Глутаминсинтетаза
| глутамат-аммиачная лигаза | |||
|---|---|---|---|
 Активный центр между двумя мономерами глутаминсинтетазы Salmonella typhimurium . Сайты связывания катионов окрашены в желтый и оранжевый цвета; АДФ — розовый; фосфинотрицин имеет синий цвет. [1] | |||
| Идентификаторы | |||
| Номер ЕС. | 6.3.1.2 | ||
| Номер CAS. | 9023-70-5 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| глутаминсинтетаза, бета-домен Grasp | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | Gln-synth_N | ||
| Пфам | PF03951 | ||
| ИнтерПро | ИПР008147 | ||
| PROSITE | PDOC00162 | ||
| СКОП2 | 2глс / СКОПе / СУПФАМ | ||
| |||
| глутаминсинтетаза, каталитический домен | |||
|---|---|---|---|
 12-субъединичный фермент глутаминсинтетаза Salmonella typhimurium . [2] | |||
| Идентификаторы | |||
| Символ | Gln-synt_C | ||
| Пфам | PF00120 | ||
| Пфам Клан | CL0286 | ||
| ИнтерПро | ИПР008146 | ||
| PROSITE | PDOC00162 | ||
| СКОП2 | 2глс / СКОПе / СУПФАМ | ||
| |||
| глутамат-аммиачная лигаза (глутаминсинтетаза) | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ГУЛ | ||
| Альт. символы | ГЛНС | ||
| ген NCBI | 2752 | ||
| HGNC | 4341 | ||
| МОЙ БОГ | 138290 | ||
| ПДБ | 2qc8 | ||
| RefSeq | НМ_002065 | ||
| ЮниПрот | P15104 | ||
| Другие данные | |||
| Номер ЕС | 6.3.1.2 | ||
| Локус | Хр. 1 q31 | ||
| |||
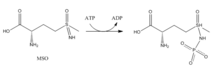
Глутаминсинтетаза ( GS ) ( EC 6.3.1.2 ) [3] Фермент , который играет важную роль в метаболизме азота , катализируя конденсацию глутамата и аммиака с образованием глутамина :
Глутамат + АТФ + NH 3 → Глутамин + АДФ + фосфат

Глутаминсинтетаза использует аммиак, образующийся в результате восстановления нитратов, расщепления аминокислот и фотодыхания . [4] Амидная группа глутамата является источником азота для синтеза метаболитов глутаминового пути . [5]
Другие реакции могут происходить через GS. Конкуренция между ионами аммония и водой, их сродство к связыванию и концентрация ионов аммония влияют на синтез и гидролиз глутамина. Глутамин образуется, если ион аммония атакует промежуточное соединение ацилфосфата, а глутамат перерабатывается, если вода атакует промежуточное соединение. [6] [7] Ион аммония связывается с GS сильнее, чем вода, за счет электростатических сил между катионом и отрицательно заряженным карманом. [4] Другая возможная реакция - при связывании NH 2 OH с GS, а не NH 4 +, образуется γ-глутамилгидроксамат. [6] [7]
Структура
[ редактировать ]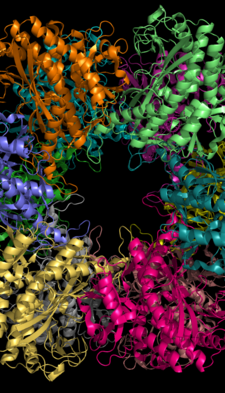
Глутаминсинтетаза может состоять из 8, 10 или 12 идентичных субъединиц, разделенных на два обращенных друг к другу кольца. [6] [8] [9] [10] Бактериальные ГС представляют собой додекамеры с 12 активными центрами между каждым мономером . [6] Каждый активный сайт создает «туннель», который является местом расположения трех различных сайтов связывания субстрата: нуклеотида , иона аммония и аминокислоты. [4] [6] [10] [11] АТФ связывается с вершиной двойной воронки, которая открывается на внешнюю поверхность GS. [4] Глутамат связывается внизу активного центра. [7] В середине биворонки находятся два места связывания двухвалентных катионов (Mn+2 или Mg+2). Один сайт связывания катионов участвует в фосфорильном переносе АТФ на глутамат, а второй стабилизирует активный GS и способствует связыванию глутамата. [6]
