Человеческий лейкоцитарный антиген
| Человеческий лейкоцитарный антиген | |
|---|---|
 Схематическое изображение MHC класса I | |
| Идентификаторы | |
| Символ | HLA |
| ИнтерПро | ИПР037055 |
| Мембраном | 63 |

Система человеческого лейкоцитарного антигена ( HLA ) или комплекс генов на хромосоме 6 человека, которые кодируют белки клеточной поверхности, ответственные за регуляцию иммунной системы . [1] Система HLA также известна как человеческая версия главного комплекса гистосовместимости (MHC), обнаруженного у многих животных. [2]
Мутации в генах HLA могут быть связаны с аутоиммунными заболеваниями , такими как диабет I типа и целиакия . Генный комплекс HLA расположен на участке длиной 3 Мб в хромосоме 6, p-плечо в положении 21,3. Гены HLA высоко полиморфны , а это означает, что они имеют множество различных аллелей , что позволяет им точно настраивать адаптивную иммунную систему . Белки, кодируемые определенными генами, также известны как антигены в результате их исторического открытия в качестве факторов при трансплантации органов . [3]
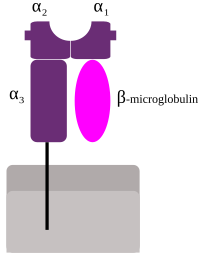
HLA, соответствующие классу MHC I ( A , B и C ), все из которых относятся к группе HLA Class1, представляют пептиды изнутри клетки. Например, если клетка заражена вирусом, система HLA переносит фрагменты вируса на поверхность клетки, чтобы иммунная система могла уничтожить клетку. Эти пептиды производятся из переваренных белков, которые расщепляются в протеасомах . В общем, эти конкретные пептиды представляют собой небольшие полимеры около 8-10 аминокислот . длиной [4] Чужеродные антигены, представленные MHC класса I, привлекают Т-лимфоциты, называемые Т-клетками-киллерами (также называемые CD8 -положительными или цитотоксическими Т-клетками), которые разрушают клетки. В некоторых новых работах было высказано предположение, что антигены длиной более 10 аминокислот, 11-14 аминокислот, могут быть представлены на MHC I, вызывая цитотоксический Т-клеточный ответ. [5] Белки MHC класса I ассоциированы с β2-микроглобулином , который, в отличие от белков HLA, кодируется геном на хромосоме 15 .
HLA, соответствующие классу MHC II ( DP , DM , DO , DQ и DR ), представляют антигены извне клетки Т-лимфоцитам. Эти конкретные антигены стимулируют размножение Т-хелперных клеток (также называемых CD4 -положительными Т-клетками), которые, в свою очередь, стимулируют антитела продуцирующие В-клетки, , к выработке антител к этому специфическому антигену. Аутоантигены подавляются регуляторными Т-клетками . Предсказать, какие антигены (фрагменты) будут представлены иммунной системе определенным типом HLA, сложно, но используемая технология совершенствуется. [6]
HLA, соответствующие MHC класса III, кодируют компоненты системы комплемента .
У HLA есть и другие роли. Они важны для защиты от болезней. Они являются основной причиной отторжения трансплантата . Они могут защитить от рака или не защитить (если их уровень снижается из-за инфекции). [7] HLA также может быть связан с восприятием людьми запаха других людей и может участвовать в выборе партнера, поскольку по крайней мере одно исследование выявило более низкий, чем ожидалось, уровень сходства HLA между супругами в изолированном сообществе. [8]
Помимо генов, кодирующих шесть основных антигенпрезентирующих белков, в комплексе HLA расположены многие другие гены, многие из которых участвуют в иммунной функции. Разнообразие HLA в человеческой популяции является одним из аспектов защиты от болезней, и в результате вероятность появления двух неродственных людей с идентичными молекулами HLA во всех локусах чрезвычайно мала. Гены HLA исторически были идентифицированы как результат способности успешно трансплантировать органы между HLA-подобными людьми. [9]
Функции [ править ]
Белки , кодируемые HLA, находятся на внешней части клеток тела и (по сути) уникальны для этого человека. Иммунная система использует HLA для дифференциации собственных и чужих клеток. Любая клетка, отображающая тип HLA этого человека, принадлежит этому человеку и, следовательно, не является захватчиком.

При инфекционных заболеваниях [ править ]
Когда чужеродный патоген попадает в организм, специфические клетки, называемые антигенпрезентирующими клетками (АПК), поглощают возбудитель посредством процесса, называемого фагоцитозом . Белки патогена перевариваются на небольшие кусочки ( пептиды ) и загружаются на антигены HLA (точнее, MHC класса II ). Затем они экспонируются антигенпредставляющими клетками клеткам CD4+ Т-хелперным . [10] которые затем производят различные эффекты и межклеточные взаимодействия для устранения патогена.
Посредством аналогичного процесса белки (как нативные, так и чужеродные, такие как белки вирусов), вырабатываемые внутри большинства клеток, отображаются на HLA (точнее, MHC класса I ) на поверхности клетки. Инфицированные клетки могут быть распознаны и уничтожены CD8+ Т-клетками . [10]
На изображении сбоку показан фрагмент ядовитого бактериального белка (пептид SEI), связанный внутри связывающей щели молекулы HLA-DR1. На иллюстрации ниже, другой вид: можно увидеть целый DQ со связанным пептидом в аналогичной щели, если смотреть сбоку. Пептиды, связанные с болезнями, вписываются в эти «слоты» так же, как рука в перчатку.
При связывании пептиды презентируются Т-клеткам. Т-клеткам требуется презентация через молекулы MHC для распознавания чужеродных антигенов — требование, известное как ограничение MHC . Т-клетки имеют рецепторы, аналогичные рецепторам В-клеток, и каждая Т-клетка распознает только несколько комбинаций пептидов MHC класса II. Как только Т-клетка распознает пептид в молекуле MHC класса II, она может стимулировать В-клетки, которые также распознают ту же молекулу в своих В-клеточных рецепторах. Таким образом, Т-клетки помогают В-клеткам вырабатывать антитела к тем же чужеродным антигенам. Каждый HLA может связывать множество пептидов, и каждый человек имеет 3 типа HLA и может иметь 4 изоформы DP, 4 изоформы DQ и 4 изоформы DR (2 DRB1 и 2 DRB3, DRB4 или DRB5), всего 12 изоформ. У таких гетерозигот белкам, связанным с заболеванием, трудно избежать обнаружения.
