Партеногенез

Партеногенез ( / ˌ p ɑːr θ ɪ n oʊ ˈ dʒ ɛ n ɪ s ɪ s , - θ ɪ n ə -/ ; [ 1 ] [ 2 ] от греческого parthenos , , «девственница» + génesis génesis , «творение». [ 3 ] ) — естественная форма бесполого размножения , при которой рост и развитие зародыша происходят непосредственно из яйцеклетки , без необходимости оплодотворения . [ 4 ] [ 5 ] У животных партеногенез означает развитие зародыша из неоплодотворенной яйцеклетки . У растений партеногенез является составной частью процесса апомиксиса . У водорослей партеногенез может означать развитие зародыша либо из отдельного спермия, либо из отдельной яйцеклетки. [ 4 ]
Партеногенез происходит естественным путем у некоторых растений, водорослей , беспозвоночных видов животных (включая нематод , некоторых тихоходок , водяных блох , некоторых скорпионов , тли , некоторых клещей, некоторых пчел , некоторых фазматод и паразитических ос ) и некоторых позвоночных животных (например, некоторых рыб) . , [ 6 ] амфибии , рептилии , [ 7 ] [ 8 ] [ 9 ] и птицы [ 10 ] [ 11 ] [ 12 ] ). Этот тип размножения был искусственно вызван у ряда видов животных, которые естественным образом размножаются половым путем, включая рыб, амфибий и мышей. [ 13 ] [ 14 ]
Нормальные яйцеклетки образуются в процессе мейоза и являются гаплоидными , с вдвое меньшим количеством хромосом, чем клетки тела матери. Однако гаплоидные особи обычно нежизнеспособны, а партеногенетическое потомство обычно имеет диплоидный набор хромосом. В зависимости от механизма восстановления диплоидного числа хромосом партеногенетическое потомство может иметь от всех до половины материнских аллелей . При некоторых типах партеногенеза потомство, имеющее весь генетический материал матери, называется полными клонами , а потомство, имеющее только половину, - полуклонами. Полные клоны обычно образуются без мейоза. Если происходит мейоз, потомство получит только часть аллелей матери, поскольку кроссинговер ДНК во время мейоза происходит , создавая вариации.
Партеногенетическое потомство видов, использующих либо XY , либо X0, систему определения пола имеет две Х-хромосомы и является самкой. У видов, использующих систему определения пола ZW , они имеют либо две Z-хромосомы (самцы), либо две W-хромосомы (в основном нежизнеспособные, но редко самки), либо они могут иметь одну Z и одну W-хромосому (самки).
Типы истории жизни
[ редактировать ]
Некоторые виды размножаются исключительно партеногенезом (например, бделлоидные коловратки ), тогда как другие могут переключаться между половым размножением и партеногенезом. Это называется факультативный партеногенез (другие термины — циклический партеногенез, гетерогамия). [ 15 ] [ 16 ] или гетерогония [ 17 ] [ 18 ] ). Переключение между сексуальностью и партеногенезом у таких видов может быть вызвано сезоном ( тля , некоторые галловые осы ), или отсутствием самцов, или условиями, благоприятствующими быстрому росту популяции ( коловратки и ветвистоусые типа дафний ). У этих видов бесполое размножение происходит либо летом (тля), либо при благоприятных условиях. Это связано с тем, что при бесполом размножении успешный генотип может быстро распространяться без изменения пола или траты ресурсов на потомство мужского пола, которое не будет рожать. Некоторые виды могут производить потомство как половым путем, так и посредством партеногенеза, а потомство в одной кладке тропической ящерицы может представлять собой смесь потомства, полученного половым путем, и потомства, полученного партеногенным путем. [ 19 ] У калифорнийских кондоров факультативный партеногенез может происходить даже тогда, когда самец присутствует и доступен самке для размножения. [ 20 ] Во время стресса потомство, полученное половым размножением, может оказаться более приспособленным, поскольку у него есть новые, возможно, полезные комбинации генов. Кроме того, половое размножение обеспечивает мейотическую рекомбинацию между несестринскими хромосомами — процесс, связанный с восстановлением ДНК и других повреждений ДНК, которые могут быть вызваны стрессовыми условиями. двухцепочечных разрывов [ 21 ]
Многие таксоны с гетерогонией содержат виды, утратившие половую фазу и теперь полностью бесполые. Многие другие случаи облигатного партеногенеза (или гиногенеза) встречаются среди полиплоидов и гибридов, у которых хромосомы не могут спариваться для мейоза. [ нужна ссылка ]
Производство потомства женского пола путем партеногенеза называется телитокией (например, у тли), тогда как образование самцов путем партеногенеза называется арренотокией (например, у пчел). Когда из неоплодотворенных яиц развиваются как самцы, так и самки, это явление называется дейтеротокией. [ 22 ]
Виды и механизмы
[ редактировать ]Партеногенез может происходить без мейоза посредством митотического оогенеза. Это называется апомиктическим партеногенезом . Зрелые яйцеклетки образуются в результате митотических делений, и эти клетки непосредственно превращаются в эмбрионы. У цветковых растений клетки гаметофита этому процессу могут подвергаться . Потомство, полученное в результате апомиктического партеногенеза, является полными клонами своей матери. Примеры включают тлю. [ нужна ссылка ]
Партеногенез с участием мейоза более сложен. В некоторых случаях потомство гаплоидное (например, самцы муравьев ) . В других случаях, называемых аутомиктическим партеногенезом , плоидность восстанавливается до диплоидии различными способами. Это связано с тем, что у большинства видов гаплоидные особи нежизнеспособны. При аутомиктическом партеногенезе потомство отличается друг от друга и от матери. Их называют полуклонами своей матери. [ нужна ссылка ]
Автомиктический
[ редактировать ]
Автомикс [ 23 ] это термин, охватывающий несколько репродуктивных механизмов, некоторые из которых являются партеногенетическими. [ 24 ]
Диплоидия может быть восстановлена путем удвоения хромосом без деления клеток до начала мейоза или после его завершения. Это называется эндомитотическим циклом. Это также может произойти за счет слияния первых двух бластомеров . Другие виды восстанавливают свою плоидность за счет слияния продуктов мейоза. Хромосомы могут не разделиться в одной из двух анафаз (так называемый реституционный мейоз ), или образовавшиеся ядра могут слиться, или одно из полярных телец может слиться с яйцеклеткой на каком-то этапе ее созревания. [ нужна ссылка ]
Некоторые авторы считают все формы аутомиксиса половыми, поскольку они связаны с рекомбинацией. Многие другие классифицируют эндомитотические варианты как бесполые и считают образующиеся эмбрионы партеногенетическими. У этих авторов порог отнесения аутомиксиса к половому процессу зависит от того, когда соединяются продукты анафазы I или анафазы II. Критерий «сексуальности» варьируется во всех случаях реституционного мейоза. [ 25 ] к тем, где ядра сливаются, или только к тем, где гаметы созревают к моменту слияния. [ 24 ] сравниваются с самооплодотворением . Те случаи аутомиксиса, которые относят к половому размножению , по механизму и последствиям [ нужна ссылка ]
Генетический состав потомства зависит от того, какой тип аутомиксиса имеет место. Когда эндомитоз возникает до мейоза [ 26 ] [ 27 ] или при центральном слиянии (реституционный мейоз анафазы I или слияние его продуктов) потомство получает все [ 26 ] [ 28 ] более половины генетического материала матери и преимущественно сохраняется гетерозиготность [ 29 ] (если у матери есть два аллеля локуса, вполне вероятно, что потомство унаследует оба). Это связано с тем, что в анафазе I гомологичные хромосомы разделяются. Гетерозиготность не сохраняется полностью при кроссинговере при центральном слиянии. [ 30 ] В случае премейотического удвоения рекомбинация, если она и происходит, происходит между идентичными сестринскими хроматидами. [ 26 ]
Если происходит терминальное слияние (реституционный мейоз анафазы II или слияние его продуктов), в потомстве присутствует немногим более половины генетического материала матери, и потомство в основном гомозиготно. [ 31 ] Это связано с тем, что в анафазе II сестринские хроматиды разделяются, и любая гетерозиготность возникает из-за кроссинговера. В случае эндомитоза после мейоза потомство полностью гомозиготно и имеет только половину генетического материала матери.
Это может привести к тому, что партеногенетическое потомство будет отличаться друг от друга и от своей матери. [ нужна ссылка ]
Пол потомства
[ редактировать ]При апомиктическом партеногенезе потомство представляет собой клоны матери и, следовательно (за исключением тлей), обычно является самками. В случае тли партеногенетически полученные самцы и самки представляют собой клоны своей матери, за исключением того, что у самцов отсутствует одна из Х-хромосом (ХО). [ 32 ]
При мейозе пол потомства будет зависеть от типа системы определения пола и типа апомиксиса. У видов, использующих систему определения пола XY , партеногенетическое потомство будет иметь две Х-хромосомы и быть самками. У видов, использующих систему определения пола ZW, генотип потомства может быть одним из ZW (самка), [ 28 ] [ 29 ] ZZ (самец), или WW (нежизнеспособен у большинства видов, [ 31 ] но плодородный, [ сомнительно – обсудить ] жизнеспособная самка у немногих (например, удавов ). [ 31 ] Потомство ZW производится путем эндорепликации перед мейозом или путем центрального слияния. [ 28 ] [ 29 ] Потомство ZZ и WW происходит либо путем терминального слияния [ 31 ] или эндомитозом яйцеклетки.
У полиплоидных облигатных партеногенов, таких как ящерица-хлыстохвост, все потомство бывает женского пола. [ 27 ]
У многих перепончатокрылых насекомых, таких как медоносные пчелы, женские яйца производятся половым путем с использованием спермы отца-трутня, в то время как производство дальнейших трутней (самцов) зависит от того, что матка (а иногда и рабочие) производит неоплодотворенные яйца. Это означает, что самки (рабочие и королевы) всегда диплоидны, а самцы (трутни) всегда гаплоидны и производятся партеногенетически.
