Октопамин
 | |
 | |
| Клинические данные | |
|---|---|
| Другие имена | OCT, норсимпатол, норсинефрин, пара -октопамин, бета-гидрокситирамин, пара-гидроксифенилэтаноламин, α-(аминометил)-4 гидроксибензолметанол, 1-(п-гидроксифенил)-2-аминоэтанол |
| Маршруты администрация | Оральный |
| код АТС | |
| Физиологические данные | |
| Исходные ткани | нервные системы беспозвоночных; следы амина у позвоночных |
| Целевые ткани | общесистемный у беспозвоночных |
| Рецепторы | TAAR1 (млекопитающие) OctαR, OctβR, TyrR (беспозвоночные), Oct-TyrR |
| Агонисты | Формамидины (амитраз (АМЗ) и хлордимеформ (CDM)) |
| Антагонисты | эпинастин (3-амино-9,13b-дигидро-1H-дибенз(c,f)имидазо(1,5а)азепина гидрохлорид) |
| Предшественник | тирамин |
| Биосинтез | тирамин-β-гидроксилаза ; дофамин-β-гидроксилаза |
| Метаболизм | п -гидроксиминдальная кислота; [1] [2] N-ацетилтрансферазы ; фенилэтаноламин N-метилтрансфераза |
| Юридический статус | |
| Юридический статус |
|
| Фармакокинетические данные | |
| Биодоступность | 99.42 % |
| Метаболизм | п -гидроксиминдальная кислота; [1] [2] N-ацетилтрансферазы ; фенилэтаноламин N-метилтрансфераза |
| Период полувыведения | 15 минут в насекомых. У человека от 76 до 175 минут. |
| Экскреция | До 93% принятого октопамина выводится почками в течение 24 часов. [1] |
| Идентификаторы | |
| Номер CAS | |
| ПабХим CID | |
| ИЮФАР/БПС | |
| ХимическийПаук | |
| НЕКОТОРЫЙ | |
| КЕГГ | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| Панель управления CompTox ( EPA ) | |
| Информационная карта ECHA | 100.002.890 |
| Химические и физические данные | |
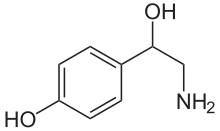
| Формула | С 8 Н 11 Н О 2 |
| Молярная масса | 153.181 g·mol −1 |
| 3D model ( JSmol ) | |
| | |
Октопамин (молекулярная формула C 8 H 11 NO 2 ; также известный как ОА, а также норсинефрин, пара -октопамин и другие) представляет собой органическое химическое вещество, тесно связанное с норэпинефрином и синтезируемое биологически гомологичным путем. Октопамин часто считают основным нейрогормоном «бей или беги» беспозвоночных. Его название происходит от того факта, что он впервые был обнаружен в слюнных железах осьминога.
У многих видов беспозвоночных октопамин является важным нейромедиатором и гормоном . У протостом — членистоногих, моллюсков и некоторых видов червей — он заменяет норадреналин и выполняет функции, по-видимому, сходные с функциями норадреналина у млекопитающих, функции, которые были описаны как мобилизация тела и нервной системы к действию. У млекопитающих октопамин обнаружен лишь в следовых количествах, и его биологическая функция не установлена. В природе он также содержится во многих растениях, включая горький апельсин . [4] [5] Октопамин продается под торговыми названиями, такими как Эпиренор , Норден и Норфен, для использования в качестве симпатомиметического препарата , доступного по рецепту.