Водородные связи и гидрофобные взаимодействия удерживают два кольца GS вместе. Каждая субъединица имеет С-конец и N-конец в своей последовательности. С-конец (спиральный ремешок) стабилизирует структуру GS, встраиваясь в гидрофобную область субъединицы поперек другого кольца. N-конец подвергается воздействию растворителя. Кроме того, центральный канал формируется за счет шести четырехцепочечных β-листов, состоящих из антипараллельных петель двенадцати субъединиц. [6]
Механизм
[ редактировать ]GS катализирует АТФ-зависимую конденсацию глутамата с аммиаком с образованием глутамина. [4] Гидролиз АТФ приводит к [8] первый шаг двухчастного согласованного механизма. [4] [6] АТФ фосфорилирует глутамат с образованием АДФ и промежуточного ацилфосфата, γ-глутамилфосфата, который реагирует с аммиаком, образуя глутамин и неорганический фосфат. АДФ и P i не диссоциируют до тех пор, пока не свяжется аммиак и не высвободится глутамин. [6]
АТФ сначала связывается с верхней частью активного сайта рядом с сайтом связывания катионов, тогда как глутамат связывается со вторым сайтом связывания катионов в нижней части активного сайта. [5] [7] Присутствие АДФ вызывает конформационный сдвиг GS, который стабилизирует фрагмент γ-глутамилфосфата. Аммоний прочно связывается с GS только в том случае, если присутствует промежуточный ацилфосфат. Аммоний, а не аммиак, связывается с GS, поскольку сайт связывания полярен и подвергается воздействию растворителя. [7] На втором этапе депротонирование аммония позволяет аммиаку атаковать промежуточное соединение из близлежащего участка с образованием глутамина. [12] Фосфат уходит через верхнюю часть активного центра, а глютамин — через нижнюю часть (между двумя кольцами). [13] [7]

Биологическая функция
[ редактировать ]GS присутствует преимущественно в головном мозге, почках и печени. [4] [10] ГС в головном мозге участвует в метаболической регуляции глутамата, детоксикации аммиака головного мозга, ассимиляции аммиака, рециклизации нейротрансмиттеров и терминации сигналов нейромедиаторов. [4] [14] GS в головном мозге обнаруживается преимущественно в астроцитах . [15] Астроциты защищают нейроны от эксайтотоксичности, поглощая избыток аммиака и глутамата. [14] В гипераммониемической среде (высокий уровень аммиака) возникает отек астроглии. [14] [16] [17] К проблеме набухания астроглии подошли разные точки зрения. Одно исследование показывает, что происходят морфологические изменения, которые увеличивают экспрессию GS в глутаматергических областях или другие адаптации, которые снижают высокие уровни глутамата и аммиака. [14] Другая точка зрения заключается в том, что набухание астроцитов происходит из-за накопления глутамина. Чтобы предотвратить повышение уровня коркового глютамата и содержания кортикальной воды, было проведено исследование по предотвращению активности GS у крыс с помощью MSO. [16]
Классы
[ редактировать ]Кажется, существует три разных класса GS: [18] [19] [20]
- Ферменты класса I (GSI) специфичны для прокариот и представляют собой олигомеры из 12 идентичных субъединиц . [21] Активность фермента GSI-типа контролируется аденилированием остатка тирозина . Аденилированный фермент неактивен. [22]
- Ферменты класса II (GSII) обнаружены у эукариот и бактерий, принадлежащих к семействам Rhizobiaceae , Frankiaceae и Streptomycetaceae (эти бактерии также имеют GS класса I). ГСИИ представляют собой декамеры идентичных субъединиц. [10] ВВП : 2OJW .