При отторжении трансплантата [ править ]
Любая клетка, отображающая какой-либо другой тип HLA, является «чужой» и рассматривается иммунной системой организма как захватчик, что приводит к отторжению ткани, несущей эти клетки. Это особенно важно в случае трансплантированной ткани, поскольку может привести к отторжению трансплантата . Из-за важности HLA при трансплантации локусы HLA являются одними из наиболее часто типируемых с помощью серологии и ПЦР. Было показано, что типирование HLA с высоким разрешением (HLA-A, HLA-B, HLA-C, HLA-DRB1, HLA-DQB1 и HLA-DPB1) может быть актуальным при трансплантации для выявления полного совпадения, даже если донор связанный. [11]
| HLA аллель | Заболевания с повышенным риском | Относительный риск |
|---|---|---|
| HLA-B27 | Анкилозирующий спондилоартрит | 12 [12] |
| Реактивный артрит | 14 [12] | |
| Острый передний увеит | 15 [12] | |
| HLA-B47 | Дефицит 21-гидроксилазы | 15 [12] |
| HLA-B51 | Болезнь Бехчета | |
| HLA-DR2 | Системная красная волчанка | 2 к 3 [13] |
| HLA-DR3 | Аутоиммунный гепатит | 14 [12] |
| Первичный синдром Шегрена | 10 [12] | |
| Сахарный диабет 1 типа | 5 [12] | |
| Системная красная волчанка | 2 к 3 [13] | |
| HLA-DR4 | Ревматоидный артрит | 4 [12] |
| Сахарный диабет 1 типа | 6 [12] | |
| HLA-DR3 и -DR4 комбинированный | Сахарный диабет 1 типа | 15 [12] |
| HLA-DQ2 и HLA-DQ8 | Целиакия | 7 [14] |
При аутоиммунитете [ править ]
Типы HLA передаются по наследству, а некоторые из них связаны с аутоиммунными нарушениями и другими заболеваниями. Люди с определенными антигенами HLA более склонны к развитию определенных аутоиммунных заболеваний, таких как диабет I типа , анкилозирующий спондилит , ревматоидный артрит , [15] целиакия , СКВ (системная красная волчанка), миастения , миозит с включенными тельцами , синдром Шегрена и нарколепсия . [16]
Типирование HLA привело к некоторому улучшению и ускорению диагностики целиакии и диабета 1 типа; однако для того, чтобы типирование DQ2 было полезным, требуется либо типирование B1* с высоким разрешением (разрешение *02:01 из *02:02), типирование DQA1*, либо серотипирование DR . Текущее серотипирование позволяет решить проблему DQ8 за один этап. Типирование HLA при аутоиммунитете все чаще используется в качестве инструмента диагностики. При целиакии это единственный эффективный способ отличить родственников первой степени родства, находящихся в группе риска, от тех, кто не подвержен риску, до появления иногда необратимых симптомов, таких как аллергия и вторичное аутоиммунное заболевание.
При раке [ править ]
Некоторые HLA-опосредованные заболевания напрямую способствуют развитию рака. Чувствительная к глютену энтеропатия связана с увеличением распространенности Т-клеточной лимфомы, связанной с энтеропатией, а гомозиготы DR3-DQ2 относятся к группе самого высокого риска: около 80% случаев Т-клеточной лимфомы, связанной с глютен-чувствительной энтеропатией. Однако чаще молекулы HLA играют защитную роль, распознавая увеличение количества антигенов, которые не переносятся из-за низких уровней в нормальном состоянии. Аномальные клетки могут стать мишенью апоптоза, который, как считается, является посредником во многих случаях рака до постановки диагноза.
В выборе партнера [ править ]
Существуют доказательства неслучайного выбора партнера в отношении определенных генетических характеристик. [17] [18] Это привело к появлению области, известной как генетическое сватовство .
Классификация [ править ]
Белки MHC класса I образуют функциональный рецептор на большинстве ядросодержащих клеток организма. [1]
В HLA есть три основных и три второстепенных гена MHC класса I.
Майор MHC I класса
Минорные гены — HLA-E , HLA-F и HLA-G . β 2 -микроглобулин связывается с основными и второстепенными субъединицами гена, образуя гетеродимер.

Существует три основных и два второстепенных белка MHC класса II, кодируемых HLA.Гены класса II объединяются с образованием гетеродимерного (αβ) белка.рецепторы, которые обычно экспрессируются на поверхности антигенпрезентирующих клеток .
Основные белки MHC класса II встречаются только на антигенпрезентирующих клетках , В-клетках и Т-клетках . [1]
- HLA-DP
- α-цепь, кодируемая локусом HLA-DPA1
- β-цепь, кодируемая HLA-DPB1 локусом
- HLA-DQ
- HLA-DR
- α-цепь, кодируемая локусом HLA-DRA
- 4 β-цепи (возможно только 3 на человека), кодируемые локусами HLA-DRB1, DRB3, DRB4, DRB5
Другие белки MHC класса II, DM и DO, используются во внутреннем процессинге антигенов, загружая антигенные пептиды, образующиеся из патогенов, на молекулы HLA антигенпредставляющей клетки .