Факультативный
[ редактировать ]Факультативный партеногенез — это термин, обозначающий ситуацию, когда самка может производить потомство половым или бесполым путем. [ 33 ] Факультативный партеногенез встречается в природе крайне редко, и лишь несколько примеров таксонов животных способны к факультативному партеногенезу. [ 33 ] Одним из наиболее известных примеров таксонов, демонстрирующих факультативный партеногенез, являются подёнки ; предположительно, это стандартный способ размножения всех видов этого отряда насекомых. [ 34 ] Обычно считается, что факультативный партеногенез является реакцией на отсутствие жизнеспособного самца. Самка может подвергнуться факультативному партеногенезу, если самец отсутствует в среде обитания или если он не может произвести жизнеспособное потомство. Однако калифорнийские кондоры и тропическая ящерица Lepidophyma smithii могут производить партеногенное потомство в присутствии самцов, что указывает на то, что факультативный партеногенез может быть более распространенным, чем считалось ранее, и не является просто реакцией на отсутствие самцов. [ 19 ] [ 11 ]
У тлей поколение, зачатое половым путем самцом и самкой, производит только самок. Причиной этого является неслучайное разделение половых хромосом «Х» и «О» во время сперматогенеза . [ 35 ]
Факультативный партеногенез часто используется для описания случаев спонтанного партеногенеза у нормально половых животных. [ 36 ] Например, многие случаи спонтанного партеногенеза у акул , некоторых змей , варанов Комодо и множества домашних птиц широко приписывались факультативному партеногенезу. [ 37 ] Эти случаи являются примерами спонтанного партеногенеза. [ 33 ] [ 36 ] Появление таких бесполых яиц у половых животных можно объяснить мейотической ошибкой, приводящей к образованию яиц посредством аутомиксиса . [ 36 ] [ 38 ]
Обязать
[ редактировать ]Облигатный партеногенез — это процесс, при котором организмы размножаются исключительно бесполым путем. [ 39 ] Было показано, что многие виды с течением времени переходят к облигатному партеногенезу. Хорошо документированные переходы к облигатному партеногенезу были обнаружены у многих таксонов многоклеточных животных, хотя и с помощью весьма разнообразных механизмов. Эти переходы часто происходят в результате инбридинга или мутации внутри больших популяций. [ 40 ] Существует ряд зарегистрированных видов, особенно саламандры и гекконы, которые полагаются на облигатный партеногенез как на основной метод размножения. Таким образом, в природе существует более 80 видов однополых рептилий (в основном ящериц, но включая один вид змей), земноводных и рыб, у которых самцы больше не являются частью репродуктивного процесса. [ 41 ] Самка производит яйцеклетку с полным набором (два набора генов), предоставленным исключительно матерью. Таким образом, самцу не требуется предоставлять сперму для оплодотворения яйцеклетки. Считается, что в некоторых случаях эта форма бесполого размножения представляет собой серьезную угрозу биоразнообразию из-за последующего отсутствия изменчивости генов и потенциально снижения приспособленности потомства. [ 39 ]
Обнаружено, что некоторые виды беспозвоночных, которые имеют (частичное) половое размножение в своем естественном ареале, размножаются исключительно путем партеногенеза в районах, в которые они были интродуцированы . [ 42 ] [ 43 ] Опора исключительно на партеногенетическое размножение имеет несколько преимуществ для инвазивных видов : это избавляет особей в очень редкой исходной популяции от необходимости искать себе пару; а распределение исключительно женского пола позволяет населению размножаться и вторгаться быстрее (потенциально в два раза быстрее). Примеры включают несколько тли . видов [ 42 ] и ивовый пилильщик Nematus oligospilus , который является половым в своей естественной среде обитания в Голарктике, но партеногенетическим там, где он был завезен в Южное полушарие. [ 43 ]
Естественное явление
[ редактировать ]Партеногенез не распространяется на изогамные виды. [ 44 ] Видно, что партеногенез происходит в природе у тлей , дафний , коловраток , нематод и некоторых других беспозвоночных, а также у многих растений. Среди позвоночных строгий партеногенез известен только у ящериц, змей, [ 45 ] птицы, [ 46 ] и акулы, [ 47 ] у рыб, амфибий и рептилий наблюдаются различные формы гиногенеза и гибридогенеза (неполная форма партеногенеза). [ 48 ] Первое женское (однополое) размножение позвоночных было описано у рыб Poecilia formosa в 1932 году. [ 49 ] С тех пор было описано не менее 50 видов однополых позвоночных, в том числе не менее 20 рыб, 25 ящериц, один вид змей, лягушек и саламандр. [ 48 ] Другие обычно половые виды могут иногда размножаться партеногенетически; Дракон Комодо , акула-молот и черноперая акула - недавние дополнения к известному списку спонтанно партеногенетических позвоночных. Как и во всех типах бесполого размножения , с партеногенезом связаны как издержки (низкое генетическое разнообразие и, следовательно, восприимчивость к неблагоприятным мутациям), так и выгоды (воспроизведение без необходимости в самце).
Партеногенез отличается от искусственного клонирования животных — процесса, при котором новый организм обязательно генетически идентичен клеткам-донорам. При клонировании ядро диплоидной , в результате чего образуется организм , клетки донорского организма вставляется в энуклеированную яйцеклетку, а затем клетка стимулируется к продолжению митоза генетически идентичный донору. Партеногенез отличается тем, что он возникает из генетического материала, содержащегося в яйцеклетке, и новый организм не обязательно генетически идентичен родительскому.
Партеногенез может быть достигнут посредством искусственного процесса, как описано ниже при обсуждении млекопитающих.
Оомицеты
[ редактировать ]Апомиксис, по-видимому, может возникнуть у Phytophthora . [ 50 ] оомицет . Ооспоры от экспериментального скрещивания прорастали, и часть потомства была генетически идентична тому или иному родителю, что означает, что мейоза не произошло и ооспоры развивались путем партеногенеза.
Бархатные черви
[ редактировать ]Самцов Epiperipatus imthurni не обнаружено, а экземпляры из Тринидада размножаются партеногенетически. Этот вид — единственный известный бархатный червь , размножающийся партеногенезом. [ 51 ]
Коловушки
[ редактировать ]У бделлоидных коловраток самки размножаются исключительно партеногенезом (облигатный партеногенез), [ 52 ] у моногононтных коловраток самки могут чередовать половое и бесполое размножение (циклический партеногенез). По крайней мере, у одного нормально цикличного партеногенетического вида может передаваться по наследству облигатный партеногенез: рецессивный аллель приводит к потере полового размножения у гомозиготного потомства. [ 53 ]
Плоские черви
[ редактировать ]По крайней мере, два вида рода Dugesia , плоских червей подразделения Platyhelminthes Turbellaria типа , включают полиплоидных особей , которые размножаются путем партеногенеза. [ 54 ] Этот тип партеногенеза требует спаривания, но сперма не участвует в генетике потомства (партеногенез псевдогамный , иначе называемый гиногенетическим). Сложный цикл спаривания диплоидных половых и полиплоидных партеногенетических особей приводит к образованию новых партеногенетических линий.
Улитки
[ редактировать ]Несколько видов партеногенетических брюхоногих моллюсков были изучены, особенно в отношении их статуса инвазивных видов . К таким видам относятся новозеландская грязевая улитка ( Potamopyrgus antipodarum ), [ 55 ] мелания с красной каймой ( Melanoides Tuberculata ), [ 56 ] и Мелания Стеганая ( Tarebia granifera ). [ 57 ]
Насекомые
[ редактировать ]Партеногенез у насекомых может охватывать широкий спектр механизмов. [ 58 ] Потомство, образующееся в результате партеногенеза, может быть обоих полов, только женского пола ( телитоки , например, у тлей и некоторых перепончатокрылых) . [ 59 ] ) или только самцы ( арренотокии , например, у большинства перепончатокрылых ). как истинный партеногенез, так и псевдогамия ( гиногенез или спермозависимый партеногенез ). Известно, что встречаются [ 33 ] Яйцеклетки, в зависимости от вида, могут образовываться без мейоза (апомиктически) или с помощью одного из нескольких аутомиктических механизмов.
Связанное с этим явление, полиэмбриония, представляет собой процесс, в результате которого из одной яйцеклетки образуется множественное клональное потомство. Это известно у некоторых перепончатокрылых паразитоидов и у Strepsiptera . [ 58 ]
У аутомиктических видов потомство может быть гаплоидным или диплоидным. Диплоиды образуются путем удвоения или слияния гамет после мейоза. Слияние наблюдается у Phasmatodea , Hemiptera ( Aleurodids и Coccidae ), Diptera и некоторых перепончатокрылых . [ 58 ]
Помимо этих форм существует гермафродитизм, при котором и яйцеклетки , и сперматозоиды производятся одной и той же особью, но не является разновидностью партеногенеза. Это наблюдается у трех видов Icerya . щитовок [ 58 ]
паразитические бактерии, такие как Wolbachia, Было отмечено, что вызывают аутомиктическую телитокию у многих видов насекомых с гаплодиплоидной системой. Они также вызывают удвоение гамет в неоплодотворенных яйцеклетках, в результате чего из них развивается потомство женского пола. [ 58 ]

У видов с гаплодиплоидной системой определения пола , таких как перепончатокрылые (муравьи, пчелы, осы) и тизаноптераны (трипсы), гаплоидные самцы образуются из неоплодотворенных яиц. Обычно яйца откладывает только королева, но не спаренные рабочие могут также откладывать гаплоидные мужские яйца либо регулярно (например, пчелы без жала), либо при особых обстоятельствах. Пример нежизнеспособного партеногенеза распространен среди домашних медоносных пчел . Пчелиная матка — единственная плодовитая самка в улье; если она умирает, не имея возможности заменить жизнеспособную королеву, рабочие пчелы нередко откладывают яйца. Это результат отсутствия феромонов матки и феромонов, выделяемых расплодом без колпачка , которые обычно подавляют развитие яичников у рабочих. Рабочие пчелы не способны к спариванию, и из неоплодотворенных яиц рождаются только трутни (самцы), которые могут спариваться только с маткой. Таким образом, за сравнительно короткий период все рабочие пчелы отмирают, а за ними следуют новые трутни, если они не успели спариться до распада семьи. Считается, что такое поведение развилось, чтобы позволить обреченной колонии производить трутней, которые могут спариваться с девственной королевой и таким образом сохранять генетическое потомство колонии.
Некоторые муравьи и пчелы способны партеногенетически производить диплоидное потомство женского пола. К ним относятся подвид медоносных пчел из Южной Африки, Apis mellifera capensis , рабочие которого способны производить диплоидные яйца партеногенетически и заменять матку в случае ее смерти; другие примеры включают некоторые виды мелких пчел-плотников (род Ceratina ). Известно, что многие осы-паразиты являются партеногенетическими, иногда вследствие заражения Wolbachia .
Рабочие в пяти [ 30 ] Известно, что виды муравьев и королевы некоторых муравьев размножаются путем партеногенеза. У Cataglyphis курсора , европейского муравья-муравья , королевы и рабочие могут производить новых маток путем партеногенеза. Рабочие производятся половым путем. [ 30 ]
У электрических муравьев Центральной и Южной Америки , Wasmannia auropunctata , королевы производят больше маток посредством аутомиктического партеногенеза с центральным слиянием. Стерильные рабочие обычно получаются из яиц, оплодотворенных самцами. Однако в некоторых яйцеклетках, оплодотворенных самцами, оплодотворение может привести к удалению женского генетического материала из зиготы. Таким образом, самцы передают только свои гены, чтобы стать плодовитым потомством мужского пола. Это первый признанный пример вида животных, у которого и самки, и самцы могут воспроизводиться клонально, что приводит к полному разделению мужского и женского генофонда. [ 60 ] Как следствие, у самцов будут только отцы, а у королев - только матери, в то время как бесплодные рабочие - единственные, у кого есть оба родителя обоих полов.
Эти муравьи получают преимущества как бесполого, так и полового размножения. [ 30 ] [ 60 ] — дочери, которые могут воспроизводить потомство (матки), имеют все гены матери, тогда как бесплодные рабочие, чья физическая сила и устойчивость к болезням имеют важное значение, производятся половым путем.