Функции
[ редактировать ]Клеточные эффекты
[ редактировать ]Октопамин оказывает свое действие путем связывания и активации рецепторов, расположенных на поверхности клеток. Эти рецепторы в основном изучались у насекомых, где их можно разделить на отдельные типы:
- OctαR ( альфа-адренергические -подобные) структурно и функционально аналогичны норадренергическим альфа-1-рецепторам млекопитающих. Существует несколько подтипов рецептора OctαR. Например, у целующегося клопа ( Rhodnius prolixus ) есть Octα 1 -R, Octα 2 R. [6]
- OctβR ( бета-адренергические -подобные) структурно и функционально аналогичны норадренергическим бета-рецепторам млекопитающих. Существует несколько подтипов рецептора OctβR. Например, у плодовой мушки ( Drosophila melanogaster ) есть DmOctβ1R, DmOctβ2R и DmOctβ3R. [7]
- ОАМБ. Разнообразие этого рецептора относительно неизвестно. Плодовая мушка (Drosophila melanogaster) имеет две отдельные изоформы, которые функционально различаются: Oamb. К3 и Оамб КАК . [8]
- TyrR (смешанные октопамин/тираминовые рецепторы), которые структурно и функционально аналогичны норадренергическим альфа-2-рецепторам млекопитающих. [9] Однако рецепторы класса TyrR обычно более сильно активируются тирамином , чем октопамином. [9]
Филогенетические исследования утверждают, что у древних билатерий, таких как Platynereis dumerilii, существует сосуществование передачи сигналов рецепторов норадреналина , тирамина и октопамина. Однако из-за частичного перекрытия их сигнальной функции рецепторы тирамина и октопамина у позвоночных утрачены. [10]
У позвоночных не выявлено специфичных для октопамина рецепторов. Октопамин слабо связывается с рецепторами норадреналина и адреналина , но неясно, имеет ли это какое-либо функциональное значение. Он более прочно связывается с рецепторами, связанными с амином (TAAR), особенно с TAAR1 . [9]
Беспозвоночные
[ редактировать ]Октопамин был впервые открыт итальянским учёным Витторио Эрспамером в 1948 году. [11] в слюнных железах осьминогов и с тех пор было обнаружено, что он действует как нейромедиатор , нейрогормон и нейромодулятор у беспозвоночных . Хотя Эрспамер обнаружил его естественное происхождение и дал ему название, октопамин на самом деле существовал в течение многих лет как фармацевтический продукт. [12] Он широко используется всеми насекомыми, ракообразными (крабы, омары, раки) и пауками при энергозатратном поведении. Такое поведение включает в себя модуляцию мышечного напряжения, [13] летающий, [14] овуляция и яйцекладка, [15] [16] [17] [18] [19] [20] и прыжки. [21] [22]
У беспозвоночных, не являющихся насекомыми
[ редактировать ]У омаров октопамин, по-видимому, в некоторой степени направляет и координирует нейрогормоны в центральной нервной системе, и было замечено, что введение октопамина омарам и ракам приводило к растяжению конечностей и живота. [23]
У нематод октопамин обнаруживается в высоких концентрациях у взрослых особей, снижая яйцекладку и глоточное насосное поведение, оказывая антагонистическое действие на серотонин . [24]
Октопаминергические нервы моллюска могут присутствовать в сердце, с высокими концентрациями в нервной системе. [25]
У не принадлежащих к дрозофилам насекомых,
[ редактировать ]У насекомых октопамин высвобождается определенным количеством нейронов, но действует широко на весь центральный мозг, на все органы чувств и на некоторые ненейрональные ткани. [26] [27] В грудных ганглиях октопамин в первую очередь высвобождается нейронами DUM (дорсальный непарный срединный) и VUM (вентральный непарный срединный) нейронами, которые высвобождают октопамин на нервные, мышечные и периферические мишени. [28] [29] Эти нейроны важны для реализации энергозатратного двигательного поведения, такого как прыжки и полеты, вызванные бегством. Например, нейрон DUMeti саранчи высвобождает октопамин в мышцу-разгибатель большеберцовой кости, чтобы увеличить мышечное напряжение и увеличить скорость расслабления. Эти действия способствуют эффективному сокращению мышц ног при прыжках. [26] Во время полета нейроны DUM также активны и выделяют октопамин по всему телу для синхронизации энергетического обмена, дыхания, мышечной активности и активности летающих интернейронов. [14] Октопамина у саранчи саранчи в четыре раза больше концентрируется в аксоне, чем в соме, и он снижает миогенный ритм . [30]
У медоносных пчел октопамин играет важную роль в обучении и памяти. У светлячка высвобождение октопамина приводит к производству света в фонаре. [31] [32]
У личинок восточной совки октопамин иммунологически полезен, увеличивая выживаемость в популяциях с высокой плотностью. [33]
Изумрудная оса-таракан жалит хозяина своей личинкой (тараканом) в головной ганглий (мозг). Яд блокирует рецепторы октопамина. [34] а таракан не проявляет нормальных реакций на побег и чрезмерно ухаживает за собой. Он становится послушным, и оса ведет его к осиному логову, дергая за антенну, как за поводок. [35]
У дрозофилы
[ редактировать ]Октопамин влияет почти на все процессы плодовой мухи и широко присутствует как у взрослых особей, так и у личинок. Неполный список некоторых областей, в которых модулируется октопамин:
- Обучение и память [36] [37] [38]
- Овуляция и яйцекладка [15] [16] [17] [18] [19] [20]
- Передвижение [39] [40] [41]
- Мышечная физиология [13] [42]
- Агрессия [43] [44] [45]
- Толерантность к алкоголю и наркотикам [46] [47] [48] [49]
- Кормление [50] [51]
- Микробиом и физиология кишечника [52] [53]
- Спать [54] [55]
- Модулирующий эффект физических упражнений [56] [57]
- Метаболизм [58] [59]
Позвоночные животные
[ редактировать ]У позвоночных октопамин заменяет норадреналин в симпатических нейронах при хроническом применении ингибиторов моноаминоксидазы . Это может быть причиной частого побочного эффекта ортостатической гипотензии при применении этих препаратов, хотя есть также доказательства того, что он на самом деле опосредован повышением уровня N -ацетилсеротонина .
