Фенилаланингидроксилаза
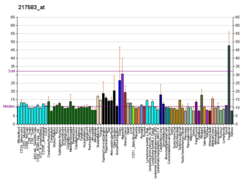
| ПАУ | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | PAH , PH, PKU, PKU1, фенилаланингидроксилаза | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 612349 ; МГИ : 97473 ; Гомологен : 234 ; GeneCards : ПАУ ; ОМА : ПАУ – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





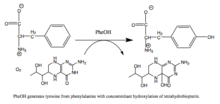
Фенилаланингидроксилаза ( ПАУ ) ( EC 1.14.16.1 ) представляет собой фермент , который катализирует гидроксилирование ароматической боковой цепи фенилаланина с образованием тирозина . ПАУ является одним из трех членов биоптерин -зависимых гидроксилаз ароматических аминокислот , класса монооксигеназ , которые используют тетрагидробиоптерин (BH 4 , птеридиновый кофактор) и негемовое железо для катализа. В ходе реакции молекулярный кислород гетеролитически расщепляется с последовательным включением одного атома кислорода в BH 4 и фенилаланиновый субстрат. [5] [6] У людей мутации в кодирующем его гене PAH могут привести к метаболическому расстройству фенилкетонурии .
 |
Ферментативный механизм
[ редактировать ]Предполагается, что реакция протекает в следующие стадии:
- образование мостика Fe(II)-OO-BH 4 .
- гетеролитический разрыв связи OO с образованием феррилоксогидроксилирующего промежуточного соединения Fe(IV)=O
- атака Fe(IV)=O с образованием гидроксила фенилаланинового субстрата до тирозина. [7]

Формирование и расщепление железо-пероксиптеринового мостика. Хотя данные убедительно подтверждают, что Fe(IV)=O является промежуточным продуктом гидроксилирования, [8] детали механизма, лежащие в основе образования мостика Fe(II)-OO-BH 4 перед гетеролитическим расщеплением, остаются спорными. Два пути были предложены на основе моделей, которые различаются близостью железа к кофактору птерина и количеством молекул воды, которые, как предполагается, координируются железом во время катализа. Согласно одной из моделей, первоначально образуется и стабилизируется дикислородный комплекс железа как резонансный гибрид Fe. 2+ О 2 и Fe 3+ Около 2 − . Активированный O 2 затем атакует BH 4 , образуя переходное состояние , характеризующееся разделением зарядов между электронодефицитным птериновым кольцом и богатыми электронами дикислородными частицами. [9] Fe(II)-OO-BH 4 Впоследствии образуется мостик . С другой стороны, образование этого мостика было смоделировано в предположении, что BH4 расположен в первой координационной сфере железа и что железо не координировано ни с какими молекулами воды. Эта модель предсказывает другой механизм, включающий радикал птерина и супероксид в качестве критических промежуточных продуктов. [10] После образования мостик Fe(II)-OO-BH 4 разрывается за счет гетеролитического разрыва связи ОО до Fe(IV)=O и 4а-гидрокситетрагидробиоптерина; таким образом, молекулярный кислород является источником как атомов кислорода, используемых для гидроксилирования птеринового кольца, так и фенилаланина.
