Протеасома


Протеасомы — это белковые комплексы , которые разрушают меченные убиквитином белки путем протеолиза — химической реакции , разрывающей пептидные связи . Ферменты , которые помогают таким реакциям, называются протеазами .
Протеасомы являются частью основного механизма, с помощью которого клетки регулируют концентрацию определенных белков и разрушают неправильно свернутые белки . Белки помечены для деградации небольшим белком, называемым убиквитином . Реакция мечения катализируется ферментами, называемыми убиквитинлигазами . Как только белок помечен одной молекулой убиквитина, это сигнал для других лигаз о присоединении дополнительных молекул убиквитина. В результате образуется полиубиквитиновая цепь , которая связывается с протеасомой, что позволяет ей разрушать меченый белок. [1] В результате процесса деградации образуются пептиды длиной от семи до восьми аминокислот , которые затем можно разложить на более короткие аминокислотные последовательности и использовать для синтеза новых белков. [1]
Протеасомы обнаружены внутри всех эукариот и архей , а также у некоторых бактерий .У эукариот протеасомы расположены как в ядре , так и в цитоплазме . [2]
По структуре протеасома представляет собой цилиндрический комплекс, содержащий «ядро» из четырех уложенных друг на друга колец, образующих центральную пору. Каждое кольцо состоит из семи отдельных белков. Два внутренних кольца состоят из семи β-субъединиц , которые содержат от трех до семи активных центров протеазы . Эти сайты расположены на внутренней поверхности колец, так что целевой белок должен войти в центральную пору, прежде чем он разрушится. Каждое из двух внешних колец содержит по семь α-субъединиц , функция которых заключается в поддержании «ворот», через которые белки попадают в ствол. Эти α-субъединицы контролируются путем связывания с «кэп»-структурами или регуляторными частицами , которые распознают полиубиквитиновые метки, прикрепленные к белковым субстратам, и инициируют процесс деградации. Общая система убиквитинирования и протеасомной деградации известна как система убиквитин-протеасома . [3]
Путь протеасомной деградации важен для многих клеточных процессов, включая клеточный цикл , регуляцию экспрессии генов и реакцию на окислительный стресс . Важность протеолитической деградации внутри клеток и роль убиквитина в протеолитических путях была признана при присуждении Нобелевской премии по химии 2004 года Аарону Чехановеру , Авраму Гершко и Ирвину Роузу . [4]
Открытие
[ редактировать ]До открытия системы убиквитин-протеасома считалось, что деградация белков в клетках происходит главным образом за счет лизосом , мембраносвязанных органелл с кислой и заполненной протеазами внутренней частью, которые могут разлагать, а затем перерабатывать экзогенные белки и старые или поврежденные органеллы. [1] Однако работа Джозефа Этлингера и Альфреда Л. Голдберга в 1977 году по АТФ-зависимой деградации белков в ретикулоцитах , у которых отсутствуют лизосомы, предположила наличие второго механизма внутриклеточной деградации. [5] В 1978 году было показано, что он состоит из нескольких отдельных белковых цепей, что было новинкой среди протеаз того времени. [6] Более поздние работы по модификации гистонов привели к идентификации неожиданной ковалентной модификации белка-гистона за счет связи между лизиновой боковой цепью гистона и C-концевым глициновым остатком убиквитина , белка, функция которого неизвестна. [7] Затем было обнаружено, что ранее идентифицированный белок, связанный с протеолитической деградацией, известный как АТФ-зависимый фактор протеолиза 1 (APF-1), был тем же белком, что и убиквитин. [8] Протеолитическая активность этой системы была выделена в виде мультибелкового комплекса, первоначально названного Шервином Уилком и Мэрион Орловски мультикаталитическим протеиназным комплексом. [9] Позднее был открыт АТФ -зависимый протеолитический комплекс, ответственный за убиквитин-зависимую деградацию белков, получивший название 26S протеасома. [10] [11]
Большая часть ранних работ, приведших к открытию протеасомной системы убиквитина, была проведена в конце 1970-х и начале 1980-х годов в Технионе в лаборатории Аврама Гершко , где Аарон Чехановер работал аспирантом. Годовой творческий отпуск Гершко в лаборатории Ирвина Роуза в онкологическом центре Фокс Чейз позволил ему получить ключевые концептуальные идеи, хотя позже Роуз преуменьшил свою роль в этом открытии. [12] Все трое получили Нобелевскую премию по химии 2004 года за работу по открытию этой системы. [4]
Хотя данные электронной микроскопии, раскрывающие многоуровневую кольцевую структуру протеасомы, стали доступны в середине 1980-х годов, [13] первая структура коровой частицы протеасомы не была решена с помощью рентгеновской кристаллографии до 1994 года. [14] 26S протеасомы человека были расшифрованы первые атомные структуры голофермента в комплексе с полиубиквитилированным белковым субстратом В 2018 году с помощью криогенной электронной микроскопии , что выявило механизмы, с помощью которых субстрат распознается, деубиквитилируется, разворачивается и деградируется 26S протеасомой человека. [15]
Структура и организация
[ редактировать ]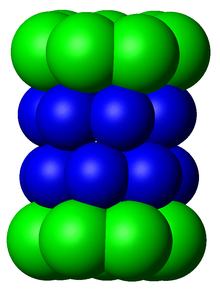
Субкомпоненты протеасом часто обозначаются их коэффициентом седиментации Сведберга (обозначается S ). Протеасома, наиболее часто используемая у млекопитающих, - это цитозольная 26S протеасома, которой составляет около 2000 килодальтон (кДа), молекулярная масса содержащая одну субъединицу белка 20S и две субъединицы регуляторного кэпа 19S. Ядро полое и представляет собой закрытую полость, в которой разлагаются белки; отверстия на двух концах ядра позволяют проникать целевому белку. Каждый конец сердцевинной частицы связан с регуляторной субъединицей 19S, которая содержит несколько АТФазы активных сайтов и сайтов связывания убиквитина; именно эта структура распознает полиубиквитинированные белки и переносит их в каталитическое ядро. [15] Альтернативная форма регуляторной субъединицы, называемая частицей 11S, может связываться с ядром практически так же, как частица 19S; 11S может играть роль в деградации чужеродных пептидов, например, тех, которые образуются после заражения вирусом . [16]
Основная частица 20S
[ редактировать ]Количество и разнообразие субъединиц, содержащихся в коровой частице 20S, зависит от организма; число отдельных и специализированных субъединиц больше у многоклеточных, чем у одноклеточных организмов, и больше у эукариот, чем у прокариот. Все частицы 20S состоят из четырех сложенных друг на друга гептамерных кольцевых структур, которые сами состоят из двух разных типов субъединиц; α-субъединицы имеют структурную природу, тогда как β-субъединицы преимущественно каталитические . Субъединицы α представляют собой псевдоферменты, гомологичные субъединицам β. Они собраны так, что их N-концы примыкают к концам β-субъединиц. [17] Два внешних кольца в стопке состоят из семи α-субъединиц каждое, которые служат стыковочными доменами для регуляторных частиц, а N-концы альфа-субъединиц ( Pfam PF10584 ) образуют ворота, которые блокируют нерегулируемый доступ субстратов во внутреннюю полость. [18] Каждое из двух внутренних колец состоит из семи β-субъединиц, а на их N-концах содержатся активные центры протеазы, которые осуществляют реакции протеолиза. [19] В очищенном комплексе были идентифицированы три различные каталитические активности: химотрипсиноподобная, трипсиноподобная и пептидилглутамилпептид-гидролизующая. [20] Размер протеасомы относительно консервативен и составляет около 150 ангстрем (Å) на 115 Å. Ширина внутренней камеры не превышает 53 Å, хотя вход может достигать 13 Å, что позволяет предположить, что для входа белки-субстраты должны быть хотя бы частично развернуты. [21]
У архей, таких как Thermoplasma acidophilum , все α- и все β-субъединицы идентичны, тогда как эукариотические протеасомы, такие как протеасомы дрожжей, содержат семь различных типов каждой субъединицы. У млекопитающих субъединицы β1, β2 и β5 являются каталитическими; хотя они имеют общий механизм, они имеют три различные субстратные специфичности: химотрипсиноподобные , трипсиноподобные и гидролизующие пептидилглутамилпептид (PHGH). [22] Альтернативные формы β, обозначаемые β1i, β2i и β5i, могут экспрессироваться в гемопоэтических клетках в ответ на воздействие провоспалительных сигналов , таких как цитокины , в частности интерферон гамма . Протеасома, собранная с этими альтернативными субъединицами, известна как иммунопротеасома , чья субстратная специфичность изменена по сравнению с нормальной протеасомой. [21] Недавно в клетках человека была обнаружена альтернативная протеасома, лишенная основной субъединицы α3. [23] Эти протеасомы (известные как протеасомы α4-α4) вместо этого образуют коровые частицы 20S, содержащие дополнительную субъединицу α4 вместо отсутствующей субъединицы α3. Ранее было известно, что эти альтернативные протеасомы «α4-α4» существуют у дрожжей. [24] Хотя точная функция этих изоформ протеасом до сих пор в значительной степени неизвестна, клетки, экспрессирующие эти протеасомы, демонстрируют повышенную устойчивость к токсичности, индуцированной ионами металлов, таких как кадмий. [23] [25]
Регуляторная частица 19S
[ редактировать ]Частица 19S у эукариот состоит из 19 отдельных белков и делится на две части: основание из 9 субъединиц, которое непосредственно связывается с α-кольцом коровой частицы 20S, и крышку из 10 субъединиц. Шесть из девяти основных белков представляют собой субъединицы АТФазы из семейства ААА, и у архей существует эволюционный гомолог этих АТФаз, называемый PAN (нуклеотидаза, активирующая протеасомы). [26] Ассоциация частиц 19S и 20S требует связывания АТФ с субъединицами 19S АТФазы, а гидролиз АТФ необходим для того, чтобы собранный комплекс разлагал свернутые и убиквитинированные белки. Обратите внимание, что только стадия разворачивания субстрата требует энергии гидролиза АТФ, тогда как одно лишь связывание АТФ может поддерживать все остальные этапы, необходимые для деградации белка (например, сборку комплекса, открытие ворот, транслокацию и протеолиз). [27] [28] Фактически, связывание АТФ с АТФазами само по себе способствует быстрой деградации развернутых белков. Однако, хотя гидролиз АТФ необходим только для разворачивания, пока неясно, можно ли использовать эту энергию для сопряжения некоторых из этих стадий. [28] [29]
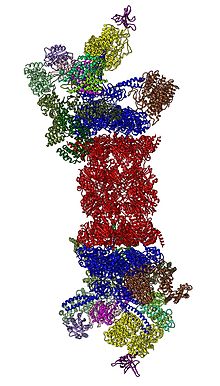
В 2012 году две независимые попытки выяснили молекулярную архитектуру протеасомы 26S с помощью одночастичной электронной микроскопии . [31] [32] В 2016 году три независимых усилия определили первую структуру человеческой 26S протеасомы с почти атомным разрешением в отсутствие субстратов с помощью криоЭМ. [33] [34] [35] В 2018 году были предприняты серьезные усилия для выяснения детальных механизмов деубиквитилирования, инициации транслокации и процессивного разворачивания субстратов путем одновременного определения семи атомных структур 26S-протеасомы, связанной с субстратом. [15] В сердце 19S, непосредственно примыкающем к 20S, находятся ААА-АТФазы ( белки ААА ), которые собираются в гетерогексамерное кольцо порядка Rpt1/Rpt2/Rpt6/Rpt3/Rpt4/Rpt5. Это кольцо представляет собой тример димеров: Rpt1/Rpt2, Rpt6/Rpt3 и Rpt4/Rpt5 димеризуются через свои N-концевые спиральные спирали. Эти спиральные катушки выступают из гексамерного кольца. Самые крупные регуляторные частицы, не относящиеся к АТФазам, Rpn1 и Rpn2, связываются с кончиками Rpt1/2 и Rpt6/3 соответственно. Убиквитиновый рецептор Rpn13 связывается с Rpn2 и завершает базовый подкомплекс. Крышка покрывает половину гексамера ААА-АТФазы (Rpt6/Rpt3/Rpt4) и неожиданно напрямую контактирует с 20S через Rpn6 и в меньшей степени Rpn5. Субъединицы Rpn9, Rpn5, Rpn6, Rpn7, Rpn3 и Rpn12, структурно родственные между собой и субъединицам комплекса COP9 и eIF3 (поэтому называемые субъединицами PCI), собираются в подковообразную структуру, включающую гетеродимер Rpn8/Rpn11. Rpn11, деубиквитинирующий фермент , помещается в устье гексамера ААА-АТФазы и идеально расположен для удаления фрагментов убиквитина непосредственно перед транслокацией субстратов в 20S. Второй идентифицированный на сегодняшний день убиквитиновый рецептор, Rpn10, расположен на периферии века, рядом с субъединицами Rpn8 и Rpn9.