В одном исследовании было отмечено, что октопамин может быть важным амином, который влияет на терапевтические эффекты ингибиторов, таких как ингибиторы моноаминоксидазы , особенно потому, что при лечении животных этим ингибитором наблюдалось значительное увеличение уровней октопамина. Октопамин был положительно идентифицирован в образцах мочи млекопитающих, таких как люди, крысы и кролики, получавших ингибиторы моноаминоксидазы . Очень небольшие количества октопамина также были обнаружены в некоторых тканях животных. Было замечено, что в организме кролика сердце и почки содержат самые высокие концентрации октопамина. Было обнаружено, что октопамин на 93% элюируется мочой в течение 24 часов после его образования в организме в качестве побочного продукта ипрониазида у кроликов. [12]
Фармакология
[ редактировать ]Октопамин продается под торговыми названиями, такими как Эпиренор , Норден и Норфен, для использования в медицине в качестве симпатомиметического препарата , доступного по рецепту. Однако существует очень мало информации о его клинической полезности и безопасности. [60]
У млекопитающих октопамин может мобилизовать высвобождение жира из адипоцитов (жировых клеток), что привело к его рекламе в Интернете как средства для похудения. Однако высвободившийся жир, скорее всего, будет быстро поглощен другими клетками, и нет никаких доказательств того, что октопамин способствует снижению веса. Октопамин также может значительно повышать кровяное давление в сочетании с другими стимуляторами , например, с некоторыми добавками для похудания . [61]
Всемирное антидопинговое агентство вносит октопамин в список запрещенных веществ для использования на соревнованиях и в качестве «специфического стимулятора». [62] в Запрещенном списке 2019 года.
Инсектициды
[ редактировать ]Рецептор октопамина является мишенью инсектицидов, поскольку его блокировка приводит к снижению уровня циклического аденозинмонофосфата (цАМФ). Эфирные масла могут оказывать такое нейроинсектицидное действие, [63] и этот механизм рецептора октопамина естественным образом используется растениями с активными инсектицидными фитохимическими веществами. [64]
Биохимические механизмы
[ редактировать ]Млекопитающие
[ редактировать ]Октопамин является одним из четырех основных эндогенных агонистов человеческого рецептора 1, связанного с следами аминов, вместе с 3-йодтиронамином , дофамином и тирамином . [65] [66]
Беспозвоночные
[ редактировать ]Октопамин связывается с соответствующими рецепторами, связанными с G-белком (GPCR), инициируя путь передачи клеточного сигнала. Были определены по крайней мере три группы октопаминовых GPCR. OctαR (рецепторы октопамина 1) более тесно связаны с α-адренергическими рецепторами, тогда как OctβR (рецепторы октопамина 2) более тесно связаны с β-адренергическими рецепторами. Рецепторы октопамина/тирамина (включая Oct-TyrR) могут связывать оба лиганда и проявлять специфическое для агониста связывание. Oct-TyrR включен в генные группы как ОКТОПАМИНОВЫХ, так и ТИРАМИНОВЫХ РЕЦЕПТОРОВ. [67]
Биосинтез
[ редактировать ]
У насекомых
[ редактировать ]Октопамин действует как у насекомых эквивалент норадреналина и участвует в регулировании агрессии у беспозвоночных, оказывая разное воздействие на разные виды. Исследования показали, что снижение уровня нейромедиатора октопамина и предотвращение кодирования тирамин-бета-гидроксилазы (фермента, превращающего тирамин в октопамин) снижает агрессию у дрозофилы, не влияя на другое поведение. [68]
У людей
[ редактировать ]См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Хенгстманн Дж.Х., Конен В., Конен С., Эйхельбаум М., Денглер Х.Дж. (1974). «Физиологическое расположение п-октопамина у человека». Архив фармакологии Наунина-Шмидеберга . 283 (1): 93–106. дои : 10.1007/bf00500148 . ПМИД 4277715 . S2CID 35523412 .
- ^ Д'Андреа Г., Нордера Г., Пиццолато Г., Болнер А., Колавито Д., Флаибани Р. и др. (январь 2010 г.). «Метаболизм следовых аминов при болезни Паркинсона: низкие уровни циркулирующего октопамина на ранних стадиях заболевания». Письма по неврологии . 469 (3): 348–351. дои : 10.1016/j.neulet.2009.12.025 . ПМИД 20026245 . S2CID 12797090 .
- ^ «Справочник новых ингредиентов пищевых добавок FDA | Foley & Lardner LLP» . www.foley.com . 5 июня 2023 г. Проверено 10 июня 2023 г.
