Маленький, без хлорофилла | Большой, с хлорофиллом | Большой, с хлорофиллом |
| Упорство | Эфемерный | Настойчивый | Настойчивый |
| Рост | Определенный | Определенный | Непрерывный |
| Апикальный рост [48] | Отсутствующий | Подарок | Отсутствующий |
| Сета | Подарок | Подарок | Отсутствующий |
| Капсульная форма | Простой | Дифференцированный ( жаберная крышка , перистом ) | удлиненный |
| Созревание спор | Одновременный | Одновременный | Постепенный |
| Распространение спор | Элатерс | Перистомные зубы | Псевдо-элатеры |
| Колумелла | Отсутствующий | Подарок | Подарок |
| Раскрытие | Продольный или неравномерный | поперечный | Продольный |
Использование
[ редактировать ]Относящийся к окружающей среде
- Кондиционирование почвы
- Биоиндикаторы
- Моховые сады
- Пестициды
Характеристики мохообразных делают их полезными для окружающей среды. Было показано, что в зависимости от конкретной текстуры растения мохообразные помогают улучшить удержание воды и воздушное пространство в почве. [49] Мохообразные используются в исследованиях загрязнения, чтобы указать на загрязнение почвы (например, наличие тяжелых металлов), загрязнение воздуха и УФ-B-излучение. [49] Сады в Японии созданы из мха, чтобы создать умиротворяющее убежище. [49] Было обнаружено, что некоторые мохообразные производят природные пестициды. Печеночник Plagiochila производит химическое вещество, ядовитое для мышей. [49] Другие мохообразные производят химические вещества, являющиеся антифидантами, которые защищают их от поедания слизняками. [49] Когда Phythium sphagnum посыпают почву прорастающими семенами, он подавляет рост «затухающих грибов», которые в противном случае могли бы убить молодые всходы. [50]

Коммерческий
- Топливо
- Упаковка
- Повязка на рану
Торф — это топливо, получаемое из высушенных мохообразных, обычно сфагнума . Антибиотические свойства мохообразных и способность удерживать воду делают их полезным упаковочным материалом для овощей, цветов и луковиц. [49] Кроме того, из-за своих антибиотических свойств сфагнум использовался в качестве хирургической повязки во время Первой мировой войны. [49]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «бриофит» . Dictionary.com Полный (онлайн). nd
- ^ Jump up to: а б Хеджес, С. Блэр (ноябрь 2002 г.). «Происхождение и эволюция модельных организмов». Обзоры природы Генетика . 3 (11): 838–849. дои : 10.1038/nrg929 . ПМИД 12415314 . S2CID 10956647 .
- ^ Леветин, Эстель; МакМахон, Карен (2012). Растения и общество . Нью-Йорк, штат Нью-Йорк: МакГроу-Хилл. п. 139. ИСБН 978-0-07-352422-1 .
- ^ «Мохообразные (мхи и печеночники) — Список растений» . theplantlist.org . Проверено 11 апреля 2017 г.
- ^ «Что такое мохообразные» . Университет Южного Иллинойса в Карбондейле . Архивировано из оригинала 19 октября 2018 г. Проверено 31 мая 2009 г.
- ^ Вандерпоортен, Ален; Гоффине, Бернар (2009). Знакомство с мохообразными . Кембридж: Издательство Кембриджского университета . п. 3. ISBN 978-0-511-54013-4 .
- ^ Jump up to: а б Шимпер, WP (1879). «Мохообразные». В Зиттеле, К.А. (ред.). Справочник по палеонтологии . Том 2. Р. Ольденбург.
- ^ Лукас, Уильям Дж.; Грувер, Эндрю; Лихтенбергер, Рафаэль; Фурута, Каори; Ядав, Шри-Рам; Хелариутта, Юка; Он, Синь-Цян; Фукуда, Хироо; Канг, Джули; Брэди, Шивон М .; Патрик, Джон В. (апрель 2013 г.). «Сосудистая система растений: эволюция, развитие и функции F». Журнал интегративной биологии растений . 55 (4): 294–388. дои : 10.1111/jipb.12041 . hdl : 10261/76903 . ПМИД 23462277 .
- ^ Лепп, Хейно (28 февраля 2008 г.). «Места обитания» . Австралийские мохообразные . Австралийский национальный ботанический сад.
- ^ Лигроне, Роберто; Дакетт, Джеффри Г.; Ренцалья, Карен С. (апрель 2012 г.). «Основные переходы в эволюции ранних наземных растений: бриологический взгляд» . Анналы ботаники . 109 (5): 851–871. дои : 10.1093/aob/mcs017 . ПМК 3310499 . ПМИД 22356739 .