Конформационные изменения 19S
[ редактировать ]Регуляторная частица 19S в составе голофермента 26S протеасомы наблюдалась в шести сильно различающихся конформационных состояниях в отсутствие субстратов. [36] [37] Отличительной чертой конфигурации ААА-АТФазы в этом преобладающем низкоэнергетическом состоянии является расположение ААА-доменов в виде лестницы или шайбы. [30] [31] В присутствии АТФ , но в отсутствие субстрата, принимаются три альтернативные, менее распространенные конформации 19S, отличающиеся главным образом расположением крышки относительно модуля ААА-АТФазы. [33] [37] В присутствии АТФ-γS или субстрата наблюдалось значительно больше конформаций, демонстрирующих резкие структурные изменения модуля ААА-АТФазы. [15] [36] [38] [39] Некоторые из связанных с субстратом конформаций очень похожи на бессубстратные, но они не полностью идентичны, особенно в модуле ААА-АТФазы. [15] [36] До сборки 26S регуляторная частица 19S в свободной форме также наблюдалась в семи конформационных состояниях. [40] Примечательно, что все эти конформеры несколько различаются и имеют разные особенности. Таким образом, регуляторная частица 19S может выбирать по меньшей мере 20 конформационных состояний в различных физиологических условиях.
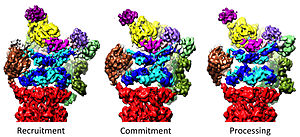
Регулирование 20-х годов 19-м
[ редактировать ]Регуляторная частица 19S отвечает за стимулирование 20S к расщеплению белков. Основная функция регуляторных АТФаз 19S — открыть ворота в 20S, которые блокируют вход субстратов в камеру разложения. [41] Недавно был выяснен механизм, с помощью которого протеасомная АТФаза открывает эти ворота. [18] Для открытия 20S-ворот и, следовательно, для деградации субстрата необходимы С-концы протеасомальных АТФаз, которые содержат специфический мотив (т.е. мотив HbYX). С-концы АТФазы связываются с карманами в верхней части 20S и привязывают комплекс АТФазы к протеолитическому комплексу 20S, соединяя таким образом оборудование для разворачивания субстрата с механизмом деградации 20S. Связывание этих С-концов с карманами 20S само по себе стимулирует открытие ворот в 20S почти так же, как «ключ в замке» открывает дверь. [18] Точный механизм функционирования этого механизма «ключ в замке» был структурно выяснен в контексте 26S протеасомы человека с разрешением, близким к атомному, что позволяет предположить, что вставка пяти С-концев субъединиц АТФазы Rpt1/2/ Для полного открытия ворот 20S необходимы 3/5/6 карманов на поверхности 20S. [36] [15] [33]
Другие регуляторные частицы
[ редактировать ]20S протеасомы также могут ассоциироваться со вторым типом регуляторных частиц, регуляторными частицами 11S, гептамерной структурой, которая не содержит никаких АТФаз и может способствовать деградации коротких пептидов , но не полных белков. Предполагается, что это связано с тем, что комплекс не может разворачивать более крупные субстраты. Эта структура также известна как PA28, REG или PA26. [17] Механизмы, с помощью которых он связывается с основной частицей через C-концевые хвосты ее субъединиц и вызывает конформационные изменения α-кольца , открывающие ворота 20S, предполагают аналогичный механизм для частицы 19S. [42] Экспрессия частицы 11S индуцируется гамма-интерфероном и вместе с β-субъединицами иммунопротеасомы ответственна за образование пептидов, которые связываются с главным комплексом гистосовместимости . [16]
Еще одним типом регуляторных частиц, не связанных с АТФазой, является Blm10 (дрожжи) или PA200/ PSME4 (человеческий). Он открывает только одну α-субъединицу в воротах 20S и сам сворачивается в купол с очень маленькой порой над ним. [17]
Сборка
[ редактировать ]Сборка протеасомы представляет собой сложный процесс из-за количества субъединиц, которые должны объединиться для образования активного комплекса. Субъединицы β синтезируются с N-концевыми «пропептидами», которые посттрансляционно модифицируются во время сборки частицы 20S, чтобы обнажить протеолитический активный центр. Частица 20S собрана из двух полупротеасом, каждая из которых состоит из семичленного про-β-кольца, присоединенного к семичленному α-кольцу. Объединение β-колец двух полупротеасом запускает треонин -зависимый аутолиз пропептидов с обнажением активного центра. Эти β-взаимодействия опосредуются главным образом солевыми мостиками и гидрофобными взаимодействиями между консервативными альфа-спиралями, разрушение которых в результате мутации повреждает способность протеасомы к сборке. [43] Сборка полупротеасом, в свою очередь, инициируется сборкой α-субъединиц в их гептамерное кольцо, образуя матрицу для ассоциации соответствующего про-β-кольца. Сборка α-субъединиц не охарактеризована. [44]
Лишь недавно процесс сборки регуляторной частицы 19S был в значительной степени выяснен. Регуляторная частица 19S состоит из двух отдельных подкомпонентов: основания и крышки. Сборке базового комплекса способствуют четыре шаперона сборки : Hsm3/S5b, Nas2/p27, Rpn14/PAAF1 и Nas6/ ганкирин (названия дрожжей/млекопитающих). [45] Эти шапероны сборки связываются с субъединицами ААА- АТФазы , и их основная функция, по-видимому, заключается в обеспечении правильной сборки гетерогексамерного кольца ААА- АТФазы . На сегодняшний день все еще обсуждается, собирается ли базовый комплекс отдельно, шаблонируется ли сборка основной частицей 20S или существуют ли альтернативные пути сборки. В дополнение к четырем шаперонам сборки, деубиквитинирующий фермент Ubp6/ Usp14 также способствует сборке оснований, но это не является существенным. [46] Крышка собирается отдельно в определенном порядке и не требует сопровождающих для сборки. [47]
Процесс деградации белка
[ редактировать ]
Убиквитинирование и таргетинг
[ редактировать ]Белки подвергаются деградации протеасомой с ковалентной модификацией остатка лизина, что требует скоординированных реакций трех ферментов . На первом этапе фермент, активирующий убиквитин (известный как E1), гидролизует АТФ и аденилирует молекулу убиквитина . в активном центре E1 Затем он переносится на остаток цистеина одновременно с аденилированием второго убиквитина. [48] Этот аденилированный убиквитин затем переносится на цистеин второго фермента, убиквитин-конъюгирующего фермента (Е2). На последнем этапе член весьма разнообразного класса ферментов, известных как убиквитинлигазы (Е3), распознает конкретный белок, подлежащий убиквитинированию, и катализирует перенос убиквитина от Е2 к этому белку-мишени. Целевой белок должен быть помечен как минимум четырьмя мономерами убиквитина (в форме полиубиквитиновой цепи), прежде чем он будет распознан крышкой протеасомы. [49] Следовательно, именно E3 придает субстратную специфичность. этой системе [50] Количество экспрессируемых белков E1, E2 и E3 зависит от организма и типа клеток, но у человека присутствует множество различных ферментов E3, что указывает на огромное количество мишеней для протеасомной системы убиквитина.
Механизм, с помощью которого полиубиквитинированный белок попадает в протеасому, до конца не изучен. Несколько снимков с высоким разрешением протеасомы, связанной с полиубиквитинированным белком, позволяют предположить, что рецепторы убиквитина могут координироваться с деубиквитиназой Rpn11 для первоначального нацеливания и взаимодействия с субстратом. [15] Белки убиквитиновых рецепторов имеют N-концевой убиквитин-подобный (UBL) домен и один или несколько убиквитин-ассоциированных (UBA) доменов. Домены UBL распознаются протеасомными кэпами 19S, а домены UBA связывают убиквитин посредством трехспиральных пучков . Эти рецепторные белки могут сопровождать полиубиквитинированные белки к протеасоме, хотя особенности этого взаимодействия и его регуляции неясны. [51]
Сам белок убиквитин последовательность имеет длину 76 аминокислот и получил такое название из-за своей повсеместной природы, поскольку он имеет высококонсервативную и встречается во всех известных эукариотических организмах. [52] Гены, кодирующие убиквитин у эукариот, расположены в виде тандемных повторов , возможно, из-за тяжелых требований транскрипции этих генов для производства достаточного количества убиквитина для клетки. Было высказано предположение, что убиквитин является самым медленно эволюционирующим белком, обнаруженным на сегодняшний день. [53] Убиквитин содержит семь остатков лизина, с которыми может быть связан другой убиквитин, в результате чего образуются различные типы полиубиквитиновых цепей. [54] Цепи, в которых каждый дополнительный убиквитин связан с лизином 48 предыдущего убиквитина, играют роль в нацеливании на протеасомы, тогда как другие типы цепей могут участвовать в других процессах. [55] [56]

Деубиквитилирование
[ редактировать ]Цепи убиквитина, конъюгированные с белком, предназначенным для протеасомной деградации, обычно удаляются любым из трех ассоциированных с протеасомами деубиквитилирующих ферментов (DUB), которыми являются Rpn11, Ubp6/USP14 и UCH37. Этот процесс перерабатывает убиквитин и необходим для поддержания резервуара убиквитина в клетках. [56] Rpn11 представляет собой внутреннюю стехиометрическую субъединицу регуляторной частицы 19S и необходим для функционирования протеасомы 26S. Активность DUB Rpn11 повышена в протеасоме по сравнению с его мономерной формой. То, как Rpn11 удаляет убиквитиновую цепь целиком из белкового субстрата, было зафиксировано атомной структурой связанной с субстратом человеческой протеасомы в конформации, названной EB . [15] Интересно, что эта структура также показывает, как активность DUB связана с распознаванием субстрата протеасомной AAA-АТФазой. В отличие от Rpn11, USP14 и UCH37 представляют собой DUB, которые не всегда связаны с протеасомой. Было обнаружено, что в клетках около 10-40% протеасом связаны с USP14. И Ubp6/USP14, и UCH37 в значительной степени активируются протеасомой и сами по себе демонстрируют очень низкую активность DUB. Было обнаружено, что после активации USP14 подавляет функцию протеасом за счет своей активности DUB и индуцирования параллельных путей конформационных переходов протеасом, один из которых, как оказалось, напрямую запрещает вставку субстрата в AAA-АТФазу, что интуитивно наблюдалось с помощью криогенной электронной микроскопии с временным разрешением. . [57] Похоже, что USP14 регулирует функцию протеасом во множественных контрольных точках, как каталитически конкурируя с Rpn11, так и аллостерически перепрограммируя состояния AAA-АТФазы, что довольно неожиданно для DUB. [57] Эти наблюдения подразумевают, что регуляция протеасомы может зависеть от ее динамических переходов конформационных состояний.
