Фенилэтаноламин N-метилтрансфераза
| фенилэтаноламин N-метилтрансфераза | |||
|---|---|---|---|
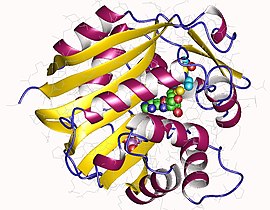 Мономер фенилэтаноламина N -метилтрансферазы, человека | |||
| Идентификаторы | |||
| Символ | ПНМТ | ||
| Альт. символы | ПЕНТ | ||
| ген NCBI | 5409 | ||
| HGNC | 9160 | ||
| МОЙ БОГ | 171190 | ||
| RefSeq | НМ_002686 | ||
| ЮниПрот | P11086 | ||
| Другие данные | |||
| Номер ЕС | 2.1.1.28 | ||
| Локус | Хр. 17 q21-q22 | ||
| |||
Фенилэтаноламин -N -метилтрансфераза ( PNMT ) представляет собой фермент , обнаруженный преимущественно в мозговом веществе надпочечников , который превращает норадреналин (норадреналин) в адреналин (адреналин). [1] Он также экспрессируется в небольших группах нейронов человеческого мозга. [2] и в избранных популяциях кардиомиоцитов . [3]
Структура
[ редактировать ]

PNMT представляет собой белок, кодирующий ген которого находится на хромосоме 17 у человека. Он состоит из 4 экзонов и представляет собой белок массой 30 кДа. Он разделяет многие свойства, обнаруженные среди других метилтрансфераз . По последовательности она наиболее близка к глицин -N -метилтрансферазе ( GNMT ). Он также имеет много общих структурных свойств, таких как форма складчатой губы, с катехол-О-метилтрансферазой (COMT), хотя его последовательность менее идентична. Некоторые особенности структуры, такие как эта складчатая губа, позволяют предположить, что PNMT представляет собой недавнюю адаптацию к семейству ферментов, синтезирующих катехоламины, развивающуюся позже, чем COMT, но раньше других метилтрансфераз, таких как GNMT. [5]
S- аденозил- L -метионин (SAM) является необходимым кофактором. [6] Область связывания активного сайта кофактора SAM содержит большое количество пи-связей от фенилаланина, а остатки тирозина в активном сайте помогают удерживать его в кармане связывания за счет пи-стэкинга . Среди всех известных в природе вариантов PNMT в активном центре консервативны 7 важнейших ароматических остатков. [5]
Остаток глутамин 185 необходим для связывания катехоламинового субстрата. Замена этого остатка другим снижает каталитическую эффективность ПНМТ в десятки-триста раз. [7]
В отсутствие ингибитора или лиганда фосфатная группа связывается с активным центром для стабилизации этой области. [8]
Человеческий ПНМТ образует в растворе димеры. При выращивании кристаллов ПНМТ в невосстанавливающих растворах между цистеинами 48 и 139 на противоположных цепях образуются две дисульфидные связи. Эта димеризация не влияет на каталитическую активность фермента. [9]
Механизм
[ редактировать ]PNMT катализирует перенос метильной группы от SAM к норэпинефрину, превращая ее в адреналин. Он работает путем объединения кофактора SAM и субстрата в непосредственной близости, так что реактивная метильная группа может быть атакована первичным амином молекулы норадреналина или другим субстратом катехоламина . Метильная группа SAM очень реакционноспособна, поэтому структура и расположение как норадреналина, так и SAM имеют решающее значение для правильного характера метилирования продукта.
