Катехоламин
| Катехоламины |
|---|
 Эпинефрин (Адреналин) |
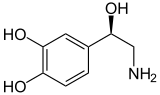 Норадреналин (Норадреналин) |
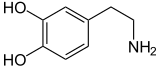 Дофамин |

Катехоламин , ( / ˌ k æ t ə ˈ k oʊ l ə m iː n / ; сокращенно CA ) — моноаминовый нейромедиатор , органическое соединение которое имеет катехин ( бензол с двумя гидроксильными боковыми группами рядом друг с другом) и боковую цепной амин . [1]
Катехол может быть либо свободной молекулой, либо заместителем более крупной молекулы, где он представляет собой 1,2-дигидроксибензольную группу.
Катехоламины образуются из аминокислоты тирозина , которая поступает из пищевых источников, а также синтезируется из фенилаланина . [2] Катехоламины водорастворимы и на 50% связаны с белками плазмы крови.
К катехоламинам относятся адреналин (адреналин), норадреналин (норадреналин) и дофамин . Выброс гормонов адреналина и норадреналина из мозгового слоя является надпочечников частью реакции «бей или беги» . [3]
Тирозин создается из фенилаланина путем гидроксилирования ферментом фенилаланингидроксилазой . Тирозин также поступает непосредственно из пищевого белка. Клетки, секретирующие катехоламины, используют несколько реакций для последовательного преобразования тирозина в L -ДОФА, а затем в дофамин. В зависимости от типа клеток дофамин может превращаться в норадреналин или даже в адреналин. [4]
Различные стимулирующие препараты (например, ряд замещенных амфетаминов ) являются аналогами катехоламинов.
Структура
[ редактировать ]Катехоламины имеют четкую структуру бензольного кольца с двумя гидроксильными группами, промежуточной этильной цепью и концевой аминогруппой . Фенилэтаноламины, такие как норэпинефрин, имеют гидроксильную группу в этильной цепи. [ нужна ссылка ]
Производство и деградация
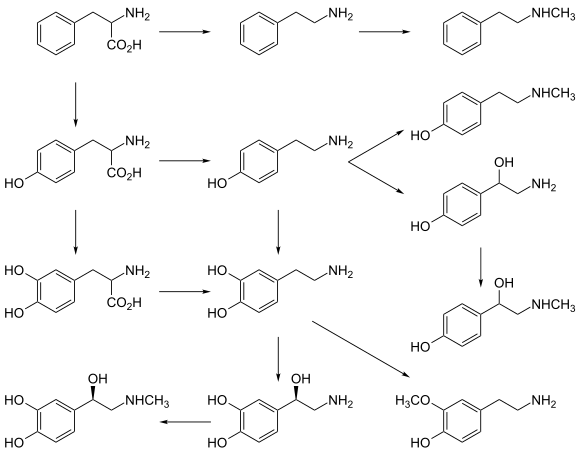
[ редактировать ]L -фенилаланин превращается в L -тирозин под действием фермента гидроксилазы ароматических аминокислот (АААН) ( фенилаланин-4-гидроксилазы ) с молекулярным кислородом (O 2 ) и тетрагидробиоптерином в качестве кофакторов . L -тирозин превращается в L -ДОФА другим ферментом АААХ ( тирозин-3-гидроксилазой ) с тетрагидробиоптерином , O 2 и двухвалентным железом (Fe 2+ ) в качестве кофакторов. L -ДОФА превращается в дофамин под действием фермента декарбоксилазы ароматических L -аминокислот (AADC) с пиридоксальфосфатом в качестве кофактора. Сам дофамин также используется в качестве предшественника в синтезе нейромедиаторов норадреналина и адреналина . Дофамин превращается в норадреналин под действием фермента дофамин-β-гидроксилазы (DBH) с участием O 2 и L -аскорбиновой кислоты в качестве кофакторов. Норадреналин превращается в адреналин под действием фермента фенилэтаноламин -N -метилтрансферазы (ПНМТ) с S -аденозил- L -метионином в качестве кофактора. |
Расположение
[ редактировать ]Катехоламины продуцируются главным образом хромаффинными клетками и мозгового слоя надпочечников постганглионарными волокнами симпатической нервной системы . Дофамин , который действует как нейромедиатор в центральной нервной системе , в основном вырабатывается в телах нейронов в двух областях ствола мозга: вентральной покрышке и черной субстанции , последняя из которых содержит нейромеланин -пигментированные нейроны. пигментированные аналогичным нейромеланином, Тела клеток голубого пятна, производят норадреналин . Адреналин вырабатывается в небольших группах нейронов головного мозга человека, которые экспрессируют синтезирующий его фермент фенилэтаноламин -N -метилтрансферазу ; [8] эти нейроны отходят от ядра, прилежащего (вентролатерально) к постремной области, и от ядра в дорсальной области одиночного тракта . [8]
Биосинтез
[ редактировать ]Дофамин – первый катехоламин, синтезированный из ДОФА. В свою очередь, норадреналин и адреналин образуются в результате дальнейшей метаболической модификации дофамина. Ферменту дофамингидроксилазе требуется медь в качестве кофактора (не показана на схеме), а ДОФА-декарбоксилазе требуется PLP (не показан на схеме). Стадией, ограничивающей скорость биосинтеза катехоламинов по преобладающему метаболическому пути, является гидроксилирование L -тирозина до L -ДОФА. [ нужна ссылка ]