Разворачивание и перемещение
[ редактировать ]После убиквитинирования белка он распознается регуляторной частицей 19S на этапе АТФ-зависимого связывания. [15] [28] Затем белок-субстрат должен войти внутрь субъединицы 20S, чтобы войти в контакт с протеолитическими активными центрами. Поскольку центральный канал частицы 20S узок и закрыт N-концевыми хвостами субъединиц α-кольца, субстраты должны быть хотя бы частично развернуты, прежде чем они войдут в ядро. [15] Переход развернутого субстрата в ядро называется транслокацией и обязательно происходит после деубиквитинирования. [15] [28] Однако порядок, в котором субстраты деубиквитинируются и разворачиваются, еще не ясен. [58] Какой из этих процессов является стадией, лимитирующей скорость общей реакции протеолиза, зависит от конкретного субстрата; для некоторых белков процесс разворачивания ограничивает скорость, тогда как деубиквитинирование является самым медленным этапом для других белков. [27] Согласно атомной структуре задействованной в субстрате 26S протеасомы в состоянии, совместимом с деубиквитилированием, степень, в которой субстраты должны быть развернуты перед транслокацией, составляет около 20 аминокислотных остатков. [15] но существенная третичная структура и, в частности, нелокальные взаимодействия, такие как дисульфидные связи , достаточны для ингибирования деградации. [59] Было также предложено, что наличие внутренне неупорядоченных белковых сегментов достаточного размера либо на конце белка, либо внутри него способствует эффективному инициированию деградации. [60] [61]
Ворота, образованные α-субъединицами, предотвращают проникновение пептидов длиной более четырех остатков внутрь 20S-частицы. Молекулы АТФ, связанные до начальной стадии узнавания, гидролизуются перед транслокацией. Хотя для разворачивания субстрата необходима энергия, для транслокации она не требуется. [27] [28] Собранная 26S протеасома может разрушать развернутые белки в присутствии негидролизуемого аналога АТФ , но не может разрушать свернутые белки, что указывает на то, что энергия гидролиза АТФ используется для разворачивания субстрата. [27] Прохождение развернутого субстрата через открытые ворота происходит посредством облегченной диффузии , если кэп 19S находится в АТФ-связанном состоянии. [62]
Механизм разворачивания глобулярных белков обязательно общий, но в некоторой степени зависит от аминокислотной последовательности . Было показано, что длинные последовательности чередующихся глицина и аланина ингибируют разворачивание субстрата, снижая эффективность протеасомной деградации; это приводит к высвобождению частично разложенных побочных продуктов, возможно, из-за разделения стадий гидролиза АТФ и разворачивания. [63] Такие глицин-аланиновые повторы встречаются и в природе, например, в шелка фиброине ; в частности, некоторые генные продукты вируса Эпштейна-Барра, несущие эту последовательность, могут останавливать протеасому, помогая вирусу размножаться, предотвращая презентацию антигена в главном комплексе гистосовместимости. [64]
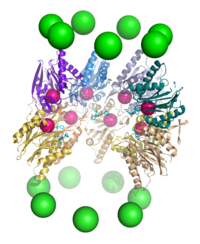
Протеолиз
[ редактировать ]Протеасома функционирует как эндопротеаза . [65] [66] [67] [68] Механизм протеолиза β-субъединицами коровой частицы 20S заключается в треонин-зависимой нуклеофильной атаке . Этот механизм может зависеть от связанной молекулы воды для депротонирования реакционноспособного гидроксила треонина . Деградация происходит внутри центральной камеры, образованной объединением двух β-колец, и обычно не высвобождает частично расщепленные продукты, вместо этого восстанавливая субстрат до коротких полипептидов, обычно длиной 7–9 остатков, хотя их длина может варьироваться от 4 до 25 остатков, в зависимости от организм и субстрат. Биохимический механизм, определяющий длину продукта, полностью не описан. [69] Хотя три каталитические β-субъединицы имеют общий механизм, они имеют несколько разные субстратные специфичности, которые считаются химотрипсин-подобными, трипсин-подобными и пептидил-глутамил-гидролизующими пептидами (PHGH). Эти изменения специфичности являются результатом межатомных контактов с локальными остатками вблизи активных центров каждой субъединицы. Каждая каталитическая β-субъединица также содержит консервативный остаток лизина, необходимый для протеолиза. [22]
Хотя протеасома обычно производит очень короткие пептидные фрагменты, в некоторых случаях эти продукты сами по себе являются биологически активными и функциональными молекулами. Некоторые факторы транскрипции, регулирующие экспрессию специфических генов, включая один компонент комплекса NF-κB млекопитающих , синтезируются в виде неактивных предшественников, убиквитинирование которых и последующая протеасомная деградация переводят их в активную форму. Такая активность требует, чтобы протеасома расщепляла белок-субстрат внутри, а не процессивно разрушала его с одного конца. Было высказано предположение, что длинные петли на поверхности этих белков служат субстратами протеасом и входят в центральную полость, тогда как большая часть белка остается снаружи. [70] Подобные эффекты наблюдались в дрожжевых белках; этот механизм избирательной деградации известен как регулируемый убиквитин/протеасомозависимый процессинг (RUP). [71]
Убиквитин-независимая деградация
[ редактировать ]Хотя большинство протеосомных субстратов должны быть убиквитинированы перед деградацией, существуют некоторые исключения из этого общего правила, особенно когда протеасома играет нормальную роль в посттрансляционном процессинге белка. Одним из основных примеров является протеасомная активация NF-κB путем процессинга p105 в p50 посредством внутреннего протеолиза. [70] Некоторые белки, которые предположительно нестабильны из-за внутренне неструктурированных областей, [72] деградируют убиквитин-независимым образом. Наиболее известным примером убиквитин-независимого протеасомного субстрата является фермент орнитиндекарбоксилаза . [73] о убиквитин-независимых механизмах, нацеленных на ключевые регуляторы клеточного цикла , такие как р53 , хотя р53 также подвержен убиквитин-зависимой деградации. Также сообщалось [74] Наконец, структурно аномальные, неправильно свернутые или сильно окисленные белки также подвержены убиквитин-независимой и 19S-независимой деградации в условиях клеточного стресса. [75]
Эволюция
[ редактировать ]
Протеасома 20S широко распространена и необходима эукариотам и археям. Бактериальный , чьи генные порядок Actinomycetales также имеет общие гомологи 20S протеасомы, тогда как большинство бактерий обладают теплового шока генами hslV и hslU продукты представляют собой мультимерную протеазу, расположенную в двухслойном кольце, и АТФазу. [76] Было высказано предположение, что белок hslV напоминает вероятного предка протеасомы 20S. [77] В целом, HslV не является существенным для бактерий, и не все бактерии им обладают, тогда как некоторые протисты обладают как 20S, так и системами hslV. [76] Многие бактерии также обладают другими гомологами протеасомы и связанной с ней АТФазы, особенно ClpP и ClpX . Эта избыточность объясняет, почему система HslUV не является необходимой.
Анализ последовательности показывает, что каталитические β-субъединицы разошлись в эволюции раньше, чем преимущественно структурные α-субъединицы. У бактерий, которые экспрессируют 20S протеасому, β-субъединицы имеют высокую идентичность последовательности с β-субъединицами архей и эукариот, тогда как идентичность последовательности α значительно ниже. Присутствие 20S протеасом у бактерий может быть результатом латерального переноса генов , тогда как диверсификация субъединиц среди эукариот приписывается множественным событиям дупликации генов . [76]
Контроль клеточного цикла
[ редактировать ]Развитие клеточного цикла контролируется упорядоченным действием циклин-зависимых киназ (CDK), активируемых специфическими циклинами , которые разграничивают фазы клеточного цикла . Митотические циклины, которые сохраняются в клетке всего несколько минут, имеют один из самых коротких периодов жизни среди всех внутриклеточных белков. [1] После того, как комплекс CDK-циклин выполнил свою функцию, связанный циклин полиубиквитинируется и разрушается протеасомой, что обеспечивает направленность клеточного цикла. В частности, выход из митоза требует протеасомо-зависимой диссоциации регуляторного компонента циклина B от комплекса факторов, способствующих митозу . [78] В клетках позвоночных «проскальзывание» через контрольную точку митоза, ведущее к преждевременному выходу из М-фазы , может происходить, несмотря на задержку этого выхода контрольной точкой веретена . [79]
Более ранние контрольные точки клеточного цикла, такие как проверка точки пострестрикции между G1 , убиквитинирование которого фазой и фазой S , аналогичным образом включают протеосомную деградацию циклина А обеспечивается комплексом, способствующим анафазе E3 (APC), убиквитинлигазой . [80] APC и белковый комплекс Skp1/Cul1/F-box ( комплекс SCF ) являются двумя ключевыми регуляторами деградации циклина и контроля контрольных точек; сам SCF регулируется APC посредством убиквитинирования адаптерного белка Skp2, который предотвращает активность SCF перед переходом G1-S. [81]
Отдельные компоненты частицы 19S выполняют свою регуляторную роль. Ганкирин , недавно идентифицированный онкопротеин , является одним из субкомпонентов 19S, который также прочно связывает циклин-зависимую киназу CDK4 и играет ключевую роль в распознавании убиквитинированного р53 благодаря его сродству к убиквитинлигазе MDM2 . Ганкирин является антиапоптотическим и , как было показано, сверхэкспрессируется в некоторых типах опухолевых клеток, таких как гепатоцеллюлярная карцинома . [82]
Подобно эукариотам, некоторые археи также используют протеасому для контроля клеточного цикла, в частности, путем контроля деления клеток, опосредованного ESCRT -III. [83]
Регулирование роста растений
[ редактировать ]У растений передача сигналов с помощью ауксинов или фитогормонов , которые определяют направление и тропизм роста растений, индуцирует нацеливание на класс репрессоров факторов транскрипции, известных как белки Aux/IAA, для протеасомной деградации. Эти белки убиквитинируются SCFTIR1 или SCF в комплексе с рецептором ауксина TIR1. Деградация белков Aux/IAA вызывает подавление транскрипционных факторов в семействе факторов ответа на ауксин (ARF) и индуцирует ARF-направленную экспрессию генов. [84] Клеточные последствия активации ARF зависят от типа растения и стадии развития, но участвуют в управлении ростом корней и жилок листьев. Считается, что специфический ответ на дерепрессию ОПН опосредован специфичностью спаривания отдельных белков АРФ и Aux/IAA. [85]
Апоптоз
[ редактировать ]Как внутренние, так и внешние сигналы могут привести к индукции апоптоза или запрограммированной гибели клеток. Результирующая деконструкция клеточных компонентов в основном осуществляется специализированными протеазами, известными как каспазы , но протеасома также играет важную и разнообразную роль в апоптотическом процессе. Об участии протеасомы в этом процессе свидетельствует как увеличение убиквитинирования белка, так и ферментов Е1, Е2 и Е3, которое наблюдается задолго до апоптоза. [86] [87] [88] Также наблюдалось, что во время апоптоза протеасомы, локализованные в ядре, перемещаются в пузырьки внешней мембраны , что характерно для апоптоза. [89]
Ингибирование протеасом по-разному влияет на индукцию апоптоза в разных типах клеток. В общем, протеасома не требуется для апоптоза, хотя ее ингибирование является проапоптотическим в большинстве изученных типов клеток. Апоптоз опосредован нарушением регулируемой деградации белков клеточного цикла, способствующих росту. [90] Однако некоторые клеточные линии, в частности первичные культуры покоящихся и дифференцированных клеток , таких как тимоциты и нейроны , не подвергаются апоптозу при воздействии ингибиторов протеасом. Механизм этого эффекта не ясен, но предполагается, что он специфичен для клеток в состоянии покоя или является результатом дифференциальной активности проапоптотической киназы JNK . [91] Способность ингибиторов протеасом индуцировать апоптоз в быстро делящихся клетках была использована в нескольких недавно разработанных химиотерапевтических агентах, таких как бортезомиб и салиноспорамид А.
Реакция на клеточный стресс
[ редактировать ]В ответ на клеточные стрессы, такие как инфекция , тепловой шок или окислительное повреждение , белки теплового шока экспрессируются , которые идентифицируют неправильно свернутые или развернутые белки и нацеливают их на протеосомную деградацию. И Hsp27 , и Hsp90 — белки- шапероны участвуют в повышении активности убиквитин-протеасомной системы, хотя они не являются непосредственными участниками этого процесса. [92] Hsp70 , с другой стороны, связывает открытые гидрофобные участки на поверхности неправильно свернутых белков и привлекает E3 убиквитинлигазы, такие как CHIP, для маркировки белков для протеасомной деградации. [93] Белок CHIP (карбоксильный конец белка, взаимодействующего с Hsp70) сам по себе регулируется посредством ингибирования взаимодействий между ферментом E3 CHIP и его партнером по связыванию E2. [94]
Подобные механизмы существуют, чтобы способствовать деградации окислительно-поврежденных белков через систему протеасом. В частности, протеасомы, локализованные в ядре, регулируются PARP и активно деградируют ненадлежащим образом окисленные гистоны . [95] Окисленные белки, которые часто образуют большие аморфные агрегаты в клетке, могут разлагаться непосредственно под действием коровой частицы 20S без регуляторного колпачка 19S и не требуют гидролиза АТФ или мечения убиквитином. [75] Однако высокий уровень окислительного повреждения увеличивает степень перекрестных связей между фрагментами белка, делая агрегаты устойчивыми к протеолизу. Большие количества и размеры таких сильно окисленных агрегатов связаны со старением . [96]
Нарушение регуляции протеасомной системы убиквитина может способствовать развитию ряда нервных заболеваний. Это может привести к опухолям головного мозга, таким как астроцитомы . [97] При некоторых нейродегенеративных заболеваниях с поздним началом, общим признаком которых является агрегация неправильно свернутых белков, таких как болезнь Паркинсона и болезнь Альцгеймера , могут образовываться большие нерастворимые агрегаты неправильно свернутых белков, которые затем приводят к нейротоксичности посредством механизмов, которые еще не до конца изучены. . Снижение активности протеасом было предложено как причина агрегации и образования телец Леви при болезни Паркинсона. [98] Эта гипотеза подтверждается наблюдением, что дрожжевые модели болезни Паркинсона более восприимчивы к токсичности α-синуклеина , основного белкового компонента телец Леви, в условиях низкой активности протеасом. [99] Нарушение протеасомальной активности может лежать в основе когнитивных расстройств, таких как расстройства аутистического спектра , а также заболеваний мышц и нервов, таких как миопатия с включениями . [97]
Роль в иммунной системе
[ редактировать ]Протеасома играет простую, но важную роль в функционировании адаптивной иммунной системы . Пептидные антигены экспонируются белками главного комплекса гистосовместимости класса I (MHC) на поверхности антигенпрезентирующих клеток . Эти пептиды являются продуктами протеасомной деградации белков, образующихся вторгшимся патогеном . Хотя в этом процессе могут участвовать конститутивно экспрессируемые протеасомы, специализированный комплекс, состоящий из белков, экспрессия которых индуцируется гамма-интерфероном , является первичным продуцентом пептидов, оптимальных по размеру и составу для связывания MHC. Эти белки, экспрессия которых увеличивается во время иммунного ответа, включают регуляторную частицу 11S, основная известная биологическая роль которой заключается в регулировании продукции лигандов MHC, а также специализированные β-субъединицы, называемые β1i, β2i и β5i, с измененной субстратной специфичностью. Комплекс, образованный специализированными β-субъединицами, известен как иммунопротеасома . [16] Другая субъединица варианта β5i, β5t, экспрессируется в тимусе, что приводит к образованию специфичной для тимуса « тимопротеасомы », функция которой пока неясна. [100]
Сила связывания лиганда MHC класса I зависит от состава С-конца лиганда , поскольку пептиды связываются за счет водородных связей и тесных контактов с областью, называемой «карманом B» на поверхности MHC. Многие аллели MHC класса I предпочитают гидрофобные С-концевые остатки, а иммунопротеасомный комплекс с большей вероятностью генерирует гидрофобные С-концевые остатки. [101]
Благодаря своей роли в генерации активированной формы NF-κB , антиапоптотического и провоспалительного регулятора экспрессии цитокинов , активность протеасом связана с воспалительными и аутоиммунными заболеваниями . Повышенные уровни активности протеасом коррелируют с активностью заболевания и связаны с аутоиммунными заболеваниями, включая системную красную волчанку и ревматоидный артрит . [16]
Протеасома также участвует во внутриклеточном антитело-опосредованном протеолизе связанных с антителами вирионов. В этом пути нейтрализации TRIM21 (белок семейства трехчастных мотивов) связывается с иммуноглобулином G , направляя вирион к протеасоме, где он разрушается. [102]
Ингибиторы протеасом
[ редактировать ]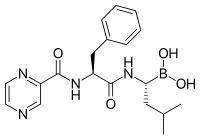
Ингибиторы протеасом обладают эффективной противоопухолевой активностью в культуре клеток , индуцируя апоптоз , нарушая регулируемую деградацию белков клеточного цикла, способствующих росту. [90] Этот подход избирательной индукции апоптоза в опухолевых клетках доказал свою эффективность на животных моделях и в испытаниях на людях.
