Комплекс, стимулирующий анафазу

Комплекс, способствующий анафазе (также называемый циклосомой или APC/C E3 ), представляет собой убиквитинлигазу , которая маркирует белки-мишени клеточного цикла для деградации с помощью 26S протеасомы . APC/C представляет собой большой комплекс из 11–13 белков- субъединиц , включая субъединицу куллина ( Apc2 ) и RING ( Apc11 ), очень похожую на SCF . Другие части APC/C имеют неизвестные функции, но хорошо сохранились . [ 1 ]
Открытие APC/C (и SCF ) и их ключевой роли в регуляции клеточного цикла эукариот установило важность убиквитин -опосредованного протеолиза в клеточной биологии. воспринимались как система, участвующая исключительно в удалении поврежденного белка из клетки, Когда-то убиквитинирование и последующая деградация белка протеасомой теперь воспринимается как универсальный регуляторный механизм передачи сигнала , важность которого приближается к значению фосфорилирования белка .
В 2014 году APC/C был нанесен на карту в 3D с разрешением менее нанометра , что также выявило его вторичную структуру. Это открытие может улучшить понимание рака и открыть новые места связывания для будущих лекарств от рака. [ 2 ] [ 3 ]
Функция
[ редактировать ]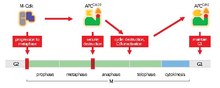
Основная функция APC/C — инициировать переход от метафазы к анафазе путем маркировки определенных белков для деградации. Тремя основными объектами деградации APC/C являются секурин и S- и M -циклины . При разложении секурин высвобождает сепаразу , протеазу. Затем сепараза запускает расщепление когезина , белкового комплекса, который связывает сестринские хроматиды вместе. Во время метафазы сестринские хроматиды соединяются интактными когезиновыми комплексами. Когда секурин подвергается убиквитинированию с помощью APC/C и высвобождает сепаразу, которая разрушает когезин, сестринские хроматиды могут свободно перемещаться к противоположным полюсам для анафазы. APC/C также нацелен на деградацию митотических циклинов , что приводит к инактивации комплексов M-CDK (митотическая циклин-зависимая киназа ), способствуя выходу из митоза и цитокинезу . [ 1 ]
В отличие от SCF, субъединицы-активаторы контролируют APC/C. Cdc20 и Cdh1 являются двумя активаторами, имеющими особое значение для клеточного цикла. Эти белки направляют APC/C на определенные наборы субстратов в разные моменты клеточного цикла, тем самым продвигая его вперед. APC/C также играет важную роль в поддержании метаболизма хроматина, особенно в G 1 и G 0 , и играет ключевую роль в фосфорилировании H3 посредством разрушения киназы Aurora A . [ 4 ]
Критическими субстратами APC/C являются секурин и циклины типа B. Это сохраняется у млекопитающих и дрожжей. Фактически, дрожжи жизнеспособны в отсутствие APC/C, если устранена необходимость нацеливания на эти два субстрата. [ 5 ]
Субъединицы
[ редактировать ]Субъединиц APC/C, которые служат в основном адаптерами, не проводилось большого количества обширных исследований. Исследования субъединиц APC в основном проводятся на дрожжах, и исследования показывают, что большинство субъединиц APC дрожжей также присутствуют у позвоночных, что предполагает их сохранение у эукариот. У позвоночных обнаружено одиннадцать основных субъединиц APC по сравнению с тринадцатью у дрожжей. [ 1 ] Субъединицы активатора связываются с APC на различных стадиях клеточного цикла, чтобы контролировать его активность убиквитинирования, часто направляя APC на целевые субстраты, предназначенные для убиквитинирования. Предполагается, что специфичность лигаз APC контролируется путем включения факторов специфичности в лигазный комплекс вместо фосфорилирования субстрата. т.е.: субъединица CDC20 позволяет APC разрушать субстраты, такие как ингибиторы анафазы (Pdsp1) в начале анафазы, с другой стороны, когда CDC20 заменяется фактором специфичности Hct1, APC разрушает другой набор субстратов, особенно циклины митоза в конце анафазы. анафаза. Активаторы CDC20 и Cdh1 имеют особое значение и являются наиболее широко изученными и известными из субъединиц APC/C.