В то время как PNMT метилирует норэпинефрин в активное соединение адреналин , норадреналин также может быть метилирован катехол-О-метилтрансферазой (COMT), другой метилтрансферазой , которая добавляет метильную группу в другом месте, в свою очередь производя неактивное соединение метанефрин . Метилтрансферазы очень распространены в путях синтеза и дезактивации катехоламинов. [10]
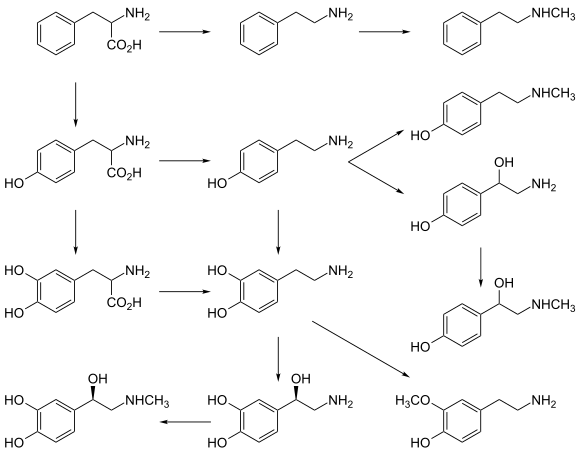
ПНМТ также участвует в биосинтезе N -метилированных следовых аминов : он метаболизирует фенэтиламин в N метилфенэтиламин ( позиционный изомер амфетамина синефрин ), п - октопамин в - и п - тирамин в N -метилтирамин . [11] [12]
Регулирование
[ редактировать ]Повышенная экспрессия PNMT является одним из способов положительной обратной связи реакции на стресс. Увеличение гормонов стресса или нервных импульсов из-за стресса может привести к тому, что PNMT преобразует больше норадреналина в адреналин. Это увеличивает эффективность системы реакции катехоламинов, увеличивая активность симпатической нервной системы и делая реакцию на стресс более глубокой. [14]
Известно, что ПНМТ регулируется глюкокортикоидами, вырабатываемыми в надпочечниках . Одним из способов регулирования экспрессии PNMT является положительное влияние кортикостерона на поддержание мРНК PNMT. [15] глюкокортикоиды Также было показано, что увеличивают биологический период полураспада фермента in vitro. [16] У животных, которым удалили гипофиз, добавление глюкокортикоидов значительно удлиняет период полувыведения ферментов PNMT. [16]
Повышенные уровни PNMT также могут быть вызваны импульсами висцеральных нервов. Нервные импульсы увеличивают синтез мРНК PNMT, воздействуя на определенные последовательности промотора. [16]
Также было показано, что стрессовая иммобилизация в течение нескольких часов увеличивает активность PNMT у крыс. Это лечение занимает около одной недели, чтобы выявить разницу в уровнях PNMT. [17]
SAM не только действует как кофактор PNMT, но также помогает стабилизировать фермент и увеличить период полураспада, делая его более устойчивым к расщеплению протеазой трипсина . [16]
Локализация
[ редактировать ]Было обнаружено, что синтез адреналина и, следовательно, расположение PNMT в основном локализуются в мозговом веществе надпочечников или надпочечниках у большинства видов. PNMT локализован у большинства взрослых млекопитающих в цитоплазме этих медуллярных клеток. [1]
Более новые исследования также показывают, что мРНК и белок PNMT экспрессируются и в других частях тела. Определенные нервные пути, сетчатка , [18] и как в предсердиях , так и в желудочках сердца в настоящее время выясняется, что они являются местами экспрессии PNMT. [19] Адреналин вырабатывается в небольших группах нейронов головного мозга человека, экспрессирующих PNMT; [2] эти нейроны отходят от ядра, прилежащего (вентролатерально) к постремной области, и от ядра в дорсальной области одиночного тракта . [2]
Болезнь
[ редактировать ]Нормальная функция и дефекты PNMT связаны с множеством заболеваний и расстройств.
Витилиго
[ редактировать ]Снижение уровня активности PNMT, измеряемого адреналином и норадреналином, наблюдается в коже пациентов с витилиго в кератиноцитах , которые обычно имеют более высокую активность PNMT. [20]
Интоксикация этанолом
[ редактировать ]Два мощных ингибитора PNMT (LY134046 и LY78335) были длительными антагонистами как интоксикации этанолом, так и седативного эффекта. Это предполагает центральную роль, которую PNMT и адреналин играют в синтезе седативного эффекта и интоксикации, вызванных этанолом и пентобарбиталом . [21]
болезнь Альцгеймера
[ редактировать ]Болезнь Альцгеймера также связана со снижением активности PNMT человека в областях мозга, наиболее связанных с дегенерацией при этом заболевании. Также были обнаружены значительные ассоциации с полиморфизмом PNMT и ранним началом болезни Альцгеймера. [22]
Торможение
[ редактировать ]Классические ингибиторы ПНМТ включают бензимидазолы , хинолоны и пурины . [8] Ингибирование также может быть вызвано добавлением S-дезоксиаденозил-L-гомоцистеина , замены кофактора SAM, который похож на него, но в нем отсутствует метильная группа, поэтому перенос метила невозможен. [23] Другой пример — CGS19281A. [24]
Ссылки
[ редактировать ]- ^ Jump up to: а б Гольдштейн М., Фуксе К., Хёкфельт Т. (июнь 1972 г.). «Характеристика и тканевая локализация ферментов, синтезирующих катехоламины». Фармакологические обзоры . 24 (2): 293–309. ПМИД 4564603 .