Лактацистин , природный продукт, синтезируемый Streptomyces бактериями , был первым обнаруженным непептидным ингибитором протеасом. [103] и широко используется в качестве исследовательского инструмента в биохимии и клеточной биологии. Лицензия на лактацистин была передана компании Myogenics/Proscript, которая была приобретена компанией Millennium Pharmaceuticals , которая теперь является частью Takeda Pharmaceuticals . Лактацистин ковалентно модифицирует аминоконцевой треонин каталитических β-субъединиц протеасомы, особенно субъединицы β5, ответственной за химотрипсиноподобную активность протеасомы. Это открытие помогло утвердить протеасому как механически новый класс протеаз: аминоконцевую треониновую протеазу .
Бортезомиб (боронатный MG132), молекула, разработанная Millennium Pharmaceuticals и продаваемая под торговой маркой Velcade, является первым ингибитором протеасом, получившим клиническое применение в качестве химиотерапевтического агента. [104] Бортезомиб применяется при лечении множественной миеломы . [105] Примечательно, что множественная миелома приводит к повышению уровня пептидов, полученных из протеасом, в сыворотке крови , которые снижаются до нормальных уровней в ответ на успешную химиотерапию. [106] Исследования на животных показали, что бортезомиб может также оказывать клинически значимое воздействие при раке поджелудочной железы . [107] [108] Начаты доклинические и ранние клинические исследования для изучения эффективности бортезомиба в лечении других видов рака, связанных с В-клетками . [109] особенно некоторые типы неходжкинской лимфомы . [110] Клинические результаты также, по-видимому, оправдывают использование ингибитора протеасом в сочетании с химиотерапией при остром В-клеточном лимфобластном лейкозе. [111] Ингибиторы протеасом могут уничтожать некоторые типы культивируемых лейкозных клеток, устойчивых к глюкокортикоидам. [112]
Молекула ритонавир , продаваемая как Норвир, была разработана как ингибитор протеазы и использовалась для борьбы с ВИЧ- инфекцией. Однако было показано, что он ингибирует протеасомы, а также свободные протеазы; конкретно, ритонавир ингибирует химотрипсин -подобную активность протеасомы, тогда как трипсин -подобная активность несколько усиливается. [113] Исследования на животных моделях показывают, что ритонавир может оказывать ингибирующее действие на рост клеток глиомы . [114]
Ингибиторы протеасом также показали многообещающую эффективность в лечении аутоиммунных заболеваний на животных моделях. Например, исследования на мышах с трансплантатами человеческой кожи обнаружили уменьшение размера поражений псориаза после лечения ингибитором протеасом. [115] Ингибиторы также показывают положительные эффекты на на грызунах моделях астмы . [116]
Мечение и ингибирование протеасомы также представляют интерес в лабораторных условиях для как in vitro , так и in vivo изучения протеасомной активности в клетках . Наиболее часто используемыми лабораторными ингибиторами являются лактацистин и пептидный альдегид MG132, первоначально разработанный лабораторией Голдберга. флуоресцентные ингибиторы для специфической маркировки активных участков собранной протеасомы. Также были разработаны [117]
Клиническое значение
[ редактировать ]Протеасома и ее субъединицы имеют клиническое значение, по крайней мере, по двум причинам: (1) нарушенная комплексная сборка или дисфункциональная протеасома могут быть связаны с основной патофизиологией конкретных заболеваний и (2) они могут использоваться в качестве мишеней для лекарственных препаратов для терапевтических целей. вмешательства. Совсем недавно были предприняты дополнительные усилия по рассмотрению протеасом для разработки новых диагностических маркеров и стратегий. Улучшенное и всестороннее понимание патофизиологии протеасом должно привести к их клиническому применению в будущем.
Протеасомы образуют ключевой компонент системы убиквитин-протеасома (UPS). [118] и соответствующий контроль качества клеточного белка (PQC). белка Убиквитинирование и последующий протеолиз и деградация протеасомой являются важными механизмами регуляции клеточного цикла , роста и дифференцировки клеток , транскрипции генов, передачи сигнала и апоптоза . [119] Дефекты протеасом приводят к снижению протеолитической активности и накоплению поврежденных или неправильно свернутых белков, что может способствовать нейродегенеративным заболеваниям. [120] [121] сердечно-сосудистые заболевания, [122] [123] [124] воспалительные реакции и аутоиммунные заболевания, [125] и системные реакции на повреждение ДНК, приводящие к злокачественным новообразованиям . [126]
Исследования показали, что дефекты ИБС участвуют в патогенезе нейродегенеративных и миодегенеративных заболеваний, включая болезнь Альцгеймера . [127] болезнь Паркинсона [128] и болезнь Пика , [129] боковой амиотрофический склероз (АЛС), [129] болезнь Гентингтона , [128] болезнь Крейтцфельдта-Якоба , [130] и двигательных нейронов, полиглутаминовые (PolyQ) болезни, мышечные дистрофии [131] и несколько редких форм нейродегенеративных заболеваний, связанных с деменцией . [132] Как часть убиквитин-протеасомной системы (UPS), протеасома поддерживает гомеостаз сердечных белков и, таким образом, играет значительную роль в ишемическом повреждении сердца. [133] гипертрофия желудочков [134] и сердечная недостаточность . [135] Кроме того, накапливаются данные о том, что ИБС играет важную роль в злокачественной трансформации. Протеолиз UPS играет важную роль в ответах раковых клеток на стимулирующие сигналы, которые имеют решающее значение для развития рака. Соответственно, экспрессия генов путем деградации факторов транскрипции , таких как p53 , c-jun , c-Fos , NF-κB , c-Myc , HIF-1α, MATα2, STAT3 , белки, связывающие элементы, регулируемые стеролом, и рецепторы андрогенов являются контролируется UPS и, таким образом, участвует в развитии различных злокачественных новообразований. [136] Более того, UPS регулирует деградацию продуктов генов-супрессоров опухолей, таких как аденоматозный полипоз кишечной палочки (APC) при колоректальном раке, ретинобластоме (Rb). и супрессор опухоли фон Хиппель-Линдау (VHL), а также ряд протоонкогенов ( Raf , Myc , Myb , Rel , Src , Mos , ABL ). ИБП также участвует в регуляции воспалительных реакций. Эту активность обычно связывают с ролью протеасом в активации NF-κB, который дополнительно регулирует экспрессию провоспалительных цитокинов, таких как TNF-α , IL-β, IL-8 , молекулы адгезии ( ICAM-1 , VCAM-1). , Р-селектин ) и простагландины и оксид азота (NO). [125] Кроме того, УПС также играет роль в воспалительных реакциях в качестве регуляторов пролиферации лейкоцитов, главным образом за счет протеолиза циклинов и деградации ингибиторов CDK . [137] Наконец, аутоиммунными заболеваниями, у пациентов с такими как СКВ , синдром Шегрена и ревматоидный артрит (РА), преимущественно обнаруживаются циркулирующие протеасомы, которые можно применять в качестве клинических биомаркеров. [138]
См. также
[ редактировать ]- Карта протеолиза
- Семейство белков DSS1/SEM1
- Экзосомный комплекс
- Деградация белков, связанных с эндоплазматической сетью
- JUNQ и IPOD
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Лодиш Х., Берк А., Мацудайра П., Кайзер К.А., Кригер М., Скотт М.П., Зипурски С.Л., Дарнелл Дж. (2004). «3» . Молекулярно-клеточная биология (5-е изд.). Нью-Йорк: WH Freeman and CO., стр. 66–72 . ISBN 978-0-7167-4366-8 .
- ^ Петерс Дж. М., Франке В. В., Кляйншмидт Дж. А. (март 1994 г.). «Отличные 19 S и 20 S субкомплексы 26 S протеасомы и их распределение в ядре и цитоплазме» . Журнал биологической химии . 269 (10): 7709–18. дои : 10.1016/S0021-9258(17)37345-3 . ПМИД 8125997 .
- ^ Нассиф, Николас Д.; Камбрей, Саманта Э.; Краут, Дэниел А. (май 2014 г.). «Проскальзывание: частичная деградация субстрата АТФ-зависимыми протеазами» . ИУБМБ Жизнь . 66 (5): 309–317. дои : 10.1002/iub.1271 . ПМИД 24823973 . S2CID 29860298 .
- ^ Jump up to: а б Нобелевский комитет (2004). «Лауреаты Нобелевской премии по химии 2004 г.» . Проверено 11 декабря 2006 г.
- ^ Этлингер Дж. Д., Голдберг А. Л. (январь 1977 г.). «Растворимая АТФ-зависимая протеолитическая система, ответственная за деградацию аномальных белков в ретикулоцитах» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (1): 54–8. Бибкод : 1977PNAS...74...54E . дои : 10.1073/pnas.74.1.54 . ПМЦ 393195 . ПМИД 264694 .
- ^ Цехановер А., Ход Ю., Гершко А. (апрель 1978 г.). «Термостабильный полипептидный компонент АТФ-зависимой протеолитической системы ретикулоцитов». Связь с биохимическими и биофизическими исследованиями . 81 (4): 1100–5. дои : 10.1016/0006-291X(78)91249-4 . ПМИД 666810 .
- ^ Гольдкнопф И.Л., Буш Х. (март 1977 г.). «Изопептидная связь между негистоновыми и гистоновыми полипептидами 2А хромосомного конъюгата-белка А24» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (3): 864–8. Бибкод : 1977PNAS...74..864G . дои : 10.1073/pnas.74.3.864 . ПМК 430507 . ПМИД 265581 .
- ^ Чехановер А. (сентябрь 2005 г.). «Ранние работы над протеасомной системой убиквитина, интервью с Аароном Чехановером. Интервью CDD» . Смерть клеток и дифференцировка . 12 (9): 1167–77. дои : 10.1038/sj.cdd.4401691 . ПМИД 16094393 .
- ^ Уилк С., Орловски М. (ноябрь 1980 г.). «Катион-чувствительная нейтральная эндопептидаза: выделение и специфичность фермента бычьего гипофиза». Журнал нейрохимии . 35 (5): 1172–82. дои : 10.1111/j.1471-4159.1980.tb07873.x . ПМИД 6778972 . S2CID 9028201 .
- ^ Арриго А.П., Танака К., Голдберг Ф., Уэлч В.Дж. (1988). «Идентичность просомной частицы 19S с большим многофункциональным протеазным комплексом клеток млекопитающих». Природа . 331 (6152): 192–94. дои : 10.1038/331192a0 . ПМИД 3277060 . S2CID 97688 . Танака К., Ваксман Л., Голдберг А.Л. (июнь 1983 г.). «АТФ выполняет две различные роли в деградации белка в ретикулоцитах: одна требует, а другая не зависит от убиквитина» . Журнал клеточной биологии . 96 (6): 1580–5. дои : 10.1083/jcb.96.6.1580 . ПМК 2112434 . ПМИД 6304111 .
- ^ Хаф Р., Пратт Г., Рехштайнер М. (июнь 1987 г.). «Очистка двух высокомолекулярных протеаз из лизата ретикулоцитов кролика» . Журнал биологической химии . 262 (17): 8303–13. дои : 10.1016/S0021-9258(18)47564-3 . ПМИД 3298229 .