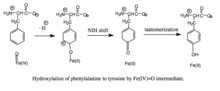
Гидроксилирование фенилаланина феррилоксо-интермедиатом. Поскольку механизм включает в себя промежуточное гидроксилирование Fe(IV)=O (в отличие от пероксиптерина), окисление кофактора BH 4 и гидроксилирование фенилаланина могут быть разделены, что приводит к непроизводительному потреблению BH 4 и образованию H 2 O 2 . [7] Однако при продуктивности промежуточное соединение Fe (IV) = O добавляется к фенилаланину в реакции электрофильного ароматического замещения, которая восстанавливает железо из феррильного состояния в двухвалентное состояние. [7] Хотя первоначально был предложен ареноксид или радикальный промежуточный продукт, анализ родственных триптофан- и тирозингидроксилаз показал, что вместо этого реакция протекает через катионный промежуточный продукт, который требует координации Fe(IV)=O с водным лигандом, а не с гидроксогруппой. . [7] [11] Этот катионный промежуточный продукт впоследствии подвергается 1,2-гидридному сдвигу NIH, в результате чего образуется диеноновый промежуточный продукт, который затем таутомеризуется с образованием тирозинового продукта. [12] Кофактор птерина регенерируется путем гидратации карбиноламинового продукта ПАУ до хиноноидного дигидробиоптерина (qBH 2 ), который затем восстанавливается до BH 4 . [13]
Регуляция ферментов
[ редактировать ]Для ФАГ предложено использовать морфеиновую модель аллостерической регуляции . [14] [15]
ПАУ млекопитающих существует в равновесии, состоящем из тетрамеров двух различных архитектур, с одной или несколькими димерными формами как часть равновесия. Такое поведение согласуется с диссоциативным аллостерическим механизмом. [15]
Многие исследования показывают, что ПАУ млекопитающих демонстрирует поведение, сравнимое с поведением порфобилиногенсинтазы (PBGS), при этом, как сообщается, различные факторы, такие как pH и связывание лиганда, влияют на активность фермента и стабильность белка. [15]
Структура
[ редактировать ]Мономер PAH (51,9 кДа) состоит из трех отдельных доменов: регуляторного N-концевого домена (остатки 1–117), который содержит Phe-связывающий субдомен ACT, каталитического домена (остатки 118–427) и C-концевого домена. (остатки 428–453), ответственные за олигомеризацию идентичных мономеров. Был проведен обширный кристаллографический анализ, особенно каталитического домена, координируемого птерином и железом, для изучения активного центра. Также была определена структура N-концевого регуляторного домена и вместе с решенной структурой гомологичного С-концевого тетрамеризационного домена тирозингидроксилазы предложена структурная модель тетрамерного ПАУ. [13] С помощью рентгеновской кристаллографии структура полноразмерного крысиного ПАУ была определена экспериментально и показала автоингибированную форму фермента или форму покоя. [16] Форма состояния покоя (RS-PAH) архитектурно отличается от активированной формы (A-PAH). [17] Полноразмерная структура A-PAH в настоящее время отсутствует, но стабилизированная Phe граница раздела ACT-ACT была определена, что характерно для A-PAH, и была предложена структурная модель A-PAH, основанная на анализе SAXS. [18] [19]
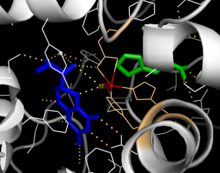
Каталитический домен
[ редактировать ]Раскрытые кристаллические структуры каталитического домена показывают, что активный центр состоит из открытого и просторного кармана, выстланного в основном гидрофобными остатками, хотя также присутствуют три остатка глутаминовой кислоты, два гистидина и тирозин, связывающие железо. [13] Существуют противоречивые данные о координационном состоянии атома железа и его близости к BH4 внутри активного центра. По данным кристаллографического анализа, Fe(II) координируется водой, His285, His290 и Glu330 (граневая триада 2-гис-1-карбоксилата) с октаэдрической геометрией. [20] Включение аналога Phe в кристаллическую структуру изменяет как железо из шести- в пятикоординированное состояние, включающее одну молекулу воды и бидентатную координацию с Glu330, так и открытие места для связывания кислорода. BH4 при этом смещается в сторону атома железа, хотя кофактор птерина остается во второй координационной сфере. [21] С другой стороны, конкурирующая модель, основанная на анализе ЯМР и молекулярного моделирования, предполагает, что все координированные молекулы воды вытесняются из активного центра во время каталитического цикла, в то время как BH4 становится непосредственно координированным с железом. [22] Как обсуждалось выше, разрешение этого несоответствия будет важно для определения точного механизма катализа ПАУ.