Синтез катехоламинов ингибируется альфа-метил- п -тирозином ( АМРТ ), который ингибирует тирозингидроксилазу . [ нужна ссылка ]
Аминокислоты фенилаланин и тирозин являются предшественниками катехоламинов. Обе аминокислоты обнаруживаются в высоких концентрациях в плазме крови и мозге. У млекопитающих тирозин может образовываться из пищевого фенилаланина с помощью фермента фенилаланингидроксилазы , обнаруженного в больших количествах в печени. Недостаточное количество фенилаланингидроксилазы приводит к фенилкетонурии , метаболическому расстройству, которое приводит к умственному дефициту, если его не лечить диетическими манипуляциями. [ нужна ссылка ] Обычно считается, что синтез катехоламинов начинается с тирозина. Фермент тирозингидроксилаза (ТН) превращает аминокислоту L -тирозин в 3,4-дигидроксифенилаланин ( L -ДОФА). Гидроксилирование L -тирозина ТГ приводит к образованию предшественника DA L -ДОФА, который метаболизируется декарбоксилазой ароматических L -аминокислот (AADC; см. Cooper et al., 2002). [ нужна ссылка ] ) к передатчику дофамина. Этот этап происходит настолько быстро, что трудно измерить уровень L -ДОФА в мозге без предварительного ингибирования AADC. [ нужна ссылка ] В нейронах , которые используют ДА в качестве передатчика, декарбоксилирование L -ДОФА до дофамина является заключительным этапом формирования передатчика; однако в тех нейронах, которые используют норадреналин (норадреналин) или адреналин (адреналин) в качестве передатчиков, также присутствует фермент дофамин-β-гидроксилаза (DBH), который превращает дофамин в норадреналин. В других нейронах, в которых передатчиком является адреналин, третий фермент фенилэтаноламин- N -метилтрансфераза (PNMT) превращает норадреналин в адреналин. Таким образом, клетка, использующая адреналин в качестве медиатора, содержит четыре фермента (TH, AADC, DBH и PNMT), тогда как норадреналиновые нейроны содержат только три фермента (без PNMT), а дофаминовые клетки - только два (TH и AADC). [ нужна ссылка ]
Деградация
[ редактировать ]Катехоламины имеют период полураспада в несколько минут при циркуляции в крови. Они могут разлагаться либо путем метилирования катехол -О -метилтрансферазами (КОМТ), либо путем дезаминирования моноаминоксидазами (МАО) .
ИМАО связываются с МАО, тем самым предотвращая расщепление катехоламинов и других моноаминов.
Катаболизм катехоламинов опосредован двумя основными ферментами: катехол- О -метилтрансферазой (КОМТ), которая присутствует в синаптической щели и цитозоле клетки, и моноаминоксидазой (МАО), которая локализована в митохондриальной мембране. Оба фермента требуют кофакторов: COMT использует Mg. 2+ в качестве кофактора, тогда как МАО использует FAD . Первый этап катаболического процесса опосредован либо МАО, либо КОМТ, что зависит от ткани и местоположения катехоламинов (например, деградация катехоламинов в синаптической щели опосредуется КОМТ, поскольку МАО является митохондриальным ферментом). Следующие этапы катаболизма включают алкогольдегидрогеназу , альдегиддегидрогеназу и альдегидредуктазу . Конечным продуктом адреналина и норадреналина является ванилилминдальная кислота (ВМА), которая выводится с мочой . Катаболизм дофамина приводит к выработке гомованилиновой кислоты (ГВА) . [9]
Функция
[ редактировать ]модальность
[ редактировать ]Два катехоламина, норадреналин и дофамин , действуют как нейромодуляторы в центральной нервной системе и как гормоны в кровообращении. Катехоламин норадреналин является нейромодулятором периферической симпатической нервной системы, но также присутствует в крови (в основном за счет «перехода» из синапсов симпатической системы). [ нужна ссылка ]
Высокие уровни катехоламинов в крови связаны со стрессом , который может быть вызван психологическими реакциями или стрессорами окружающей среды, такими как повышенный уровень звука , яркий свет или низкий уровень сахара в крови . [10]
Чрезвычайно высокие уровни катехоламинов (также известные как токсичность катехоламинов) могут возникать при травмах центральной нервной системы из-за стимуляции или повреждения ядер ствола мозга , в частности тех ядер, которые влияют на симпатическую нервную систему . В неотложной медицине это явление широко известно как «свалка катехоламинов».