- ^ Гершко А (сентябрь 2005 г.). «Ранние работы над убиквитиновой протеасомной системой, интервью с Аврамом Гершко. Интервью CDD» . Смерть клеток и дифференцировка . 12 (9): 1158–61. дои : 10.1038/sj.cdd.4401709 . ПМИД 16094391 .
- ^ Копп Ф., Штайнер Р., Дальманн Б., Кюн Л., Рейнауэр Х. (август 1986 г.). «Размер и форма мультикаталитической протеиназы скелетных мышц крысы». Biochimica et Biophysical Acta (BBA) - Структура белка и молекулярная энзимология . 872 (3): 253–60. дои : 10.1016/0167-4838(86)90278-5 . ПМИД 3524688 .
- ^ Лёве Дж., Сток Д., Яп Б., Цвикль П., Баумайстер В., Хубер Р. (апрель 1995 г.). «Кристаллическая структура протеасомы 20S археи T. acidophilum при разрешении 3,4 А». Наука . 268 (5210): 533–9. Бибкод : 1995Sci...268..533L . дои : 10.1126/science.7725097 . ПМИД 7725097 .
- ^ Jump up to: а б с д и ж г час я дж к л Донг Й, Чжан С, Ву З, Ли Х, Ван ВЛ, Чжу Й, Стойлова-Макфи С, Лу Й, Финли Д, Мао Й (ноябрь 2018 г.). «Крио-ЭМ структуры и динамика протеасомы 26S человека, связанной с субстратом» . Природа . 565 (7737): 49–55. дои : 10.1038/s41586-018-0736-4 . ПМК 6370054 . ПМИД 30479383 .
- ^ Jump up to: а б с д Ван Дж., Мальдонадо, Массачусетс (август 2006 г.). «Убиквитин-протеасомная система и ее роль при воспалительных и аутоиммунных заболеваниях». Клеточная и молекулярная иммунология . 3 (4): 255–61. ПМИД 16978533 .
- ^ Jump up to: а б с Штадтмюллер, Б.М.; Хилл, CP (7 января 2011 г.). «Протеасомные активаторы» . Молекулярная клетка . 41 (1): 8–19. doi : 10.1016/j.molcel.2010.12.020 . ПМК 3040445 . ПМИД 21211719 .
- ^ Jump up to: а б с Смит Д.М., Чанг С.К., Парк С., Финли Д., Ченг Ю., Голдберг А.Л. (сентябрь 2007 г.). «Стыковка карбоксильных концов протеасомных АТФаз с альфа-кольцом 20S протеасомы открывает ворота для входа субстрата» . Молекулярная клетка . 27 (5): 731–44. doi : 10.1016/j.molcel.2007.06.033 . ПМК 2083707 . ПМИД 17803938 .
- ^ «Семья МЕРОПС Т1» . ЭМБЛ-ЭБИ . Проверено 16 февраля 2019 г.
- ^ Уилк С., Орловски М. (март 1983 г.). «Доказательства того, что катион-чувствительная нейтральная эндопептидаза гипофиза представляет собой мультикаталитический протеазный комплекс». Журнал нейрохимии . 40 (3): 842–9. дои : 10.1111/j.1471-4159.1983.tb08056.x . ПМИД 6338156 . S2CID 23508675 .
- ^ Jump up to: а б Нанди Д., Тахилиани П., Кумар А., Чанду Д. (март 2006 г.). «Система убиквитин-протеасома» (PDF) . Журнал биологических наук . 31 (1): 137–55. дои : 10.1007/BF02705243 . ПМИД 16595883 . S2CID 21603835 .
- ^ Jump up to: а б Хайнемейер В., Фишер М., Криммер Т., Стахон У., Вольф Д.Х. (октябрь 1997 г.). «Активные сайты эукариотической 20 S протеасомы и их участие в процессинге предшественников субъединиц» . Журнал биологической химии . 272 (40): 25200–9. дои : 10.1074/jbc.272.40.25200 . ПМИД 9312134 .
- ^ Jump up to: а б Падманабхан А., Вуонг С.А., Хохштрассер М. (март 2016 г.). «Сборка эволюционно консервативной альтернативной изоформы протеасомы в клетках человека» . Отчеты по ячейкам . 14 (12): 2962–74. дои : 10.1016/j.celrep.2016.02.068 . ПМЦ 4828729 . ПМИД 26997268 .
- ^ Величутина И., Коннерли П.Л., Арендт К.С., Ли Х, Хохштрассер М. (февраль 2004 г.). «Пластичность сборки кольца 20S протеасомы эукариот, выявленная путем делеции субъединицы у дрожжей» . Журнал ЭМБО . 23 (3): 500–10. дои : 10.1038/sj.emboj.7600059 . ПМЦ 1271798 . ПМИД 14739934 .
- ^ Кусмерчик А.Р., Кунджаппу М.Ю., Фунакоси М., Хохштрассер М. (март 2008 г.). «Мультимерный фактор сборки контролирует образование альтернативных 20S протеасом». Структурная и молекулярная биология природы . 15 (3): 237–44. дои : 10.1038/nsmb.1389 . ПМИД 18278055 . S2CID 21181637 .
- ^ Цвикл П., Нг Д., Ву К.М., Кленк Х.П., Голдберг А.Л. (сентябрь 1999 г.). «Архебактериальная АТФаза, гомологичная АТФазам в 26S-протеасоме эукариот, активирует расщепление белка 20S-протеасомами» . Журнал биологической химии . 274 (37): 26008–14. дои : 10.1074/jbc.274.37.26008 . ПМИД 10473546 .
- ^ Jump up to: а б с д Смит Д.М., Кафри Г., Ченг Ю., Нг Д., Уолц Т., Голдберг А.Л. (декабрь 2005 г.). «Связывание АТФ с PAN или 26S-АТФазами вызывает ассоциацию с 20S-протеасомой, открытие ворот и транслокацию развернутых белков» . Молекулярная клетка . 20 (5): 687–98. doi : 10.1016/j.molcel.2005.10.019 . ПМИД 16337593 .
- ^ Jump up to: а б с д и Лю К.В., Ли Х, Томпсон Д., Вудинг К., Чанг Т.Л., Тан З., Ю Х., Томас П.Дж., ДеМартино Г.Н. (октябрь 2006 г.). «Связывание АТФ и гидролиз АТФ играют разные роли в функции 26S протеасомы» . Молекулярная клетка . 24 (1): 39–50. doi : 10.1016/j.molcel.2006.08.025 . ПМЦ 3951175 . ПМИД 17018291 .
- ^ Лам Ю.А., Лоусон Т.Г., Велаютам М., Цвайер Дж.Л., Пиккарт К.М. (апрель 2002 г.). «Протеасомная субъединица АТФазы распознает сигнал деградации полиубиквитина». Природа . 416 (6882): 763–7. Бибкод : 2002Natur.416..763L . дои : 10.1038/416763а . ПМИД 11961560 . S2CID 4421764 .
- ^ Jump up to: а б Бек Ф., Унвердорбен П., Бон С., Швейцер А., Пфайфер Г., Саката Е., Никелл С., Плитцко Дж.М., Вилла Е., Баумайстер В., Фёрстер Ф. (сентябрь 2012 г.). «Структурная модель 26S протеасомы дрожжей с почти атомным разрешением» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (37): 14870–5. Бибкод : 2012PNAS..10914870B . дои : 10.1073/pnas.1213333109 . ПМЦ 3443124 . ПМИД 22927375 .
- ^ Jump up to: а б Ландер Г.К., Эстрин Э., Матыскела М.Е., Башор С., Ногалес Е., Мартин А. (февраль 2012 г.). «Полная субъединичная архитектура регуляторной частицы протеасомы» . Природа . 482 (7384): 186–91. Бибкод : 2012Natur.482..186L . дои : 10.1038/nature10774 . ПМЦ 3285539 . ПМИД 22237024 .
- ^ Ласкер К., Фёрстер Ф., Бон С., Вальцтхени Т., Вилла Е, Унвердорбен П., Бек Ф., Эберсолд Р., Сали А., Баумейстер В. (январь 2012 г.). «Молекулярная архитектура протеасомного голокомплекса 26S, определяемая интегративным подходом» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (5): 1380–7. дои : 10.1073/pnas.1120559109 . ПМК 3277140 . ПМИД 22307589 .
- ^ Jump up to: а б с Чен С., Ву Дж, Лу Й, Ма ЙБ, Ли БХ, Ю З, Оуян Кью, Финли DJ, Киршнер М.В., Мао Й (ноябрь 2016 г.). «Структурная основа динамической регуляции протеасомы 26S человека» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (46): 12991–12996. Бибкод : 2016PNAS..11312991C . дои : 10.1073/pnas.1614614113 . ПМЦ 5135334 . ПМИД 27791164 .
- ^ Хуан X, Луань Б, Ву Дж, Ши Ю (сентябрь 2016 г.). «Атомная структура протеасомы 26S человека». Структурная и молекулярная биология природы . 23 (9): 778–785. дои : 10.1038/nsmb.3273 . ПМИД 27428775 . S2CID 21909333 .
- ^ Швейцер А., Ауфдерхайде А., Рудак Т., Бек Ф., Пфайфер Г., Плитцко Дж. М., Саката Е., Шультен К., Фёрстер Ф., Баумейстер В. (июль 2016 г.). «Структура протеасомы 26S человека с разрешением 3,9 Å» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (28): 7816–7821. Бибкод : 2016PNAS..11312991C . дои : 10.1073/pnas.1614614113 . ПМЦ 5135334 . ПМИД 27791164 .
- ^ Jump up to: а б с д Чжу Ю, Ван ВЛ, Ю Д, Оуян Ц, Лу Ю, Мао Ю (апрель 2018 г.). «Структурный механизм нуклеотидного ремоделирования анфолдазы AAA-АТФазы в активированной 26S протеасоме человека» . Природные коммуникации . 9 (1): 1360. Бибкод : 2018NatCo...9.1360Z . doi : 10.1038/s41467-018-03785-w . ПМЦ 5893597 . ПМИД 29636472 .
- ^ Jump up to: а б с Унвердорбен П., Бек Ф., Следьц П., Швейцер А., Пфайфер Г., Плитцко Ю.М., Баумайстер В., Фёрстер Ф. (апрель 2014 г.). «Глубокая классификация большого набора крио-ЭМ данных определяет конформационный ландшафт протеасомы 26S» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (15): 5544–9. Бибкод : 2014PNAS..111.5544U . дои : 10.1073/pnas.1403409111 . ПМЦ 3992697 . ПМИД 24706844 .
- ^ Следз П., Унвердорбен П., Бек Ф., Пфайфер Г., Швейцер А., Фёрстер Ф., Баумайстер В. (апрель 2013 г.). «Структура 26S протеасомы, связанной с АТФ-γS, дает представление о механизме нуклеотид-зависимой транслокации субстрата» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (18): 7264–7269. Бибкод : 2013PNAS..110.7264S . дои : 10.1073/pnas.1305782110 . ПМЦ 3645540 . ПМИД 23589842 .
- ^ Матыскела М.Е., Ландер Г.К., Мартин А. (июль 2013 г.). «Конформационное переключение протеасомы 26S обеспечивает деградацию субстрата» . Структурная и молекулярная биология природы . 20 (7): 781–788. дои : 10.1038/nsmb.2616 . ПМЦ 3712289 . ПМИД 23770819 .
- ^ Лу Й, Ву Дж, Донг Й, Чен С, Сунь С, Ма ЙБ, Оуян Ц, Финли Д., Киршнер М.В., Мао Й (июль 2017 г.). «Конформационный ландшафт регуляторной частицы протеасомы человека, связанной с p28» . Молекулярная клетка . 67 (2): 322–333.e6. doi : 10.1016/j.molcel.2017.06.007 . ПМК 5580496 . ПМИД 28689658 .
- ^ Кёлер А., Касио П., Леггетт Д.С., Ву К.М., Голдберг А.Л., Финли Д. (июнь 2001 г.). «Аксиальный канал коровой частицы протеасомы закрывается АТФазой Rpt2 и контролирует как вход субстрата, так и высвобождение продукта» . Молекулярная клетка . 7 (6): 1143–52. дои : 10.1016/S1097-2765(01)00274-X . ПМИД 11430818 .
- ^ Фёрстер А., Мастерс Э.И., Уитби Ф.Г., Робинсон Х., Хилл С.П. (май 2005 г.). «Структура 1.9 A активаторного комплекса протеасома-11S и последствия для взаимодействий протеасома-PAN/PA700» . Молекулярная клетка . 18 (5): 589–99. doi : 10.1016/j.molcel.2005.04.016 . ПМИД 15916965 .
- ^ Витт С., Квон Ю.Д., Шэрон М., Фельдерер К., Беттлер М., Робинсон К.В., Баумайстер В., Япончик Б.К. (июль 2006 г.). «Сборка протеасом запускает переключатель, необходимый для созревания активного сайта» . Структура . 14 (7): 1179–88. дои : 10.1016/j.str.2006.05.019 . ПМИД 16843899 .