- ^ Jump up to: а б с Китахама К., Пирсон Дж., Денорой Л., Копп Н., Ульрих Дж., Маэда Т., Жуве М. (февраль 1985 г.). «Адренергические нейроны в мозге человека, продемонстрированные с помощью иммуногистохимии с антителами к фенилэтаноламин-N-метилтрансферазе (PNMT): открытие новой группы в ядре одиночного тракта». Письма по неврологии . 53 (3): 303–8. дои : 10.1016/0304-3940(85)90555-5 . ПМИД 3885079 . S2CID 2578817 .
- ^ Ван Ю, Лин В.К., Кроуфорд В., Ни Х., Болтон Э.Л., Хан Х. и др. (январь 2017 г.). «+ Клетки мышиного сердца» . Научные отчеты . 7 (1): 40687. дои : 10.1038/srep40687 . ПМК 5234027 . ПМИД 28084430 .
- ^ Jump up to: а б PDB : 4MQ4 ; Барт А.Г., Скотт Э.Э. Кристаллическая структура hPNMT в комплексе с бисубстратным ингибитором N-(3-((((2S,3S,4R,5R)-5-(6-амино-9H-пурин-9-ил)-3,4-дигидрокситетрагидрофуран-2 -ил)метил)тио)пропил)-1,2,3,4-тетрагидроизохинолин-3-карбоксамид (Отчет). Банк данных белков RCSB.
- ^ Jump up to: а б Мартин Дж.Л., Бегун Дж., Маклиш М.Дж., Кейн Дж.М., Грюневальд Г.Л. (октябрь 2001 г.). «Выработка адреналина: кристаллическая структура синтезирующего адреналин фермента PNMT» . Структура . 9 (10): 977–85. дои : 10.1016/s0969-2126(01)00662-1 . ПМИД 11591352 .
- ^ Вонг Д.Л., Лесаж А., Сиддалл Б., Фундер Дж.В. (ноябрь 1992 г.). «Глюкокортикоидная регуляция фенилэтаноламин-N-метилтрансферазы in vivo» . Журнал ФАСЭБ . 6 (14): 3310–5. дои : 10.1096/fasebj.6.14.1426768 . ПМИД 1426768 . S2CID 23761885 .
- ^ Дринкуотер Н., Джи К.Л., Пури М., Крисционе К.Р., Маклиш М.Дж., Грюневальд Г.Л., Мартин Дж.Л. (август 2009 г.). «Молекулярное распознавание физиологического субстрата норадреналина адреналинсинтезирующим ферментом ПНМТ и факторы, влияющие на его метилтрансферазную активность» . Биохимический журнал . 422 (3): 463–71. дои : 10.1042/bj20090702 . hdl : 1808/26489 . ПМК 5940352 . ПМИД 19570037 .
- ^ Jump up to: а б Дринкуотер Н., Ву Х., Ловелл К.М., Крисционе К.Р., Коллинз Б.М., Призинцано Т.Э. и др. (октябрь 2010 г.). «Скрининг на основе фрагментов методами рентгеновской кристаллографии, МС и изотермической титровальной калориметрии для идентификации ингибиторов PNMT (фенилэтаноламин-N-метилтрансферазы)». Биохимический журнал . 431 (1): 51–61. дои : 10.1042/bj20100651 . ПМИД 20642456 .
- ^ Джи К.Л., Нурс А., Синь А.Ю., Ву К., Тиндалл Дж.Д., Грюневальд Г.Л. и др. (июнь 2005 г.). «Дисульфидно-связанные димеры человеческого фермента, синтезирующего адреналин PNMT, являются каталитически активными». Biochimica et Biophysical Acta (BBA) - Белки и протеомика . 1750 (1): 82–92. дои : 10.1016/j.bbapap.2005.03.006 . ПМИД 15893506 .
- ^ Брандт. «Медулла надпочечников» (PDF) .
- ^ Jump up to: а б Бродли К.Дж. (март 2010 г.). «Сосудистые эффекты следовых аминов и амфетаминов». Фармакология и терапия . 125 (3): 363–375. doi : 10.1016/j.pharmthera.2009.11.005 . ПМИД 19948186 .
- ^ Jump up to: а б Линдеманн Л., Хонер MC (май 2005 г.). «Ренессанс следовых аминов, вдохновленный новым семейством GPCR». Тенденции в фармакологических науках . 26 (5): 274–281. дои : 10.1016/j.tips.2005.03.007 . ПМИД 15860375 .
- ^ Ван X, Ли Дж, Донг Дж, Юэ Дж (февраль 2014 г.). «Эндогенные субстраты CYP2D мозга». Европейский журнал фармакологии . 724 : 211–218. дои : 10.1016/j.ejphar.2013.12.025 . ПМИД 24374199 .