Каталитическое ядро APC/C состоит из субъединицы куллина Apc2 и субъединицы домена RING H2 Apc11. Эти две субъединицы катализируют убиквитинирование субстратов, когда С-концевой домен Apc2 образует плотный комплекс с Apc11. RING/APc11 связывается с конъюгатом E2-убиквитин, который катализирует перенос убиквитина в активный сайт E2. [ 1 ] Помимо каталитической функциональности, другие основные белки APC состоят из множественных повторяющихся мотивов, основная цель которых - обеспечение поддержки молекулярного каркаса. К ним относятся Apc1, самая большая субъединица, которая содержит 11 тандемных повторов из 35–40 аминокислотных последовательностей, и Apc2, которая содержит три повтора куллина , в общей сложности примерно 130 аминокислот. [ 6 ] Основные мотивы в субъединицах APC включают мотивы тетратрикопептида (TPR) и повторы WD40 1. [ 1 ] С-концевые области CDC20 и Cdh1 имеют домен WD40, который, как предполагается, образует связывающую платформу, которая связывает субстраты APC, тем самым способствуя способности APC нацеливаться на эти субстраты, хотя точный механизм, посредством которого они увеличивают активность APC, неизвестен. [ 7 ] Также предполагается, что вариации в этих доменах WD40 приводят к различной субстратной специфичности, что подтверждается недавними результатами, предполагающими, что разные субстраты APC могут напрямую и специфично связываться с Cdc20 и Cdh1/Hct1. В конечном итоге различия в специфичности ответственны за время разрушение нескольких мишеней APC во время митоза. CDC20 нацелен на несколько основных субстратов в метафазе, а Cdh1 нацелен на более широкий спектр субстратов в направлении позднего митоза и G1. [ 8 ]
В частности, 4 субъединицы дрожжевого APC/C почти полностью состоят из множественных повторов мотива из 34 аминокислотных остатков тетратрикопептида (TPR). Эти субъединицы TPR, Cdc16, [ 9 ] CDC27 , [ 10 ] Cdc23 и Apc5 в основном обеспечивают каркас и поддержку для осуществления других межбелковых взаимодействий. Было показано, что Cdc27 и Cdc23 поддерживают связывание Cdc20 и Cdh1, поскольку мутации в ключевых остатках этих субъединиц приводят к усилению диссоциации активаторов. Было показано, что Apc10/Doc1 способствует связыванию субстрата, опосредуя их взаимодействие с Cdh1 и Cdc20. [ 11 ]
В частности, CDC20 (также известный как p55CDC, Fizzy или Slp1) инактивирует CDK1 посредством убиквитинирования циклинов B-типа. Это приводит к активации Cdh1 (также известного как Fizzy-родственный, Hct1, Ste9 или Srw1), который взаимодействует с APC во время позднего митоза и G1/G0. Cdh1 инактивируется посредством фосфорилирования во время S, G2 и ранней M-фазы. В эти моменты цикла его невозможно собрать. [ 12 ]
Данные показывают, что APC3 и APC7 служат для привлечения Cdh1 в комплекс, способствующий анафазе. [ 13 ] Это также подтверждает, что Cdh1 отвечает за поддержание активности APC во время G1. Cdh1 не требует фосфорилирования APC для связывания, фактически фосфорилирование Cdh1 с помощью Cdks предотвращает его связывание с APC из фазы S в фазу M. После разрушения M-Cdk теперь может произойти высвобождение CDC20 из APC и связывание Cdh1, что позволяет активности APC продолжаться во время входа в G1. [ 1 ] Хотя Cdh1 распознает циклины M и S, позволяя их разрушать до тех пор, пока вся клетка не перейдет к новому циклу, он не распознает циклины G1/S, и во время фазы G1/S их циклиновая активность может беспрепятственно повышаться и фосфорилироваться и, таким образом, инактивируя Cdh1 и, следовательно, APC.