N-концевой регуляторный домен
[ редактировать ]Регуляторная природа N-концевого домена (остатки 1–117) обусловлена его структурной гибкостью. [23] Анализ обмена водорода/дейтерия показывает, что аллостерическое связывание Phe глобально изменяет конформацию ПАУ, так что активный центр менее закупорен, поскольку граница раздела между регуляторным и каталитическими доменами все больше подвергается воздействию растворителя. [23] [24] [25] Это наблюдение согласуется с кинетическими исследованиями, которые показывают изначально низкую скорость образования тирозина для полноразмерных ПАУ. Однако это время задержки не наблюдается для укороченного ПАУ, лишенного N-концевого домена, или если полноразмерный фермент предварительно инкубируется с Phe. Удаление N-концевого домена также устраняет время задержки, одновременно увеличивая сродство к Phe почти в два раза; никакой разницы в V max или K m для кофактора тетрагидробиоптерина не наблюдается. [26] Дополнительную регуляцию обеспечивает Ser16; фосфорилирование этого остатка не меняет конформацию фермента, но снижает концентрацию Phe, необходимую для аллостерической активации. [25] Этот N-концевой регуляторный домен не наблюдается в бактериальных ПАУ, но демонстрирует значительную структурную гомологию с регуляторным доменом фосфогилцератдегидрогеназы, фермента пути биосинтеза серина. [25]
Домен тетрамеризации
[ редактировать ]Прокариотический ПАУ является мономерным, тогда как эукариотический ПАУ существует в равновесии между гомотетрамерной и гомодимерной формами. [7] [13] Интерфейс димеризации состоит из связанных по симметрии петель, которые связывают идентичные мономеры, в то время как перекрывающийся С-концевой домен тетрамеризации опосредует ассоциацию конформационно различных димеров, которые характеризуются различной относительной ориентацией каталитического и тетрамеризационного доменов (Flatmark, Erlandsen). Возникающее в результате нарушение симметрии тетрамера проявляется в различной площади поверхности границ раздела димеризации и отличает ПАУ от тетрамерно-симметричной тирозингидроксилазы. [13] Был предложен механизм замены доменов, опосредующий образование тетрамера из димеров, в котором С-концевые альфа-спирали взаимно изменяют свою конформацию вокруг гибкой С-концевой шарнирной области с пятью остатками, образуя структуру спиральной спирали, смещая равновесие. в сторону тетрамерной формы. [7] [13] [27] Хотя как гомодимерная, так и гомотетрамерная формы ПАУ каталитически активны, они обладают различной кинетикой и регуляцией. Помимо сниженной каталитической эффективности, димер не проявляет положительной кооперативной активности по отношению к L-Phe (который при высоких концентрациях активирует фермент), что позволяет предположить, что L-Phe аллостерически регулирует ФАГ, влияя на взаимодействие димер-димер. [27]
Биологическая функция
[ редактировать ]ПАУ является важнейшим ферментом в метаболизме фенилаланина и катализирует лимитирующую стадию его полного катаболизма до углекислого газа и воды. [13] [28] Регулирование потока через связанные с фенилаланином пути имеет решающее значение для метаболизма млекопитающих, о чем свидетельствует токсичность высоких уровней этой аминокислоты в плазме, наблюдаемая при фенилкетонурии (см. ниже). Основным источником фенилаланина являются поступающие в организм белки, но относительно небольшая часть этого пула используется для синтеза белка. [28] Вместо этого большая часть поступившего в организм фенилаланина катаболизируется через ПАУ с образованием тирозина ; добавление гидроксильной группы позволяет разорвать бензольное кольцо на последующих стадиях катаболизма. Трансаминирование до фенилпирувата , метаболиты которого выводятся с мочой, представляет собой еще один путь обмена фенилаланина, но преобладает катаболизм через ПАУ. [28]