Другие примеры партеногенеза насекомых можно найти у тлей, образующих галлы (например, Pemphigus betae ), самки которых размножаются партеногенетически во время фазы образования галлов их жизненного цикла, а также у травяных трипсов . У травяных трипсов рода Aptinothrips , несмотря на очень ограниченное число видов в этом роде, наблюдалось несколько переходов к асексуальности. [ 61 ]
Ракообразные
[ редактировать ]Размножение ракообразных варьируется как между видами, так и внутри видов. У водяной блохи Daphnia pulex происходит чередование полового и партеногенетического размножения. [ 62 ] Среди наиболее известных крупных десятиногих ракообразных некоторые раки размножаются партеногенезом. « Марморкребы » — партеногенетические раки , которые были обнаружены в торговле домашними животными в 1990-х годах. [ 63 ] Потомство генетически идентично родителю, что указывает на то, что оно размножается путем апомиксиса, то есть партеногенеза, при котором яйца не подвергаются мейозу. [ 64 ] Раки колючие ( Orconectes limosus ) могут размножаться как половым путем, так и партеногенезом. [ 65 ] Красный болотный рак Луизианы ( Procambarus clarkii ), который обычно размножается половым путем, также предположительно размножается партеногенезом. [ 66 ] хотя ни одна особь этого вида не выращивалась таким образом в лаборатории. Artemia parthenogenetica — вид или серия популяций партеногенетических артемий . [ 67 ]
Пауки
[ редактировать ]По крайней мере, два вида пауков семейства Oonopidae (пауки-гоблины), Heteroonops spinimanus и Triaeris stenaspis , считаются партеногенетическими, поскольку самцов никогда не собирали. Партеногенетическое размножение было продемонстрировано в лаборатории T. stenaspis . [ 68 ]
Акулы
[ редактировать ]Партеногенез у акул подтвержден как минимум у трех видов: чепчикообразной , [ 47 ] черноперая акула , [ 69 ] и акула-зебра , [ 70 ] и сообщалось в других.
акула Было обнаружено, что капотоголовая , разновидность маленькой акулы-молота , произвела на свет детеныша, родившегося живым в 2001 году в зоопарке Генри Дурли в Небраске, в аквариуме, в котором находились три самки акул-молотов, но не было самцов. Считалось, что щенок был зачат в результате партеногенеза. После анализа ДНК был сделан вывод, что размножение было партеногенетическим, поскольку ДНК щенка-самки соответствовала только одной самке, которая жила в резервуаре, а ДНК самца у щенка не присутствовала. Щенок не был близнецом или клоном своей матери, а содержал только половину ДНК матери (« аутомиктический партеногенез »). Этот тип размножения ранее наблюдался у костистых рыб, но не у хрящевых рыб, таких как акулы. [ 71 ]
В том же году самка атлантической черноперой акулы в Вирджинии размножилась посредством партеногенеза. [ 72 ] 10 октября 2008 года ученые подтвердили второй случай «непорочного зачатия» у акулы. Журнал «Рыбная биология» сообщил об исследовании, в котором ученые заявили, что анализ ДНК доказал, что детеныш, вынесенный самкой атлантической черноперой акулы в Аквариуме и Центре морских наук Вирджинии, не содержит генетического материала самца. [ 69 ]
В 2002 году две белопятнистые бамбуковые акулы родились в Аквариуме Бель-Айл в Детройте . Они вылупились через 15 недель после того, как их заложили в аквариум, в котором находились только две самки акул. [ 73 ]
В 2011 году повторяющийся партеногенез акул в течение нескольких лет был продемонстрирован у содержащейся в неволе зебровой акулы , разновидности ковровой акулы. [ 70 ] [ 74 ] Генотипирование ДНК показало, что отдельные акулы-зебры могут переключаться с полового размножения на партеногенетическое. [ 75 ]
Лучи
[ редактировать ]В 2024 году было зарегистрировано, что самка круглого ската ( Urobatis Halleri ), находившаяся в неволе вместе со всеми самцами в течение восьми лет, была беременна. [ 76 ] В июне 2024 года в аквариуме, где проживал скат, сообщили, что она не беременна, а страдает редким репродуктивным заболеванием. [ 77 ]
Земноводные
[ редактировать ]Крокодилы
[ редактировать ]В июне 2023 года открытие было сделано в зоопарке Коста-Рики , где исследователи выявили первый задокументированный случай самобеременной крокодилы . Эта самка американского крокодила, обитающая в Парке Рептилания, произвела на свет генетически идентичный плод, сходный с ней на 99,9%. Ученые предполагают, что эта уникальная способность могла быть унаследована от эволюционного предка, предполагая, что даже динозавры могли обладать способностью к самовоспроизведению. 18-летний крокодил отложил яйцо в январе 2018 года, полностью сформировавшийся плод не вылупился и родился мертвым. Примечательно, что этого крокодила держали отдельно от других крокодилов на протяжении всей ее жизни с тех пор, как он был приобретен в возрасте двух лет. [ 78 ] [ 79 ]
Чешуйчатый
[ редактировать ]
Большинство рептилий отряда чешуйчатых ( ящерицы и змеи ) размножаются половым путем , но партеногенез наблюдался естественным путем у некоторых видов хлыстохвостов , некоторых гекконов , скальных ящериц , [ 7 ] [ 80 ] [ 81 ] комодские драконы , [ 82 ] и змеи. [ 83 ] Некоторые из них, такие как траурный геккон Lepidodactylus lugubris , индо-тихоокеанский домашний геккон Hemidactylus garnotii , гибридные хлыстохвосты Cnemidophorus , кавказские скальные ящерицы Darevskia и браминная слепая змея , Indotyphlops braminus являются однополыми и облигатно партеногенетическими. Другие рептилии, такие как комодский варан, другие вараны, [ 84 ] и некоторые виды удавов , [ 13 ] [ 31 ] [ 85 ] питоны , [ 29 ] [ 86 ] файлызмеи , [ 87 ] [ 88 ] подвязочные змеи , [ 89 ] и гремучие змеи [ 90 ] [ 91 ] ранее рассматривались как случаи факультативного партеногенеза, но могут быть и случаями случайного партеногенеза. [ 36 ]
В 2012 году американские исследователи впервые сообщили о факультативном партеногенезе у диких позвоночных среди отловленных беременных самок медноголовой и хлопковой гадюки. [ 92 ] Дракон Комодо, который обычно размножается половым путем, также оказался способен к бесполому размножению путем партеногенеза. [ 93 ] Был задокументирован случай размножения варана Комодо путем полового размножения после известного партеногенетического события. [ 94 ] подчеркивая, что эти случаи партеногенеза являются репродуктивными несчастными случаями, а не адаптивным факультативным партеногенезом. [ 36 ]
Некоторые виды рептилий используют хромосомную систему ZW, которая производит либо самцов (ZZ), либо самок (ZW). До 2010 года считалось, что хромосомная система ZW, используемая рептилиями, неспособна производить жизнеспособное потомство WW, но было обнаружено, что самка удава (ZW) произвела жизнеспособное потомство женского пола с хромосомами WW. [ 95 ]
Партеногенез широко изучался у хлыстовоста Нью-Мексико из рода Aspidoscelis , 15 видов которого размножаются исключительно партеногенезом. Эти ящерицы обитают в сухом и порой суровом климате юго-запада США и севера Мексики. Все эти бесполые виды, по-видимому, возникли в результате гибридизации двух или трех половых видов рода, что привело к образованию полиплоидных особей. Механизм, с помощью которого смешение хромосом двух или трех видов может привести к партеногенетическому размножению, неизвестен. Недавно в лаборатории была выведена гибридная партеногенетическая ящерица-хлыстохвост в результате скрещивания бесполого и полового хлыстохвоста. [ 96 ] Поскольку могут происходить множественные события гибридизации, отдельные партеногенетические виды хлыстохвоста могут состоять из нескольких независимых бесполых линий. Внутри линий очень мало генетического разнообразия, но разные линии могут иметь совершенно разные генотипы.
Интересным аспектом размножения этих бесполых ящериц является то, что все еще наблюдается брачное поведение, хотя все популяции состоят из самок. Одна самка играет роль самца у близкородственных видов и садится на самку, которая собирается отложить яйца. Такое поведение обусловлено гормональными циклами самок, которые заставляют их вести себя как самцы вскоре после откладки яиц, когда уровень прогестерона высок, и брать на себя женскую роль в спаривании перед откладкой яиц, когда доминирует эстроген. Ящерицы, которые разыгрывают ритуал ухаживания, имеют большую плодовитость, чем те, которые содержатся в изоляции, из-за увеличения уровня гормонов, сопровождающего подъем. Таким образом, хотя в популяциях и не хватает самцов, для максимального репродуктивного успеха они все равно нуждаются в стимулах сексуального поведения. [ 97 ]
Некоторые партеногены ящериц демонстрируют образец географического партеногенеза, занимая высокогорные районы, где их предковые формы обладают худшей конкурентной способностью. [ 98 ] У кавказских скальных ящериц рода Darevskia , имеющих шесть партеногенетических форм гибридного происхождения. [ 80 ] [ 81 ] [ 99 ] гибридная партеногенетическая форма D. «dahli» имеет более широкую нишу, чем любой из ее двуполых предков, и ее распространение по Центральному Малому Кавказу вызвало сокращение ареалов как ее материнского, так и отцовского вида. [ 100 ]
Птицы
[ редактировать ]Партеногенез у птиц известен главным образом из исследований на домашних индейках и курах , хотя он отмечен и у домашнего голубя . [ 46 ] В большинстве случаев яйцо не развивается нормально или полностью до вылупления. [ 46 ] [ 101 ] Первое описание партеногенетического развития воробьиных было продемонстрировано у содержащихся в неволе зебровых амадин , хотя делящиеся клетки имели неравномерные ядра и яйца не вылуплялись. [ 46 ]
Партеногенез у индеек, по-видимому, является результатом превращения гаплоидных клеток в диплоидные ; [ 101 ] большинство эмбрионов, полученных таким способом, погибают на ранних стадиях развития. В результате этого процесса редко появляются жизнеспособные птицы, и скорость, с которой это происходит у индеек, может быть увеличена путем селекционного разведения . [ 102 ] однако индейки-самцы, полученные в результате партеногенеза, имеют меньшие семенники и пониженную плодовитость. [ 103 ]
В 2021 году зоопарк Сан-Диего сообщил, что у них было два неоплодотворенных яйца от вылупления калифорнийской программы разведения кондоров . Это первый известный пример партеногенеза у этого вида, а также один из единственных известных примеров партеногенеза, происходящего там, где все еще присутствуют самцы. [ 11 ]
Млекопитающие
[ редактировать ]Случаев естественного партеногенеза млекопитающих в дикой природе не известно. Партеногенетическое потомство млекопитающих будет иметь две Х-хромосомы и, следовательно, будет генетически женским.