Субъединица Apc15 играет важную роль в APC/C. Cdc20 активация после биориентации сестринских хроматид через метафазную пластинку. Когда кинетохоры не прикреплены к веретенам, комплексы митотических контрольных точек (MCC) и ингибируют APC. При отсутствии Apc15, MCC и Cdc20 остаются заблокированными на APC/C, предотвращая его активность после выполнения требований контрольной точки шпинделя. Apc15 опосредует обмен Cdc20 и MCC, предоставляя информацию о состоянии прикрепления кинетохор. [ 14 ]
CDC27/APC3
[ редактировать ]Было идентифицировано, что одна из субъединиц, демонстрирующих мотив TPR, CDC27, взаимодействует с белками митотических контрольных точек, такими как Mad2, p55CDC и BUBR1, что позволяет предположить, что она может участвовать в определении времени М-фазы. [ 15 ] Данные показывают, что CDC27 участвует в тройном комплексе с SMAD2/3 и Cdh1, который создается в ответ на передачу сигналов TGFβ. В частности, из-за его взаимодействия с Cdh1 он может играть потенциальную роль в определении сродства между APC и его активаторами Cdc20 и Cdh1. Исследование показывает, что фосфорилирование Cdc27, индуцированное TGF-β, усиливает взаимодействие между cdc27 и Cdh1, которое непосредственно участвует в активации APC. [ 16 ] CDC27 может служить средством, посредством которого передача сигналов TGFβ может активировать APC. Индуцированное гиперфосфорилирование CDC27 с помощью TGFβ показало повышенную активность APC.
CDC23, CDC16, CDC27
[ редактировать ]CDC23, другая субъединица TPR, взаимодействует с SWM1, связываясь с D-боксом CLB2. На основании гибридных анализов in vivo и коиммунопреципитации in vitro предполагается, что Cdc16p, Cdc23p и Cdc27p (аналоги в Sacchyromyces cerevisiae) взаимодействуют и образуют макромолекулярный комплекс. Предполагается, что их общая тема TPR опосредует эти взаимодействия. [ 17 ] Что касается Cdc27 и Cdc16 у дрозофилы, их функции были проверены с помощью РНК-интерференции (RNAi). [ 18 ] Результаты показывают, что они могут опосредовать активность всего комплекса посредством разных механизмов в разных сайтах. В дальнейших исследованиях дрозофилы Cdk16 и cdk23, по-видимому, активируются посредством фосфорилирования Polo-подобной киназой 1 (Plk1), а ее аналог делящихся дрожжей, по-видимому, особенно связывается с Cdc23. [ 19 ]
Предполагается, что комплекс регулируется активаторами CDC20 и Cdh1 во время митоза. Их роль в деградации циклина B продемонстрирована скринингом мутантов Saccharomyces cerevisiae, дефектных по деградации циклина B, у которых были обнаружены мутации в генах CDC16 и CDC23. Мутанты CDC27, CDC23 и CDC27 приводили к остановке клеточного цикла в метафазе. [ 20 ]
Распознавание субстрата
[ редактировать ]Субстраты APC/C имеют аминокислотные последовательности распознавания, которые позволяют APC/C идентифицировать их. Наиболее распространенная последовательность известна как ящик разрушения или D-бокс. APC/C объединяет фермент, конъюгирующий убиквитин E2 , и D-бокс, а не является промежуточным ковалентным носителем. [ 21 ] D-бокс должен иметь версию следующей аминокислотной последовательности: RXXLXXXXN, где R — аргинин , X — любая аминокислота, L — лейцин , а N — аспарагин . Ящик Кена — еще один важный мотив. Его последовательность должна напоминать следующую: KENXXXN, где K — лизин , а E — глутамат . Последняя позиция аминокислоты в Кен-боксе сильно варьируется. Хотя было показано, что мутации в последовательностях действительно ингибируют разрушение белков «in vivo», еще многое предстоит узнать о том, как APC/C воздействует на белки. [ 1 ]