У человека этот фермент экспрессируется как в печени, так и в почках, и есть некоторые признаки того, что в этих тканях он может по-разному регулироваться. [29] ПАУ необычен среди гидроксилаз ароматических аминокислот своим участием в катаболизме; С другой стороны, тирозин- и триптофангидроксилазы экспрессируются преимущественно в центральной нервной системе и катализируют этапы, ограничивающие скорость биосинтеза нейромедиаторов/гормонов. [13]
Актуальность заболевания
[ редактировать ]Дефицит активности ФАГ из-за мутаций ФАГ вызывает гиперфенилаланинемию (ГФА), а когда уровень фенилаланина в крови увеличивается более чем в 20 раз по сравнению с нормальной концентрацией, возникает метаболическое заболевание фенилкетонурия (ФКУ). [28] ФКУ является как генотипически, так и фенотипически гетерогенной: было идентифицировано более 300 различных патогенных вариантов, большинство из которых соответствуют миссенс-мутациям, которые картируются в каталитическом домене. [13] [20] Когда группа идентифицированных мутантов ФАГ экспрессировалась в рекомбинантных системах, ферменты проявляли измененное кинетическое поведение и/или пониженную стабильность, что согласуется со структурным картированием этих мутаций как в каталитическом, так и в тетрамеризационном доменах фермента. [13] BH4 4 применялся в качестве фармакологического лечения, и было показано, что он снижает уровень фенилаланина в крови у части пациентов с фенилкетонурией, чьи генотипы приводят к некоторой остаточной активности ЛАГ, но не имеют дефектов в синтезе или регенерации BH4 4 . Последующие исследования показывают, что в случае некоторых мутантов PAH избыток BH4 4 действует как фармакологический шаперон для стабилизации мутантных ферментов с нарушенной сборкой тетрамеров и повышенной чувствительностью к протеолитическому расщеплению и агрегации. [30] Мутации, выявленные в локусе PAH, задокументированы в Базе знаний локуса фенилаланингидроксилазы (PAHdb, https://web.archive.org/web/20130718162051/http://www.pahdb.mcgill.ca/ ).
Поскольку фенилкетонурия может вызвать необратимые повреждения, крайне важно выявить дефицит фенилаланингидроксилазы на ранних стадиях развития. Первоначально это делалось с использованием анализа ингибирования бактерий, известного как тест Гатри . В настоящее время ФКУ является частью скрининга новорожденных во многих странах, а повышенные уровни фенилаланина выявляются вскоре после рождения путем измерения с помощью тандемной масс-спектрометрии . Помещение человека на диету с низким содержанием фенилаланина и высоким содержанием тирозина может помочь предотвратить любой долгосрочный ущерб его развитию.
Модель нокаута ПАУ
[ редактировать ]О первой попытке создания модели мыши Pah -KO сообщалось в исследовательской статье, опубликованной в 2021 году. [31] Эта нокаутная мышь была создана как гомозиготная путем ее развития в штамме C57BL/6 J с использованием CRISPR / Cas9 . [32] Кодон 7, GAG, в гене Pah был заменен на стоп-кодон TAG, что указывает на преднамеренную точечную мутацию . Гомозиготных мышей мужского пола в возрасте от двух до шести месяцев изучали с помощью научных методов, таких как поведенческие и биохимические анализы, МРТ и гистопатология. Гомозиготных мышей обычно сравнивали с контрольными мышами, гетерозиготными мышами Pah -KO, соответствующими по возрасту и полу, которые экспрессировали достаточную активность фермента PAH для обеспечения уровней фенилаланина и тирозина, аналогичных мышам дикого типа .