Номенклатура [ править ]
Современные аллели HLA обычно отмечаются с разной степенью детализации. Большинство обозначений начинаются с HLA- и названия локуса, затем * и некоторого (четного) количества цифр, определяющих аллель. Первые две цифры обозначают группу аллелей, также известную как супертипы. Старые методы типирования часто не могли полностью различить аллели и поэтому останавливались на этом уровне. Цифры с третьей по четвертую обозначают несинонимичный аллель. Цифры с пятой по шестую обозначают любые синонимичные мутации в кодирующей рамке гена. Седьмая и восьмая цифры различают мутации за пределами кодирующей области. Такие буквы, как L, N, Q или S, могут следовать за обозначением аллели, чтобы указать уровень экспрессии или другие известные о нем негеномные данные. Таким образом, полностью описанный аллель может иметь длину до 9 цифр, не считая HLA-префикса и обозначения локуса. [19]
Вариативность [ править ]

Локусы MHC являются одними из наиболее генетически вариабельных кодирующих локусов у млекопитающих, и локусы HLA человека не являются исключением. Несмотря на то, что человеческая популяция за свою историю несколько раз переживала сужение, способное фиксировать многие локусы, локусы HLA, по-видимому, пережили такое сужение с большим количеством вариаций. [20] Из 9 упомянутых выше локусов большинство сохранило дюжину или более аллельных групп для каждого локуса, что представляет собой гораздо более сохранившуюся вариацию, чем подавляющее большинство локусов человека. Это согласуется с гетерозиготным или балансирующим коэффициентом отбора для этих локусов. Кроме того, некоторые локусы HLA относятся к числу наиболее быстро развивающихся кодирующих областей генома человека. Один механизм диверсификации был отмечен при изучении амазонских племен Южной Америки, которые, по-видимому, претерпели интенсивную конверсию генов между вариабельными аллелями и локусами внутри каждого класса генов HLA. [21] Реже были отмечены продуктивные рекомбинации большего радиуса действия через гены HLA с образованием химерных генов.
Шесть локусов содержат более 100 аллелей, обнаруженных в человеческой популяции. Из них наиболее вариабельными являются HLA B и HLA DRB1. По состоянию на 2012 год количество определенных аллелей указано в таблице ниже. Чтобы интерпретировать эту таблицу, необходимо учитывать, что аллель представляет собой вариант последовательности нуклеотидов (ДНК) в локусе, такой, что каждая аллель отличается от всех других аллелей по крайней мере в одном положении (однонуклеотидный полиморфизм, SNP). Большинство этих изменений приводят к изменению аминокислотных последовательностей, что приводит к незначительным или значительным функциональным различиям в белке.
Существуют проблемы, которые ограничивают этот вариант. Определенные аллели, такие как DQA1*05:01 и DQA1*05:05, кодируют белки с идентично обработанными продуктами. Другие аллели, такие как DQB1*0201 и DQB1*0202, производят белки, функционально схожие. Для класса II (DR, DP и DQ) варианты аминокислот в пептидсвязывающей щели рецептора имеют тенденцию продуцировать молекулы с различной способностью связывания.
Однако частоты генов наиболее распространенных аллелей (>5%) HLA-A, -B, -C и HLA-DPA1, -DPB1, -DQA1, -DQB1 и -DRB1 из Южной Америки были получены из Южной Америки. типирование и секвенирование, проводимые в ходе исследований генетического разнообразия, а также случаев и контроля. [22] Кроме того, собрана информация о частотах аллелей генов HLA-I и HLA-II для европейского населения. [23] [24] В обоих случаях распределение частот аллелей демонстрирует региональные различия, связанные с историей популяций.
Таблицы вариантов аллелей
Количество вариантных аллелей в локусах класса I по данным базы данных IMGT-HLA, последнее обновление — октябрь 2018 г.:
| МХК I класса | |
|---|---|
| локус | # [25] [26] |
| Основные антигены | |
| ПЕСНИ А | 4,340 |
| HLA Б | 5,212 |
| HLA С | 3,930 |
| Минорные антигены | |
| HLA E | 27 |
| HLA Ф | 31 |
| HLA G | 61 |
Количество вариантов аллелей в локусах класса II (DM, DO, DP, DQ и DR):
| МХК класс II | ||||
|---|---|---|---|---|
| HLA | -А1 | -B1 | от -B3 до -B5 1 | Теор. возможный |
| локус | # [26] | # [26] | # [26] | комбинации |
| ДМ- | 7 | 13 | 91 | |
| ДЕЛАТЬ- | 12 | 13 | 156 | |
| ДП- | 67 | 1,014 | 16,036 | |
| DQ- | 95 | 1,257 | 34,528 | |
| ДР- | 7 | 2,593 | 312 | 11,431 |
| 1 DRB3, DRB4, DRB5 присутствуют у людей по-разному. | ||||
Тип варианта объекта последовательности (SFVT) [ править ]
Большая степень изменчивости генов HLA создает серьезные проблемы при исследовании роли генетических вариаций HLA в заболеваниях. Исследования ассоциации заболеваний обычно рассматривают каждый аллель HLA как единую целостную единицу, которая не освещает части молекулы, связанные с заболеванием. Карп Д.Р. и др. описывает новый подход типа варианта признака последовательности (SFVT) для генетического анализа HLA, который классифицирует белки HLA на биологически значимые меньшие признаки последовательности (SF) и их варианты типов (VT). [27] Характеристики последовательности представляют собой комбинации аминокислотных участков, определенные на основе структурной информации (например, бета-лист 1), функциональной информации (например, связывания пептидного антигена) и полиморфизма. Эти особенности последовательности могут перекрываться, быть непрерывными или прерывистыми в линейной последовательности. Типы вариантов для каждого признака последовательности определяются на основе всех известных полиморфизмов в описываемом локусе HLA. Классификация HLA по SFVT применяется при анализе генетических ассоциаций, чтобы можно было идентифицировать эффекты и роль эпитопов, общих для нескольких аллелей HLA. Особенности последовательностей и их варианты описаны для всех классических белков HLA; международный репозиторий HLA SFVT будет храниться в базе данных IMGT/HLA. [28] Инструмент для преобразования аллелей HLA в составляющие их SFVT можно найти на веб-сайте портала базы данных и анализа иммунологии (ImmPort). [29]
, хорошо документированные и редкие Распространенные аллели
Хотя количество идентифицированных отдельных аллелей HLA велико, примерно 40% этих аллелей кажутся уникальными, поскольку были идентифицированы только у отдельных людей. [30] [31] Примерно треть аллелей была зарегистрирована более трех раз у неродственных лиц. [31] [32] Из-за этой разницы в скорости обнаружения отдельных аллелей HLA были предприняты попытки классифицировать аллели в каждом экспрессируемом локусе HLA с точки зрения их распространенности. Результатом стал каталог распространенных и хорошо документированных (CWD) аллелей HLA. [32] [33] и каталог редких и очень редких аллелей HLA. [30] [31]
Общие аллели HLA определяются как встречающиеся с частотой не менее 0,001 в контрольных популяциях, насчитывающих не менее 1500 человек. [32] [33] Хорошо документированные аллели HLA первоначально определялись как обнаруженные как минимум трижды у неродственных лиц. [32] и теперь определяются как обнаруженные как минимум пять раз у неродственных людей с помощью метода типирования на основе последовательностей (SBT) или как минимум три раза с помощью метода SBT и в определенном гаплотипе у неродственных людей. [33] Редкие аллели определяются как те, о которых сообщалось от одного до четырех раз, а очень редкие аллели — как те, о которых сообщалось только один раз. [30] [31]
Таблица аллелей HLA в распространенности каждой категории
Хотя нынешние CWD и редкие или очень редкие обозначения были разработаны с использованием разных наборов данных и разных версий базы данных IMGT/HLA , [31] [33] приблизительная доля аллелей в каждом локусе HLA в каждой категории показана ниже.