Растения имеют два и более изоферментов GSII, один из изоферментов транслоцируется в хлоропласт . Другая форма — цитозольная . Трансляция цитозольного гена GS регулируется его 5'-нетранслируемой областью (UTR), тогда как его 3'-UTR играет роль в обмене транскриптов. [23]
- Ферменты класса III (GSIII) в настоящее время обнаружены только у Bacteroides fragilis и Butyrivibrio fibrisolvens . Это двухкольцевой додекамер из одинаковых цепочек. [24] Он намного больше (около 700 аминокислот), чем ферменты GSI (от 450 до 470 аминокислот) или GSII (от 350 до 420 аминокислот).
Хотя три класса GS явно структурно родственны, сходство последовательностей не столь велико.
Регулирование и ингибирование
[ редактировать ]GS подвержен обратимой ковалентной модификации. Тир 397 из всех 12 субъединиц могут подвергаться аденилированию или деаденилированию с помощью аденилаттрансферазы (АТ), бифункционального регуляторного фермента. [25] Аденилирование — это посттрансляционная модификация, включающая ковалентное присоединение АМФ к боковой цепи белка. Для каждого аденилирования требуется АТФ , а для полного ингибирования GS требуется 12 АТФ. Деаденилирование с помощью AT включает фосфоролитическое удаление Tyr-связанных аденилильных групп в виде ADP . На активность АТ влияет связанный с ней регуляторный белок: P II массой 44 кДа , тример . [25] P II также подвергается посттрансляционной модификации уридилилтрансферазой , поэтому P II имеет две формы. Состояние P II диктует активность аденилаттрансферазы. Если P II не уридилирован, то он примет форму P IIA . Комплекс AT:P IIA дезактивирует GS путем аденилирования. Если P II уридилирован, то он примет форму P IID . Комплекс AT:P IID активирует GS путем деаденилирования. [25] AT:P IIA и AT:P IID Комплексы аллостерически регулируются реципрокным образом с помощью α-кетоглутарата (α-KG) и глутамина (Gln). Gln активирует активность AT:P IIA и ингибирует AT:P IID , что приводит к аденилированию и последующей дезактивации GS. Кроме того, Gln выступает за преобразование P IID в P IIA . Эффекты α-KG на комплексы противоположны. [25] У большинства грамотрицательных бактерий GS может модифицироваться путем аденилирования (некоторые цианобактерии и зеленые водоросли или исключения). [26]
Ингибирование GS в основном сосредоточено на лигандах аминосайтов. [6] Другие ингибиторы являются результатом метаболизма глютамина: триптофан, гистидин, карбамоилфосфат, глюкозамин-6-фосфат, цитидинтрифосфат (ЦТФ) и аденозинмонофосфат (АМФ). [5] [8] [27] Другими ингибиторами/регуляторами являются глицин и аланин. Аланин, глицин и серин связываются с субстратом глутамата. GDP, AMP, ADP связываются с сайтом АТФ. [6] L-серин, L-аланин и глицин связываются с сайтом L-глутамата в неаденилированном GS. Четыре аминокислоты связываются с сайтом своими общими атомами, «основной цепью» аминокислот. [5] Глутамат — еще один продукт метаболизма глютамина; однако глутамат является субстратом для GS, ингибируя его действие в качестве регулятора GS.2 Каждый ингибитор может снижать активность фермента; как только все конечные метаболиты глютамина связываются с GS, активность GS почти полностью ингибируется. [8] Многие тормозные входные сигналы позволяют точно настроить GS, отражая уровень азота в организме.