Мышиная модель Pah -KO демонстрировала высокий уровень фенилаланина в крови и низкий уровень тирозина, гипохолестеринемию , высокий уровень фенилаланина и низкий уровень нейротрансмиттеров и тирозина в мозге, гипомиелинизацию, низкую массу мозга и тела, высокую степень глазной патологии, гипопигментацию и прогрессирующее поведенческое поведение. дефицит по сравнению с гетерозиготными мышами. [31] После уничтожения гомозиготных мышей с помощью вестерн-блоттинга белки печени ФАГ не были обнаружены . [33] Повышенное количество фенилаланина в гомогенатах цельного мозга гомозиготных мышей было сходным с таковым у пациентов с ФКУ. Цвет шерсти обеих мышей зависел от количества пигмента меланина в стержнях волос; таким образом, гомозиготные мыши были склонны иметь более светло-коричневый цвет из-за меньшего количества меланина. Поведение мышей проверяли с помощью анализа гнездового строительства. [31]
Эта модель открывает возможности для обширного исследования биологии ФКУ, напоминающей пациентов с ФКУ, и для оценки современных терапевтических стратегий лечения ФКУ.
Родственные ферменты
[ редактировать ]Фенилаланингидроксилаза тесно связана с двумя другими ферментами:
- триптофангидроксилаза (номер ЕС 1.14.16.4), контролирующая уровень серотонина в головном мозге и желудочно-кишечном тракте.
- тирозингидроксилаза (номер ЕС 1.14.16.2), которая контролирует уровни дофамина , адреналина и норадреналина в мозге и мозговом веществе надпочечников.
Эти три фермента гомологичны, то есть полагают, что они произошли от одной и той же древней гидроксилазы.
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000171759 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: Ensembl, выпуск 89: ENSMUSG00000020051 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Фицпатрик П.Ф. (1999). «Гидроксилазы тетрагидроптерин-зависимых аминокислот». Ежегодный обзор биохимии . 68 : 355–81. doi : 10.1146/annurev.biochem.68.1.355 . ПМИД 10872454 .
- ^ Кауфман С. (февраль 1958 г.). «Новый кофактор, необходимый для ферментативного превращения фенилаланина в тирозин» . Журнал биологической химии . 230 (2): 931–9. дои : 10.1016/S0021-9258(18)70516-4 . ПМИД 13525410 .
- ^ Jump up to: а б с д и ж Фитцпатрик П.Ф. (декабрь 2003 г.). «Механизм гидроксилирования ароматических аминокислот» . Биохимия . 42 (48): 14083–91. дои : 10.1021/bi035656u . ПМЦ 1635487 . ПМИД 14640675 .
- ^ Панай А.Дж., Ли М., Кребс С., Боллинджер Дж.М., Фитцпатрик П.Ф. (март 2011 г.). «Доказательства наличия высокоспиновых разновидностей Fe (IV) в каталитическом цикле бактериальной фенилаланингидроксилазы» . Биохимия . 50 (11): 1928–33. дои : 10.1021/bi1019868 . ПМЦ 3059337 . ПМИД 21261288 .
- ^ Бассан А., Бломберг М.Р., Зигбан П.Е. (январь 2003 г.). «Механизм расщепления дикислорода в тетрагидробиоптеринзависимых гидроксилазах аминокислот» . Химия: Европейский журнал . 9 (1): 106–15. дои : 10.1002/chem.200390006 . ПМИД 12506369 .
- ^ Олссон Э., Мартинес А., Тейген К., Дженсен В.Р. (март 2011 г.). «Образование железо-оксогидроксилирующих частиц в каталитическом цикле гидроксилаз ароматических аминокислот». Химия: Европейский журнал . 17 (13): 3746–58. дои : 10.1002/chem.201002910 . ПМИД 21351297 .
- ^ Бассан А., Бломберг М.Р., Зигбан П.Е. (сентябрь 2003 г.). «Механизм ароматического гидроксилирования активированным ядром FeIV = O в тетрагидробиоптерин-зависимых гидроксилазах». Химия: Европейский журнал . 9 (17): 4055–67. дои : 10.1002/chem.200304768 . ПМИД 12953191 .
- ^ Павон Дж. А., Фитцпатрик П.Ф. (сентябрь 2006 г.). «Понимание каталитических механизмов фенилаланин- и триптофангидроксилазы на основе кинетического изотопного воздействия на ароматическое гидроксилирование» . Биохимия . 45 (36): 11030–7. дои : 10.1021/bi0607554 . ЧВК 1945167 . ПМИД 16953590 .