| HLA Локус | № общий аллели [33] | % общий аллели [33] | Нет. хорошо документировано аллели [33] | % хорошо документировано аллели [33] | Нет. Редкий аллели [31] | % редкий аллели [31] | Нет, очень редко аллели [31] | % очень редко аллели [31] | % аллелей категоризированный |
|---|---|---|---|---|---|---|---|---|---|
| А | 68 | 3.4% | 178 | 8.8% | 145 | 21.5% | 280 | 41.6% | ~75% |
| Б | 125 | 4.8% | 242 | 9.3% | 190 | 17.6% | 468 | 43.5% | ~75% |
| С | 44 | 2.8% | 102 | 6.6% | 77 | 21.4% | 154 | 42.8% | ~74% |
| ДРБ1 | 79 | 6.8% | 147 | 12.7% | 133 | 22.7% | 206 | 35.2% | ~77% |
| ДРБ3 | 5 | 8.6% | 7 | 12.1% | ~21% | ||||
| ДРБ4 | 6 | 40.0% | 2 | 13.3% | ~53% | ||||
| ДРБ5 | 5 | 25.0% | 3 | 15.0% | ~40% | ||||
| DQA1 | 15 | 31.9% | 4 | 8.5% | 9 | 26.5% | 7 | 20.6% | ~88% |
| DQB1 | 22 | 12.5% | 8 | 4.5% | 26 | 28.9% | 42 | 45.2% | ~91% |
| ДПА1 | 6 | 17.6% | 0 | 0.0% | 4 | 14.8% | 15 | 55.6% | ~88% |
| ДПБ1 | 40 | 28.8% | 14 | 9.0% | 29 | 22.7% | 29 | 32.8% | ~90% |
| Все локусы | 415 | 5.3% | 707 | 9.0% | 613 | 20.6% | 1214 | 40.8% | ~76% |
HLA Изучение типов
Названия серотипов и аллелей [ править ]
К HLA применяются две параллельные системы номенклатуры. Первая и самая старая система основана на серологическом распознавании (на основе антител). В этой системе антигенам в конечном итоге были присвоены буквы и цифры (например, HLA-B27 или сокращенно B27). Была разработана параллельная система, позволившая более точно определить аллели. В этой системе «HLA» используется в сочетании с буквой * и четырехзначным или более цифровым номером (например, HLA-B*08:01, A*68:01, A*24:02). :01N N=Null) для обозначения конкретной аллели в данном локусе HLA . Локусы HLA можно дополнительно разделить на MHC класса I и MHC класса II (или, реже, локус D). Каждые два года создается номенклатура, которая помогает исследователям интерпретировать серотипы и аллели. [25]
Серотипирование [ править ]
Для создания реагента для типирования берется кровь животных или людей, клетки крови отделяются от сыворотки, а сыворотка разбавляется до оптимальной чувствительности и используется для типирования клеток от других людей или животных. Таким образом, серотипирование стало способом грубой идентификации рецепторов HLA и изоформ рецепторов. С годами серотипирующие антитела стали более совершенными по мере совершенствования методов повышения чувствительности и постоянного появления новых серотипирующих антител. Одна из целей анализа серотипов — заполнить пробелы в анализе. Можно прогнозировать на основе «квадратного корня», метода «максимального правдоподобия» или анализа семейных гаплотипов для учета адекватно типизированных аллелей. Эти исследования с использованием методов серотипирования часто выявляли, особенно в популяциях за пределами Европы или Северо-Восточной Азии, множество нулевых или пустых серотипов. До недавнего времени это было особенно проблематично для локуса Cw, и почти половина серотипов Cw не была типизирована в ходе обследования человеческой популяции в 1991 году.
Существует несколько типов серотипов. Широкий серотип антигена является грубым показателем идентичности клеток. Например, серотип HLA A9 распознает клетки людей, несущих A23 и A24. Он также может распознавать клетки, которые A23 и A24 пропускают из-за небольших различий. А23 и А24 являются расщепленными антигенами, но антитела, специфичные к каждому из них, обычно используются чаще, чем антитела к широким антигенам.
Сотовая типизация [ править ]
Репрезентативным клеточным анализом является смешанная культура лимфоцитов (MLC), используемая для определения типов HLA класса II. [34] Клеточные анализы более чувствительны при обнаружении различий HLA, чем серотипирование. Это связано с тем, что незначительные различия, не распознаваемые аллоантисыворотками, могут стимулировать Т-клетки. Такая типизация обозначается как типы Dw. Серотипированный DR1 на клеточном уровне определяется как Dw1 или Dw20 и так далее для других серотипированных DR. Стол [35] демонстрирует ассоциированную клеточную специфичность для аллелей DR. Однако клеточное типирование имеет противоречивые реакции между индивидуумами клеточного типа, иногда приводящие к результатам, отличным от прогнозируемых. Наряду с трудностью клеточного анализа в создании и поддержании реагентов для клеточного типирования, клеточный анализ заменяется методом типирования на основе ДНК. [34]
Секвенирование генов [ править ]
Незначительные реакции на субрегионы, демонстрирующие сходство с другими типами, можно наблюдать на генные продукты аллелей серотипической группы. Последовательность антигенов определяет реактивность антител, поэтому наличие хороших возможностей секвенирования (или типирования на основе последовательности) устраняет необходимость в серологических реакциях. Следовательно, реакции различных серотипов могут указывать на необходимость секвенирования HLA человека для определения новой последовательности гена.