Регуляция по принципу обратной связи различает два эукариотических типа ГС: мозговые и немозговые ткани. Немозговой GS реагирует на ингибирование обратной связи конечного продукта, а мозговой GS - нет. [6] Высокие концентрации глутамин-зависимых метаболитов должны ингибировать активность GS, тогда как низкие концентрации должны активировать активность GS. [6]
Ингибиторы:
- Метионинсульфоксимин (МСО): МСО является ингибитором, который связывается с сайтом глутамата. Связанный с GS, MSO фосфорилируется АТФ, что приводит к необратимому нековалентному ингибированию GS. Конфигурация S-изомера является более ингибирующей. [6] Вход глутамата в активный сайт блокируется за счет стабилизации гибкой петли в активном сайте с помощью MSO. [7]
- фосфинотрицин [1] (PPT, глюфосинат): фосфинотрицин является ингибитором, который связывается с глутаматным участком. Глюфосинат используется как гербицид. Растения, обработанные глюфосинатом, погибают из-за накопления аммиака и прекращения фотосинтеза. [10]
- Сегодня доступно множество синтетических ингибиторов. [6]
Исследования E. coli показали, что GS регулируется посредством экспрессии генов. Ген, кодирующий субъединицу GS, обозначается glnA . Транскрипция glnA зависит от NR I (специфического усилителя транскрипции ). Активная транскрипция происходит, если NR I находится в фосфорилированной форме, обозначенной NR I -P . Фосфорилирование NR I катализируется NR , протеинкиназой II . Если NR II образует комплекс с P IIA то он будет функционировать как фосфатаза , и NR I -P преобразуется обратно в NR I. , В этом случае транскрипция glnA прекращается. [25]
GS подчиняется совершенно другим регуляторным механизмам У цианобактерий . [28] Вместо общей двухкомпонентной системы NtrC-NtrB, [29] [30] цианобактерии содержат регулятор транскрипции NtcA, который ограничен этой кладой и контролирует экспрессию GS и множества генов, участвующих в азотистом метаболизме. [31] [32] Более того, GS у цианобактерий не модифицируется ковалентно, чтобы повысить чувствительность к ингибированию по принципу обратной связи. [30] Вместо этого GS у цианобактерий ингибируется небольшими белками, называемыми факторами, инактивирующими GS (IF), транскрипция которых отрицательно регулируется NtcA. [33] [34] Эти инактивирующие факторы, кроме того, регулируются различными некодирующими РНК : мРНК NsiR4 взаимодействует с 5'UTR мРНК инактивирующего фактора GS IF7 ( мРНК gifA ) и снижает его экспрессию. Экспрессия NsiR4 находится под положительным контролем фактора транскрипции NtcA, контролирующего азот. [35] Кроме того, экспрессия фактора инактивации GS IF17 контролируется глутамин-связывающим рибопереключателем . [36]
Ссылки
[ редактировать ]- ^ Jump up to: а б с ВВП : 1 FPY ; Гилл Х.С., Айзенберг Д. (февраль 2001 г.). «Кристаллическая структура фосфинотрицина в активном центре глутаминсинтетазы проливает свет на механизм ферментативного ингибирования». Биохимия . 40 (7): 1903–12. дои : 10.1021/bi002438h . ПМИД 11329256 .
- ^ ПДБ : 2ГЛС ; Ямашита М.М., Алмасси Р.Дж., Янсон К.А., Касио Д., Айзенберг Д. (октябрь 1989 г.). «Уточненная атомная модель глутаминсинтетазы с разрешением 3,5 А». Ж. Биол. Хим . 264 (30): 17681–90. дои : 10.2210/pdb2gls/pdb . ПМИД 2572586 .
- ^ Айзенберг Д., Алмасси Р.Дж., Янсон К.А., Чепмен М.С., Су С.В., Касио Д., Смит В.В. (1987). «Некоторые эволюционные связи первичных биологических катализаторов глутаминсинтетазы и RuBisCO». Холодный источник Харб. Симп. Квант. Биол . 52 : 483–90. дои : 10.1101/sqb.1987.052.01.055 . ПМИД 2900091 .
- ^ Jump up to: а б с д и ж г час Лиав Ш., Куо И., Айзенберг Д. (ноябрь 1995 г.). «Открытие сайта аммониевого субстрата на глутаминсинтетазе, третьего сайта связывания катионов» . Белковая наука . 4 (11): 2358–65. дои : 10.1002/pro.5560041114 . ПМК 2143006 . ПМИД 8563633 .