- ^ Тан Ф., Тао Л., Луо Икс, Дин Л., Го М., Не Л. и др. (сентябрь 2006 г.). «Определение октопамина, синефрина и тирамина в цитрусовых травах с помощью ионной жидкости, улучшенной« зеленой »хроматографии». Журнал хроматографии А. 1125 (2): 182–188. дои : 10.1016/j.chroma.2006.05.049 . ПМИД 16781718 .
- ^ Ягелло-Войтович Э (1979). «Механизм центрального действия октопамина». Польский журнал фармакологии и фармации . 31 (5): 509–516. ПМИД 121158 .
- ^ Хана С., Lange AB (26 сентября 2017 г.). «Клонирование и функциональная характеристика рецептора Octβ2 и рецептора Tyr1 у переносчика болезни Шагаса, Rhodnius prolixus » . Границы в физиологии . 8 : 744. doi : 10.3389/fphys.2017.00744 . ПМК 5623054 . ПМИД 29018364 .
- ^ Макейра Б., Чатвин Х., Эванс П.Д. (июль 2005 г.). «Идентификация и характеристика нового семейства рецепторов, связанных с бета-адренергически подобными октопамину G-белком дрозофилы». Журнал нейрохимии . 94 (2): 547–560. дои : 10.1111/j.1471-4159.2005.03251.x . ПМИД 15998303 . S2CID 83666118 .
- ^ Ли Х.Г., Рохила С., Хан К.А. (5 марта 2009 г.). «Рецептор октопамина OAMB опосредует овуляцию через Ca2+/кальмодулин-зависимую протеинкиназу II в эпителии яйцевода дрозофилы» . ПЛОС ОДИН . 4 (3): е4716. Бибкод : 2009PLoSO...4.4716L . дои : 10.1371/journal.pone.0004716 . ПМК 2650798 . ПМИД 19262750 .
- ^ Jump up to: а б с Пфлюгер Х.Дж., Стивенсонб, Пенсильвания (2005). «Эволюционные аспекты октопаминэргических систем с акцентом на членистоногих» . Строение и развитие членистоногих . 34 (3): 379–396. дои : 10.1016/j.asd.2005.04.004 .
- ^ Баукнехт П., Йекели Г (январь 2017 г.). «Древнее сосуществование передачи сигналов норадреналина, тирамина и октопамина у билатерий» . БМК Биология . 15 (1): 6. дои : 10.1186/s12915-016-0341-7 . ПМК 5282848 . ПМИД 28137258 .
- ^ Эрспамер В (2009). «Активные вещества в задних слюнных железах осьминоги. II. Тирамин и октопамин (оксиоктопамин)». Acta Pharmacologica et Toxicologica . 4 (3–4): 224–47. дои : 10.1111/j.1600-0773.1948.tb03345.x .
- ^ Jump up to: а б Какимото Ю., Армстронг, доктор медицины (февраль 1962 г.). «Об идентификации октопамина у млекопитающих» . Журнал биологической химии . 237 (2): 422–427. дои : 10.1016/S0021-9258(18)93937-2 . ПМИД 14453200 .
- ^ Jump up to: а б Ормерод К.Г., Хадден Дж.К., Дэди Л.Д., Мерсье А.Дж., Кранс Дж.Л. (октябрь 2013 г.). «Действие октопамина и тирамина на мышцы личинок Drosophila melanogaster». Журнал нейрофизиологии . 110 (8): 1984–1996. дои : 10.1152/jn.00431.2013 . hdl : 10464/6361 . ПМИД 23904495 .
- ^ Jump up to: а б Орчард I, Рамирес Х.М., Ланге А.Б. (январь 1993 г.). «Многофункциональная роль октопамина в бегстве саранчи». Ежегодный обзор энтомологии . 38 (1): 227–249. doi : 10.1146/annurev.en.38.010193.001303 . ISSN 0066-4170 .
- ^ Jump up to: а б Ли Х.Г., Сон К.С., Ким Ю.К., Дэвис Р.Л., Хан К.А. (декабрь 2003 г.). «Рецептор октопамина OAMB необходим для овуляции у Drosophila melanogaster» . Биология развития . 264 (1): 179–190. дои : 10.1016/j.ydbio.2003.07.018 . ПМИД 14623240 .
- ^ Jump up to: а б Ли Ю, Финк С., Эль-Холи С., Редер Т. (март 2015 г.). «Рецептор октопамина Octoberß2R необходим для овуляции и оплодотворения у плодовой мухи Drosophila melanogaster». Архив биохимии и физиологии насекомых . 88 (3): 168–178. дои : 10.1002/arch.21211 . ПМИД 25353988 .