Широкие типы антигенов по-прежнему полезны, например, для типирования очень разнообразных популяций со многими неидентифицированными аллелями HLA (Африка, Аравия, [36] Юго-Восточный Иран [37] и Пакистан, Индия [38] ). Африка, Южный Иран и Аравия демонстрируют трудности с типизацией территорий, которые были заселены ранее. Аллельное разнообразие делает необходимым использовать широкое типирование антигенов с последующим секвенированием генов, поскольку существует повышенный риск ошибочной идентификации с помощью методов серотипирования.
В конце концов, семинар, основанный на последовательности, решает, какой новый аллель входит в какую серогруппу либо по последовательности, либо по реактивности. После проверки последовательности ей присваивается номер. Например, новый аллель B44 может получить серотип (т.е. B44) и идентификатор аллели, т.е. B*44:65, поскольку это 65-й обнаруженный аллель B44. Марш и др. (2005) [25] можно считать сборником кодов серотипов и генотипов HLA, а также новой книгой, выпускаемой два раза в год с ежемесячными обновлениями в разделе « Тканевые антигены» .
Фенотипирование [ править ]
Типирование генов отличается от секвенирования генов и серотипирования.При этой стратегии используются праймеры для ПЦР, специфичные к вариантному участку ДНК (так называемые праймеры, специфичные для последовательности). Если обнаружен продукт нужного размера, предполагается, что аллель HLA идентифицирован. Новые последовательности генов часто приводят к усилению неопределенности. Поскольку типирование генов основано на SSP-PCR, возможно, что новые варианты, особенно в локусах класса I и DRB1, могут быть пропущены.
Например, SSP-PCR в клинической ситуации часто используется для выявления фенотипов HLA. Примером расширенного фенотипа человека может быть:
A *01:01 / *03:01 , C *07:01 / *07:02 , B *07:02 / *08:01 , DRB1 *03:01 / *15:01 , DQA1 *05:01 / *01:02 , DQB1 *02:01 / *06:02
В целом он идентичен расширенному серотипу:А1,А3,Б7,Б8,ДР3,ДР15(2), ДК2,ДК6(1)
Во многих популяциях, таких как японская или европейская популяция, было типировано так много пациентов, что новые аллели встречаются относительно редко, и, таким образом, SSP-PCR более чем достаточна для разрешения аллелей. Гаплотипы можно получить путем типирования членов семьи в тех регионах мира, где SSP-PCR не может распознать аллели, а типирование требует секвенирования новых аллелей. Районы мира, где SSP-PCR или серотипирование могут оказаться неадекватными, включают Центральную Африку, Восточную Африку, части южной Африки, Аравию, Южный Иран, Пакистан и Индию.
Гаплотипы [ править ]
Гаплотип HLA представляет собой серию «генов» HLA (локус-аллелей) в хромосоме, один из которых перешел от матери, а другой - от отца.
Приведенный выше фенотип является одним из наиболее распространенных в Ирландии и является результатом двух общих генетических гаплотипов :
А *01:01 ; С *07:01 ; Б *08:01 ; ДРБ1 *03:01 ; ДКА1 *05:01 ; DQB1 *02:01 (путем серотипирования A1-Cw7-B8-DR3-DQ2 )
который называется «супер B8» или «предковый гаплотип» и
А *03:01 ; С *07:02 ; Б *07:02 ; ДРБ1 *15:01 ; ДКА1 *01:02 ; DQB1 *06:02 (Набрав серотип A3-Cw7-B7-DR15-DQ6 или более старую версию «A3-B7-DR2-DQ1»)
Эти гаплотипы можно использовать для отслеживания миграций человеческой популяции, поскольку они часто напоминают отпечатки пальцев событий, произошедших в эволюции. Гаплотип Super-B8 увеличивается у западных ирландцев, снижается по мере удаления от этого региона и встречается только в тех регионах мира, куда мигрировали западные европейцы. «A3-B7-DR2-DQ1» распространен более широко: от Восточной Азии до Иберии. Гаплотип Super-B8 связан с рядом аутоиммунных заболеваний, связанных с питанием. Существует 100 000 расширенных гаплотипов, но лишь немногие из них демонстрируют видимый и узловой характер в человеческой популяции.
Роль аллельной вариации
Исследования на людях и животных предполагают, что механизм гетерозиготного отбора, действующий на эти локусы, объясняет эту изменчивость. [39] Одним из предложенных механизмов является половой отбор, при котором самки способны обнаруживать самцов с HLA, отличным от их собственного типа. [40] Хотя локусы, кодирующие DQ и DP, имеют меньше аллелей, комбинации A1:B1 могут давать теоретический потенциал 7755 гетеродимеров DQ и 5270 DP αβ соответственно. Хотя в человеческой популяции не существует такого количества изоформ, каждый человек может нести 4 варьируемые изоформы DQ и DP, увеличивая потенциальное количество антигенов, которые эти рецепторы могут представить иммунной системе.
Исследования вариабельных положений DP, DR и DQ показывают, что контактные остатки пептида с антигеном на молекулах класса II чаще всего являются местом изменения первичной структуры белка. Таким образом, благодаря сочетанию интенсивных вариаций аллелей и/или спаривания субъединиц пептидные рецепторы класса II способны связывать почти бесконечные вариации пептидов длиной 9 аминокислот или более, защищая межпородные субпопуляции от зарождающихся или эпидемических заболеваний. Особи в популяции часто имеют разные гаплотипы, и это приводит к множеству комбинаций, даже в небольших группах. Это разнообразие увеличивает выживаемость таких групп и препятствует эволюции эпитопов патогенов, которые в противном случае могли бы быть защищены от иммунной системы.