После связывания с APC/C Cdc20 и Cdh1 служат рецепторами D- и KEN-бокса для различных субстратов APC. Крафт и др. показали, что D-боксы субстратов связываются непосредственно с высококонсервативной областью пропеллера повторов WD40 на активаторах APC. Важно отметить, что консервативная площадь пропеллера Cdh1 намного больше, чем у Cdc20, что позволяет Cdh1 иметь более широкую субстратную специфичность, что согласуется с тем фактом, что APC/C Cdh1 также активирует APC-опосредованное разрушение KEN-бокса, содержащего субстраты. D-бокс дополнительно усиливает деградацию белка, поскольку остатки лизина в непосредственной близости от D-бокса служат мишенями убиквитилирования. Было обнаружено, что остаток Lys, расположенный непосредственно на С-конце D-бокса, может функционировать как акцептор убиквитина. [ 22 ]
Многие подложки APC содержат как D, так и KEN-боксы с их убиквитилированием либо APC/C. Cdc20 или АПК/С Cdh1 зависит от обеих последовательностей, однако некоторые субстраты содержат только D-бокс или KEN-бокс в одной или нескольких копиях. Наличие двух различных последовательностей деградации обеспечивает высокий уровень субстратной специфичности APC/C. Cdc20 большая зависимость от D-box и APC/C Cdh1 больше зависит от коробки KEN. Например, АПК/С Cdh1 способен убиквитилировать субстраты, содержащие только KEN-бокс, такие как Tome-1 и Sororin. [ 6 ]
Хотя Cdc20 и Cdh1 могут служить рецепторами D- и KEN-бокса, низкое сродство этих взаимодействий коактиватор-субстрат предполагает, что маловероятно, что одних коактиваторов достаточно для обеспечения высокоаффинного связывания субстрата с APC/C. Cdc20 и АПК/С Cdh1 . [ 6 ] Следовательно, основные субъединицы APC/C, такие как Apc10, также вносят вклад в ассоциацию субстрата. В конструкциях APC/C, лишенных субъединицы Apc10/Doc1, такие субстраты, как Clb2, не могут связываться с APC. Δdoc1 –Cdh1, при добавлении очищенного Doc1 к APC Δdoc1 Конструкция –Cdh1 восстанавливает способность связывания субстрата. [ 11 ]
Переход от метафазы к анафазе
[ редактировать ]Когда начинается метафаза, контрольная точка веретена ингибирует APC/C до тех пор, пока все сестринские кинетохоры не прикрепятся к противоположным полюсам митотического веретена - процесс, известный как биоориентация хромосом. Когда все кинетохоры правильно прикреплены, контрольная точка веретена замолкает, и APC/C может стать активным. M-Cdks фосфорилирует субъединицы APC/C, которые способствуют связыванию с Cdc20. Секурин и M-циклины (циклин A и циклин B) затем подвергаются воздействию APC/C. Cdc20 на деградацию. После деградации высвобождается сепарин, когезин разрушается, и сестринские хроматиды готовятся переместиться к своим соответствующим полюсам для анафазы. [ 1 ]
Вполне вероятно, что в клетках животных по крайней мере некоторая активация APC/C Cdc20 происходит на ранних стадиях клеточного цикла (профаза или прометафаза) в зависимости от времени деградации его субстратов. Циклин А разлагается на ранних стадиях митоза, что подтверждает теорию, но циклин В и секурин не разлагаются до метафазы. Молекулярная основа задержки неизвестна, но считается, что она является ключом к правильному выбору времени начала анафазы. В животных клетках система контрольных точек веретена способствует задержке, если необходимо исправить биориентацию хромосом. Однако неизвестно, каким образом система контрольных точек веретена ингибирует разрушение циклина B и секурина, одновременно позволяя разлагать циклин А. Задержку также можно объяснить неизвестными взаимодействиями с регуляторами, изменениями локализации и фосфорилирования. [ 1 ]