- ^ Jump up to: а б Лим Дж., Сабандал П.Р., Фернандес А., Сабандал Дж.М., Ли Х.Г., Эванс П. и др. (6 августа 2014 г.). Бротон С. (ред.). «Рецептор октопамина Octβ2R регулирует овуляцию у Drosophila melanogaster» . ПЛОС ОДИН . 9 (8): e104441. Бибкод : 2014PLoSO...9j4441L . дои : 10.1371/journal.pone.0104441 . ПМК 4123956 . ПМИД 25099506 .
- ^ Jump up to: а б Ли Х.Г., Рохила С., Хан К.А. (5 марта 2009 г.). Луи М. (ред.). «Рецептор октопамина OAMB опосредует овуляцию через Ca2+/кальмодулин-зависимую протеинкиназу II в эпителии яйцевода дрозофилы» . ПЛОС ОДИН . 4 (3): е4716. Бибкод : 2009PLoSO...4.4716L . дои : 10.1371/journal.pone.0004716 . ПМК 2650798 . ПМИД 19262750 .
- ^ Jump up to: а б Монастириоти М (декабрь 2003 г.). «Отличная популяция октопаминовых клеток, проживающая в абдоминальном ганглие ЦНС, контролирует овуляцию у Drosophila melanogaster» . Биология развития . 264 (1): 38–49. дои : 10.1016/j.ydbio.2003.07.019 . ПМИД 14623230 .
- ^ Jump up to: а б Дэди Л.Д., Сан Дж. (октябрь 2015 г.). Вольфнер М.Ф. (ред.). «Анализ разрыва фолликула показывает важную роль фолликулярной адренергической передачи сигналов в овуляции дрозофилы» . ПЛОС Генетика . 11 (10): e1005604. дои : 10.1371/journal.pgen.1005604 . ПМЦ 4608792 . ПМИД 26473732 .
- ^ Поллак А.Дж., Ритцманн Р.Э., Вестин Дж. (сентябрь 1988 г.). «Активация интернейронов клеток DUM вентральными гигантскими интернейронами у таракана Periplaneta americana». Журнал нейробиологии . 19 (6): 489–497. дои : 10.1002/neu.480190602 . ПМИД 3171574 .
- ^ Фруктовый сад I (1 апреля 1982 г.). «Октопамин у насекомых: нейромедиатор, нейрогормон и нейромодулятор». Канадский журнал зоологии . 60 (4): 659–669. дои : 10.1139/z82-095 . ISSN 0008-4301 .
- ^ Ливингстон М.С., Харрис-Уоррик Р.М., Кравиц Э.А. (апрель 1980 г.). «Серотонин и октопамин вызывают у омаров противоположные позы». Наука . 208 (4439): 76–79. Бибкод : 1980Sci...208...76L . дои : 10.1126/science.208.4439.76 . ПМИД 17731572 . S2CID 32141532 .
- ^ Хорвиц Х.Р., Чалфи М., Трент С., Салстон Дж.Э., Эванс П.Д. (май 1982 г.). «Серотонин и октопамин у нематоды Caenorhabditis elegans». Наука . 216 (4549): 1012–1014. Бибкод : 1982Sci...216.1012H . дои : 10.1126/science.6805073 . ПМИД 6805073 .
- ^ Дуган Д.Ф., Даффилд П.Х., Уэйд Д.Н., Даффилд А.М. (январь 1981 г.). «Возникновение и синтез октопамина в сердце и ганглиях моллюска Tapes watlingi». Сравнительная биохимия и физиология. Часть C: Сравнительная фармакология . 70 (2): 277–280. дои : 10.1016/0306-4492(81)90064-2 . ISSN 0306-4492 .
- ^ Jump up to: а б Этвуд Х.Л., Клозе М.К. (1 января 2009 г.), «Модуляция нервно-мышечной передачи в нервно-мышечных соединениях беспозвоночных» , в Squire LR (редактор), Энциклопедия нейронаук , Oxford: Academic Press, стр. 671–690, doi : 10.1016/B978- 008045046-9.01262-6 , ISBN 978-0-08-045046-9 , получено 10 июля 2020 г.
- ^ Редер Т. (декабрь 1999 г.). «Октопамин у беспозвоночных». Прогресс нейробиологии . 59 (5): 533–561. дои : 10.1016/s0301-0082(99)00016-7 . ПМИД 10515667 . S2CID 25654298 .
- ^ Эккерт М., Рапус Дж., Нюрнбергер А., Пенцлин Х. (август 1992 г.). «Новое специфическое антитело обнаруживает подобную октопамину иммунореактивность в вентральном нервном канатике таракана». Журнал сравнительной неврологии (на французском языке). 322 (1): 1–15. дои : 10.1002/cne.903220102 . ПМИД 1430305 . S2CID 41099770 .