Антитела [ править ]
Антитела HLA обычно не встречаются в природе и, за некоторыми исключениями, образуются в результате иммунологического заражения инородным материалом, содержащим чужеродные HLA, при переливании крови, беременности (антигены, унаследованные по отцовской линии) или трансплантации органов или тканей.
Антитела против ассоциированных с заболеванием гаплотипов HLA были предложены для лечения тяжелых аутоиммунных заболеваний. [41]
Было обнаружено, что донор-специфические антитела HLA связаны с недостаточностью трансплантата при трансплантации почек, сердца, легких и печени. Эти донор-специфичные HLA-антитела могут существовать до трансплантации вследствие сенсибилизации к предшествующим трансплантатам или во время беременности, но также могут возникать de novo после трансплантации. Существует четкая связь между риском сенсибилизации HLA-антител и несоответствием HLA (молекулярного) донора-реципиента. [42]
Сопоставление HLA у больных братьев и сестер [ править ]
При некоторых заболеваниях, требующих трансплантации гемопоэтических стволовых клеток , может использоваться преимплантационная генетическая диагностика для рождения брата или сестры с совпадающим HLA, хотя существуют этические соображения. [43]
См. также [ править ]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с Чу С.Ю. (февраль 2007 г.). «Система HLA: генетика, иммунология, клинические испытания и клинические последствия» . Медицинский журнал Йонсей . 48 (1): 11–23. дои : 10.3349/ymj.2007.48.1.11 . ПМК 2628004 . ПМИД 17326240 .
- ^ «Человеческий лейкоцитарный антиген (HLA)» . МС Траст . декабрь 2020 года . Проверено 7 июня 2021 г.
- ^ Накамура Т., Ширузу Т., Наката К., Ёсимура Н., Ушигоме Х. (сентябрь 2019 г.). «Роль главного комплекса гистосовместимости в трансплантации органов: анализ донорских специфических антител к главному комплексу гистосовместимости переходит на следующий этап» . Международный журнал молекулярных наук . 20 (18): 4544. doi : 10.3390/ijms20184544 . ПМК 6769817 . ПМИД 31540289 .
- ^ Мацумура М., Фремонт Д.Х., Петерсон П.А., Уилсон И.А. (август 1992 г.). «Новые принципы распознавания пептидных антигенов молекулами MHC класса I». Наука . 257 (5072): 927–34. Бибкод : 1992Sci...257..927M . дои : 10.1126/science.1323878 . ПМИД 1323878 .
- ^ Берроуз С.Р., Россджон Дж., МакКласки Дж. (январь 2006 г.). «Не слишком ли мы ограничились картированием эпитопов CTL?». Тенденции в иммунологии . 27 (1): 11–6. дои : 10.1016/j.it.2005.11.001 . ПМИД 16297661 .
- ^ Бузид Р., де Бейер М.Т., Луитен Р.Дж., Безстарости К., Кесслер А.Л., Бруно М.Дж., Пеппеленбош М.П., Деммерс Дж.А., Бушоу С.И. (май 2021 г.). «Эмпирическая оценка использования вычислительного связывания HLA в качестве раннего фильтра для рабочего процесса обнаружения эпитопов на основе масс-спектрометрии» . Раки . 13 (10): 2307. doi : 10.3390/cancers13102307 . ПМК 8150281 . ПМИД 34065814 .
- ^ Гэлбрейт В., Вагнер М.К., Чао Дж., Абаза М., Эрнст Л.А., Недерлоф М.А. и др. (1991). «Визуальная цитометрия методом многопараметрической флуоресценции». Цитометрия . 12 (7): 579–96. дои : 10.1002/cyto.990120702 . ПМИД 1782829 .
- ^ Бреннан П.А., Кендрик К.М. (декабрь 2006 г.). «Социальные запахи млекопитающих: привлекательность и индивидуальное узнавание» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 361 (1476): 2061–78. дои : 10.1098/rstb.2006.1931 . ПМЦ 1764843 . ПМИД 17118924 .
- ^ Сингал Д.П., Микки М.Р., Миттал К.К., Терасаки П.И. (ноябрь 1968 г.). «Серотипирование для гомотрансплантации. XVII. Предварительные исследования субъединиц и аллелей HL-A» . Трансплантация . 6 (8): 904–912. дои : 10.1097/00007890-196811000-00005 . ПМИД 5696819 . S2CID 32428268 .
- ^ Jump up to: Перейти обратно: а б Тейлор С.Дж., Болтон Э.М., Брэдли Дж.А. (август 2011 г.). «Иммунологические соображения по банкированию эмбриональных и индуцированных плюрипотентных стволовых клеток» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 366 (1575): 2312–22. дои : 10.1098/rstb.2011.0030 . ПМК 3130422 . ПМИД 21727137 .
- ^ Агарвал Р.К., Кумари А., Седай А., Пармар Л., Дханья Р., Фолкнер Л. (сентябрь 2017 г.). «Обоснование расширенного типирования HLA с 6 локусами с высоким разрешением для идентификации родственных доноров на Индийском субконтиненте» . Биология трансплантации крови и костного мозга . 23 (9): 1592–1596. дои : 10.1016/j.bbmt.2017.05.030 . ПМИД 28603069 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Таблица 5-7 в: Митчелл Р.С., Кумар В., Аббас А.К., Фаусто Н. (2007). Основная патология Роббинса . Филадельфия: Сондерс. ISBN 978-1-4160-2973-1 . 8-е издание.
- ^ Jump up to: Перейти обратно: а б Значения даны для европеоидов согласно странице 61 (правый столбец) в: Салмон Дж., Уоллес DJ, Дюбуа Э.Л., Киру К.А., Хан Б., Леман Т.А. (2007). Красная волчанка Дюбуа . Филадельфия: Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-9394-0 .