- ^ CM Шон Кэррингтон (04 ноября 2013 г.). «Мохообразные» . Университет Вест-Индии. Архивировано из оригинала 5 февраля 2020 г. Проверено 05 марта 2020 г.
- ^ Джонсон, Миннесота; Шоу, AJ (24 февраля 2016 г.). «Влияние количественной плодовитости на гаплоидной стадии на репродуктивный успех и диплоидную приспособленность водного торфяного мха Sphagnum macrophyllum» . Наследственность . 116 (6): 523–530. дои : 10.1038/hdy.2016.13 . ПМЦ 4868265 . ПМИД 26905464 .
- ^ Кронберг, Н.; Начева Р.; Хедлунд, К. (2006). «Микроартроподы опосредуют передачу спермы у мхов». Наука . 313 (5791): 1255. doi : 10.1126/science.1128707 . ПМИД 16946062 . S2CID 11555211 .
- ^ Ули Гроссниклаус (2019). Развитие и эволюция растений . Академическая пресса. п. 13. ISBN 9780128098059 .
- ^ Jump up to: а б с д и Глайм, Дж. М. и Бисанг, И. (2014). «Сексуальность: ее определение (гл. 3-1)» (PDF) . В Glime, JM (ред.). Экология мохообразных . Том. 1 Физиологическая экология. Мичиганский технологический университет и Международная ассоциация бриологов . Проверено 9 ноября 2014 г.
- ^ Jump up to: а б Уотсон, Э.В. (1981). Британские мхи и печеночники (3-е изд.). Издательство Кембриджского университета. п. 7. ISBN 9780521285360 . (Ватсон использует термины «ойси», а не термины «ойси».)
- ^ «Тип Bryophyta – Иерархия» . Таксономикон.
- ^ Jump up to: а б с де Соуза, Филипе; и др. (2019). «Филогения ядерных белков поддерживает монофилию трех групп мохообразных (Bryophyta Schimp.)». Новый фитолог . 222 (1): 565–575. дои : 10.1111/nph.15587 . hdl : 1983/0b471d7e-ce54-4681-b791-1da305d9e53b . ПМИД 30411803 . S2CID 53240320 .
- ^ Смит, генеральный директор (1955). Криптогаминая ботаника . Том. 2 (2-е изд.). Нью-Йорк: МакГроу-Хилл.
- ^ Горемыкин В.В. и Хеллвиг Ф.Х. (2005). «Доказательства самого базального разделения наземных растений, разделяющих линии мохообразных и трахеофитов». Систематика и эволюция растений . 254 (1–2): 93–103. дои : 10.1007/s00606-005-0337-1 . S2CID 41403901 .
- ^ Конрат, М.; Шоу, Эй Джей; Рензалья, Канзас (2010). «Специальный выпуск Phytotaxa, посвященный мохообразным: ближайшим ныне живущим родственникам ранних наземных растений» . Фитотаксы . 9 :5–10. дои : 10.11646/phytotaxa.9.1.3 .
- ^ Троицкий А.В.; Игнатов, М.С.; Боброва В.К.; Милютина И.А. (декабрь 2007 г.). «Вклад геносистематики в современные представления о филогении и классификации мохообразных». Биохимия (Москва) . 72 (12): 1368–1376. дои : 10.1134/s0006297907120115 . ПМИД 18205621 . S2CID 13509400 .
- ^ Кнуп, Волкер (31 декабря 2010 г.). «В поисках смысла в ерунде: краткий обзор некодирующей органеллярной ДНК, объясняющей филогению мохообразных». Разнообразие и эволюция мохообразных . 31 (1): 51–60. дои : 10.11646/bde.31.1.10 .
- ^ Jump up to: а б с д Кокс, Саймон Дж.; и др. (2014). «Противоречивые филогении ранних наземных растений вызваны предвзятостью состава синонимических замен» . Систематическая биология . 63 (2): 272–279. дои : 10.1093/sysbio/syt109 . ПМЦ 3926305 . ПМИД 24399481 .
- ^ Jump up to: а б с Путтик, Марк Н.; и др. (2018). «Взаимоотношения наземных растений и природа предкового эмбриофита». Современная биология . 28 (5): 733–745.e2. дои : 10.1016/j.cub.2018.01.063 . hdl : 1983/ad32d4da-6cb3-4ed6-add2-2415f81b46da . ПМИД 29456145 . S2CID 3269165 .