- ^ Синакевич И.Г., Джеффард М., Пелате М., Лапьед Б. (апрель 1994 г.). «Октопаминоподобная иммунореактивность в дорсальных непарных срединных (DUM) нейронах, иннервирующих добавочную железу самца таракана Periplaneta americana». Исследования клеток и тканей . 276 (1): 15–21. дои : 10.1007/bf00354779 . ISSN 0302-766X . S2CID 23485136 .
- ^ Эванс П.Д., О'Ши М. (апрель 1978 г.). «Идентификация октопаминэргического нейрона и модуляция миогенного ритма у саранчи». Журнал экспериментальной биологии . 73 : 235–260. дои : 10.1242/jeb.73.1.235 . ПМИД 25941 .
- ^ Гринфилд, доктор медицины (ноябрь 2001 г.). «Обнаружено недостающее звено в биолюминесценции светлячков: НЕТ регуляции дыхания фотоцитов». Биоэссе . 23 (11): 992–995. дои : 10.1002/bies.1144 . ПМИД 11746215 .
- ^ Триммер Б.А., Април Дж.Р., Дудзински Д.М., Лагас С.Дж., Льюис С.М., Мишель Т. и др. (июнь 2001 г.). «Оксид азота и контроль мигания светлячка». Наука . 292 (5526): 2486–2488. дои : 10.1126/science.1059833 . ПМИД 11431567 . S2CID 1095642 .
- ^ Конг Х., Юань Л., Донг С., Чжэн М., Цзин В., Тянь З. и др. (декабрь 2020 г.). «Иммунологическая регуляция с помощью гена β-адренергически подобного рецептора октопамина у скученных личинок восточной совки Mythmina separata». Развивающая и сравнительная иммунология . 113 : 103802. doi : 10.1016/j.dci.2020.103802 . ПМИД 32712170 . S2CID 220797641 .
- ^ Хопкин М (2007). «Как сделать таракана-зомби». Природа . дои : 10.1038/news.2007.312 .
- ^ Гал Р., Розенберг Л.А., Libersat F (декабрь 2005 г.). «Оса-паразитоид использует ядовитый коктейль, вводимый в мозг, чтобы манипулировать поведением и метаболизмом своей жертвы-таракана». Архив биохимии и физиологии насекомых . 60 (4): 198–208. дои : 10.1002/arch.20092 . ПМИД 16304619 .
- ^ Сабандал Дж. М., Сабандал П. Р., Ким Ю. К., Хан К. А. (май 2020 г.). «Согласованные действия октопаминовых и дофаминовых рецепторов стимулируют обонятельное обучение» . Журнал неврологии . 40 (21): 4240–4250. doi : 10.1523/JNEUROSCI.1756-19.2020 . ПМК 7244198 . ПМИД 32277043 .
- ^ Берк С.Дж., Хюттерот В., Овальд Д., Перисс Э., Крашес М.Дж., Дас Г. и др. (декабрь 2012 г.). «Многослойная передача сигналов вознаграждения через октопамин и дофамин у дрозофилы» . Природа . 492 (7429): 433–437. Бибкод : 2012Natur.492..433B . дои : 10.1038/nature11614 . ПМЦ 3528794 . ПМИД 23103875 .
- ^ Шварцель М., Монастириоти М., Шольц Х., Фригги-Грелин Ф., Бирман С., Гейзенберг М. (ноябрь 2003 г.). «Дофамин и октопамин различают аверсивные и аппетитные обонятельные воспоминания у дрозофилы» . Журнал неврологии . 23 (33): 10495–10502. doi : 10.1523/JNEUROSCI.23-33-10495.2003 . ПМК 6740930 . ПМИД 14627633 .
- ^ Шютцлер Н., Гирверт С., Хюгли И., Мохана Г., Руанант Ю., Риглевски С. и др. (февраль 2019 г.). «Действие тирамина на возбудимость мотонейронов и адаптируемое соотношение тирамин/октопамин регулируют передвижение дрозофилы в зависимости от состояния питания» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (9): 3805–3810. дои : 10.1073/pnas.1813554116 . ПМК 6397572 . ПМИД 30808766 .
- ^ Сельчо М., Паулс Д., Эль Джунди Б., Стокер Р.Ф., Тум А.С. (ноябрь 2012 г.). «Роль октопамина и тирамина в передвижении личинок дрозофилы» . Журнал сравнительной неврологии . 520 (16): 3764–3785. дои : 10.1002/cne.23152 . ПМИД 22627970 . S2CID 17014658 .
- ^ Сарасвати С., Фокс Л.Е., Солл Д.Р., Ву К.Ф. (март 2004 г.). «Тирамин и октопамин оказывают противоположное влияние на передвижение личинок дрозофилы». Журнал нейробиологии . 58 (4): 425–441. дои : 10.1002/neu.10298 . ПМИД 14978721 .