Это запускает петлю отрицательной обратной связи . При активации APC/C Cdc20 требуется M-Cdk, комплекс также отвечает за разрушение циклина и деактивацию M-CdK. Это означает, что APC/C Cdc20 способствует собственной дезактивации. Возможно, что эта отрицательная обратная связь является основой активности Cdk, контролируемой колебаниями концентрации M и S циклинов. [ 1 ]
от M к G 1 Переход
[ редактировать ]Важно, чтобы по завершении митоза клетки (за исключением эмбриональных) прошли период роста, известный как G 1 фаза , для роста и производства факторов, необходимых для следующего клеточного цикла. Вход в следующий раунд митоза предотвращается путем ингибирования активности Cdk. Хотя за это ингибирование ответственны разные процессы, важным из них является активация APC/C с помощью Cdh1. Эта продолжающаяся активация предотвращает накопление циклина, который запускает новый раунд митоза, и вместо этого приводит к выходу из митоза. [ 1 ]
В начале клеточного цикла Cdh1 фосфорилируется с помощью M-Cdk, предотвращая его присоединение к APC/C. Затем APC/C может свободно прикрепиться к Cdc20 и начать переход от метафазы к анафазе. Поскольку M-Cdk позже разрушается в ходе митоза, Cdc20 высвобождается, и Cdh1 может связываться с APC/C, поддерживая его активацию посредством перехода M/ G1 . Ключевое отличие, которое следует отметить, заключается в том, что хотя связывание Cdc20 с APC/C зависит от фосфорилирования APC/C митотическими Cdks, связывание Cdh1 не зависит. Таким образом, как АПК Cdc20 становится инактивированным во время метафазы из-за дефосфорилирования в результате неактивных митотических Cdks, Cdh1 способен немедленно связываться с APC/C, занимая место Cdc20. Cdc20 также является целью APC/C. Cdh1 , гарантируя, что APC/C Cdc20 выключен. БТР/С Cdh1 затем продолжает работу в G 1, чтобы пометить циклины S и M для уничтожения. Однако циклины G 1 /S не являются субстратами APC/C. Cdh1 и, следовательно, накапливаются на протяжении этой фазы и фосфорилируют Cdh1. К концу G 1 достаточное количество циклинов G 1 /S накопилось и фосфорилировало Cdh1, чтобы инактивировать APC/C до следующей метафазы. [ 1 ]
Оказавшись в G1 , APC Cdh1 отвечает за деградацию различных белков, которые способствуют правильному развитию клеточного цикла. Геминин — это белок, который связывается с Cdt1, что предотвращает его связывание с комплексом распознавания происхождения (ORC). БТР Cdh1 нацелен на геминин для убиквитинирования в G 1 , сохраняя его уровни на низком уровне. Это позволяет Cdt1 выполнять свою функцию во время сборки перед RC. Когда АПК Cdh1 становится неактивным вследствие фосфорилирования Cdh1 циклинами G 1 /S, активность геминина снова повышается. Кроме того, Dbf4 стимулирует активность протеинкиназы, связанной с 7-м циклом деления клеток (Cdc7), что способствует активации источников репликации. Предполагается, что APCCdh1 нацелен на уничтожение Dbf4. Это может дать ответ на вопрос, как Cdc7 активируется в начале нового клеточного цикла. Его активность, вероятно, соответствует инактивации APC/C. Cdh1 G/S-циклинами. [ 1 ]
Дополнительное регулирование
[ редактировать ]БТР/С Cdc20 инактивация на ранних стадиях клеточного цикла частично достигается белком Emi1. Первоначальные эксперименты показали, что добавление Emi1 к экстрактам цикла Xenopus может предотвратить разрушение эндогенного циклина A, циклина B и выход из митоза, что позволяет предположить, что Emi1 способен противодействовать активности APC. Более того, истощение Emi1 в соматических клетках приводит к отсутствию накопления циклина B. Отсутствие Emi1, вероятно, приводит к отсутствию ингибирования APC, предотвращающему накопление циклина B. [ 23 ]