В 1936 году Грегори Гудвин Пинкус сообщил об успешной индукции партеногенеза у кроликов . [ 104 ]
В апреле 2004 года ученые Токийского сельскохозяйственного университета успешно использовали партеногенез для создания мыши, лишенной отца . Используя генное нацеливание, они смогли манипулировать двумя импринтированными локусами H19/IGF2 и DLK1/ MEG3 для получения мышей с двойной матерью с высокой частотой. [ 105 ] и впоследствии показали, что мыши, оставшиеся без отца, имеют увеличенную продолжительность жизни . [ 106 ]
Индуцированный партеногенез у мышей и обезьян часто приводит к аномальному развитию. Это связано с тем, что у млекопитающих есть импринтированные генетические области, в которых либо материнская, либо отцовская хромосома инактивируется у потомства, чтобы развитие протекало нормально. Млекопитающие, созданные в результате партеногенеза, будут иметь двойные дозы генов, импринтированных по материнской линии, и не иметь генов, импринтированных по отцовской линии, что приведет к аномалиям развития. Было предложено [ 107 ] что дефекты складчатости или переплетения плаценты являются одной из причин абортивного развития партенота свиней . Как следствие, исследования партеногенеза человека сосредоточены на производстве эмбриональных стволовых клеток для использования в медицинских целях, а не в качестве репродуктивной стратегии. В 2022 году исследователи сообщили, что им удалось добиться партеногенеза у мышей для получения жизнеспособного потомства, рожденного из неоплодотворенных яиц, решив проблемы геномного импринтинга путем «целевого переписывания метилирования ДНК семи контрольных областей импринтинга». [ 14 ]
Методы
[ редактировать ]Эта статья может быть слишком технической для понимания большинства читателей . ( декабрь 2021 г. ) |
Использование электрического или химического раздражителя может вызвать начало процесса партеногенеза при бесполом развитии жизнеспособного потомства. [ 108 ]

Во время развития ооцитов высокая активность фактора, способствующего метафазе (MPF), приводит к остановке ооцитов млекопитающих на стадии метафазы II до момента оплодотворения спермой. Событие оплодотворения вызывает внутриклеточные колебания кальция и целенаправленную деградацию циклина B, регуляторной субъединицы MPF, что позволяет ооциту, задержанному по MII, пройти мейоз.
Чтобы инициировать партеногенез ооцитов свиней, существуют различные методы, вызывающие искусственную активацию, имитирующую проникновение сперматозоидов, такие как обработка ионофором кальция, микроинъекция ионов кальция или электрическая стимуляция. Лечение циклогексимидом, неспецифическим ингибитором синтеза белка, усиливает развитие партенотов у свиней, предположительно, за счет постоянного ингибирования MPF/циклина B. [ 109 ] По мере развития мейоза экструзия второго поляра блокируется воздействием цитохалазина B. Такая обработка приводит к образованию диплоидного (2 материнских генома) партенота. [ 107 ] Партеноты могут быть хирургическим путем перенесены в яйцевод реципиента для дальнейшего развития, но они погибнут из-за задержки развития примерно через 30 дней беременности. Плацента свиного партенота часто выглядит гиповаскулярной: см. бесплатное изображение (рис. 1) в связанной ссылке. [ 107 ]
Люди
[ редактировать ]Сообщения о партеногенезе человека известны с древних времен и занимают видное место в христианстве и различных других религиях . Совсем недавно Хелен Спервей , генетик, специализирующийся на репродуктивной биологии гуппи ( Lebistes reticulatus ), в 1955 году заявила, что партеногенез, который происходит у гуппи в природе, может также происходить (хотя и очень редко) у человеческого вида, приводя к так называемые «непорочные роды». Это произвело фурор среди ее коллег и широкой общественности. [ 110 ] Иногда эмбрион может начать делиться без оплодотворения, но полностью развиваться самостоятельно он не может; поэтому, хотя он может создавать некоторые клетки кожи и нервные клетки, он не может создавать другие (например, скелетные мышцы) и становится разновидностью доброкачественной опухоли, называемой тератомой яичника . [ 111 ] Спонтанная активация яичников не является редкостью и известна с 19 века. Некоторые тератомы могут даже превратиться в примитивные плоды (фетиформные тератомы) с несовершенными головами, конечностями и другими структурами, но нежизнеспособны.
В 1995 г. был зарегистрирован случай частичного партеногенеза человека; Было обнаружено, что у мальчика в некоторых клетках (например, лейкоцитах ) отсутствует какой-либо генетический контент его отца. Ученые полагают, что неоплодотворенная яйцеклетка начала самоделиться, но затем некоторые (но не все) ее клетки были оплодотворены сперматозоидом; это должно было произойти на ранних стадиях развития, поскольку самоактивирующиеся яйца быстро теряют способность к оплодотворению. Неоплодотворенные клетки в конечном итоге дублировали свою ДНК, увеличивая количество хромосом до 46. Когда неоплодотворенные клетки столкнулись с блокировкой развития, оплодотворенные клетки взяли верх и развили эту ткань. У мальчика были асимметричные черты лица и трудности с обучением, но в остальном он был здоров. Это сделало бы его партеногенетической химерой (ребенком с двумя клеточными линиями в теле). [ 112 ] Хотя с тех пор было зарегистрировано более дюжины подобных случаев (обычно обнаруживаемых после того, как у пациентки были выявлены клинические отклонения), не было научно подтвержденных сообщений о нехимерном, клинически здоровом партеноте человека (т.е. полученном из одного партеногенетически активированного ооцита). ). [ 111 ]
26 июня 2007 года Международная корпорация стволовых клеток (ISCC), калифорнийская компания по исследованию стволовых клеток, объявила, что ее ведущий ученый, доктор Елена Ревазова, и ее исследовательская группа были первыми, кто намеренно создал человеческие стволовые клетки из неоплодотворенных человеческих клеток. яйца с помощью партеногенеза. Этот процесс может предложить способ создания стволовых клеток, генетически соответствующих конкретной женщине, для лечения дегенеративных заболеваний, которые могут у нее возникнуть. В декабре 2007 года д-р Ревазова и ISCC опубликовали статью. [ 113 ] иллюстрирующий прорыв в использовании партеногенеза для производства стволовых клеток человека, гомозиготных по HLA- области ДНК. Эти стволовые клетки называются HLA-гомозиготными партеногенетическими стволовыми клетками человека (hpSC-Hhom) и обладают уникальными характеристиками, которые позволят имплантировать производные этих клеток миллионам людей без иммунного отторжения. [ 114 ] При правильном отборе доноров ооцитов по гаплотипу HLA можно создать банк клеточных линий, тканевые производные которых в совокупности могут быть сопоставлены по MHC со значительным количеством индивидуумов в человеческой популяции.
2 августа 2007 года после независимого расследования выяснилось, что дискредитированный южнокорейский учёный Хван У Сук по незнанию произвел первые человеческие эмбрионы, возникшие в результате партеногенеза. Первоначально Хван утверждал, что он и его команда извлекли стволовые клетки из клонированных человеческих эмбрионов, но позже выяснилось, что этот результат был сфабрикован. Дальнейшее исследование хромосом этих клеток показывает показатели партеногенеза в этих извлеченных стволовых клетках, аналогичные тем, которые были обнаружены у мышей, созданных токийскими учеными в 2004 году. Хотя Хван обманул мир, утверждая, что он первым создал искусственно клонированные человеческие эмбрионы, он внес свой вклад. крупный прорыв в исследованиях стволовых клеток путем создания человеческих эмбрионов с использованием партеногенеза. [ 115 ] Истина была открыта в 2007 году, спустя много времени после того, как эмбрионы были созданы им и его командой в феврале 2004 года. Это сделало Хвана первым, сам того не зная, успешно осуществившим процесс партеногенеза для создания человеческого эмбриона и, в конечном итоге, человеческого партеногенетического стебля. клеточная линия. [ нужна ссылка ]
Подобные явления
[ редактировать ]Гиногенез
[ редактировать ]Одной из форм бесполого размножения, связанной с партеногенезом, является гиногенез. что и при партеногенезе, но с требованием, чтобы яйцеклетка просто стимулировалась присутствием сперматозоида для Здесь потомство производится по тому же механизму , развития. Однако сперматозоид не передает никакого генетического материала потомству. Поскольку все гиногенетические виды являются самками, активация их яиц требует спаривания с самцами близкородственного вида для получения необходимого стимула. Некоторые саламандры рода Ambystoma являются гиногенетическими и, по-видимому, оставались таковыми уже более миллиона лет. Считается, что [ кем? ] что успех этих саламандр может быть обусловлен редким оплодотворением яиц самцами, вносящим новый материал в генофонд, что может произойти, возможно, в результате только одного спаривания из миллиона. Кроме того, известно, что амазонская молли размножается гиногенезом. [ 116 ]
Гибридогенез
[ редактировать ]Гибридогенез — способ размножения гибридов . Гибридогенные гибриды (например, геном АВ ), обычно самки, в ходе гаметогенеза исключают один из родительских геномов (А) и образуют гаметы с некомбинированными [ 117 ] геном второго родительского вида (B), вместо того, чтобы содержать смешанные рекомбинированные родительские геномы. Первый геном (А) восстанавливается путем оплодотворения этих гамет гаметами первого вида (АА, половой хозяин, [ 117 ] обычно мужчина). [ 117 ] [ 118 ] [ 119 ]
Таким образом, гибридогенез не является полностью бесполым, а гемиклональным: половина генома передается следующему поколению клонально , в некомбинированном, неповрежденном виде (Б), другая половина - половым путем , рекомбинированная (А). [ 117 ] [ 120 ]
Этот процесс продолжается, так что каждое поколение является наполовину (или геми) клональным со стороны матери и имеет половину нового генетического материала со стороны отца.
Такая форма размножения наблюдается у некоторых живородящих рыб рода Poeciliopsis. [ 118 ] [ 121 ] а также у некоторых представителей Pelophylax spp. («зеленые лягушки» или «водяные лягушки»):
- П. кл. esculentus (съедобная лягушка): P. Lessae × P.ridibundus , [ 117 ] [ 122 ] [ 123 ]
- П. кл. grafi (гибридная лягушка Графа): P. perezi × P.ridibundus [ 117 ]
- П. кл. hispanicus (итальянская съедобная лягушка) - неизвестного происхождения: P. bergeri × P.ridibundus или P. kl. эскулентус [ 117 ]
и, возможно, у P. demarchii .
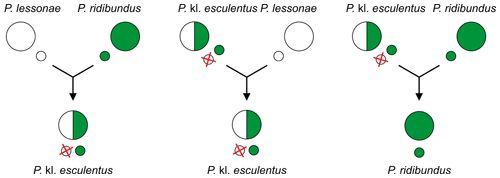
Другие примеры, когда гибридогенез является по меньшей мере одним из способов размножения, включают, например,
- Иберийский гольян Tropidophoxinellus alburnoides ( Squaliuspyrenaicus × гипотетический предок, родственный Anecypris hispanica ) [ 125 ]
- щиповки Cobitis hankugensis × C. longicorpus [ 126 ]
- Палочники B. rossius × Bacillus grandii benazzii [ 127 ]
Партеногенез в мифологии
[ редактировать ]Примеры партеногенеза в форме размножения от одного человека (обычно бога) распространены в мифологии и фольклоре всего мира, в том числе в древнегреческом мифе ; например, Афина родилась из головы Зевса . [ 128 ]
См. также
[ редактировать ]- Андрогенез - форма квазиполового размножения, при которой мужчина является единственным источником ядерного генетического материала эмбриона.
- Телескопические поколения
- Шарль Бонне - женевский ботаник (1720–1793) - провел эксперименты, которые установили то, что сейчас называется партеногенезом у тлей.