- ^ Ормерод К.Г., Юнг Дж., Мерсье А.Дж. (сентябрь 2018 г.). «Модуляция нервно-мышечных синапсов и сокращение у личинок дрозофилы 3-го возраста» . Журнал нейрогенетики . 32 (3): 183–194. дои : 10.1080/01677063.2018.1502761 . ПМИД 30303434 . S2CID 52948972 .
- ^ Эндрюс Дж.К., Фернандес, член парламента, Ю К, Лири Г.П., Люнг А.К., Кавано, член парламента и др. (май 2014 г.). Кландинин Т. (ред.). «Нейромодуляция октопамина регулирует связанные с Gr32a пути агрессии и ухаживания у самцов дрозофилы» . ПЛОС Генетика . 10 (5): e1004356. дои : 10.1371/journal.pgen.1004356 . ПМК 4031044 . ПМИД 24852170 .
- ^ Луо Дж., Лущак О.В., Гёрген П., Уильямс М.Дж., Нэссель Д.Р. (12 июня 2014 г.). Бротон С. (ред.). «Инсулин-продуцирующие клетки дрозофилы по-разному модулируются рецепторами серотонина и октопамина и влияют на социальное поведение» . ПЛОС ОДИН . 9 (6): e99732. Бибкод : 2014PLoSO...999732L . дои : 10.1371/journal.pone.0099732 . ПМК 4055686 . ПМИД 24923784 .
- ^ Уильямс М.Дж., Герген П., Раджендран Дж., Клокарс А., Касаяннис А., Фредрикссон Р. и др. (январь 2014 г.). «Регуляция агрессии генами TfAP-2 и Twz, связанными с ожирением, посредством передачи сигналов октопамина у дрозофилы» . Генетика . 196 (1): 349–362. дои : 10.1534/genetics.113.158402 . ПМЦ 3872196 . ПМИД 24142897 .
- ^ Хеберляйн Ю, Вольф Ф.В., Ротенфлю А., Гварниери DJ (август 2004 г.). «Молекулярно-генетический анализ интоксикации этанолом у Drosophila melanogaster». Интегративная и сравнительная биология . 44 (4): 269–274. CiteSeerX 10.1.1.536.262 . дои : 10.1093/icb/44.4.269 . ПМИД 21676709 . S2CID 14762870 .
- ^ Текотт Л.Х., Хеберлейн У (декабрь 1998 г.). «Да мы пьем?» . Клетка . 95 (6): 733–735. дои : 10.1016/S0092-8674(00)81695-5 . ПМИД 9865690 .
- ^ Уильямс Р. (22 июня 2005 г.). «Барные мухи: чему наши родственники-насекомые могут научить нас о толерантности к алкоголю» . Голый учёный .
- ^ Винс Дж. (22 августа 2005 г.). « Ген похмелья — ключ к толерантности к алкоголю» . Новый учёный .
- ^ Сельчо М., Паулс Д. (декабрь 2019 г.). «Связь физиологических процессов и пищевого поведения с помощью октопамина». Современное мнение в области науки о насекомых . 36 : 125–130. дои : 10.1016/j.cois.2019.09.002 . ПМИД 31606580 . S2CID 203470883 .
- ^ Сайин С., Де Бакер Дж. Ф., Сиджу К. П., Восняк М. Е., Льюис Л. П., Фриш Л. М. и др. (ноябрь 2019 г.). «Нерная цепь решает между настойчивостью и замкнутостью у голодной дрозофилы» . Нейрон . 104 (3): 544–558.e6. дои : 10.1016/j.neuron.2019.07.028 . ПМК 6839618 . ПМИД 31471123 .
- ^ Цзя Ю, Джин С., Ху К., Гэн Л., Хан С., Кан Р. и др. (май 2021 г.). «Микробиом кишечника модулирует агрессию дрозофилы посредством передачи сигналов октопамина» . Природные коммуникации . 12 (1): 2698. Бибкод : 2021NatCo..12.2698J . дои : 10.1038/s41467-021-23041-y . ПМЦ 8113466 . ПМИД 33976215 .
- ^ Шреттер С.Э., Вилметтер Дж., Бартос И., Марка З., Марка С., Аргаде С. и др. (ноябрь 2018 г.). «Кишечный микробный фактор модулирует двигательное поведение дрозофилы» . Природа . 563 (7731): 402–406. Бибкод : 2018Natur.563..402S . дои : 10.1038/s41586-018-0634-9 . ПМК 6237646 . ПМИД 30356215 .
- ^ Налл А., Сегал А. (июнь 2014 г.). «Моноамины и сон у дрозофилы». Поведенческая нейронаука . 128 (3): 264–272. дои : 10.1037/a0036209 . ПМИД 24886188 .