- ^ Jump up to: а б с д и ж г час я дж Флэтмарк Т., Стивенс Р.К. (август 1999 г.). «Структурное понимание гидроксилаз ароматических аминокислот и их мутантных форм, связанных с заболеваниями». Химические обзоры . 99 (8): 2137–2160. дои : 10.1021/cr980450y . ПМИД 11849022 .
- ^ Селвуд Т., Яффе ЭК (март 2012 г.). «Динамическая диссоциация гомоолигомеров и контроль функции белка» . Архив биохимии и биофизики . 519 (2): 131–43. дои : 10.1016/j.abb.2011.11.020 . ПМЦ 3298769 . ПМИД 22182754 .
- ^ Jump up to: а б с Джаффе Э.К., Стит Л., Лоуренс Ш., Андрейк М., Данбрэк Р.Л. (февраль 2013 г.). «Новая модель аллостерической регуляции фенилаланингидроксилазы: значение для болезней и терапии» . Архив биохимии и биофизики . 530 (2): 73–82. дои : 10.1016/j.abb.2012.12.017 . ПМК 3580015 . ПМИД 23296088 .
- ^ Артуро Э.К., Гупта К., Эру А., Стит Л., Кросс П.Дж., Паркер Э.Дж., Лолл П.Дж., Яффе Е.К. (март 2016 г.). «Первая структура полноразмерной фенилаланингидроксилазы млекопитающих раскрывает архитектуру автоингибированного тетрамера» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (9): 2394–9. Бибкод : 2016PNAS..113.2394A . дои : 10.1073/pnas.1516967113 . ПМК 4780608 . ПМИД 26884182 .
- ^ Яффе ЭК (август 2017 г.). «Новые белковые структуры обеспечивают обновленное понимание фенилкетонурии» . Молекулярная генетика и обмен веществ . 121 (4): 289–296. дои : 10.1016/j.ymgme.2017.06.005 . ПМЦ 5549558 . ПМИД 28645531 .
- ^ Патель Д., Копец Дж., Фитцпатрик Ф., МакКорви Т.Дж., Юэ У.В. (апрель 2016 г.). «Структурная основа лиганд-зависимой димеризации регуляторного домена фенилаланингидроксилазы» . Научные отчеты . 6 (1): 23748. Бибкод : 2016НатСР...623748П . дои : 10.1038/srep23748 . ПМЦ 4822156 . ПМИД 27049649 .
- ^ Мейсбургер С.П., Тейлор А.Б., Хан К.А., Чжан С., Фитцпатрик П.Ф., Андо Н. (май 2016 г.). «Движения доменов при активации фенилаланингидроксилазы, характеризуемые методами кристаллографии и хроматографически-связанного малоуглового рентгеновского рассеяния» . Журнал Американского химического общества . 138 (20): 6506–16. дои : 10.1021/jacs.6b01563 . ПМЦ 4896396 . ПМИД 27145334 .
- ^ Jump up to: а б Эрландсен Х., Фузетти Ф., Мартинес А., Хаф Э., Флэтмарк Т., Стивенс Р.К. (декабрь 1997 г.). «Кристаллическая структура каталитического домена фенилаланингидроксилазы человека раскрывает структурную основу фенилкетонурии». Структурная биология природы . 4 (12): 995–1000. дои : 10.1038/nsb1297-995 . ПМИД 9406548 . S2CID 6293946 .
- ^ Андерсен О.А., Флэтмарк Т., Хаф Э. (июль 2002 г.). «Кристаллическая структура тройного комплекса каталитического домена фенилаланингидроксилазы человека с тетрагидробиоптерином и 3-(2-тиенил)-L-аланином и ее значение для механизма катализа и активации субстрата». Журнал молекулярной биологии . 320 (5): 1095–108. дои : 10.1016/S0022-2836(02)00560-0 . ПМИД 12126628 .