- Ян Дзержон - польский пчеловод (1811–1906). - польский пчеловод и пионер партеногенеза среди пчел.
- Жак Леб - американский физиолог и биолог немецкого происхождения - заставил яйца морских ежей начать эмбриональное развитие без спермы.
- Чудесные рождения - Зачатия и рождения при чудесных обстоятельствах.
- Партенокарпия - Получение бессемянных плодов без внесения удобрений - растения с бессемянными плодами.
Ссылки
[ редактировать ]- ^ «партеногенез» . Словарь Merriam-Webster.com . Мерриам-Вебстер.
- ^ «партеногенез» . Оксфордский словарь OxfordDictionaries.com . Английское определение. Архивировано из оригинала 12 сентября 2012 года . Проверено 20 января 2016 г.
- ^ Лидделл; Скотт; Джонс, ред. (1940). « γένεσις А.II» . Греко-английский лексикон . Оксфорд, Великобритания: Clarendon Press – через Perseus / Tufts U. , Медфорд и Сомервилл, Массачусетс. кв .
- ^ Jump up to: а б Хееш, Свенья ; Серрано-Серрано, Марта; Баррера-Редондо, Жозуэ; Лютерингер, Реми; Питерс, Акира Ф.; Дестомб, Кристоф; и др. (июль 2021 г.). «Эволюция жизненных циклов и репродуктивных качеств: данные о бурых водорослях» . Журнал эволюционной биологии . 34 (7): 992–1009. дои : 10.1111/jeb.13880 . ISSN 1010-061X . ПМИД 34096650 . S2CID 92334399 .
- ^
Престон, Элизабет (13 февраля 2024 г.). «Любовь к себе важна, но мы, млекопитающие, застряли в сексе» . Нью-Йорк Таймс . Архивировано из оригинала 13 февраля 2024 года . Проверено 16 февраля 2024 г.
Некоторые самки птиц, рептилий и других животных могут выносить потомство самостоятельно. Но для таких млекопитающих, как мы, яйцеклетки и сперматозоиды нуждаются друг в друге.
- ^ «Самки акул могут размножаться в одиночку, обнаружили исследователи» , The Washington Post , 23 мая 2007 г.; п. А02
- ^ Jump up to: а б Холлидей, Тим Р. (1986). Крейг Адлер (ред.). Рептилии и амфибии . Книги Торстара. п. 101. ИСБН 978-0-920269-81-7 .
- ^ Уокер, Брайан (11 ноября 2010 г.). «Ученые обнаружили неизвестные виды ящериц за обеденным столом» . CNN . Проверено 11 ноября 2010 г.
- ^ Аллен, Л.; Сандерс, КЛ; Томсон, Вирджиния (февраль 2018 г.). «Молекулярные доказательства первых записей факультативного партеногенеза у элапидных змей» . Королевское общество открытой науки . 5 (2): 171901. Бибкод : 2018RSOS....571901A . дои : 10.1098/rsos.171901 . ISSN 2054-5703 . ПМЦ 5830781 . ПМИД 29515892 .
- ^ Сэвидж, Томас Ф. (11 февраля 2008 г.) [12 сентября 2005 г.]. Руководство по распознаванию партеногенеза в инкубированных индюшачьих яйцах (Отчет). Департамент зоотехники. Университет штата Орегон . Архивировано из оригинала 16 июля 2012 года . Проверено 19 марта 2024 г.
- ^ Jump up to: а б с Райдер, Оливер А.; Томас, Стивен; Джадсон, Джессика Мартин; Романов Михаил Н.; Дандекар, Сугандха; Папп, Жанетт С.; и др. (17 декабря 2021 г.). «Факультативный партеногенез у калифорнийских кондоров» . Журнал наследственности . 112 (7): 569–574. doi : 10.1093/jhered/esab052 . ISSN 1465-7333 . ПМЦ 8683835 . ПМИД 34718632 .
- ^ Рамачандран, Р.; Насименту душ Сантуш, М.; Паркер, HM; Макдэниел, компакт-диск (сентябрь 2018 г.). «Родительский половой эффект партеногенеза на производство потомства и продуктивность китайского расписного перепела ( Coturnix chinensis )» . Териогенология . 118 : 96–102. doi : 10.1016/j.theriogenology.2018.05.027 . ПМИД 29886358 . S2CID 47008147 .
- ^ Jump up to: а б Бут, В.; Джонсон, Д.Х.; Мур, С.; Шаль, К.; Варго, Э.Л. (2010). «Доказательства существования жизнеспособных, неклональных, но лишенных отца удавов» . Письма по биологии . 7 (2): 253–256. дои : 10.1098/rsbl.2010.0793 . ПМК 3061174 . ПМИД 21047849 .
- ^ Jump up to: а б Вэй Ю, Ян ЧР, Чжао ЗА (7 марта 2022 г.). «Жизнеспособное потомство, полученное из одиночных неоплодотворенных ооцитов млекопитающих» . ПНАС . 119 (12): e2115248119. Бибкод : 2022PNAS..11915248W . дои : 10.1073/pnas.2115248119 . ПМЦ 8944925 . PMID 35254875 .
- ^ Скотт, Томас (1996). Краткая энциклопедия биологии . Вальтер де Грюйтер. ISBN 978-3-11-010661-9 .
- ^ Пойнар, Джордж О-младший; Тревор А. Джексон; Найджел Л. Белл; Мохд Б-Асри Вахид (июль 2002 г.). « Elaeolenchus parthenonema ng, n. sp. (Nematoda: Sphaerularioidea: Anandranematidae n. fam.), паразитирующий на долгоносике-опылителе пальм Elaeidobius kamerunicus Faust, с филогенетическим обзором Sphaerularioidea Lubbock, Систематическая паразитология . 52 (3): 219–225. дои : 10.1023/А:1015741820235 . ISSN 0165-5752 . ПМИД 12075153 . S2CID 6405965 .
- ^ Уайт, Майкл Джей Ди (1984). «Хромосомные механизмы размножения животных». Боллетино ди Зоологии . 51 (1–2): 1–23. дои : 10.1080/11250008409439455 . ISSN 0373-4137 .
- ^ Пужад-Виллард, июль; Беллидо, Д.; Сегу, Г.; Мелика, Джордж (2001). «Современное состояние знаний о гетерогонии Cynipidae (Hymenoptera, Cynipoidea)». Совместная сессия ICHNSCL по энтомологии . 11 (1999): 87–107.
- ^ Jump up to: а б Краточвил, Лукаш; Вукич, Ясна; Червенка, Ян; Кубичка, Лукаш; Джонсон Покорна, Мартина; Кукачкова, Доминика; и др. (ноябрь 2020 г.). «Смешанное потомство, полученное в результате загадочного партеногенеза у ящерицы» . Молекулярная экология . 29 (21): 4118–4127. Бибкод : 2020MolEc..29.4118K . дои : 10.1111/mec.15617 . ISSN 0962-1083 . ПМИД 32881125 . S2CID 221474843 .
- ^ Райдер, Оливер А; Томас, Стивен; Джадсон, Джессика Мартин; Романов Михаил Н.; Дандекар, Сугандха; Папп, Жанетт С.; и др. (17 декабря 2021 г.). Мерфи, Уильям Дж. (ред.). «Факультативный партеногенез у калифорнийских кондоров» . Журнал наследственности . 112 (7): 569–574. doi : 10.1093/jhered/esab052 . ISSN 0022-1503 . ПМЦ 8683835 . ПМИД 34718632 .
- ^ Бернштейн, Х; Хопф, ФА; Мишод, RE (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–370. дои : 10.1016/s0065-2660(08)60012-7 . ISBN 978-0-12-017624-3 . ПМИД 3324702 .
- ^ Гаврилов И.А.; Кузнецова, В.Г. (2007). «О некоторых терминах, используемых в цитогенетике и репродуктивной биологии щитовок (Homoptera: Coccinea)» (PDF) . Сравнительная цитогенетика . 1 (2): 169–174. ISSN 1993-078X .
- ^ Энгельштедтер, Ян (2017). «Бесексуальный, но не клональный: эволюционные процессы в аутомиктических популяциях | Генетика» . Генетика . 206 (2): 993–1009. doi : 10.1534/genetics.116.196873 . ПМК 5499200 . ПМИД 28381586 .
- ^ Jump up to: а б Моги, Майкл (1986). «Автомиксис: его распространение и статус». Биологический журнал Линнеевского общества . 28 (3): 321–329. дои : 10.1111/j.1095-8312.1986.tb01761.x .
- ^ Захаров И.А. (апрель 2005 г.). «Интратретрадное спаривание и его генетические и эволюционные последствия». Российский генетический журнал . 41 (4): 402–411. дои : 10.1007/s11177-005-0103-z . ISSN 1022-7954 . ПМИД 15909911 . S2CID 21542999 .
- ^ Jump up to: а б с Косин, Дарио Х. Диас, Марта Ново и Роза Фернандес. «Размножение дождевых червей: половой отбор и партеногенез». В «Биологии дождевых червей» под редакцией Айтена Караджи, 24:69–86. Берлин, Гейдельберг: Springer Berlin Heidelberg, 2011. https://doi.org/10.1007%2F978-3-642-14636-7_5 .
- ^ Jump up to: а б Куэльяр, Орландо (1 февраля 1971 г.). «Размножение и механизм мейотической реституции у партеногенетической ящерицы Cnemidophorus uniparens». Журнал морфологии . 133 (2): 139–165. дои : 10.1002/jmor.1051330203 . ISSN 1097-4687 . ПМИД 5542237 . S2CID 19729047 .
- ^ Jump up to: а б с Локки, Юхани; Эско Суомалайнен; Ансси Саура; Пекка Ланкинен (1 марта 1975 г.). «Генетический полиморфизм и эволюция партеногенетических животных. Ii. Диплоидные и полиплоидные Solenobia Triquetrella (lepidoptera: Psychidae)» . Генетика . 79 (3): 513–525. дои : 10.1093/генетика/79.3.513 . ПМЦ 1213290 . ПМИД 1126629 . Проверено 20 декабря 2011 г.
- ^ Jump up to: а б с д Грут, ТВМ; Э Брюинз; Джей Джей Бреувер (28 февраля 2003 г.). «Молекулярно-генетические доказательства партеногенеза у бирманского питона, Python molars bivittatus ». Наследственность . 90 (2): 130–135. CiteSeerX 10.1.1.578.4368 . дои : 10.1038/sj.hdy.6800210 . ISSN 0018-067X . ПМИД 12634818 . S2CID 2972822 .
- ^ Jump up to: а б с д Пирси, М.; Арон, С; Думс, К; Келлер, Л. (2004). «Условное использование пола и партеногенеза для производства рабочих и маток у муравьев» . Наука . 306 (5702): 1780–1783. Бибкод : 2004Sci...306.1780P . дои : 10.1126/science.1105453 . ПМИД 15576621 . S2CID 37558595 .
- ^ Jump up to: а б с д и Бут, Уоррен; Ларри Миллион; Р. Грэм Рейнольдс; Гордон М. Бургхардт; Эдвард Л. Варго; Коби Шал ; Афанасия К. Цика; Гордон В. Шуэтт (декабрь 2011 г.). «Последовательные девственные роды в Новом Свете Боид Змея, колумбийский радужный удав, Epicrates maurus » . Журнал наследственности . 102 (6): 759–763. CiteSeerX 10.1.1.414.384 . doi : 10.1093/jhered/esr080 . ПМИД 21868391 .