- ^ Эрион Р., ДиАнджело-младший, Крокер А., Сегал А. (сентябрь 2012 г.). «Взаимодействие между сном и обменом веществ у дрозофилы с измененной передачей сигналов октопамина» . Журнал биологической химии . 287 (39): 32406–32414. дои : 10.1074/jbc.M112.360875 . ПМЦ 3463357 . ПМИД 22829591 .
- ^ Суйковски А., Гретцингер А., Соаве Н., Тоди С.В., Уэсселс Р. (июнь 2020 г.). Бай Х (ред.). «Альфа- и бета-адренергические октопаминовые рецепторы в мышцах и сердце необходимы для адаптации дрозофилы к физическим нагрузкам» . ПЛОС Генетика . 16 (6): e1008778. дои : 10.1371/journal.pgen.1008778 . ПМЦ 7351206 . ПМИД 32579604 .
- ^ Кобб Т., Суйковски А., Мортон С., Рамеш Д., Уэсселс Р. (июль 2020 г.). «Различия в подвижности и адаптации к физическим упражнениям между видами дрозофилы» . Журнал сравнительной физиологии А. 206 (4): 611–621. дои : 10.1007/s00359-020-01421-x . ПМЦ 7314734 . ПМИД 32335730 .
- ^ Ахмед М.А., Vogel CF (август 2020 г.). «Опасное воздействие агонистов рецепторов октопамина на изменение генов, связанных с метаболизмом, и поведения Drosophila melanogaster» . Хемосфера . 253 : 126629. Бибкод : 2020Chmsp.25326629A . doi : 10.1016/j.chemSphere.2020.126629 . ПМЦ 9888421 . ПМИД 32283422 . S2CID 215757990 .
- ^ Ли Ю, Хоффманн Дж, Ли Ю, Стефано Ф, Бруххаус И, Финк С и др. (октябрь 2016 г.). «Октопамин контролирует устойчивость к голоданию, продолжительность жизни и метаболические особенности дрозофилы» . Научные отчеты . 6 (1): 35359. Бибкод : 2016NatSR...635359L . дои : 10.1038/srep35359 . ПМК 5069482 . ПМИД 27759117 .
- ^ Стохс С.Дж. (январь 2015 г.). «Физиологические функции и фармакологические и токсикологические эффекты п-октопамина». Лекарственная и химическая токсикология . 38 (1): 106–112. дои : 10.3109/01480545.2014.900069 . ПМИД 24654910 . S2CID 21901553 .
- ^ Халлер К.А., Беновиц Н.Л., Джейкоб П. (сентябрь 2005 г.). «Гемодинамические эффекты добавок для похудания без эфедры у людей». Американский медицинский журнал . 118 (9): 998–1003. doi : 10.1016/j.amjmed.2005.02.034 . ПМИД 16164886 .
- ^ «Запрещено на соревнованиях – стимуляторы» . ВАДА. Архивировано из оригинала 6 мая 2019 года . Проверено 6 мая 2019 г.
- ^ Энан Э (ноябрь 2001 г.). «Инсектицидная активность эфирных масел: октопаминэргические места действия». Сравнительная биохимия и физиология. Токсикология и фармакология . 130 (3): 325–337. дои : 10.1016/S1532-0456(01)00255-1 . ПМИД 11701389 .
- ^ Ротанг RS (1 сентября 2010 г.). «Механизм действия инсектицидных вторичных метаболитов растительного происхождения». Защита урожая . 29 (9): 913–920. дои : 10.1016/j.cropro.2010.05.008 . ISSN 0261-2194 .
- ^ Магуайр Джей Джей, Davenport AP (20 февраля 2018 г.). «Рецептор следовых аминов: рецептор ТА 1 » . Руководство IUPHAR/BPS по ФАРМАКОЛОГИИ . Международный союз фундаментальной и клинической фармакологии . Проверено 16 июля 2018 г.
- ^ Хеффернан М.Л., Герман Л.В., Браун С., Джонс П.Г., Шао Л., Хьюитт М.К. и др. (январь 2022 г.). «Улотаронт: агонист TAAR1 для лечения шизофрении» . Письма ACS по медицинской химии . 13 (1) (опубликовано 6 декабря 2021 г.): 92–98. doi : 10.1021/acsmedchemlett.1c00527 . ПМЦ 8762745 . ПМИД 35047111 .
- ^ «Геновая группа: ОКТОПАМИНОВЫЕ РЕЦЕПТОРЫ» . ФлайБейс . 16 октября 2018 г.
- ^ Чжоу С., Рао Ю, Рао Ю (сентябрь 2008 г.). «Подмножество октопаминэргических нейронов важно для агрессии дрозофилы». Природа Нейронауки . 11 (9): 1059–1067. дои : 10.1038/nn.2164 . ПМИД 19160504 . S2CID 1134848 .