- ^ Тейген К., Фрёйстейн Н.А., Мартинес А. (декабрь 1999 г.). «Структурные основы распознавания кофакторов фенилаланина и птерина фенилаланингидроксилазой: значение для каталитического механизма». Журнал молекулярной биологии . 294 (3): 807–23. дои : 10.1006/jmbi.1999.3288 . ПМИД 10610798 .
- ^ Jump up to: а б Ли Дж., Данготт Л.Дж., Фитцпатрик П.Ф. (апрель 2010 г.). «Регуляция фенилаланингидроксилазы: конформационные изменения при связывании фенилаланина, обнаруженные с помощью водородно-дейтериевого обмена и масс-спектрометрии» . Биохимия . 49 (15): 3327–35. дои : 10.1021/bi1001294 . ПМЦ 2855537 . ПМИД 20307070 .
- ^ Ли Дж., Илангован Ю., Даубнер С.К., Хинк А.П., Фитцпатрик П.Ф. (январь 2011 г.). «Прямое доказательство наличия фенилаланинового сайта в регуляторном домене фенилаланингидроксилазы» . Архив биохимии и биофизики . 505 (2): 250–5. дои : 10.1016/j.abb.2010.10.009 . ПМК 3019263 . ПМИД 20951114 .
- ^ Jump up to: а б с Коби Б., Дженнингс И.Г., Хаус СМ, Мичелл Б.Дж., Гудвилл К.Е., Сантарсиеро Б.Д., Стивенс Р.К., Коттон Р.Г., Кемп Б.Е. (май 1999 г.). «Структурные основы ауторегуляции фенилаланингидроксилазы». Структурная биология природы . 6 (5): 442–8. дои : 10.1038/8247 . ПМИД 10331871 . S2CID 11709986 .
- ^ Даубнер С.К., Хиллас П.Дж., Фитцпатрик П.Ф. (декабрь 1997 г.). «Экспрессия и характеристика каталитического домена фенилаланингидроксилазы человека». Архив биохимии и биофизики . 348 (2): 295–302. дои : 10.1006/abbi.1997.0435 . ПМИД 9434741 .
- ^ Jump up to: а б Бьорго Э., де Карвалью Р.М., Флэтмарк Т. (февраль 2001 г.). «Сравнение кинетических и регуляторных свойств тетрамерных и димерных форм дикого типа и Thr427 → Pro-мутантной фенилаланингидроксилазы человека: вклад гибкой шарнирной области Asp425-Gln429 в тетрамеризацию и кооперативное связывание субстрата». Европейский журнал биохимии . 268 (4): 997–1005. дои : 10.1046/j.1432-1327.2001.01958.x . ПМИД 11179966 .
- ^ Jump up to: а б с д Кауфман С. (март 1999 г.). «Модель метаболизма фенилаланина человека у нормальных людей и у пациентов с фенилкетонурией» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (6): 3160–4. Бибкод : 1999PNAS...96.3160K . дои : 10.1073/pnas.96.6.3160 . ПМК 15912 . ПМИД 10077654 .
- ^ Лихтер-Конецкий У., Хипке С.М., Конецкий Д.С. (август 1999 г.). «Экспрессия гена фенилаланингидроксилазы человека в почках и других непеченочных тканях». Молекулярная генетика и обмен веществ . 67 (4): 308–16. дои : 10.1006/mgme.1999.2880 . ПМИД 10444341 .
- ^ Мунтау AC, Герстинг SW (декабрь 2010 г.). «Фенилкетонурия как модель заболеваний, связанных с неправильным сворачиванием белков, и для разработки орфанных лекарств следующего поколения для пациентов с врожденными нарушениями метаболизма». Журнал наследственных метаболических заболеваний . 33 (6): 649–58. дои : 10.1007/s10545-010-9185-4 . ПМИД 20824346 . S2CID 20843095 .