- ^ Крюгер Э., Клёцель П.М., Эненкель С. (2001). «Биогенез 20S протеасом». Биохимия . 83 (3–4): 289–93. дои : 10.1016/S0300-9084(01)01241-X . ПМИД 11295488 .
- ^ Мурата С., Яширода Х., Танака К. (февраль 2009 г.). «Молекулярные механизмы сборки протеасом». Nature Reviews Молекулярно-клеточная биология . 10 (2): 104–115. дои : 10.1038/nrm2630 . ПМИД 19165213 . S2CID 21263837 .
- ^ Саката Э., Стенгель Ф., Фукунага К., Чжоу М., Саэки Ю., Фёрстер Ф., Баумайстер В., Танака К., Робинсон К.В. (июнь 2011 г.). «Каталитическая активность Ubp6 усиливает созревание протеасомальных регуляторных частиц» . Молекулярная клетка . 42 (5): 637–649. doi : 10.1016/j.molcel.2011.04.021 . ПМИД 21658604 .
- ^ Фукунага К., Кудо Т., Тохе А., Танака К., Саэки Ю. (июнь 2010 г.). «Рассечение пути сборки крышки протеасомы у Saccharomyces cerevisiae». Связь с биохимическими и биофизическими исследованиями . 396 (4): 1048–1053. дои : 10.1016/j.bbrc.2010.05.061 . ПМИД 20471955 .
- ^ Хаас А.Л., Вармс СП, Гершко А., Роуз И.А. (март 1982 г.). «Фермент, активирующий убиквитин. Механизм и роль в конъюгации белок-убиквитин» . Журнал биологической химии . 257 (5): 2543–8. дои : 10.1016/S0021-9258(18)34958-5 . ПМИД 6277905 .
- ^ Троуэр Дж.С., Хоффман Л., Рехштайнер М., Пиккарт К.М. (январь 2000 г.). «Распознавание протеолитического сигнала полиубиквитина» . Журнал ЭМБО . 19 (1): 94–102. дои : 10.1093/emboj/19.1.94 . ПМЦ 1171781 . ПМИД 10619848 .
- ^ Риссеев Е.П., Даскальчук Т.Е., Бэнкс Т.В., Лю Э., Котелесаж Дж., Хеллманн Х., Эстель М., Сомерс Д.Э., Кросби В.Л. (июнь 2003 г.). «Анализ белкового взаимодействия субъединиц убиквитин E3-лигазы SCF из Arabidopsis» . Заводской журнал . 34 (6): 753–67. дои : 10.1046/j.1365-313X.2003.01768.x . ПМИД 12795696 .
- ^ Эльзассер С., Финли Д. (август 2005 г.). «Доставка убиквитинированных субстратов в машины для разворачивания белков». Природная клеточная биология . 7 (8): 742–9. дои : 10.1038/ncb0805-742 . ПМИД 16056265 . S2CID 21069699 .
- ^ Саданандом А., Бейли М., Юэн Р., Ли Дж., Нелис С. (октябрь 2012 г.). «Убиквитин-протеасомная система: центральный модификатор передачи сигналов растений» . Новый фитолог . 196 (1): 13–28. дои : 10.1111/j.1469-8137.2012.04266.x . ПМИД 22897362 .
- ^ Sharp PM, Ли WH (1987). «Гены убиквитина как парадигма согласованной эволюции тандемных повторов». Журнал молекулярной эволюции . 25 (1): 58–64. Бибкод : 1987JMolE..25...58S . дои : 10.1007/BF02100041 . ПМИД 3041010 . S2CID 7929162 .
- ^ Пиккарт CM, Фушман Д. (декабрь 2004 г.). «Полюбиквитиновые цепи: сигналы полимерных белков». Современное мнение в области химической биологии . 8 (6): 610–16. дои : 10.1016/j.cbpa.2004.09.009 . ПМИД 15556404 .
- ^ Сюй П., Дуонг Д.М., Сейфрид Н.Т., Ченг Д., Се Ю, Роберт Дж., Раш Дж., Хохштрассер М., Финли Д., Пэн Дж. (апрель 2009 г.). «Количественная протеомика раскрывает функцию нетрадиционных цепей убиквитина в протеасомной деградации» . Клетка . 137 (1): 133–45. дои : 10.1016/j.cell.2009.01.041 . ПМК 2668214 . ПМИД 19345192 .
- ^ Jump up to: а б Пиккарт CM (ноябрь 2000 г.). «Убиквитин в цепочках». Тенденции биохимических наук . 25 (11): 544–8. дои : 10.1016/S0968-0004(00)01681-9 . ПМИД 11084366 .
- ^ Jump up to: а б Чжан С., Цзоу С., Инь Д., Чжао Л., Финли Д., Ву Цзи, Мао Юй (апрель 2022 г.). «Регулируемая USP14 аллостерия протеасомы человека с помощью крио-ЭМ с временным разрешением» . Природа . 605 (7910): 567–574. Бибкод : 2022Natur.605..567Z . дои : 10.1038/s41586-022-04671-8 . ПМЦ 9117149 . ПМИД 35477760 .
- ^ Чжу Ц, Вани Дж., Ван Ц.Э., Эль-Махди М., Снапка Р.М., Вани А.А. (июль 2005 г.). «Дубиквитинирование протеасомой координируется с транслокацией субстрата для протеолиза in vivo». Экспериментальные исследования клеток . 307 (2): 436–51. doi : 10.1016/j.yexcr.2005.03.031 . ПМИД 15950624 .
- ^ Венцель Т., Баумайстер В. (март 1995 г.). «Конформационные ограничения деградации белка протеасомой 20S». Структурная биология природы . 2 (3): 199–204. дои : 10.1038/nsb0395-199 . ПМИД 7773788 . S2CID 41599619 .
- ^ Инобе Т., Фишбейн С., Пракаш С., Матушек А. (март 2011 г.). «Определение геометрии двухкомпонентного протеасомного дегрона» . Химическая биология природы . 7 (3): 161–7. дои : 10.1038/nchembio.521 . ПМК 3129032 . ПМИД 21278740 .
- ^ ван дер Ли Р., Ланг Б., Крузе К., Гспонер Дж., Санчес де Гроот Н., Хюйнен М.А., Матушек А., Фуксрайтер М., Бабу М.М. (сентябрь 2014 г.). «Внутренне неупорядоченные сегменты влияют на период полураспада белка в клетке и в ходе эволюции» . Отчеты по ячейкам . 8 (6): 1832–44. дои : 10.1016/j.celrep.2014.07.055 . ПМЦ 4358326 . ПМИД 25220455 .
- ^ Смит Д.М., Бенарудж Н., Голдберг А. (октябрь 2006 г.). «Протеасомы и связанные с ними АТФазы: разрушительная комбинация». Журнал структурной биологии . 156 (1): 72–83. дои : 10.1016/j.jsb.2006.04.012 . ПМИД 16919475 .
- ^ Хойт М.А., Зич Дж., Такеучи Дж., Чжан М., Говертс С., Коффино П. (апрель 2006 г.). «Глицин-аланиновые повторы нарушают правильное развертывание субстрата протеасомой» . Журнал ЭМБО . 25 (8): 1720–9. дои : 10.1038/sj.emboj.7601058 . ПМК 1440830 . ПМИД 16601692 .
- ^ Чжан М., Коффино П. (март 2004 г.). «Повторяющаяся последовательность белка ядерного антигена 1, кодируемого вирусом Эпштейна-Барра, прерывает обработку субстрата протеасом» . Журнал биологической химии . 279 (10): 8635–41. дои : 10.1074/jbc.M310449200 . ПМИД 14688254 .
- ^ Зеемюллер Э., Лупас А., Сток Д., Лёве Дж., Хубер Р., Баумайстер В. (апрель 1995 г.). «Протеасома из Thermoplasma acidophilum: треониновая протеаза». Наука . 268 (5210): 579–82. Бибкод : 1995Sci...268..579S . дои : 10.1126/science.7725107 . ПМИД 7725107 .
- ^ Ку О, Танака К, Голдберг А.Л. (1996). «Структура и функции 20S и 26S протеасом». Ежегодный обзор биохимии . 65 : 801–47. дои : 10.1146/annurev.bi.65.070196.004101 . ПМИД 8811196 .
- ^ Гролл М., Дитцель Л., Лёве Дж., Сток Д., Бохтлер М., Бартуник Х.Д., Хубер Р. (апрель 1997 г.). «Структура 20S протеасомы дрожжей с разрешением 2,4 А». Природа . 386 (6624): 463–71. Бибкод : 1997Natur.386..463G . дои : 10.1038/386463a0 . ПМИД 9087403 . S2CID 4261663 .
- ^ Дик Т.П., Нуссбаум А.К., Диг М., Хайнемейер В., Гролл М., Ширле М., Кейлхольц В., Стеванович С., Вольф Д.Х., Хубер Р., Раммензее Х.Г., Шильд Х. (октябрь 1998 г.). «Вклад протеасомальных бета-субъединиц в расщепление пептидных субстратов, проанализированных с помощью дрожжевых мутантов» . Журнал биологической химии . 273 (40): 25637–46. дои : 10.1074/jbc.273.40.25637 . ПМИД 9748229 .
- ^ Фогес Д., Цвикль П., Баумайстер В. (1999). «Протеасома 26S: молекулярная машина, предназначенная для контролируемого протеолиза». Ежегодный обзор биохимии . 68 (1): 1015–68. doi : 10.1146/annurev.biochem.68.1.1015 . ПМИД 10872471 .
- ^ Jump up to: а б Рап М., Йентч С. (май 2002 г.). «Откусить: обработка протеасомных белков». Природная клеточная биология . 4 (5): Е113–6. дои : 10.1038/ncb0502-e113 . ПМИД 11988749 . S2CID 7126477 .
- ^ Рап М., Йентш С. (ноябрь 2004 г.). «Продуктивный RUPture: активация факторов транскрипции путем протеасомного процессинга» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1695 (1–3): 209–13. дои : 10.1016/j.bbamcr.2004.09.022 . ПМИД 15571816 .
- ^ Ашер Дж., Реувен Н., Шауль Ю. (август 2006 г.). «20S протеасомы и деградация белков «по умолчанию» ». Биоэссе . 28 (8): 844–9. doi : 10.1002/bies.20447 . ПМИД 16927316 .
- ^ Чжан М., Пиккарт С.М., Коффино П. (апрель 2003 г.). «Детерминанты распознавания протеасом орнитиндекарбоксилазы, убиквитин-независимого субстрата» . Журнал ЭМБО . 22 (7): 1488–96. дои : 10.1093/emboj/cdg158 . ПМК 152902 . ПМИД 12660156 .
- ^ Ашер Дж., Шауль Ю. (август 2005 г.). «Протеосомная деградация p53: полиубиквитинирование – это еще не все» . Клеточный цикл . 4 (8): 1015–8. дои : 10.4161/cc.4.8.1900 . ПМИД 16082197 .
- ^ Jump up to: а б Шригарпур Р., Грюн Т., Мельхейс Дж., Дэвис К.Дж. (январь 2003 г.). «Конъюгация убиквитина не требуется для деградации окисленных белков протеасомой» . Журнал биологической химии . 278 (1): 311–8. дои : 10.1074/jbc.M206279200 . ПМИД 12401807 .
- ^ Jump up to: а б с Гилле С., Гёде А., Шлётельбург С., Прейсснер Р., Клёцель П.М., Гёбель У.Б., Фрёммель С. (март 2003 г.). «Комплексный взгляд на протеасомные последовательности: последствия для эволюции протеасомы». Журнал молекулярной биологии . 326 (5): 1437–48. дои : 10.1016/S0022-2836(02)01470-5 . ПМИД 12595256 .
- ^ Бохтлер М., Дитцель Л., Гролл М., Хартманн С., Хубер Р. (1999). «Протеасома». Ежегодный обзор биофизики и биомолекулярной структуры . 28 (1): 295–317. doi : 10.1146/annurev.biophys.28.1.295 . ПМИД 10410804 .
- ^ Чеснель Ф., Базиль Ф., Паскаль А., Кубяк Дж. З. (август 2006 г.). «Диссоциация циклина B от CDK1 предшествует его деградации при инактивации MPF в митотических экстрактах эмбрионов Xenopus laevis» . Клеточный цикл . 5 (15): 1687–98. дои : 10.4161/cc.5.15.3123 . ПМИД 16921258 .
- ^ Брито Д.А., Ридер К.Л. (июнь 2006 г.). «Проскальзывание митотической контрольной точки у людей происходит за счет разрушения циклина B в присутствии активной контрольной точки» . Современная биология . 16 (12): 1194–200. дои : 10.1016/j.cub.2006.04.043 . ПМЦ 2749311 . ПМИД 16782009 .