- ^ Jump up to: а б с д Лиав Ш., Пан С., Айзенберг Д. (июнь 1993 г.). «Ингибирование полностью аденилированной глутаминсинтетазы Salmonella typhimurium по принципу обратной связи с помощью глицина, аланина и серина» . Учеб. Натл. акад. наук. США . 90 (11): 4996–5000. Бибкод : 1993PNAS...90.4996L . дои : 10.1073/pnas.90.11.4996 . ПМК 46640 . ПМИД 8099447 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Айзенберг Д., Гилл Х.С., Пфлюгль Г.М., Ротштейн С.Х. (март 2000 г.). «Структурно-функциональные взаимоотношения глутаминсинтетаз» . Биохим Биофиз Акта . 1477 (1–2): 122–45. дои : 10.1016/S0167-4838(99)00270-8 . ПМИД 10708854 .
- ^ Jump up to: а б с д и ж г Лиав Ш., Айзенберг Д. (январь 1994 г.). «Структурная модель механизма реакции глутаминсинтетазы, основанная на пяти кристаллических структурах фермент-субстратных комплексов». Биохимия . 33 (3): 675–81. дои : 10.1021/bi00169a007 . ПМИД 7904828 .
- ^ Jump up to: а б с д Страйер Л., Берг Дж.М., Тимочко Дж.Л. (2007). Биохимия (6-е изд.). Сан-Франциско: WH Freeman. стр. 679–706 . ISBN 978-0-7167-8724-2 .
- ^ Гудселл DS (июнь 2002 г.). «Глютаминсинтетаза» . Молекула месяца . Банк данных белков RCSB. Архивировано из оригинала 31 мая 2008 г. Проверено 8 мая 2010 г.
- ^ Jump up to: а б с д и Краевски В.В., Коллинз Р., Холмберг-Шьявоне Л., Джонс Т.А., Карлберг Т., Моубрей С.Л. (январь 2008 г.). «Кристаллические структуры глутаминсинтетаз млекопитающих иллюстрируют конформационные изменения, вызванные субстратом, и открывают возможности для разработки лекарств и гербицидов» . Дж Мол Биол . 375 (1): 317–28. дои : 10.1016/j.jmb.2007.10.029 . ПМИД 18005987 .
- ^ Гинзбург А., Йе Дж., Хенниг С.Б., Дентон, доктор медицины (февраль 1970 г.). «Некоторые эффекты аденилирования на биосинтетические свойства глутаминсинтетазы Escherichia coli». Биохимия . 9 (3): 633–49. дои : 10.1021/bi00805a025 . ПМИД 4906326 .
- ^ Хант Дж.Б., Смирниотис П.З., Гинзбург А., Штадтман Э.Р. (январь 1975 г.). «Потребность в ионах металлов глютаминсинтетазой Escherichia coli в катализе переноса гамма-глутамила». Арх Биохим Биофиз . 166 (1): 102–24. дои : 10.1016/0003-9861(75)90370-7 . ПМИД 235885 .
- ^ Гудселл, DS (июнь 2002 г.). «Глютаминсинтетаза» . Банк данных белков RCSB. Архивировано из оригинала 31 мая 2008 года . Проверено 8 мая 2010 г.
- ^ Jump up to: а б с д Суарес И., Бодега Г., Фернандес Б. (август – сентябрь 2002 г.). «Глутаминсинтетаза в мозге: влияние аммиака». Нейрохим. Межд . 41 (2–3): 123–42. дои : 10.1016/S0197-0186(02)00033-5 . ПМИД 12020613 . S2CID 24661063 .
- ^ Венкатеш К., Шрикант Л., Венгамма Б., Чандрасекхар С., Сандживкумар А., Мулешвара Прасад Б.К., Сарма П.В. (2013). «Дифференцировка in vitro культивируемых клеток CD34+ человека в астроциты» . Нейрол Индия . 61 (4): 383–8. дои : 10.4103/0028-3886.117615 . ПМИД 24005729 .
- ^ Jump up to: а б Уиллард-Мак CL, Келер RC, Хирата Т и др. (март 1996 г.). «Ингибирование глутаминсинтетазы уменьшает вызванное аммиаком набухание астроцитов у крыс». Нейронаука . 71 (2): 589–99. дои : 10.1016/0306-4522(95)00462-9 . ПМИД 9053810 . S2CID 31674240 .