На основании этих ранних наблюдений было подтверждено, что при G2 и раннем митозе Emi1 связывает и ингибирует Cdc20, предотвращая его ассоциацию с субстратами APC. Cdc20 все еще может фосфорилироваться и связываться с APC/C, но связанный Emi1 блокирует взаимодействие Cdc20 с мишенями APC. [ 1 ] Ассоциация Emi1 с Cdc20 позволяет стабилизировать различные циклины на протяжении фаз S и G2, но удаление Emi1 важно для прогрессирования митоза. Таким образом, в поздней профазе Emi1 фосфорилируется Polo-подобной киназой Plk. Plk активируется во время раннего митоза за счет активности Cdk1, и его фосфорилирование сайта связывания βTrCP BTRC (гена) Emi1 делает его мишенью для SCF, что приводит к его последующему разрушению в прометафазе. [ 24 ] Разрушение Emi1 приводит к активации APC/CCdc20, что позволяет разрушить циклин А в раннем митозе. Уровни Emi1 снова начинают повышаться в G, что помогает ингибировать APC/C. Cdh1 . [ 1 ]
Регулирование APC/C Cdc20 активность по отношению к метафазным субстратам, таким как секурин и циклин B, может быть результатом внутриклеточной локализации. Предполагается, что белки контрольной точки веретена, ингибирующие APC/C, Cdc20 ассоциированы только с подмножеством популяции Cdc20, локализованной вблизи митотического веретена. Таким образом, циклин А может расщепляться, тогда как циклин В и секурин разрушаются только после того, как сестринские хроматиды достигают биориентации. [ 1 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п Морган Д.О. (2007). «Глава 3-10: Комплекс, способствующий анафазе» . Клеточный цикл: принципы управления . Лондон: New Science Press. стр. 48–49. ISBN 978-0-9539181-2-6 .
- ^ «Ученые составили карту одного из самых важных белков в жизни – и рака» . Институт исследования рака . 20 июля 2014 года . Проверено 22 июля 2014 г.
- ^ Чанг Л.Ф., Чжан З., Ян Дж., Маклафлин С.Х., Барфорд Д. (сентябрь 2014 г.). «Молекулярная архитектура и механизм комплекса, способствующего анафазе» . Природа . 513 (7518): 388–393. Бибкод : 2014Natur.513..388C . дои : 10.1038/nature13543 . ПМК 4456660 . ПМИД 25043029 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П., ред. (2002). «Глава 17. Клеточный цикл и запрограммированная смерть клеток» . Молекулярная биология клетки (4-е изд.). Гирляндная наука. ISBN 0-8153-3218-1 .
- ^ Торнтон Б.Р., Точиски Д.П. (декабрь 2003 г.). «Секурин и B-циклин/CDK являются единственными важными мишенями APC». Природная клеточная биология . 5 (12): 1090–4. дои : 10.1038/ncb1066 . ПМИД 14634663 . S2CID 30582585 .
- ^ Перейти обратно: а б с Барфорд Д. (декабрь 2011 г.). «Структурное понимание сложных функций и механизмов, способствующих анафазе» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 366 (1584): 3605–24. дои : 10.1098/rstb.2011.0069 . ПМК 3203452 . ПМИД 22084387 .
- ^ Кастро А., Бернис С., Виньерон С., Лаббе Х.К., Лорка Т. (январь 2005 г.). «Комплекс, способствующий анафазе: ключевой фактор регуляции клеточного цикла». Онкоген . 24 (3): 314–25. дои : 10.1038/sj.onc.1207973 . ПМИД 15678131 . S2CID 29467714 .
- ^ Шваб М., Нойцнер М., Мёккер Д., Зойферт В. (сентябрь 2001 г.). «Дрожжи Hct1 распознают митотический циклин Clb2 и другие субстраты убиквитинлигазы APC» . Журнал ЭМБО . 20 (18): 5165–75. дои : 10.1093/emboj/20.18.5165 . ПМК 125620 . ПМИД 11566880 .
- ^ Ван Дж., Дай Б.Т., Раджашанкар К.Р., Куринов И., Шульман Б.А. (сентябрь 2009 г.). «Понимание анафазы, способствующей сборке сложного субдомена TPR из структуры CDC26-APC6» . Структурная и молекулярная биология природы . 16 (9): 987–9. дои : 10.1038/nsmb.1645 . ПМЦ 2759704 . ПМИД 19668213 .