- ^ Хэвенс К.Г., Хо А., Йошиока Н., Дауди С.Ф. (июнь 2006 г.). «Регуляция позднего фазового перехода G1/S и APC Cdh1 активными формами кислорода» . Молекулярная и клеточная биология . 26 (12): 4701–11. дои : 10.1128/MCB.00303-06 . ПМЦ 1489138 . ПМИД 16738333 .
- ^ Башир Т., Доррелло Н.В., Амадор В., Гуардаваккаро Д., Пагано М. (март 2004 г.). «Контроль убиквитинлигазы SCF(Skp2-Cks1) с помощью убиквитинлигазы APC/C(Cdh1)». Природа . 428 (6979): 190–3. дои : 10.1038/nature02330 . ПМИД 15014502 . S2CID 4401971 .
- ^ Хигасицудзи Х., Лю Ю., Майер Р.Дж., Фудзита Дж. (октябрь 2005 г.). «Онкопротеин ганкирин отрицательно регулирует как р53, так и RB, усиливая протеасомную деградацию» . Клеточный цикл . 4 (10): 1335–7. дои : 10.4161/cc.4.10.2107 . ПМИД 16177571 .
- ^ Таррасон Риса, Габриэль; Хуртиг, Фредрик; Брей, Сиан; Хафнер, Энн Э.; Харкер-Киршнек, Лена; Фаулл, Питер; Дэвис, Колин; Папациаму, Димитра; Мутавчиев, Делян Р.; Фан, Кэтрин; Менегуэлло, Летисия; Араширо Пульшен, Андре; Дей, Гаутама; Калли, Сиан; Килкенни, Майри; Соуза, Диорж П.; Пеллегрини, Лука; де Брюин, Робертус AM; Энрикес, Рикардо; Снейдерс, Амброзиус П.; Шарич, Анжела; Линдос, Анн-Кристин; Робинсон, Николас П.; Баум, Базз (7 августа 2020 г.). «Протеасома контролирует деление клеток у археев, опосредованное ESCRT-III» . Наука . 369 (6504): eaaz2532. дои : 10.1126/science.aaz2532 . ПМК 7116001 . ПМИД 32764038 .
- ^ Дхармасири С., Эстель М. (2002). «Роль регулируемой деградации белка в ответе на ауксин». Молекулярная биология растений . 49 (3–4): 401–9. дои : 10.1023/A:1015203013208 . ПМИД 12036263 . S2CID 7669386 .
- ^ Вейерс Д., Бенкова Е., Ягер К.Е., Шлерет А., Хаманн Т., Кинц М., Уилмот Дж.К., Рид Дж.В., Юргенс Г. (май 2005 г.). «Специфичность развития реакции ауксина парами регуляторов транскрипции ARF и Aux/IAA» . Журнал ЭМБО . 24 (10): 1874–85. дои : 10.1038/sj.emboj.7600659 . ПМЦ 1142592 . ПМИД 15889151 .
- ^ Хаас А.Л., Бабошина О., Уильямс Б., Шварц Л.М. (апрель 1995 г.). «Координированная индукция пути конъюгации убиквитина сопровождает запрограммированную на этапе развития гибель скелетных мышц насекомых» . Журнал биологической химии . 270 (16): 9407–12. дои : 10.1074/jbc.270.16.9407 . ПМИД 7721865 .
- ^ Шварц Л.М., Майер А., Кош Л., Энгельштейн М., Майер С. (октябрь 1990 г.). «Активация экспрессии гена полиубиквитина во время запрограммированной гибели клеток». Нейрон . 5 (4): 411–9. дои : 10.1016/0896-6273(90)90080-Y . ПМИД 2169771 . S2CID 33829749 .
- ^ Лёв П., Бассел К., Доусон С.П., Биллетт М.А., Майер Р.Дж., Рейнольдс С.Е. (январь 1997 г.). «Экспрессия субъединицы 26S-протеасомы АТФазы, MS73, в мышцах, которые подвергаются запрограммированной в процессе развития гибели клеток, и ее контроль с помощью экдистероидных гормонов у насекомого Manduca sexta» . Письма ФЭБС . 400 (3): 345–9. дои : 10.1016/S0014-5793(96)01413-5 . ПМИД 9009228 . S2CID 10873052 .
- ^ Питцер Ф., Дантес А., Фукс Т., Баумайстер В., Амстердам А. (сентябрь 1996 г.). «Удаление протеасом из ядра и их накопление в апоптотических пузырях во время запрограммированной гибели клеток». Письма ФЭБС . 394 (1): 47–50. дои : 10.1016/0014-5793(96)00920-9 . ПМИД 8925925 . S2CID 29256092 .
- ^ Jump up to: а б Адамс Дж., Паломбелла В.Дж., Сосвилл Э.А., Джонсон Дж., Дестри А., Лазарус Д.Д., Маас Дж., Пиен К.С., Пракаш С., Эллиотт П.Дж. (июнь 1999 г.). «Ингибиторы протеасом: новый класс мощных и эффективных противоопухолевых средств». Исследования рака . 59 (11): 2615–22. ПМИД 10363983 .
- ^ Орловский Р.З. (апрель 1999 г.). «Роль убиквитин-протеасомного пути в апоптозе» . Смерть клеток и дифференцировка . 6 (4): 303–13. дои : 10.1038/sj.cdd.4400505 . ПМИД 10381632 .
- ^ Гарридо К., Брюне М., Дидело К., Цермати Ю., Шмитт Э., Кремер Г. (ноябрь 2006 г.). «Белки теплового шока 27 и 70: антиапоптотические белки с онкогенными свойствами» . Клеточный цикл . 5 (22): 2592–601. дои : 10.4161/cc.5.22.3448 . ПМИД 17106261 .
- ^ Пак С.Х., Болендер Н., Эйзель Ф., Костова З., Такеучи Дж., Коффино П., Вольф Д.Х. (январь 2007 г.). «Цитоплазматический механизм-шаперон Hsp70 подвергает неправильно свернутые и некомпетентные к импорту белки эндоплазматического ретикулума деградации через систему убиквитин-протеасома» . Молекулярная биология клетки . 18 (1): 153–65. doi : 10.1091/mbc.E06-04-0338 . ПМЦ 1751312 . ПМИД 17065559 .
- ^ Дай Цюань, Цянь С.Б., Ли Х.Х., Макдонаф Х., Борчерс С., Хуан Д., Такаяма С., Янгер Дж.М., Рен Х.И., Сир Д.М., Паттерсон С. (ноябрь 2005 г.). «Регуляция пути деградации белка контроля качества цитоплазмы с помощью BAG2» . Журнал биологической химии . 280 (46): 38673–81. дои : 10.1074/jbc.M507986200 . ПМИД 16169850 .
- ^ Бадер Н., Грюн Т. (2006). «Окисление белков и протеолиз». Биологическая химия . 387 (10–11): 1351–5. дои : 10.1515/BC.2006.169 . ПМИД 17081106 . S2CID 30385354 .
- ^ Дэвис К.Дж. (2003). «Деградация окисленных белков 20S протеасомой». Биохимия . 83 (3–4): 301–10. дои : 10.1016/S0300-9084(01)01250-0 . ПМИД 11295490 .
- ^ Jump up to: а б Lehman NL (сентябрь 2009 г.). «Убиквитиновая протеасомная система в невропатологии» . Акта Нейропатологика . 118 (3): 329–47. дои : 10.1007/s00401-009-0560-x . ПМК 2716447 . ПМИД 19597829 .
- ^ Макнот К.С., Джексон Т., ДжноБатист Р., Капустин А., Оланов К.В. (май 2006 г.). «Протеосомная дисфункция при спорадической болезни Паркинсона» . Неврология . 66 (10 Приложение 4): S37–49. дои : 10.1212/01.wnl.0000221745.58886.2e . ПМИД 16717251 .
- ^ Шарма Н., Брандис К.А., Эррера С.К., Джонсон Б.Е., Вайдья Т., Шреста Р., Деббурман С.К. (2006). «Модель почкующихся дрожжей альфа-синуклеина: токсичность усиливается за счет нарушения протеасомы и окислительного стресса». Журнал молекулярной нейронауки . 28 (2): 161–78. дои : 10.1385/JMN:28:2:161 . ПМИД 16679556 . S2CID 27762513 .
- ^ Мурата С., Сасаки К., Кишимото Т., Нива С., Хаяши Х., Такахама Ю., Танака К. (июнь 2007 г.). «Регуляция развития CD8+ Т-клеток с помощью протеасом, специфичных для тимуса». Наука . 316 (5829): 1349–53. Бибкод : 2007Sci...316.1349M . дои : 10.1126/science.1141915 . ПМИД 17540904 . S2CID 37185716 .
- ^ Касио П., Хилтон С., Киселев А.Ф., Рок К.Л., Голдберг А.Л. (май 2001 г.). «26S протеасомы и иммунопротеасомы продуцируют в основном N-расширенные версии антигенного пептида» . Журнал ЭМБО . 20 (10): 2357–66. дои : 10.1093/emboj/20.10.2357 . ПМК 125470 . ПМИД 11350924 .
- ^ Маллери Д.Л., Макьюэн В.А., Бидгуд С.Р., Тауэрс Дж.Дж., Джонсон К.М., Джеймс Л.К. (ноябрь 2010 г.). «Антитела опосредуют внутриклеточный иммунитет через трехчастный мотив, содержащий 21 (TRIM21)» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (46): 19985–19990. Бибкод : 2010PNAS..10719985M . дои : 10.1073/pnas.1014074107 . ПМЦ 2993423 . ПМИД 21045130 .
- ^ Фентеани Дж., Стандарт РФ, Лейн В.С., Чой С., Кори Э.Дж., Шрайбер С.Л. (май 1995 г.). «Ингибирование активности протеасом и субъединично-специфической модификации аминоконцевого треонина лактацистином». Наука . 268 (5211): 726–31. Бибкод : 1995Sci...268..726F . дои : 10.1126/science.7732382 . ПМИД 7732382 . S2CID 37779687 .
- ^ Пресс-релиз Управления по контролю за продуктами и лекарствами США. Архивировано 19 февраля 2007 г. в Wayback Machine 13 мая 2003 г. Дата доступа 29 декабря 2006 г. См. Также информационную страницу FDA Velcade .
- ^ Фишер Р.И., Бернштейн С.Х., Каль Б.С., Джулбегович Б., Робертсон М.Дж., де Вос С., Эпнер Э., Кришнан А., Леонард Дж.П., Лониал С., Штадтмауэр Э.А., О'Коннор О.А., Ши Х., Борал А.Л., Гой А. (октябрь 2006 г.) ). «Мультицентровое исследование фазы II бортезомиба у пациентов с рецидивирующей или рефрактерной мантийно-клеточной лимфомой» . Журнал клинической онкологии . 24 (30): 4867–74. дои : 10.1200/JCO.2006.07.9665 . ПМИД 17001068 .
- ^ Якоб С, Эгерер К, Либиш П, Тюркмен С, Заврски И, Кукелькорн У, Хайдер У, Кайзер М, Фляйсснер С, Стерц Дж, Клееберг Л, Файст Е, Бурместер Г.Р., Клёцель П.М., Сезер О (март 2007 г.). «Уровни циркулирующих протеасом являются независимым прогностическим фактором выживания при множественной миеломе» . Кровь . 109 (5): 2100–5. дои : 10.1182/blood-2006-04-016360 . ПМИД 17095627 .
- ^ Шах С.А., Поттер М.В., МакДейд Т.П., Риккарди Р., Перуджини Р.А., Эллиотт П.Дж., Адамс Дж., Каллери М.П. (2001). «Ингибирование 26S-протеасомы вызывает апоптоз и ограничивает рост рака поджелудочной железы человека». Журнал клеточной биохимии . 82 (1): 110–22. дои : 10.1002/jcb.1150 . ПМИД 11400168 . S2CID 21223980 .
- ^ Навроцкий С.Т., Суини-Готч Б., Такамори Р., МакКонки DJ (январь 2004 г.). «Ингибитор протеасом бортезомиб усиливает активность доцетаксела в ортотопических ксенотрансплантатах опухоли поджелудочной железы человека» . Молекулярная терапия рака . 3 (1): 59–70. дои : 10.1158/1535-7163.59.3.1 . ПМИД 14749476 . S2CID 38429730 .
- ^ Шенкейн Д. (июнь 2002 г.). «Ингибиторы протеасом в лечении В-клеточных злокачественных новообразований». Клиническая лимфома . 3 (1): 49–55. дои : 10.3816/CLM.2002.n.011 . ПМИД 12141956 .
- ^ О'Коннор О.А., Райт Дж., Московиц С., Маззи Дж., МакГрегор-Кортелли Б., Стабблфилд М., Штраус Д., Портлок С., Хэмлин П., Чой Е., Думетреску О., Эсселтин Д., Треху Е., Адамс Дж., Шенкейн Д., Зеленец нашей эры (февраль 2005 г.). «Клинический опыт фазы II применения нового ингибитора протеасом бортезомиба у пациентов с ленивой неходжкинской лимфомой и лимфомой мантийных клеток». Журнал клинической онкологии . 23 (4): 676–84. дои : 10.1200/JCO.2005.02.050 . ПМИД 15613699 .