Чрезвычайно высокий уровень катехоламинов может быть также вызван нейроэндокринными опухолями мозгового слоя надпочечников – излечимым заболеванием, известным как феохромоцитома .
Высокий уровень катехоламинов также может быть вызван дефицитом моноаминоксидазы А (МАО-А) , известным как синдром Бруннера . Поскольку МАО-А является одним из ферментов, ответственных за деградацию этих нейротрансмиттеров, его дефицит значительно увеличивает биодоступность этих нейротрансмиттеров. Встречается при отсутствии феохромоцитомы , нейроэндокринных опухолей и карциноидного синдрома , но внешне похож на карциноидный синдром с такими симптомами, как покраснение лица и агрессия. [11] [12]
Острая порфирия может вызывать повышение уровня катехоламинов. [13]
Эффекты
[ редактировать ]Катехоламины вызывают общие физиологические изменения, которые готовят организм к физической активности ( реакция «бей или беги» ). Некоторыми типичными эффектами являются увеличение частоты сердечных сокращений , артериального давления , уровня глюкозы в крови и общая реакция симпатической нервной системы . [ нужна ссылка ] Некоторые препараты, например толкапон (центральный ингибитор КОМТ ), повышают уровень всех катехоламинов. учащение дыхания ( тахипноэ ). Повышенный уровень катехоламинов может также вызвать у пациентов [14]
Катехоламин выделяется в мочу после расщепления, и уровень его секреции можно измерить для диагностики заболеваний, связанных с уровнем катехоламинов в организме. [15] Анализ мочи на катехоламины используется для выявления феохромоцитомы .
Функция у растений
[ редактировать ]«Они были обнаружены в 44 семействах растений, но для них не установлено никакой важной метаболической функции. Они являются предшественниками бензо[ c ]фенантридиновых алкалоидов , которые являются основными активными ингредиентами многих экстрактов лекарственных растений . Возможная защитная роль против насекомых-хищников, травм и детоксикации азота. Было показано, что они способствуют росту тканей растений, соматическому эмбриогенезу в культурах in vitro и цветению. Они ингибируют индол-3-уксусной кислоты окисление и усиливают биосинтез этилена . также было показано, что они синергически усиливают различные эффекты гиббереллинов ». [16]
Тестирование на катехоламины
[ редактировать ]Катехоламины секретируются клетками тканей различных систем организма человека, преимущественно нервной и эндокринной систем. Надпочечники выделяют в кровь определенные катехоламины, когда человек испытывает физический или умственный стресс, и это обычно здоровая физиологическая реакция. [ нужна ссылка ] Однако острый или хронический избыток циркулирующих катехоламинов потенциально может повысить кровяное давление и частоту сердечных сокращений до очень высоких уровней и в конечном итоге спровоцировать опасные последствия. Тесты на свободные метанефрины фракционированной плазмы или метанефрины мочи используются для подтверждения или исключения определенных заболеваний, когда врач выявляет признаки гипертонии и тахикардии , которые неадекватно реагируют на лечение. [17] [18] Каждый из тестов измеряет количество метаболитов адреналина и норадреналина, называемых соответственно метанефрином и норметанефрином .
Также проводятся анализы крови для анализа количества катехоламинов, присутствующих в организме.
Тесты на катехоламины проводятся для выявления редких опухолей надпочечников или нервной системы. Тесты на катехоламины предоставляют информацию о таких опухолях, как феохромоцитома, параганглиома и нейробластома. [19] [20]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Фицджеральд, Пенсильвания (2011). «Глава 11. Мозговое вещество надпочечников и параганглии» . В Гарднере, Д.Г.; Шобак, Д. (ред.). Базовая и клиническая эндокринология Гринспена (9-е изд.). Нью-Йорк: МакГроу-Хилл . Проверено 26 октября 2011 г.
- ^ Первс, Д.; Августин, Дж.Дж.; Фитцпатрик, Д.; Холл, туалет; ЛаМантия, AS; Макнамара, Джо; Уайт, Л.Е., ред. (2008). Нейронаука (4-е изд.). Синауэр Ассошиэйтс. стр. 137–138. ISBN 978-0-87893-697-7 .
- ^ «Катехоламины» . Библиотека здоровья . Сан-Диего, Калифорния: Калифорнийский университет. Архивировано из оригинала 16 июля 2011 года.