- ^ Jump up to: а б с Сингх К., Корнелл К.С., Джексон Р., Кабири М., Фиппс М., Десаи М., Фогл Р., Ин Икс, Анарат-Капиллино Г., Геллер С., Джонсон Дж., Робертс Э., Мэлли К., Девлин Т., ДеРизо М., Бертелетт П., Чжан Ю.В., Райан С., Рао С., Терберг Б.Л., Бангари Д.С., Киостио-Мур С. (31 марта 2021 г.). «CRISPR/Cas9 создал нокаутных мышей, у которых отсутствует белок фенилаланингидроксилазы, в качестве новой доклинической модели фенилкетонурии человека» . Научные отчеты . 11 (1): 7254. doi : 10.1038/s41598-021-86663-8 . ПМК 8012645 . ПМИД 33790381 .
- ^ «Информационный бюллетень о нокаутирующих мышах» . Genome.gov .
- ^ «ПАУ-фенилаланингидроксилаза [Homo sapiens (человек)] - Ген - NCBI» . www.ncbi.nlm.nih.gov .
Дальнейшее чтение
[ редактировать ]- Эйзенсмит Р.К., Ву С.Л. (1993). «Молекулярные основы фенилкетонурии и связанных с ней гиперфенилаланинемий: мутации и полиморфизмы в гене фенилаланингидроксилазы человека» . Человеческая мутация . 1 (1): 13–23. дои : 10.1002/humu.1380010104 . ПМИД 1301187 . S2CID 19476605 .
- Конецкий Д.С., Лихтер-Конецкий У (август 1991 г.). «Локус фенилкетонурии: современные знания об аллелях и мутациях гена фенилаланингидроксилазы в различных популяциях». Генетика человека . 87 (4): 377–88. дои : 10.1007/BF00197152 . ПМИД 1679029 . S2CID 25627287 .
- Коттон Р.Г. (1991). «Гетерогенность фенилкетонурии на клиническом, белковом уровне и уровне ДНК». Журнал наследственных метаболических заболеваний . 13 (5): 739–50. дои : 10.1007/BF01799577 . ПМИД 2246858 . S2CID 21931016 .
- Эрландсен Х., Фузетти Ф., Мартинес А., Хаф Э., Флэтмарк Т., Стивенс Р.К. (декабрь 1997 г.). «Кристаллическая структура каталитического домена фенилаланингидроксилазы человека раскрывает структурную основу фенилкетонурии». Структурная биология природы . 4 (12): 995–1000. дои : 10.1038/nsb1297-995 . ПМИД 9406548 . S2CID 6293946 .
- Уотерс П.Дж., Парняк М.А., Новацкий П., Скривер Ч.Р. (1998). «Анализ экспрессии мутаций фенилаланингидроксилазы in vitro: связь генотипа с фенотипом и структуры с функцией» . Человеческая мутация . 11 (1): 4–17. doi : 10.1002/(SICI)1098-1004(1998)11:1<4::AID-HUMU2>3.0.CO;2-L . ПМИД 9450897 . S2CID 20099476 .
- Уотерс П.Дж. (апрель 2003 г.). «Как мутации гена ФАГ вызывают гиперфенилаланинемию и почему механизм имеет значение: результаты экспрессии in vitro». Человеческая мутация . 21 (4): 357–69. дои : 10.1002/humu.10197 . ПМИД 12655545 . S2CID 23769500 .
Внешние ссылки
[ редактировать ]- Запись GeneReviews/NCBI/NIH/UW о дефиците фенилаланингидроксилазы
- Локус-специфическая база данных вариантов гена фенилаланингидроксилазы человека
- Молекула месяца: фенилаланингидроксилаза. Архивировано 16 октября 2015 г. в Wayback Machine.
- Обзор всей структурной информации, доступной в PDB для UniProt : P00439 (фенилаланингидроксилаза человека) на PDBe-KB .
галерея PDB |
|---|