- ^ Мессингер Ю.Х., Гайнон П.С., Спосто Р., ван дер Гиссен Дж., Экрот Э., Малвар Дж., Бостром БК (июль 2012 г.). «Бортезомиб в сочетании с химиотерапией высокоактивен при распространенном остром лимфобластном лейкозе-предшественнике B: исследование терапевтических достижений в области детской лейкемии и лимфомы (TACL)» . Кровь . 120 (2): 285–90. doi : 10.1182/blood-2012-04-418640 . ПМИД 22653976 .
- ^ Ламбру Г.И., Пападимитриу Л., Хрусос Г.П., Влахопулос С.А. (апрель 2012 г.). «Влияние глюкокортикоидов и ингибиторов протеасом на лейкемические лимфобласты: множественные разнообразные сигналы, сходящиеся на нескольких ключевых нижестоящих регуляторах». Молекулярная и клеточная эндокринология . 351 (2): 142–51. дои : 10.1016/j.mce.2012.01.003 . ПМИД 22273806 . S2CID 28749125 .
- ^ Шмидтке Г., Хольцхюттер Х.Г., Богьо М., Кайриес Н., Гролл М., де Джули Р., Эмч С., Гроеттруп М. (декабрь 1999 г.). «Как ингибитор протеазы ВИЧ-I модулирует активность протеасом» . Журнал биологической химии . 274 (50): 35734–40. дои : 10.1074/jbc.274.50.35734 . PMID 10585454 .
- ^ Лоран Н., де Буар С., Гильямо Ж.С., Христов С., Зини Р., Жуо Х., Андре П., Лотто В., Песчански М. (февраль 2004 г.). «Влияние ингибитора протеасом ритонавира на рост глиомы in vitro и in vivo» . Молекулярная терапия рака . 3 (2): 129–36. дои : 10.1158/1535-7163.129.3.2 . ПМИД 14985453 . S2CID 24423806 .
- ^ Золлнер Т.М., Подда М., Пьен С., Эллиотт П.Дж., Кауфманн Р., Бенке В.Х. (март 2002 г.). «Ингибирование протеасом снижает опосредованную суперантигеном активацию Т-клеток и тяжесть псориаза на модели SCID-hu» . Журнал клинических исследований . 109 (5): 671–9. дои : 10.1172/JCI12736 . ПМК 150886 . ПМИД 11877475 .
- ^ Эллиотт П.Дж., Пьен К.С., МакКормак Т.А., Чепмен И.Д., Адамс Дж. (август 1999 г.). «Ингибирование протеасом: новый механизм борьбы с астмой» . Журнал аллергии и клинической иммунологии . 104 (2 ч. 1): 294–300. дои : 10.1016/S0091-6749(99)70369-6 . ПМИД 10452747 .
- ^ Вердоес М., Флореа Б.И., Менендес-Бенито В., Мейнард С.Дж., Витте М.Д., ван дер Линден В.А., ван ден Ньювендейк А.М., Хофманн Т., Беркерс Ч.Р., ван Леувен Ф.В., Гротуис Т.А., Леувенбург М.А., Оваа Х., Нефджес Дж.Дж., Филиппов Д.В., ван дер Марел Г.А., Дантума Н.П., Оверклифт Х.С. (ноябрь 2006 г.). «Флуоресцентный ингибитор протеасом широкого спектра действия для мечения протеасом in vitro и in vivo» . Химия и биология . 13 (11): 1217–26. doi : 10.1016/j.chembiol.2006.09.013 . hdl : 1887/65477 . ПМИД 17114003 .
- ^ Кляйгер Г., мэр Т. (июнь 2014 г.). «Опасное путешествие: экскурсия по системе убиквитин-протеасома» . Тенденции клеточной биологии . 24 (6): 352–9. дои : 10.1016/j.tcb.2013.12.003 . ПМК 4037451 . ПМИД 24457024 .
- ^ Голдберг А.Л., Стейн Р., Адамс Дж. (август 1995 г.). «Новый взгляд на функцию протеасом: от архебактерий до разработки лекарств» . Химия и биология . 2 (8): 503–8. дои : 10.1016/1074-5521(95)90182-5 . ПМИД 9383453 .
- ^ Сулистио Я.А., Хизе К. (январь 2015 г.). «Система убиквитин-протеасома и нарушение регуляции молекулярных шаперонов при болезни Альцгеймера». Молекулярная нейробиология . 53 (2): 905–31. дои : 10.1007/s12035-014-9063-4 . ПМИД 25561438 . S2CID 14103185 .
- ^ Ортега З., Лукас Джей-Джей (2014). «Вовлечение системы убиквитин-протеасома в болезнь Гентингтона» . Границы молекулярной нейронауки . 7 : 77. doi : 10.3389/fnmol.2014.00077 . ПМК 4179678 . ПМИД 25324717 .
- ^ Сандри М., Роббинс Дж. (июнь 2014 г.). «Протеотоксичность: недооцененная патология сердечно-сосудистых заболеваний» . Журнал молекулярной и клеточной кардиологии . 71 : 3–10. дои : 10.1016/j.yjmcc.2013.12.015 . ПМК 4011959 . ПМИД 24380730 .
- ^ Дрюс О., Тэгтмейер Х. (декабрь 2014 г.). «Нацеливание на систему убиквитин-протеасома при заболеваниях сердца: основа новых терапевтических стратегий» . Антиоксиданты и окислительно-восстановительная сигнализация . 21 (17): 2322–43. дои : 10.1089/ars.2013.5823 . ПМК 4241867 . ПМИД 25133688 .
- ^ Ван З.В., Хилл Дж.А. (февраль 2015 г.). «Контроль качества белка и метаболизма: двунаправленный контроль в сердце» . Клеточный метаболизм . 21 (2): 215–26. дои : 10.1016/j.cmet.2015.01.016 . ПМЦ 4317573 . ПМИД 25651176 .
- ^ Jump up to: а б Карин М., Делхасе М. (февраль 2000 г.). «Киназа I каппа B (IKK) и NF-каппа B: ключевые элементы передачи провоспалительных сигналов». Семинары по иммунологии . 12 (1): 85–98. дои : 10.1006/smim.2000.0210 . ПМИД 10723801 .
- ^ Ермолаева М.А., Даховник А, Шумахер Б (январь 2015 г.). «Механизмы контроля качества клеточных и системных реакций на повреждение ДНК» . Обзоры исследований старения . 23 (Часть А): 3–11. дои : 10.1016/j.arr.2014.12.009 . ПМЦ 4886828 . ПМИД 25560147 .
- ^ Чеклер Ф, да Коста К.А., Анколио К., Шевалье Н., Лопес-Перес Э., Марамбо П. (июль 2000 г.). «Роль протеасомы в болезни Альцгеймера» . Biochimica et Biophysica Acta (BBA) - Молекулярные основы болезней . 1502 (1): 133–8. дои : 10.1016/s0925-4439(00)00039-9 . ПМИД 10899438 .
- ^ Jump up to: а б Чунг К.К., Доусон В.Л., Доусон Т.М. (ноябрь 2001 г.). «Роль убиквитин-протеасомного пути при болезни Паркинсона и других нейродегенеративных расстройствах». Тенденции в нейронауках . 24 (11 Приложение): С7–14. дои : 10.1016/s0166-2236(00)01998-6 . ПМИД 11881748 . S2CID 2211658 .
- ^ Jump up to: а б Икеда К., Акияма Х., Арай Т., Уэно Х., Цучия К., Косака К. (июль 2002 г.). «Морфометрическая переоценка системы двигательных нейронов при болезни Пика и боковом амиотрофическом склерозе с деменцией». Акта Нейропатологика . 104 (1): 21–8. дои : 10.1007/s00401-001-0513-5 . ПМИД 12070660 . S2CID 22396490 .
- ^ Манака Х, Като Т, Курита К, Катагири Т, Шикама Ю, Куджирай К, Каванами Т, Сузуки Ю, Нихей К, Сасаки Х (май 1992 г.). «Заметное увеличение убиквитина спинномозговой жидкости при болезни Крейтцфельдта-Якоба». Письма по неврологии . 139 (1): 47–9. дои : 10.1016/0304-3940(92)90854-з . ПМИД 1328965 . S2CID 28190967 .
- ^ Мэтьюз К.Д., Мур С.А. (январь 2003 г.). «Поясно-конечно-поясная мышечная дистрофия». Текущие отчеты по неврологии и нейробиологии . 3 (1): 78–85. дои : 10.1007/s11910-003-0042-9 . ПМИД 12507416 . S2CID 5780576 .
- ^ Майер Р.Дж. (март 2003 г.). «От нейродегенерации к нейрогомеостазу: роль убиквитина». Новости и перспективы наркотиков . 16 (2): 103–8. дои : 10.1358/dnp.2003.16.2.829327 . ПМИД 12792671 .
- ^ Калис Дж., Пауэлл С.Р. (февраль 2013 г.). «Убиквитиновая протеасомная система и ишемия миокарда» . Американский журнал физиологии. Физиология сердца и кровообращения . 304 (3): H337–49. дои : 10.1152/ajpheart.00604.2012 . ПМЦ 3774499 . ПМИД 23220331 .
- ^ Предмор Дж.М., Ван П., Дэвис Ф., Бартолоне С., Вестфолл М.В., Дайк Д.Б., Пагани Ф., Пауэлл С.Р., Дэй С.М. (март 2010 г.). «Дисфункция убиквитиновых протеасом при гипертрофических и дилатационных кардиомиопатиях человека» . Тираж . 121 (8): 997–1004. doi : 10.1161/CIRCULATIONAHA.109.904557 . ПМЦ 2857348 . ПМИД 20159828 .
- ^ Пауэлл С.Р. (июль 2006 г.). «Убиквитин-протеасомная система в физиологии и патологии сердца». Американский журнал физиологии. Физиология сердца и кровообращения . 291 (1): H1–H19. дои : 10.1152/ajpheart.00062.2006 . ПМИД 16501026 . S2CID 7073263 .
- ^ Адамс Дж. (апрель 2003 г.). «Возможность ингибирования протеасом при лечении рака». Открытие наркотиков сегодня . 8 (7): 307–15. дои : 10.1016/s1359-6446(03)02647-3 . ПМИД 12654543 .
- ^ Бен-Нерия Ю. (январь 2002 г.). «Регуляторные функции убиквитинирования в иммунной системе». Природная иммунология . 3 (1): 20–6. дои : 10.1038/ni0102-20 . ПМИД 11753406 . S2CID 26973319 .
- ^ Эгерер К., Кукелькорн У., Рудольф П.Е., Рюкерт Дж.К., Дёрнер Т., Бурместер Г.Р., Клёцель П.М., Файст Э. (октябрь 2002 г.). «Циркулирующие протеасомы являются маркерами повреждения клеток и иммунологической активности при аутоиммунных заболеваниях». Журнал ревматологии . 29 (10): 2045–52. ПМИД 12375310 .
Дальнейшее чтение
[ редактировать ]- Гликман М.Х., Адир Н. (январь 2004 г.). «Протеасома и тонкий баланс между разрушением и спасением» . ПЛОС Биология . 2 (1): е13. doi : 10.1371/journal.pbio.0020013 . ПМК 314468 . ПМИД 14737189 .
- Протеасома дрожжей 26S со списком субъединиц и изображениями
- Чехановер А. (сентябрь 2005 г.). «Ранние работы над протеасомной системой убиквитина, интервью с Аароном Чехановером. Интервью CDD» . Смерть клеток и дифференцировка . 12 (9): 1167–77. дои : 10.1038/sj.cdd.4401691 . ПМИД 16094393 .
- Гершко А (сентябрь 2005 г.). «Ранние работы над убиквитиновой протеасомной системой, интервью с Аврамом Гершко. Интервью CDD» . Смерть клеток и дифференцировка . 12 (9): 1158–61. дои : 10.1038/sj.cdd.4401709 . ПМИД 16094391 .
- Роуз I (сентябрь 2005 г.). «Ранние работы над протеасомной системой убиквитина, интервью с Ирвином Роузом. Интервью CDD» . Смерть клеток и дифференцировка . 12 (9): 1162–6. дои : 10.1038/sj.cdd.4401700 . ПМИД 16094392 .
- Цвек Б, Дворжак З (2007). «Нацеливание на ядерный фактор-каппаВ и протеасому с помощью дитиокарбаматных комплексов с металлами» . Текущий фармацевтический дизайн . 13 (30): 3155–67. дои : 10.2174/138161207782110390 . ПМИД 17979756 . Архивировано из оригинала 29 июля 2012 года.
Внешние ссылки
[ редактировать ]- Руководство по номенклатуре субъединиц протеасом
- 3D-структуры протеасом в Банке данных EM (EMDB)
- Ключевые моменты протеасомы. Архивировано 17 ноября 2020 г. в Wayback Machine. функции
| Базы данных органов управления : Национальные |
|---|