- ^ Джо, TH; Хван, О. (1987). «Дофамин-бета-гидроксилаза: биохимия и молекулярная биология». Анналы Нью-Йоркской академии наук . 493 : 342–350. дои : 10.1111/j.1749-6632.1987.tb27217.x . ПМИД 3473965 . S2CID 86229251 .
- ^ Бродли К.Дж. (март 2010 г.). «Сосудистые эффекты следовых аминов и амфетаминов». Фармакология и терапия . 125 (3): 363–375. doi : 10.1016/j.pharmthera.2009.11.005 . ПМИД 19948186 .
- ^ Линдеманн Л., Хонер MC (май 2005 г.). «Ренессанс следовых аминов, вдохновленный новым семейством GPCR». Тенденции в фармакологических науках . 26 (5): 274–281. дои : 10.1016/j.tips.2005.03.007 . ПМИД 15860375 .
- ^ Ван X, Ли Дж, Донг Дж, Юэ Дж (февраль 2014 г.). «Эндогенные субстраты CYP2D мозга». Европейский журнал фармакологии . 724 : 211–218. дои : 10.1016/j.ejphar.2013.12.025 . ПМИД 24374199 .
- ^ Jump up to: а б Китахама, К.; Пирсон, Дж.; Денорой, Л.; Копп, Н.; Ульрих, Дж.; Маэда, Т.; Жуве, М. (1985). «Адренергические нейроны в мозге человека, продемонстрированные с помощью иммуногистохимии с антителами к фенилэтаноламин -N -метилтрансферазе (PNMT): открытие новой группы в ядре одиночного тракта». Письма по неврологии . 53 (3): 303–308. дои : 10.1016/0304-3940(85)90555-5 . ПМИД 3885079 . S2CID 2578817 .
- ^ Эйзенхофер, Г.; Копин, Эй-Джей; Гольдштейн, Д.С. (2004). «Метаболизм катехоламинов: современный взгляд с последствиями для физиологии и медицины». Фармакологические обзоры . 3 (56): 331–349. дои : 10.1124/пр.56.3.1 . ПМИД 15317907 . S2CID 12825309 .
- ^ Чу, Брианна; Марваха, Комал; Санвикторес, Терренс; Авосика, Айула О.; Айерс, Дерек (2024 г.), «Физиология, реакция на стресс» , StatPearls , Остров сокровищ (Флорида): StatPearls Publishing, PMID 31082164 , получено 28 июня 2024 г.
- ^ Мэнор, И.; Тяно, С.; Мел, Э.; Айзенберг, Дж.; Бахнер-Мельман, Р.; Котлер, М.; Эбштейн, Р.П. (2002). «Семейные и ассоциативные исследования моноаминоксидазы А и синдрома дефицита внимания с гиперактивностью (СДВГ): преимущественная передача длинного повтора промоторной области и ее связь с ухудшением производительности в тесте непрерывной производительности (TOVA)» . Молекулярная психиатрия . 7 (6): 626–632. дои : 10.1038/sj.mp.4001037 . ПМИД 12140786 .
- ^ Бруннер, Х.Г. (1996). «Дефицит MAOA и аномальное поведение: перспективы ассоциации». Генетика криминального и антисоциального поведения . Симпозиум Фонда Ciba. Том. 194. Уайли. стр. 155–167. дои : 10.1002/9780470514825.ch9 . ISBN 978-0-470-51482-5 . ПМИД 8862875 .
- ^ Стюарт, МФ; Крофт, Дж.; Рид, П.; Нью, JP (2006). «Острая интермиттирующая порфирия и феохромоцитома: общие черты» . Журнал клинической патологии . 60 (8): 935–936. дои : 10.1136/jcp.2005.032722 . ЧВК 1994495 . ПМИД 17660335 .
- ^ Эстес, Мэри (2016). Оценка здоровья и медицинский осмотр (2-е изд.). Мельбурн : Сенгадж . п. 143. ИСБН 978-0-17-035484-4 .
- ^ «Катехоламины в моче» . webmd.com . Проверено 4 мая 2017 г.
- ^ Куклин А.И.; Конгер, Б.В. (1995). «Катехоламины в растениях». Журнал регулирования роста растений . 14 (2): 91–97. дои : 10.1007/BF00203119 . S2CID 41493767 .
- ^ «Метанефрины без плазмы | Лабораторные тесты онлайн» . labtestsonline.org . Проверено 24 декабря 2019 г.
- ^ «Метанефрины в моче | Лабораторные тесты онлайн» . labtestsonline.org . 6 декабря 2019 года . Проверено 24 декабря 2019 г.
- ^ «Анализ мочи и крови на катехоламины» . ВебМД . Проверено 9 октября 2019 г.
- ^ «Катехоламины» . labtestsonline.org . Проверено 9 октября 2019 г.
Внешние ссылки
[ редактировать ]- Катехоламины Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|