- ^ Хейлз, Дина Ф.; Уилсон, Алекс CC; Слоан, Мэтью А.; Симон, Жан-Кристоф; Юридический, Жан-Франсуа; Саннакс, Пол (2002). «Отсутствие обнаруживаемой генетической рекомбинации на Х-хромосоме во время партеногенетического образования самок и самцов тли» . Генетические исследования . 79 (3): 203–209. дои : 10.1017/S0016672302005657 . ПМИД 12220127 .
- ^ Jump up to: а б с д Белл, Г. (1982). Шедевр природы: эволюция и генетика сексуальности , University of California Press, Беркли, стр. 1–635 (см. стр. 295). ISBN 978-0-520-04583-5
- ^ Фанк, Дэвид Х.; Суини, Бернард В.; Джексон, Джон К. (2010). «Почему речные подёнки могут размножаться без самцов, но остаются бисексуальными: случай утраты генетической изменчивости». Журнал Североамериканского бентологического общества . 29 (4): 1258–1266. дои : 10.1899/10-015.1 . S2CID 86088826 .
- ^ Шварц, Герман (1932). «Хромосомный цикл Tetraneura ulmi de Geer» . Журнал клеточных исследований и микроскопической анатомии . 15 (4): 645–687. дои : 10.1007/BF00585855 . S2CID 43030757 .
- ^ Jump up to: а б с д и ван дер Коой, CJ; Швандер, Т. (2015). «Партеногенез: рождение новой линии или репродуктивный несчастный случай?» (PDF) . Современная биология . 25 (15): Р659–Р661. Бибкод : 2015CBio...25.R659V . дои : 10.1016/j.cub.2015.06.055 . ПМИД 26241141 .
- ^ Ламперт, КП (2008). «Факультативный партеногенез у позвоночных: репродуктивная ошибка или случайность?». Сексуальное развитие . 2 (6): 290–301. дои : 10.1159/000195678 . ПМИД 19276631 . S2CID 9137566 .
- ^ Суомалайнен, Э.; и др. (1987). Цитология и эволюция в партеногенезе . Бока-Ратон, Флорида: CRC Press.
- ^ Jump up to: а б Штельцер, К.-П.; Шмидт, Дж.; Видлройтер, А.; Рисс, С. (2010). «Потеря полового размножения и карликовость мелких многоклеточных животных» . ПЛОС . 5 (9): e12854. Бибкод : 2010PLoSO...512854S . дои : 10.1371/journal.pone.0012854 . ПМЦ 2942836 . ПМИД 20862222 .
- ^ Шойерль, Томас; и др. (2011). «Фенотипический аллель, вызывающий облигатный партеногенез» . Журнал наследственности . 102 (4): 409–415. дои : 10.1093/jhered/esr036 . ПМК 3113615 . ПМИД 21576287 . сеть. 23 октября 2012 г.
- ^ Бут, В.; Смит, CF; Эскридж, штат Пенсильвания; Хосс, СК; Мендельсон-младший; Шуэтт, GW (2012). «Факультативный партеногенез обнаружен у диких позвоночных» . Письма по биологии . 8 (6): 983–985. дои : 10.1098/rsbl.2012.0666 . ПМЦ 3497136 . ПМИД 22977071 .
- ^ Jump up to: а б Ворбургер, Кристоф (2003). «Экологически связанные модели репродуктивных способов тли Myzus persicae и преобладание двух «суперклонов» в Виктории, Австралия». Молекулярная экология . 12 (12): 3493–3504. Бибкод : 2003MolEc..12.3493V . дои : 10.1046/j.1365-294X.2003.01998.x . ПМИД 14629364 . S2CID 32192796 .
- ^ Jump up to: а б Кэрон, В.; Норгейт, М.; Эде, Ф.Дж.; Найман, Т. (2013). «Новые микросателлитные ДНК-маркеры указывают на строгий партеногенез и небольшое количество генотипов у инвазивного ивового пилильщика Nematus oligospilus » (PDF) . Бюллетень энтомологических исследований . 103 (1): 74–88. дои : 10.1017/S0007485312000429 . ПМИД 22929915 . S2CID 25210471 .
- ^ Смит, Джон Мейнард (1978). Эволюция секса . Архив Кубка. п. 42. ИСБН 978-0-521-21887-0 .
- ^ Прайс, АХ (1992). «Сравнительное поведение ящериц рода Cnemidophorus (Teiidae) с комментариями по эволюции партеногенеза у рептилий». Копейя . 1992 (2): 323–331. дои : 10.2307/1446193 . JSTOR 1446193 .
- ^ Jump up to: а б с д Шут, Э.; Хеммингс, Н.; Биркхед, Т.Р. (2008). «Партеногенез у воробьиных птиц зебры Taeniopygia Guttata ». Ибис . 150 (1): 197–199. дои : 10.1111/j.1474-919x.2007.00755.x .
- ^ Jump up to: а б Чепмен, Демиан Д.; Шивджи, Махмуд С.; Луи, Эд; Соммер, Джули; Флетчер, Хью; Продёль, Пауло А. (2007). «Непорочное зачатие у акулы-молота» . Письма по биологии . 3 (4): 425–427. дои : 10.1098/rsbl.2007.0189 . ПМК 2390672 . ПМИД 17519185 .
- ^ Jump up to: а б Вриенгук, Р.К., Р.М. Доули, С.Дж. Коул и Дж.П. Богарт. 1989. «Список известных однополых позвоночных», стр. 19–23 в : «Эволюция и экология однополых позвоночных» . Р. М. Доули и Дж. П. Богарт (ред.), Бюллетень 466, Музей штата Нью-Йорк, Олбани.
- ^ Хаббс, CL; Хаббс, LC (1932). «Видимый партеногенез в природе у рыб гибридного происхождения». Наука . 76 (1983): 628–630. Бибкод : 1932Sci....76..628H . дои : 10.1126/science.76.1983.628 . ПМИД 17730035 .
- ^ Уртадо-Гонсалес, ОП; Ламур, К.Х. (2009). «Доказательства инбридинга и апомиксиса при близких скрещиваниях Phytophthora capsici ». Патология растений . 58 (4): 715–722. дои : 10.1111/j.1365-3059.2009.02059.x .
- ^ Читайте, В.М. Дж. (июль 1988 г.). «Онихофора Тринидада, Тобаго и Малых Антильских островов». Зоологический журнал Линнеевского общества . 93 (3): 225–257. дои : 10.1111/j.1096-3642.1988.tb01362.x .
- ^ «Бделлоиды: секса нет более 40 миллионов лет» . Бесплатная библиотека. Новости науки . Проверено 30 апреля 2011 г.
- ^ Штельцер, К.-П.; Шмидт, Дж.; Видлройтер, А.; Рисс, С. (2010). «Потеря полового размножения и карликовость у мелких многоклеточных животных» . ПЛОС ОДИН . 5 (9): e12854. Бибкод : 2010PLoSO...512854S . дои : 10.1371/journal.pone.0012854 . ПМЦ 2942836 . ПМИД 20862222 .
- ^ Лентати, Дж. Бенацци (1966). «Амфимиксис и псевдогамия у пресноводных триклад: экспериментальное восстановление полиплоидных псевдогамных биотипов». Хромосома . 20 : 1–14. дои : 10.1007/BF00331894 . S2CID 21654518 .
- ^ Уоллес, К. (1992). «Артеногенез, пол и хромосомы у Potamopyrgus ». Журнал исследований моллюсков . 58 (2): 93–107. дои : 10.1093/моллюс/58.2.93 .
- ^ Бен-Ами, Ф.; Хеллер, Дж. (2005). «Пространственные и временные закономерности партеногенеза и паразитизма у пресноводной улитки Melanoides Tuberculata ». Журнал эволюционной биологии . 18 (1): 138–146. дои : 10.1111/j.1420-9101.2004.00791.x . ПМИД 15669970 . S2CID 10422561 .
- ^ Миранда, Нельсон А.Ф.; Периссинотто, Ренцо; Эпплтон, Кристофер С.; Лалуэса-Фокс, Карлес (2011). «Структура населения инвазивного партеногенетического брюхоногого моллюска в прибрежных озерах и эстуариях северного Квазулу-Натала, Южная Африка» . ПЛОС ОДИН . 6 (8): e24337. Бибкод : 2011PLoSO...624337M . дои : 10.1371/journal.pone.0024337 . ПМК 3164166 . ПМИД 21904629 .
- ^ Jump up to: а б с д и Киркендалл Л.Р. и Нормарк Б. (2003) «Партеногенез» в Энциклопедии насекомых (Винсент Х. Реш и Р.Т. Карде, ред.) Academic Press. стр. 851–856
- ^ Коупленд, Клаудия С.; Хой, Марджори А.; Джеяпракаш, Айямперумал; Алуя, Мартин; Рамирес-Ромеро, Рикардо; Сивински, Джон М. (1 сентября 2010 г.). «Генетические характеристики двуполых и женских популяций Odontosema anastrephae (Hymenoptera: Figitidae)» . Флоридский энтомолог . 93 (3): 437–443. дои : 10.1653/024.093.0318 .
- ^ Jump up to: а б Фурнье, Дени; Эступ, Арно; Оривел, Жером; Фуко, Жюльен; Журдан, Эрве; Ле Бретон, Жюльен Ле; Келлер, Лоран (2005). «Клональное размножение самцов и самок у маленького огненного муравья» (PDF) . Природа . 435 (7046): 1230–1234. Бибкод : 2005Natur.435.1230F . дои : 10.1038/nature03705 . ПМИД 15988525 . S2CID 1188960 .
- ^ CJ ван дер Коой и Т. Швандер (2014) «Эволюция асексуальности с помощью различных механизмов у травяных трипсов (Thysanoptera: Aptinothrips)» Эволюция 86: 1883–1893
- ^ Идс, Брайан Д.; Колбурн, Джон К; Богуски, Элизабет; Эндрюс, Джастен (2007). «Профилирование экспрессии генов с учетом пола во время партеногенетического размножения у Daphnia pulex» . БМК Геномика . 8 : 464. дои : 10.1186/1471-2164-8-464 . ПМК 2245944 . ПМИД 18088424 .
- ^ Шольц, Герхард; Брабанд, Анке; Толли, Лаура; Рейманн, Эндрю; Миттманн, Беате; Руководитель, Крис; и др. (2003). «Партеногенез у рака-аутсайдера» . Экология. Природа 421 (6925): 806. Бибкод : 2003Nature.421..806S . дои : 10.1038/421806a . ПМИД 12594502 . S2CID 84740187 .
- ^ Мартин, Пер; Кольманн, Клаус; Шольц, Герхард (2007). «Партеногенетические . мраморные раки производят генетически однородное потомство» естественные науки . 94 (10): 843–846. Бибкод : 2007NW.....94..843M . дои : 10.1007/s00114-007-0260-0 . ПМИД 17541537 . S2CID 21568188 .