- ^ Ямагучи М., Ю С., Цяо Р., Вайсманн Ф., Миллер Дж., ВандерЛинден Р. и др. (апрель 2015 г.). «Структура комплекса APC3-APC16: понимание сборки комплекса/циклосомы, способствующего анафазе» . Журнал молекулярной биологии . 427 (8): 1748–64. дои : 10.1016/j.jmb.2014.11.020 . ПМЦ 4444369 . ПМИД 25490258 .
- ^ Перейти обратно: а б Пассмор Л.А. , МакКормак Э.А., Ау С.В., Пол А., Уиллисон К.Р., Харпер Дж.В., Барфорд Д. (февраль 2003 г.). «Doc1 опосредует активность комплекса, способствующего анафазе, способствуя распознаванию субстрата» . Журнал ЭМБО . 22 (4): 786–96. дои : 10.1093/emboj/cdg084 . ПМК 145444 . ПМИД 12574115 .
- ^ Крамер Э.Р., Шойрингер Н., Подтелейников А.В., Манн М., Петерс Дж.М. (май 2000 г.). «Митотическая регуляция белков-активаторов APC CDC20 и CDH1» . Молекулярная биология клетки . 11 (5): 1555–69. дои : 10.1091/mbc.11.5.1555 . ПМК 14867 . ПМИД 10793135 .
- ^ Водермайер Х.К., Гифферс К., Маурер-Штро С., Эйзенхабер Ф., Петерс Дж.М. (сентябрь 2003 г.). «Субъединицы TPR комплекса, способствующего анафазе, опосредуют связывание с белком-активатором CDH1» . Современная биология . 13 (17): 1459–68. дои : 10.1016/s0960-9822(03)00581-5 . ПМИД 12956947 . S2CID 5942532 .
- ^ Мансфельд Дж., Коллин П., Коллинз М.О., Чоудхари Дж.С., Пайнс Дж. (сентябрь 2011 г.). «APC15 управляет оборотом MCC-CDC20, чтобы контрольная точка сборки шпинделя реагировала на прикрепление кинетохор» . Природная клеточная биология . 13 (10): 1234–43. дои : 10.1038/ncb2347 . ПМЦ 3188299 . ПМИД 21926987 .
- ^ «Ген Энтрез: гомолог 27 цикла клеточного деления CDC27 (S. cerevisiae)» .
- ^ Чжан Л., Фудзита Т., Ву Г, Сяо X, Ван Ю (март 2011 г.). «Фосфорилирование комплекса, способствующего анафазе/Cdc27, участвует в передаче сигналов TGF-бета» . Журнал биологической химии . 286 (12): 10041–50. дои : 10.1074/jbc.M110.205518 . ПМК 3060455 . ПМИД 21209074 .
- ^ Лэмб-младший, Мишо В.А., Сикорски Р.С., Хитер П.А. (сентябрь 1994 г.). «Cdc16p, Cdc23p и Cdc27p образуют комплекс, необходимый для митоза» . Журнал ЭМБО . 13 (18): 4321–8. дои : 10.1002/j.1460-2075.1994.tb06752.x . ПМЦ 395359 . ПМИД 7925276 .
- ^ Хуан JY, Рафф JW (июль 2002 г.). «Динамическая локализация APC/C дрозофилы: свидетельства существования множественных комплексов, которые выполняют разные функции и дифференциально локализованы» . Журнал клеточной науки . 115 (Часть 14): 2847–56. дои : 10.1242/jcs.115.14.2847 . ПМИД 12082146 .
- ^ Дик П., Дональдсон М., Гловер Д.М. (октябрь 2003 г.). «Мутации в mákos, гене дрозофилы, кодирующем субъединицу Cdc27 комплекса, способствующего анафазе, усиливают центросомные дефекты в поло и подавляются мутациями у близнецов/аар, которые кодируют регуляторную субъединицу PP2A» . Журнал клеточной науки . 116 (Часть 20): 4147–58. дои : 10.1242/jcs.00722 . ПМИД 12953067 .