- ^ Маргаритта-Жаннен П., Баброн М.К., Буржи М., Лука А.С., Клот Ф., Перкопо С. и др. (июнь 2004 г.). «Относительные риски HLA-DQ для целиакии в европейском населении: исследование Европейского генетического кластера по целиакии». Тканевые антигены . 63 (6): 562–7. дои : 10.1111/j.0001-2815.2004.00237.x . ПМИД 15140032 .
- ^ Курко Ю., Бесеньей Т., Лаки Дж., Глант Т.Т., Микеч К., Секанеч З. (октябрь 2013 г.). «Генетика ревматоидного артрита – комплексный обзор» . Клинические обзоры по аллергии и иммунологии . 45 (2): 170–9. дои : 10.1007/s12016-012-8346-7 . ПМЦ 3655138 . ПМИД 23288628 .
- ^ Миньо Э., Лин Л., Роджерс В., Хонда Ю., Цю Х., Лин Х. и др. (март 2001 г.). «Сложные взаимодействия HLA-DR и -DQ повышают риск нарколепсии-катаплексии в трех этнических группах» . Американский журнал генетики человека . 68 (3): 686–99. дои : 10.1086/318799 . ПМЦ 1274481 . ПМИД 11179016 .
- ^ Йокиниеми А., Магрис М., Ритари Дж., Куусипало Л., Лундгрен Т., Партанен Дж., Кекяляйнен Дж. (август 2020 г.). «Посткопулятивное генетическое спаривание: HLA-зависимое влияние цервикальной слизи на функцию спермы человека» . Слушания. Биологические науки . 287 (1933): 20201682. doi : 10.1098/rspb.2020.1682 . ПМЦ 7482290 . ПМИД 32811307 .
- ^ Йокиниеми А., Куусипало Л., Ритари Дж., Коскела С., Партанен Ю., Кекяляйнен Дж. (ноябрь 2020 г.). «Иммуногенетическая несовместимость на уровне гамет у человека – к более глубокому пониманию оплодотворения и бесплодия?» . Наследственность . 125 (5): 281–289. дои : 10.1038/s41437-020-0350-8 . ПМЦ 7553942 . ПМИД 32747723 . S2CID 220947699 .
- ^ «Номенклатура HLA @ hla.alleles.org» . hla.alleles.org . Архивировано из оригинала 2 мая 2018 года . Проверено 2 мая 2018 г.
- ^ Шеннан Д.Х. (2006). Эволюция и спираль технологий . Траффорд Паблишинг. ISBN 978-1-55212-518-2 .
- ^ Пархэм П., Охта Т. (апрель 1996 г.). «Популяционная биология презентации антигена молекулами MHC класса I». Наука . 272 (5258): 67–74. Бибкод : 1996Sci...272...67P . дои : 10.1126/science.272.5258.67 . ПМИД 8600539 . S2CID 22209086 . .
- ^ Рекена Д., Медико А., Чакон Р.Д., Рамирес М., Марин-Санчес О. (2020). «Идентификация новых эпитопов-кандидатов на белках SARS-CoV-2 для Южной Америки: обзор частот HLA по странам» . Границы в иммунологии . 11 : 2008. doi : 10.3389/fimmu.2020.02008 . ПМЦ 7494848 . ПМИД 33013857 .
- ^ Нуньес Х.М., Бюлер С., Россли Д., Санчес-Масас А. (май 2014 г.). «Конвейер HLA-net GENE[RATE] для эффективного анализа данных HLA и его применения к 145 выборкам населения из Европы и соседних регионов». Тканевые антигены . 83 (5): 307–23. дои : 10.1111/tan.12356 . ПМИД 24738646 .
- ^ Санчес-Масас А., Бюлер С., Нуньес Х.М. (2013). «Новая карта Европы HLA: региональные генетические вариации и их значение для истории населения, исследования ассоциаций болезней и трансплантации тканей» . Наследственность человека . 76 (3–4): 162–77. дои : 10.1159/000360855 . ПМИД 24861861 .
- ^ Jump up to: Перейти обратно: а б с Марш С.Г., Альберт Э.Д., Бодмер В.Ф., Бонтроп Р.Э., Дюпон Б., Эрлих Х.А. и др. (апрель 2010 г.). «Номенклатура факторов системы HLA, 2010» . Тканевые антигены . 75 (4): 291–455. дои : 10.1111/j.1399-0039.2010.01466.x . ПМЦ 2848993 . ПМИД 20356336 .
- ^ Jump up to: Перейти обратно: а б с д Веб-службы EBI. «Статистика <IMGT/HLA <IPD <EMBL-EBI» . www.ebi.ac.uk. Архивировано из оригинала 20 сентября 2012 года . Проверено 2 мая 2018 г.
- ^ Карп Д.Р., Мартандан Н., Марш С.Г., Ан С., Арнетт Ф.К., Делука Д.С. и др. (февраль 2010 г.). «Новый анализ вариантов последовательностей генетической ассоциации HLA при системной склеродермии» . Молекулярная генетика человека . 19 (4): 707–19. дои : 10.1093/hmg/ddp521 . ПМК 2807365 . ПМИД 19933168 .
- ^ «База данных IMGT/HLA» . Архивировано из оригинала 24 сентября 2006 года.
- ^ «Портал базы данных и анализа иммунологии (ImmPort)» . Архивировано из оригинала 26 июля 2011 года.
- ^ Jump up to: Перейти обратно: а б с Миддлтон Д., Гонсалес Ф., Фернандес-Вина М., Тирси Дж.М., Марш С.Г., Обри М. и др. (декабрь 2009 г.). «Биоинформатический подход к определению редкости аллелей HLA». Тканевые антигены . 74 (6): 480–5. дои : 10.1111/j.1399-0039.2009.01361.x . ПМИД 19793314 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я Гонсалес-Галарса Ф.Ф., Мак С.Дж., Холленбах Дж., Фернандес-Вина М., Сеттерхольм М., Кемпених Дж. и др. (февраль 2013 г.). «16-й IHIW: расширение количества ресурсов и биоинформатического анализа для исследования редких аллелей HLA». Международный журнал иммуногенетики . 40 (1): 60–5. дои : 10.1111/iji.12030 . ПМИД 23198982 . S2CID 205192491 .