- ^ Буржич, Милош; Гулак, Мартин; Куба, Антонин; Петрусек, Адам; Козак, Павел; Этгес, Уильям Дж. (31 мая 2011 г.). «Успешный захватчик раков способен к факультативному партеногенезу: новый репродуктивный способ десятиногих ракообразных» . ПЛОС Один . 6 (5): e20281. Бибкод : 2011PLoSO...620281B . дои : 10.1371/journal.pone.0020281 . ПМК 3105005 . ПМИД 21655282 .
- ^ Юэ GH, Ван GL, Чжу BQ, Ван CM, Чжу ZY, Ло LC (2008). «Открытие четырех природных клонов у вида раков Procambarus clarkii » . Международный журнал биологических наук . 4 (5): 279–282. дои : 10.7150/ijbs.4.279 . ПМЦ 2532795 . ПМИД 18781225 .
- ^ Муньос, Хоакин; Гомес, Африка; Грин, Энди Дж.; Фигерола, Хорди; Амат, Франциско; Рико, Чиро; Моро, Корри С. (4 августа 2010 г.). «Эволюционное происхождение и филогеография диплоидного облигатного партеногена Artemia parthenogenetica (Branchiopoda: Anostraca)» . ПЛОС Один . 5 (8): е11932. Бибкод : 2010PLoSO...511932M . дои : 10.1371/journal.pone.0011932 . ПМЦ 2915914 . ПМИД 20694140 .
- ^ Коренко Станислав; Шмерда, Якуб и Пекар, Стано (2009). «История жизни партеногенетического паука-оонопида Triaeris stenaspis (Araneae: Oonopidae)» . Европейский журнал энтомологии . 106 (2): 217–223. дои : 10.14411/eje.2009.028 . Проверено 30 апреля 2016 г.
- ^ Jump up to: а б Чепмен, Д.Д.; Фирчау, Б.; Шивджи, М.С. (2008). «Партеногенез у крупнотелой акулы-реквиема, черноперой». Журнал биологии рыб . 73 (6): 1473–1477. дои : 10.1111/j.1095-8649.2008.02018.x .
- ^ Jump up to: а б Робинсон, ДП; Баверсток, В.; Аль-Джару, А.; Хайленд, К.; Хазанедари, К.А. (2011). «Ежегодно повторяющийся партеногенез у акулы-зебры Stegostoma fasciatum ». Журнал биологии рыб . 79 (5): 1376–1382. Бибкод : 2011JFBio..79.1376R . дои : 10.1111/j.1095-8649.2011.03110.x . ПМИД 22026614 .
- ^ «У акул в плену было «непорочное зачатие» » . Новости Би-би-си . 23 мая 2007 года . Проверено 23 декабря 2008 г.
- ^ « Непорочное зачатие» для аквариумных акул» . Metro.co.uk . 10 октября 2008 года . Проверено 10 октября 2008 г.
- ^ «Акула рожает непорочно в Детройте» . Нэшнл Географик . Сентябрь 2002 г. Архивировано из оригинала 29 сентября 2002 г. Проверено 17 апреля 2010 г.
- ^ «Первое непорочное рождение зебровой акулы в Дубае» . Sharkyear.com . 12 декабря 2011 г.
- ^ Даджен, Кристин Л.; Коултон, Лора; Кость, Рен; Овенден, Дженнифер Р.; Томас, Северин (16 января 2017 г.). «Переход от полового размножения к партеногенетическому у зебровой акулы» . Научные отчеты . 7 : 40537. Бибкод : 2017NatSR...740537D . дои : 10.1038/srep40537 . ISSN 2045-2322 . ПМЦ 5238396 . ПМИД 28091617 .
- ^ Хьюсон, Джорджи (14 февраля 2024 г.). «Скат Шарлотта должна родить через несколько недель, несмотря на отсутствие компании самцов скатов в течение многих лет» . Австралийская радиовещательная корпорация . Проверено 17 февраля 2024 г.
- ^ Чаппелл, Билл (4 июня 2024 г.). «После того, как Шарлотта, одинокий скат, сказала, что беременна, аквариум теперь говорит, что она больна» . NPR.org . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . Проверено 5 июня 2024 г.
- ^ «Крокодил обнаружила, что забеременела» . Новости Би-би-си . 7 июня 2023 г. Проверено 7 июня 2023 г.
- ^ Бут, Уоррен; Левин, Бренна А.; Коруш, Джоэл Б.; Дэвис, Марк А.; Дуайер, Кетцаль; Де Плекер, Роэл; Шуэтт, Гордон В. (июнь 2023 г.). «Открытие факультативного партеногенеза у крокодила нового мира» . Письма по биологии . 19 (6). дои : 10.1098/rsbl.2023.0129 . hdl : 10919/117182 . ISSN 1744-957X . ПМЦ 10244963 . ПМИД 37282490 .
- ^ Jump up to: а б Даревский ИС. 1967. Скальные ящерицы Кавказа: систематика, экология и филогенез полиморфных групп кавказских скальных ящериц подрода Archaeolacerta . Наука: Ленинград.
- ^ Jump up to: а б Тархнишвили, Д.Н. (2012). «Эволюционная история, среда обитания, разнообразие и видообразование кавказских скальных ящериц». В Дженкинсе, ОП (ред.). Достижения в области зоологических исследований . Том. 2. Хауппож, Нью-Йорк: Издательство Nova Science. стр. 79–120.
- ^ Уоттс, ПК; Булей, КР; Сандерсон, С.; Бордман, В.; Чиофи, К.; Гибсон, Р. (2006). «Партеногенез у варанов Комодо». Природа . 444 (7122): 1021–1022. Бибкод : 2006Natur.444.1021W . дои : 10.1038/4441021a . ПМИД 17183308 . S2CID 4311088 .
- ^ «Самооплодотворенная змея в Миссури пережила еще одно «непорочное зачатие» » . ЮПИ.com . Юнайтед Пресс Интернэшнл . 21 сентября 2015 года . Проверено 3 октября 2015 г.
- ^ Вихманн, Р. (2012). «Наблюдения за партеногенезом у варанов» (PDF) . Биавак . 6 (1): 11–21.
- ^ Кинни, Мэн; Вак, РФ; Гран, РА; Лайонс, Л. (2013). «Партеногенез у бразильского радужного удава ( Epicrates cenchria cenchria )» Зоопарковая биология 32 (2): 172–176. дои : 10.1002/zoo.21050 . ПМИД 23086743 .
- ^ Шепард, Кайл (18 декабря 2014 г.). «Рождение непорочной змеи» (Пресс-релиз).
- ^ Магнуссон, МЫ (1979). «Производство эмбриона Acrochordus javanicus, изолированного в течение семи лет». Копейя . 1979 (4): 744–745. дои : 10.2307/1443886 . JSTOR 1443886 .
- ^ Дубач, Дж.; Саевич А.; Поли, Р. (1997). «Партеногенез у арафурской змеи ( Acrochordus arafurae )». Герпетологическое естествознание . 5 (1): 11–18.
- ^ Рейнольдс, Р.Г.; Бут, В.; Шуэтт, GW; Фитцпатрик, Б.М.; Бургхардт, генеральный директор (2012). «Последовательное непорочное рождение жизнеспособного потомства мужского пола у клетчатой подвязки Thamnophis marcianus ». Биологический журнал Линнеевского общества . 107 (3): 566–572. дои : 10.1111/j.1095-8312.2012.01954.x .
- ^ Шуэтт, GW; Фернандес, П.Дж.; Гергитс, ВФ; Касна, Нью-Джерси; Чизар, Д.; Смит, HM; и др. (1997). «Производство потомства в отсутствие самцов: доказательства факультативного партеногенеза у двуполых змей». Герпетологическое естествознание . 5 (1): 1–10.
- ^ Шуэтт, GW; Фернандес, П.Дж.; Чизар, Д.; Смит, Х.М. (1998). «Сыновья без отца: новый тип партеногенеза у змей». Фауна . 1 (3): 20–25.
- ^ «У диких змей обнаружено непорочное зачатие» . Би-би-си Природа . 12 сентября 2012 года. Архивировано из оригинала 13 сентября 2012 года . Проверено 12 сентября 2012 г.
- ^ Хайфилд, Роджер (21 декабря 2006 г.). «Пожалуйста, никакого секса, мы ящерицы» . «Дейли телеграф» . Архивировано из оригинала 11 октября 2007 года.
- ^ «Непорочное зачатие драконов» . Индус . 25 января 2007 г. Архивировано из оригинала 1 октября 2007 г. Проверено 3 февраля 2007 г.
- ^ Уокер, Мэтт (3 ноября 2010 г.). «Змея имеет уникальное «непорочное зачатие» » . Новости Би-би-си .
- ^ Лютес, Арасели А.; Бауманн, Диана П.; Нивз, Уильям Б.; Бауманн, Питер (14 июня 2011 г.). «Лабораторный синтез самостоятельно размножающихся видов позвоночных» . Труды Национальной академии наук США . 108 (24): 9910–9915. Бибкод : 2011PNAS..108.9910L . дои : 10.1073/pnas.1102811108 . ПМК 3116429 . ПМИД 21543715 .
- ^ Крюс, Д.; Грассман, М.; Линдзи, Дж. (1986). «Поведенческое облегчение размножения половых и однополых ящериц-хлыстохвостов» . Труды Национальной академии наук . 83 (24): 9547–9550. Бибкод : 1986PNAS...83.9547C . дои : 10.1073/pnas.83.24.9547 . ПМЦ 387177 . ПМИД 3467325 .
- ^ Вриенгук, Р.К.; Паркер, ЭД (2009). «Географический партеногенез: генотипы общего назначения и вариации замороженных ниш». В красоте я; Мартенс К.; ван Дейк П. (ред.). Потерянный секс . Берлин, Делавэр: Springer Publications. стр. 99–131.
- ^ Мерфи, RW; Даревский И.С.; МакКаллох, РД; Фу, Дж.; Куприянова Л.А.; Аптон, Делавэр; Даниелян, Ф. (1997). «Старость, множественные образования или генетическая пластичность? Клональное разнообразие у партеногенетической кавказской скальной ящерицы Lacerta dahli ». Генетика . 101 (2): 125–130. дои : 10.1023/А:1018392603062 . ПМИД 16220367 . S2CID 11145792 .
- ^ Тархнишвили Д.; Гавашелишвили А.; Авалиани, А.; Мурцхваладзе, М.; Мумладзе, Л. (2010). «Однополая скальная ящерица, возможно, превзойдет своих бисексуальных прародителей на Кавказе» . Биологический журнал Линнеевского общества . 101 (2): 447–460. дои : 10.1111/j.1095-8312.2010.01498.x .
- ^ Jump up to: а б Митвох, У. (1978). «Партеногенез» . Журнал медицинской генетики . 15 (3): 165–181. дои : 10.1136/jmg.15.3.165 . ПМЦ 1013674 . ПМИД 353283 .
- ^ Нестор, Карл (2009). «Партеногенез у индеек» . Потрясающая Турция. oardc.ohio-state.edu/4h . 4Ч / Птица. Университет штата Огайо . Архивировано из оригинала 14 июля 2010 года.
- ^ Сарвелла, П. (1974). «Строение семенников у нормальных и партеногенетических индеек». Журнал наследственности . 65 (5): 287–290. doi : 10.1093/oxfordjournals.jhered.a108530 . ПМИД 4373503 .