- ^ Хартвелл Л.Х., Смит Д. (июль 1985 г.). «Измененная точность передачи митотических хромосом у мутантов клеточного цикла S. cerevisiae» . Генетика . 110 (3): 381–95. дои : 10.1093/генетика/110.3.381 . ПМК 1202570 . ПМИД 3894160 .
- ^ Кинг Р.В., Деше Р.Дж., Питерс Дж.М., Киршнер М.В. (декабрь 1996 г.). «Как протеолиз управляет клеточным циклом». Наука . 274 (5293): 1652–9. Бибкод : 1996Sci...274.1652K . дои : 10.1126/science.274.5293.1652 . ПМИД 8939846 . S2CID 25369228 .
- ^ Крафт С., Водермайер Х.К., Маурер-Штро С., Эйзенхабер Ф., Петерс Дж.М. (май 2005 г.). «Пропеллерный домен WD40 Cdh1 действует как рецептор коробки разрушения для субстратов APC/C» . Молекулярная клетка . 18 (5): 543–53. doi : 10.1016/j.molcel.2005.04.023 . ПМИД 15916961 .
- ^ Рейманн Дж.Д., Фрид Э., Сюй Дж.Ю., Крамер Э.Р., Питерс Дж.М., Джексон П.К. (июнь 2001 г.). «Emi1 является митотическим регулятором, который взаимодействует с Cdc20 и ингибирует комплекс, способствующий анафазе» . Клетка . 105 (5): 645–55. дои : 10.1016/s0092-8674(01)00361-0 . ПМИД 11389834 . S2CID 16366514 .
- ^ Хансен Д.В., Локтев А.В., Бан К.Х., Джексон П.К. (декабрь 2004 г.). «Plk1 регулирует активацию комплекса, способствующего анафазе, путем фосфорилирования и запуска SCFbetaTrCP-зависимого разрушения ингибитора APC Emi1» . Молекулярная биология клетки . 15 (12): 5623–34. дои : 10.1091/mbc.e04-07-0598 . ПМК 532041 . ПМИД 15469984 .
Дальнейшее чтение
[ редактировать ]- Висинтин Р., Принц С., Амон А. (октябрь 1997 г.). «CDC20 и CDH1: семейство субстрат-специфичных активаторов APC-зависимого протеолиза». Наука . 278 (5337): 460–3. Бибкод : 1997Sci...278..460В . дои : 10.1126/science.278.5337.460 . ПМИД 9334304 .
- Сюй Ю., Рейманн Дж. Д., Соренсен К. С., Лукас Дж., Джексон П. К. (май 2002 г.). «E2F-зависимое накопление hEmi1 регулирует вход в S-фазу путем ингибирования APC(Cdh1)». Природная клеточная биология . 4 (5): 358–66. дои : 10.1038/ncb785 . ПМИД 11988738 . S2CID 25403043 .
- Захария В., Нэсмит К. (август 1999 г.). «Чья цель — разрушение: деление клеток и комплекс, способствующий анафазе» . обзор. Гены и развитие . 13 (16): 2039–58. дои : 10.1101/гад.16.13.2039 . ПМИД 10465783 .
- Харпер Дж.В., Бертон Дж.Л., Соломон М.Дж. (сентябрь 2002 г.). «Комплекс, способствующий анафазе: он больше не только для митоза» . обзор. Гены и развитие . 16 (17): 2179–206. дои : 10.1101/gad.1013102 . ПМИД 12208841 .
- Лима М., Элой Н.Б., Пегораро С., Сагит Р., Рохас С., Бретц Т. и др. (ноябрь 2010 г.). «Геномная эволюция и сложность комплекса, способствующего анафазе (APC) у наземных растений» . Биология растений BMC . 10 :254. дои : 10.1186/1471-2229-10-254 . ПМК 3095333 . ПМИД 21087491 .
Внешние ссылки
[ редактировать ]- комплекс, стимулирующий анафазу, в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- 3D-структуры электронной микроскопии комплекса, способствующего анафазе, в Банке данных EM (EMDB)