- ^ Танигами Х, Ребел А, Мартин Л.Дж., Чен Т.Ю., Брусилов С.В., Трайстман Р.Дж., Келер Р.К. (2005). «Влияние ингибирования глутаминсинтетазы на набухание астроцитов и изменение экспрессии астроглиального белка во время гипераммониемии у крыс» . Нейронаука . 131 (2): 437–49. doi : 10.1016/j.neuroscience.2004.10.045 . ПМК 1819407 . ПМИД 15708485 .
- ^ Кумада Ю., Бенсон Д.Р., Хиллеманн Д., Хостинг Т.Дж., Рошфор Д.А., Томпсон С.Дж., Воллебен В., Татено Ю. (апрель 1993 г.). «Эволюция гена глутаминсинтетазы, одного из старейших существующих и функционирующих генов» . Учеб. Натл. акад. наук. США . 90 (7): 3009–13. Бибкод : 1993PNAS...90.3009K . дои : 10.1073/pnas.90.7.3009 . ПМЦ 46226 . ПМИД 8096645 .
- ^ Шаттерс Р.Г., Кан М.Л. (ноябрь 1989 г.). «Глутаминсинтетаза II в Rhizobium: пересмотр предполагаемого горизонтального переноса ДНК от эукариот к прокариотам». Дж. Мол. Эвол . 29 (5): 422–8. Бибкод : 1989JMolE..29..422S . дои : 10.1007/BF02602912 . ПМИД 2575672 . S2CID 36704558 .
- ^ Браун-младший, Масучи Й., Робб Ф.Т., Дулиттл В.Ф. (июнь 1994 г.). «Эволюционные взаимоотношения генов бактериальной и архейной глутаминсинтетазы». Дж. Мол. Эвол . 38 (6): 566–76. Бибкод : 1994JMolE..38..566B . дои : 10.1007/BF00175876 . ПМИД 7916055 . S2CID 21493521 .
- ^ «Структура ГСИ» . Архивировано из оригинала 17 декабря 2008 г. Проверено 31 марта 2009 г.
- ^ InterPro: IPR001637 Глутаминсинтетаза класса I, сайт аденилирования
- ^ Ортега Дж.Л., Уилсон О.Л., Сенгупта-Гопалан С. (декабрь 2012 г.). «5'-нетранслируемая область гена цитозольной глутаминсинтетазы β(1) сои содержит сигналы инициации трансляции прокариот и действует как усилитель трансляции в растениях» . Молекулярная генетика и геномика . 287 (11–12): 881–93. дои : 10.1007/s00438-012-0724-6 . ПМЦ 3881598 . ПМИД 23080263 .
- ^ ван Ройен Дж. М., Абратт В. Р., Сьюэлл Б. Т. (август 2006 г.). «Трехмерная структура глутаминсинтетазы III типа путем одночастичной реконструкции». Дж. Мол. Биол . 361 (4): 796–810. дои : 10.1016/j.jmb.2006.06.026 . hdl : 11394/1617 . ПМИД 16879836 .
- ^ Jump up to: а б с д и Гаррет, Гришэм (2017). Биохимия (6-е изд.). Соединенные Штаты Америки: Cengage Learning. стр. 886–889. ISBN 978-1-305-57720-6 .
- ^ Ивановский Р.Н., Хатипов Е.А. (1994). «Доказательства ковалентной модификации глутаминсинтетазы у пурпурной серной бактерии» . Письма FEMS по микробиологии . 122 (1–2): 115–119. дои : 10.1111/j.1574-6968.1994.tb07153.x .
- ^ Кришнан И.С., Сингхал Р.К., Дуа Р.Д. (апрель 1986 г.). «Очистка и характеристика глутаминсинтетазы Clostridium Pasteurianum». Биохимия . 25 (7): 1589–99. дои : 10.1021/bi00355a021 . ПМИД 2871863 .