- ^ Либенс-Мак, Джеймс Х.; и др. (2019). «Тысяча растительных транскриптомов и филогеномика зеленых растений» . Природа . 574 (7780): 679–685. дои : 10.1038/s41586-019-1693-2 . ПМК 6872490 . PMID 31645766 .
- ^ Чжан, Цзянь; и др. (2020). «Геном роголистника и ранняя эволюция наземных растений» . Природные растения . 6 (2): 107–118. дои : 10.1038/s41477-019-0588-4 . ПМК 7027989 . ПМИД 32042158 .
- ^ Jump up to: а б Харрис, Броган Дж.; и др. (2020). «Филогеномные доказательства монофилии мохообразных и редуктивной эволюции устьиц». Современная биология . 30 (11): P2201–2012.E2. дои : 10.1016/j.cub.2020.03.048 . hdl : 1983/fbf3f371-8085-4e76-9342-e3b326e69edd . ПМИД 32302587 . S2CID 215798377 .
- ^ Ли, Фэй-Вэй; и др. (2020). «Геномы Anthoceros проливают свет на происхождение наземных растений и уникальную биологию роголистников» . Природные растения . 6 (3): 259–272. дои : 10.1038/s41477-020-0618-2 . hdl : 10261/234303 . ПМЦ 8075897 . ПМИД 32170292 .
- ^ Соуза, Филипе; и др. (2020). «Филогения наземных растений хлоропластов: анализ с использованием более подходящих моделей неоднородного состава деревьев и участков» . Границы в науке о растениях . 11 : 1062. дои : 10.3389/fpls.2020.01062 . ПМЦ 7373204 . ПМИД 32760416 .
- ^ Су, Данян; и др. (2021). «Крупномасштабный филогеномный анализ выявляет монофилию мохообразных и неопротерозойское происхождение наземных растений» . Молекулярная биология и эволюция . 38 (8): 3332–3344. дои : 10.1093/molbev/msab106 . ПМЦ 8321542 . ПМИД 33871608 .
- ^ Jump up to: а б с Соуза, Филипе; и др. (2020). «Митохондриальная филогения наземных растений демонстрирует поддержку Setaphyta в рамках моделей гетерогенного по составу замещения» . ПерДж . 8 (4): е8995. дои : 10.7717/peerj.8995 . ПМК 7194085 . ПМИД 32377448 .
- ^ «ГЛОССАРИЙ Б» . Архивировано из оригинала 2 апреля 2009 г. Проверено 26 марта 2009 г.
- ^ Jump up to: а б Кокс, Саймон Дж. (2018). «Молекулярная филогенетика наземных растений: обзор с комментариями по оценке несоответствия между филогениями». Критические обзоры по наукам о растениях . 37 (2–3): 113–127. дои : 10.1080/07352689.2018.1482443 . hdl : 10400.1/14557 . S2CID 92198979 .
- ^ Белл, Н. Э. и Хивонен, Дж. (2010). «Филогения класса мхов Polytrichopsida (BRYOPHYTA): структура родового уровня и несоответствующие генные деревья» . Молекулярная филогенетика и эволюция . 55 (2): 381–398. дои : 10.1016/j.ympev.2010.02.004 . ПМИД 20152915 .
- ^ Бодрибб, Ти Джей; и др. (2020). «У широко распространенного мха обнаружена расширенная сосудистая функция». Природные растения . 6 (3): 273–279. дои : 10.1038/s41477-020-0602-x . ПМИД 32170283 . S2CID 212641738 .
- ^ Кенрик, Пол и Крейн, Питер Р. (1997a). Происхождение и ранняя диверсификация наземных растений: кладистическое исследование . Вашингтон, округ Колумбия: Издательство Смитсоновского института. ISBN 978-1-56098-730-7 .
- ^ Кенрик, П. и Крейн, PR (1997b). «Происхождение и ранняя эволюция растений на суше». Природа . 389 (6646): 33–39. Бибкод : 1997Natur.389...33K . дои : 10.1038/37918 . S2CID 3866183 .
- ^ Крейн, Питер Р.; Херендин, Патрик; Фриис, Эльза Мари (октябрь 2004 г.). «Окаменелости и филогения растений» . Американский журнал ботаники . 91 (10): 1683–1699. дои : 10.3732/ajb.91.10.1683 . ПМИД 21652317 .
- ^ Жерриен, Филипп; Серве, Томас; Веколи, Марко (апрель 2016 г.). «Эволюция растений и террестриализация в палеозойскую эпоху - филогенетический контекст». Обзор палеоботаники и палинологии . 227 : 4–18. дои : 10.1016/j.revpalbo.2016.01.004 .