- ^ Пинкус, Грегори (2018). Яйца млекопитающих . Нью-Йорк, штат Нью-Йорк: Компания Macmillan – через Интернет-архив .
- ^ Кавахара, Манабу; Ву, Цион; Такахаши, Нозоми; Морита, Шинноске; Ямада, Каори; Ито, Мицутеру; и др. (2007). «Высокочастотное создание жизнеспособных мышей из искусственно созданных двуматеринских эмбрионов». Природная биотехнология . 25 (9): 1045–1050. дои : 10.1038/nbt1331 . ПМИД 17704765 . S2CID 7242745 .
- ^ Кавахара, М.; Коно, Т. (2009). «Долголетие мышей без отца» . Репродукция человека . 25 (2): 457–461. дои : 10.1093/humrep/dep400 . ПМИД 19952375 .
- ^ Jump up to: а б с д и Бишофф, СР; Цай, С.; Хардисон, Н.; Моцингер-Рейф, А.А.; Фрекинг, бакалавр искусств; Ноннеман, Д.; и др. (2009). «Характеристика консервативных и неконсервативных импринтированных генов свиней» . Биология размножения . 81 (5): 906–920. дои : 10.1095/biolreprod.109.078139 . ПМК 2770020 . ПМИД 19571260 .
- ^ Версирен, К; Хайндрикс, Б; Лиерман, С; Геррис, Дж; Де Саттер, П. (2010). «Развитие компетентности партеногенетических эмбрионов мыши и человека после химической или электрической активации» . Репрод Биомед . 21 (6): 769–775. дои : 10.1016/j.rbmo.2010.07.001 . ПМИД 21051286 .
- ^ Jump up to: а б Мори, Хиронори; Мизобе, Ямато; Иноуэ, Шин; Уэнохара, Акари; Такеда, Мицуру; Ёсида, Мицутоши; Миёси, Казучика (2008). «Влияние циклогексимида на партеногенетическое развитие ооцитов свиньи, активированных ультразвуковой обработкой» . Журнал воспроизводства и развития 54 (5): 364–369. дои : 10.1262/jrd.20064 . ПМИД 18635923 .
- ↑ Время , 28 ноября 1955 г.; Редакционная статья в The Lancet , 2: 967 (1955).
- ^ Jump up to: а б де Карли, Габриэль Хосе и Тьяго Кампос Перейра. «О партеногенезе человека». Медицинские гипотезы 106 (2017): 57–60.
- ↑ Филип Коэн, «Мальчик, у которого в крови нет отца» , New Scientist , 7.10.1995
- ^ Ревазова Е.С.; Туровец, Н.А.; Кочеткова, О.Д.; Киндарова, Л.Б.; Кузьмичев Л.Н.; Янус, JD; Прыжкова, М.В. (2007). «Линии стволовых клеток, специфичные для пациента, полученные из партеногенетических бластоцист человека». Клонирование и стволовые клетки . 9 (3): 432–449. дои : 10.1089/clo.2007.0033 . ПМИД 17594198 .
- ^ Ревазова Е.С.; Туровец, Н.А.; Кочеткова, О.Д.; Агапова, Л.С.; Себастьян, JL; Прыжкова М.В.; и др. (2008). «HLA-гомозиготные линии стволовых клеток, полученные из партеногенетических бластоцист человека». Клонирование и стволовые клетки . 10 (1): 11–24. дои : 10.1089/clo.2007.0063 . ПМИД 18092905 .
- ^ Уильямс, Крис. «Мошенник со стволовыми клетками совершил прорыв в области непорочного зачатия: серебряная подкладка для корейского научного скандала» , The Register , 3 августа 2007 г.
- ^ «Никакого секса для видов рыб, состоящих исключительно из девочек» . Новости Би-би-си . 23 апреля 2008 года . Проверено 11 июня 2007 г.
- ^ Jump up to: а б с д и ж г час Холсбек, Г.; Джоорис, Р. (2010). «Потенциальное влияние исключения генома чужеродными видами у гибридогенных водяных лягушек ( комплекс Pelophylax esculentus )». Биологические инвазии . 12 (1): 1–13. Бибкод : 2010BiInv..12....1H . дои : 10.1007/s10530-009-9427-2 . ISSN 1387-3547 . S2CID 23535815 .
- ^ Jump up to: а б Шульц, Р. Джек (ноябрь – декабрь 1969 г.). «Гибридизация, унисексуальность и полиплоидия костистых рыб Poeciliopsis (Poeciliidae) и других позвоночных». Американский натуралист . 103 (934): 605–619. дои : 10.1086/282629 . JSTOR 2459036 . S2CID 84812427 .
- ^ Вриенгук, Роберт К. (1998). «Партеногенез и природные клоны» (PDF) . В Кнобиле, Эрнст; Нил, Джимми Д. (ред.). Энциклопедия репродукции . Том. 3. Академическая пресса. стр. 695–702 . ISBN 978-0-12-227020-8 .
- ^ Саймон, Ж.-К.; Дельмотт, Ф.; Риспе, К.; Криз, Т. (2003). «Филогенетические отношения между партеногенами и их половыми родственниками: возможные пути партеногенеза у животных» (PDF) . Биологический журнал Линнеевского общества . 79 : 151–163. дои : 10.1046/j.1095-8312.2003.00175.x . Проверено 21 июня 2015 г.
- ^ Вриенгук, Дж. М.; Авизе, JC; Вриенгук, Р.К. (1 января 1992 г.). «Древняя клональная линия рода рыб Poeciliopsis (Atheriniformes: Poeciliidae)» . Труды Национальной академии наук США . 89 (1): 348–352. Бибкод : 1992PNAS...89..348Q . дои : 10.1073/pnas.89.1.348 . ISSN 0027-8424 . ПМЦ 48234 . ПМИД 11607248 .
- ^ «Гибридогенез у водяных лягушек» . tolweb.org . Примечание 579. Архивировано из оригинала 14 июля 2014 года . Проверено 13 марта 2007 г.
- ^ Бекебум, LW; Вриенгук, Р.К. (1998). «Эволюционная генетика и экология спермозависимого партеногенеза» . Журнал эволюционной биологии . 11 (6): 755–782. дои : 10.1046/j.1420-9101.1998.11060755.x . ISSN 1420-9101 . S2CID 85833296 .
- ^ Форбургер, Кристоф; Рейер, Хайнц-Ульрих (2003). «Генетический механизм смены видов у европейских водяных лягушек?» (PDF) . Сохраняющая генетика . 4 (2): 141–155. дои : 10.1023/А:1023346824722 . ISSN 1566-0621 . S2CID 20453910 . Проверено 21 июня 2015 г.
- ^ Инасио, А; Пиньо, Дж; Перейра, премьер-министр; Комай, Л; Коэльо, ММ (2012). «Глобальный анализ транскриптома малой РНК в различных плоидиях и геномных комбинациях комплекса позвоночных - Squalius alburnoides » . ПЛОС ОДИН . 7 (7: e41158): 359–368. Бибкод : 2012PLoSO...741158I . дои : 10.1371/journal.pone.0041158 . ПМК 3399795 . ПМИД 22815952 .
- ^ Сайто, К; Ким, И.С.; Ли, Э.Х. (2004). «Интрогрессия митохондриальных генов между щиповками посредством гибридогенеза» . Зоологическая наука . 21 (7): 795–798. дои : 10.2108/zsj.21.795 . ПМИД 15277723 . S2CID 40846660 .
- ^ Мантовани, Барбара; Скали, Валерио (1992). «Гибридогенез и андрогенез у палочников Bacillus rossius - Grandii benazzii (Insecta, Phasmatodea)». Эволюция 46 (3): 783–796. дои : 10.2307/2409646 . JSTOR 2409646 . ПМИД 28568678 .
- ^ Ригольозо, Маргарита (2010). Богини-матери древности . Нью-Йорк: Пэлгрейв Макмиллан. ISBN 978-0-230-61886-2 .
Дальнейшее чтение
[ редактировать ]- Доули, Роберт М. и Богарт, Джеймс П. (1989). Эволюция и экология однополых позвоночных . Олбани: Музей штата Нью-Йорк. ISBN 1-55557-179-4 .
- Фангерау, Х (2005). «Может ли искусственный партеногенез обойти этические ловушки при терапевтическом клонировании человека? Историческая перспектива» . Журнал медицинской этики . 31 (12): 733–735. дои : 10.1136/jme.2004.010199 . ПМК 1734065 . ПМИД 16319240 .
- Футуйма, Дуглас Дж. и Слаткин, Монтгомери. (1983). Коэволюция . Сандерленд, Массачусетс: Sinauer Associates. ISBN 0-87893-228-3 .
- Хор, Т; Рэпкинс, Р.; Грейвс, Дж (2007). «Строительство и эволюция импринтированных локусов у млекопитающих». Тенденции в генетике . 23 (9): 440–448. дои : 10.1016/j.tig.2007.07.003 . ПМИД 17683825 .
- Коно, Т.; Обата, Ю.; Ву, К.; Нива, К.; Оно, Ю.; Ямамото, Ю.; Парк, ES; Со, Ж.-С.; Огава, Х. (2004). «Рождение партеногенетических мышей, способных развиваться до взрослой жизни». Природа . 428 (6985): 860–864. Бибкод : 2004Natur.428..860K . дои : 10.1038/nature02402 . ПМИД 15103378 . S2CID 4353479 .
- Мейнард Смит, Джон. (1978). Эволюция секса . Кембридж: Издательство Кембриджского университета. ISBN 0-521-29302-2 .
- Мишод, Ричард Э. и Левин, Брюс Р. (1988). Эволюция секса . Сандерленд, Массачусетс: Sinauer Associates. ISBN 0-87893-459-6 .
- Шлупп, Инго (2005). «Эволюционная экология гиногенеза». Ежегодный обзор экологии, эволюции и систематики . 36 : 399–417. doi : 10.1146/annurev.ecolsys.36.102003.152629 .
- Саймон, Дж; Риспе, Клод; Саннакс, Пол (2002). «Экология и эволюция пола у тлей». Тенденции в экологии и эволюции . 17 : 34–39. дои : 10.1016/S0169-5347(01)02331-X .
- Стернс, Стефан К. (1988). Эволюция секса и ее последствия (Experientia Supplementum, Vol. 55). Бостон: Биркхаузер. ISBN 0-8176-1807-4 .
Внешние ссылки
[ редактировать ]- Репродуктивное поведение хлыстохвостов в лаборатории Крюса
- Типы бесполого размножения
- Партеногенез в инкубированных яйцах индейки из Университета штата Орегон.
- Новости National Geographic: На Рождество ожидается непорочное зачатие – Комодский дракон
- «Непорочное зачатие» гигантских ящериц (комодский варан)» BBC News
- Рейтер: Гордая мама (и папа) пятерых детей, варан Комодо. Архивировано 9 мая 2021 года в Wayback Machine.
- Самки акул способны к непорочному зачатию
- Ученые подтверждают «непорочное зачатие» акулы. Статья Стива Шкотака AP обновлена в 1:49 по восточноевропейскому времени, пятница, 10 октября 2008 г.