- ^ Болай П., Муро-Пастор М., Флоренсио Ф., Клен С. (27 октября 2018 г.). «Особенная регуляция цианобактериальной глутаминсинтетазы» . Жизнь . 8 (4): 52. Бибкод : 2018Жизнь....8...52Б . дои : 10.3390/life8040052 . ПМК 6316151 . ПМИД 30373240 .
- ^ Меррик М.Дж., Эдвардс Р.А. (декабрь 1995 г.). «Контроль азота у бактерий» . Микробиологические обзоры . 59 (4): 604–22. дои : 10.1128/MR.59.4.604-622.1995 . ПМК 239390 . ПМИД 8531888 .
- ^ Jump up to: а б Фишер Р., Тули Р., Хазелькорн Р. (июнь 1981 г.). «Клонированный цианобактериальный ген глутаминсинтетазы функционирует в Escherichia coli, но фермент не аденилирован» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (6): 3393–7. Бибкод : 1981PNAS...78.3393F . дои : 10.1073/pnas.78.6.3393 . ПМК 319574 . ПМИД 6115380 .
- ^ Вега-Палас М.А., Флорес Э., Эрреро А. (июль 1992 г.). «NtcA, глобальный регулятор азота из цианобактерии Synechococcus, принадлежащей к семейству бактериальных регуляторов Crp». Молекулярная микробиология . 6 (13): 1853–9. дои : 10.1111/j.1365-2958.1992.tb01357.x . ПМИД 1630321 . S2CID 32757978 .
- ^ Рейес Дж.К., Муро-Пастор М.И., Флоренсио Ф.Дж. (апрель 1997 г.). «Транкрипция генов глутаминсинтетазы (glnA и glnN) из штамма цианобактерии Synechocystis sp. PCC 6803 по-разному регулируется в ответ на доступность азота» . Журнал бактериологии . 179 (8): 2678–89. дои : 10.1128/jb.179.8.2678-2689.1997 . ПМК 179018 . ПМИД 9098067 .
- ^ Гарсиа-Домингес М., Рейес Х.К., Флоренсио Ф.Дж. (июнь 1999 г.). «Инактивация глутаминсинтетазы путем белок-белкового взаимодействия» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (13): 7161–6. Бибкод : 1999PNAS...96.7161G . дои : 10.1073/pnas.96.13.7161 . ПМК 22038 . ПМИД 10377385 .
- ^ Гарсиа-Домингес М., Рейес Х.К., Флоренсио Ф.Дж. (март 2000 г.). «NtcA подавляет транскрипцию gifA и gifB, генов, которые кодируют ингибиторы глутаминсинтетазы типа I из Synechocystis sp. PCC 6803». Молекулярная микробиология . 35 (5): 1192–201. дои : 10.1046/j.1365-2958.2000.01789.x . ПМИД 10712699 . S2CID 23804565 .
- ^ Клен С., Шаал С., Георг Дж., Баумгартнер Д., Книппен Г., Хагеманн М., Муро-Пастор А.М., Хесс В.Р. (ноябрь 2015 г.). «МрРНК NsiR4 участвует в контроле ассимиляции азота у цианобактерий путем воздействия на фактор IF7, инактивирующий глутаминсинтетазу» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (45): Е6243-52. Бибкод : 2015PNAS..112E6243K . дои : 10.1073/pnas.1508412112 . ПМЦ 4653137 . ПМИД 26494284 .
- ^ Клен С., Болай П., Райт П.Р., Атильо Р.М., Брюэр К.И., Хагеманн М., Брейкер Р.Р., Хесс В.Р. (август 2018 г.). «Глутаминовый рибопереключатель является ключевым элементом регуляции глутаминсинтетазы у цианобактерий» . Исследования нуклеиновых кислот . 46 (19): 10082–10094. дои : 10.1093/nar/gky709 . ПМК 6212724 . ПМИД 30085248 .
Внешние ссылки
[ редактировать ]- Запись ИнтерПро
- Молекула месяца RCSB PDB
- PDBe-KB предоставляет обзор всей информации о структуре, доступной в PDB, для глутаминсинтетазы человека.