- ^ Делвич, Чарльз Ф.; Тимм, Рут Э. (7 июня 2011 г.). «Растения» . Современная биология . 21 (11): Р417–Р422. дои : 10.1016/j.cub.2011.04.021 . ISSN 0960-9822 . ПМИД 21640897 . S2CID 235312105 .
- ^ Конрат, М.; Шоу, Эй Джей; Рензалья, Канзас (2010). «Специальный выпуск Phytotaxa, посвященный мохообразным: ближайшим ныне живущим родственникам ранних наземных растений» . Фитотаксы . 9 :5–10. дои : 10.11646/phytotaxa.9.1.3 .
- ^ Карол, Кеннет Г.; Арумуганатан, Катираветпиллай; Бур, Джеффри Л.; Даффи, Аарон М.; Эверетт, Карин Д.Э.; Холл, Джон Д.; Хансен, СК; Кюль, Дженнифер В.; Мандоли, Дина Ф.; Мишлер, Брент Д.; Олмстед, Ричард Г.; Рензалья, Карен С. и Вольф, Пол Г. (2010). «Полные последовательности пластома Equisetum arvense и Isoetes flaccida: значение для филогении и эволюции пластидного генома ранних линий наземных растений» . Эволюционная биология BMC . 10 (1): 321. дои : 10.1186/1471-2148-10-321 . ПМК 3087542 . ПМИД 20969798 .
- ^ Шоу, А. Джонатан; Шовеньи, Питер; Шоу, Бланка (март 2011 г.). «Разнообразие и эволюция мохообразных: окна в раннюю эволюцию наземных растений». Американский журнал ботаники . 98 (3): 352–369. дои : 10.3732/ajb.1000316 . ПМИД 21613131 .
- ^ Донохью, Филип CJ; Харрисон, К. Джилл; Папс, Хорди; Шнайдер, Харальд (11 октября 2021 г.). «Эволюционное возникновение наземных растений» . Современная биология . 31 (19): R1281–R1298. дои : 10.1016/j.cub.2021.07.038 . hdl : 1983/662d176e-fcf4-40bf-aa8c-5694a86bd41d . ПМИД 34637740 . S2CID 238588736 .
- ^ Jump up to: а б с д Эверт, Рэй; Эйххорн, Сьюзен (2013). Биология растений . Издатели WHFreeman and Company.
- ^ Jump up to: а б Перселл, Адам. «Мохообразные» . Базовая биология .
- ^ Как регулировался верхушечный рост у наземного растения предков? Результаты разработки несеменных растений
- ^ Jump up to: а б с д и ж г Глайм, Дженис. «Экономическое и этническое использование мохообразных» (PDF) . Гарвард.edu .
- ^ Вольфхехель, Х. (апрель 1988 г.). «Подавляющее действие сфагнового торфа на виды Pythium ». Acta Horticulturae (221): 217–222. дои : 10.17660/actahortic.1988.221.22 .
Библиография
[ редактировать ]- Лесица, П.; МакКьюн, Б.; Купер, СВ; Хонг, WS (1991). «Различия в сообществах лишайников и мохообразных между старовозрастными и управляемыми вторичными лесами в Лебединой долине, штат Монтана». Канадский журнал ботаники . 69 (8): 1745–1755. дои : 10.1139/b91-222 .
Внешние ссылки
[ редактировать ]- Сайт Эндрюс Мосс Фотографии мохообразных
- 27 мая 2013 г. Возрождены многовековые замороженные растения 400-летние образцы мохообразных, оставленные отступающими ледниками в Канаде. , в лаборатории возвращены к жизни
- Фардж, Кэтрин Ла; Уильямс, Криста Х.; Англия, Джон Х. (22 мая 2013 г.). «Регенерация мохообразных Малого ледникового периода, появляющихся из полярного ледника, с последствиями тотипотентности в экстремальных условиях» . Труды Национальной академии наук . 110 (24): 9839–9844. Бибкод : 2013PNAS..110.9839L . дои : 10.1073/pnas.1304199110 . ПМЦ 3683725 . ПМИД 23716658 .
- Мэгилл, RE, изд. (1990). Glossarium multiglottum bryologiae. Многоязычный глоссарий по бриологии . Монографии по систематической ботанике из Ботанического сада Миссури, т. 33, 297 стр. Интернет-версия: Интернет-архив .
Классификация Archaeplastida или Plantae sl |
|---|