- ^ Вуртман Р.Дж. (июнь 2002 г.). «Стресс и адренокортикальный контроль синтеза адреналина». Метаболизм . 51 (6 Приложение 1): 11–4. дои : 10.1053/meta.2002.33185 . ПМИД 12040535 .
- ^ Цзян В., Ухт Р., Бон MC (1989). «Регуляция мРНК фенилэтаноламин-N-метилтрансферазы (PNMT) в мозговом веществе надпочечников крысы кортикостероном». Международный журнал нейробиологии развития . 7 (5): 513–20. дои : 10.1016/0736-5748(89)90010-5 . ПМИД 2816488 . S2CID 24803398 .
- ^ Jump up to: а б с д Чиаранелло Р.Д. (1978). «Регуляция фенилэтаноламин-N-метилтрансферазы». Биохимическая фармакология . 27 (15): 1895–7. дои : 10.1016/0006-2952(78)90002-3 . ПМИД 708473 .
- ^ Кэхилл А.Л., Эртмоед А.Л., Мангура Д., Перлман Р.Л. (сентябрь 1996 г.). «Дифференциальная регуляция экспрессии фенилэтаноламин-N-метилтрансферазы в двух различных субпопуляциях хромаффинных клеток крупного рогатого скота». Журнал нейрохимии . 67 (3): 1217–24. дои : 10.1046/j.1471-4159.1996.67031217.x . ПМИД 8752129 . S2CID 26602827 .
- ^ Пак Д.Х., Тейтельман Г., Эвинджер М.Дж., Ву Джи, Руджеро Д.А., Альберт В.Р. и др. (апрель 1986 г.). «Нейроны, содержащие фенилэтаноламин N-метилтрансферазу, в сетчатке крысы: иммуногистохимия, иммунохимия и молекулярная биология» . Журнал неврологии . 6 (4): 1108–13. doi : 10.1523/JNEUROSCI.06-04-01108.1986 . ПМК 6568425 . ПМИД 2871139 .
- ^ Кризанова О, Микуткова Л, Желокова Ю, Филипенко М, Саббан Е, Кветнанский Р (сентябрь 2001 г.). «Существование сердечной мРНК PNMT у взрослых крыс: повышение при стрессе глюкокортикоид-зависимым образом». Американский журнал физиологии. Физиология сердца и кровообращения . 281 (3): H1372-9. дои : 10.1152/ajpheart.2001.281.3.H1372 . ПМИД 11514309 .
- ^ Шаллройтер К.У. , Вуд Дж.М., Питтелькоу М.Р., Баттнер Г., Суонсон Н., Корнер С., Эрке С. (1996). «Повышение активности моноаминоксидазы А в эпидермисе пациентов с витилиго». Архив дерматологических исследований . 288 (1): 14–8. дои : 10.1007/bf02505037 . ПМИД 8750929 . S2CID 31646987 .
- ^ Меффорд И.Н., Листер Р.Г., Ота М., Линнойла М. (февраль 1990 г.). «Антагонизм этаноловой интоксикации у крыс ингибиторами фенилэтаноламин-N-метилтрансферазы» . Алкоголизм: клинические и экспериментальные исследования . 14 (1): 53–7. дои : 10.1111/j.1530-0277.1990.tb00446.x . ПМИД 2178473 .
- ^ Манн М.Б., Ву С., Ростамхани М., Туртеллотт В., МакМюррей Дж., Комингс Д.Э. (май 2001 г.). «Ген фенилэтаноламин-N-метилтрансферазы (PNMT) и болезнь Альцгеймера с ранним началом». Американский журнал медицинской генетики . 105 (4): 312–6. дои : 10.1002/ajmg.1363 . ПМИД 11378842 .
- ^ Борхардт RT, Ву Й.С. (март 1975 г.). «Потенциальные ингибиторы S-аденозилметионин-зависимых метилтрансфераз. 3. Модификации сахарной части S-аденозилгомоцистеина». Журнал медицинской химии . 18 (3): 300–4. дои : 10.1021/jm00237a018 . ПМИД 1133821 .
- ^ Атобе М., Кубота М., Накагавара М., Кария Т. (1996). «Влияние ингибитора фенилэтаноламин-N-метилтрансферазы CGS19281A на функцию альфа-2-адренорецепторов в гипоталамусе крыс в сравнении с SKF29661, SKF64139 и йохимбином». Нейропсихобиология . 34 (2): 82–89. дои : 10.1159/000119297 . ПМИД 8904737 .
Внешние ссылки
[ редактировать ]- Фенилэтаноламин + N-метилтрансфераза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)