- ^ Jump up to: Перейти обратно: а б с д Кано П., Клитц В., Мак С.Дж., Майерс М., Марш С.Г., Норин Х. и др. (май 2007 г.). «Распространенные и хорошо документированные аллели HLA: отчет Специального комитета Американского общества по гистосовместимости и иммуногенетике». Иммунология человека . 68 (5): 392–417. дои : 10.1016/j.humimm.2007.01.014 . ПМИД 17462507 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Мак С.Дж., Кано П., Холленбах Дж.А., Хе Дж., Херли С.К., Миддлтон Д. и др. (апрель 2013 г.). «Распространенные и хорошо документированные аллели HLA: обновление каталога CWD за 2012 г.» . Тканевые антигены . 81 (4): 194–203. дои : 10.1111/tan.12093 . ПМЦ 3634360 . ПМИД 23510415 .
- ^ Jump up to: Перейти обратно: а б Херли СК (1997). «Типирование HLA на основе ДНК для трансплантации». В Леффелл М.С., Донненберг А.Д., Роуз Н.Р., ред. (1997) Справочник по иммунологии человека. стр. 521–55, Бока-Ратон: CRC Press , ISBN 0-8493-0134-3 .
- ^ Бодмер Дж.Г., Марш С.Г., Альберт Э.Д., Бодмер В.Ф., Дюпон Б., Эрлих Х.А. и др. (май 1992 г.). «Номенклатура факторов системы HLA, 1991». Иммунология человека . 34 (1): 4–18. дои : 10.1016/0198-8859(92)90079-3 . ПМИД 1399721 .
- ^ Валлури В., Валлуэй В., Мустафа М., Сантош А., Миддлтон Д., Альварес М. и др. (август 2005 г.). «Частоты фенотипов HLA-A, HLA-B, HLA-DR и HLA-DQ в населении Объединенных Арабских Эмиратов». Тканевые антигены . 66 (2): 107–13. дои : 10.1111/j.1399-0039.2005.00441.x . ПМИД 16029430 .
- ^ Фарджадиан С., Нарусе Т., Кавата Х., Гадери А., Бахрам С., Иноко Х. (ноябрь 2004 г.). «Молекулярный анализ частот и гаплотипов аллелей HLA у белуджей Ирана по сравнению с родственными популяциями Пакистана». Тканевые антигены . 64 (5): 581–7. дои : 10.1111/j.1399-0039.2004.00302.x . ПМИД 15496201 .
- ^ Шанкаркумар У., Прасанавар Д., Гош К., Моханти Д. (май 2003 г.). «Частоты аллелей HLA A * 02 и ассоциации гаплотипов B у жителей Западной Индии». Иммунология человека . 64 (5): 562–6. дои : 10.1016/S0198-8859(03)00032-6 . ПМИД 12691707 .
- ^ Апаниус В., Пенн Д., Слев П.Р., Рафф Л.Р., Поттс В.К. (1997). «Природа отбора по главному комплексу гистосовместимости». Критические обзоры по иммунологии . 17 (2): 179–224. doi : 10.1615/critrevimmunol.v17.i2.40 . ПМИД 9094452 . .
- ^ Ведекинд С., Зеебек Т., Беттенс Ф., Папке А.Дж. (июнь 1995 г.). «MHC-зависимые предпочтения партнера у людей». Слушания. Биологические науки . 260 (1359): 245–9. Бибкод : 1995RSPSB.260..245W . дои : 10.1098/rspb.1995.0087 . ПМИД 7630893 . S2CID 34971350 .
- ^ Осима М., Дейтикер П., Асидзава Т., Атасси М.З. (май 2002 г.). «Вакцинация пептидом MHC класса II ослабляет клеточные и гуморальные реакции против tAChR и подавляет клинический EAMG». Аутоиммунитет . 35 (3): 183–90. дои : 10.1080/08916930290022270 . ПМИД 12389643 . S2CID 5690960 .
- ^ Сенев А, Куманс М, Лерут Е, Ван Сандт В, Керкхофс Дж, Даниэльс Л, Дрише М.В., Компернолле В, Спрангерс Б, Ван Лун Е, Каллемейн Дж, Клаас Ф, Тамбур А.Р., Вербеке Г., Кайперс Д., Эмондс М.П., Нэсенс М (сентябрь 2020 г.). «Нагрузка несоответствия эплетов и возникновение de Novo донор-специфичных анти-HLA-антител, отторжение и недостаточность трансплантата после трансплантации почки: наблюдательное когортное исследование» . J Am Soc Нефрол . 31 (9): 2193–204. дои : 10.1681/ASN.2020010019 . ПМЦ 7461684 . ПМИД 32764139 .
- ^ Верлинский Ю., Речицкий С., Скулкрафт В., Стром С., Кулиев А. (июнь 2001 г.). «Преимплантационная диагностика анемии Фанкони в сочетании с соответствием HLA» . ДЖАМА . 285 (24): 3130–3. дои : 10.1001/jama.285.24.3130 . ПМИД 11427142 .
Библиография [ править ]
- Дэниел М. Дэвис, Ген совместимости , Лондон, Penguin Books , 2014 ( ISBN 978-0-241-95675-5 ).
- Жолт Харсаньи и Ричард Хаттон, Ричард, Генетическое пророчество: За пределами двойной спирали , Лондон: Гранада, 1982 ( ISBN 0-246-11760-5 ).
Внешние ссылки [ править ]
- База данных последовательностей IMGT/HLA в Европейском институте биоинформатики
- hla.alleles.org
- Группа информатики HLA в Фонде Энтони Нолана
- Британское общество гистосовместимости и иммуногенетики
- Американское общество гистосовместимости и иммуногенетики
- Инструмент HistoCheck HLA для трансплантации органов и стволовых клеток
- Частоты аллелей в вариабельных локусах, связанных с иммунитетом
- Человек + лейкоциты + антигены в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- dbMHC Home, база данных NCBI по главному комплексу гистосовместимости.
- Проект редких аллелей в базе данных AlleleFrequency Net (AFND)
| Базы данных органов управления : Национальные |
|---|