Человеческий мозг
| Человеческий мозг | |
|---|---|
 Человеческий мозг, полученный после вскрытия | |
 Человеческий мозг и череп | |
| Подробности | |
| Предшественник | Нервная трубка |
| Система | Центральная нервная система |
| Артерия | Внутренние сонные артерии , позвоночные артерии |
| вена | Внутренняя яремная вена , внутренние мозговые вены ; Наружные вены: ( верхние , средние и нижние мозговые вены ), базальные вены и мозжечковые вены. |
| Идентификаторы | |
| латинский | головной мозг |
| Греческий | энкефалос [1] |
| ТА98 | A14.1.03.001 |
| ТА2 | 5415 |
| ФМА | 50801 |
| Анатомическая терминология | |
является Головной мозг центральным органом человека нервной системы и вместе со спинным мозгом составляет центральную нервную систему . Головной мозг состоит из головного мозга , ствола мозга и мозжечка . Он контролирует большую часть деятельности тела , обрабатывая, интегрируя и координируя информацию, которую он получает от органов чувств , и принимая решения относительно инструкций, посылаемых остальной части тела. Мозг находится внутри костей черепа и защищен ими .
Головной мозг, самая большая часть человеческого мозга, состоит из двух полушарий головного мозга . Каждое полушарие имеет внутреннее ядро, состоящее из белого вещества , и внешнюю поверхность – кору головного мозга – состоящую из серого вещества . Кора имеет внешний слой, неокортекс , и внутренний аллокортекс . Неокортекс состоит из шести слоев нейронов , а аллокортекс — из трех или четырех. Каждое полушарие разделено на четыре доли — лобную , височную , теменную и затылочную . Лобная доля связана с исполнительными функциями, включая самоконтроль , планирование , рассуждение и абстрактное мышление , а затылочная доля отвечает за зрение. Внутри каждой доли корковые области связаны с определенными функциями, такими как сенсорные , двигательные и ассоциативные области. Хотя левое и правое полушария во многом схожи по форме и функциям, некоторые функции связаны с одной стороной , например, речь — слева, а зрительно-пространственные способности — справа. Полушария соединены между собой комиссуральные нервные пути , самый крупный из которых — мозолистое тело .
Головной мозг соединен стволом мозга со спинным мозгом. Ствол мозга состоит из среднего мозга , моста и продолговатого мозга . Мозжечок называемых соединен со стволом мозга тремя парами нервных путей, ножками мозжечка . Внутри головного мозга находится желудочковая система , состоящая из четырех соединенных между собой желудочков , в которых спинномозговая жидкость вырабатывается и циркулирует . Под корой головного мозга находится несколько важных структур, в том числе таламус , эпиталамус , шишковидная железа , гипоталамус , гипофиз и субталамус ; лимбические структуры , включая миндалевидное тело и гиппокамп , Клауструм , различные ядра базальных ганглиев , базальные структуры переднего мозга и три околожелудочковых органа . Структуры мозга, не находящиеся в средней плоскости, существуют парами, например, имеются два гиппокампа и две миндалевидные тела. Клетки головного мозга включают нейроны и поддерживающие глиальные клетки . В мозге более 86 миллиардов нейронов и примерно такое же количество других клеток. Мозговая деятельность становится возможной благодаря взаимосвязи нейронов и высвобождению ими нейромедиаторов в ответ на нервные импульсы . Нейроны соединяются, образуя нервные пути , нейронные цепи и сложные сетевые системы . Вся схема управляется процессом нейротрансмиссии .
Мозг защищен черепом , находится в спинномозговой жидкости и изолирован от кровотока гематоэнцефалическим барьером . Однако мозг по-прежнему подвержен повреждениям , болезням и инфекциям . Повреждение может быть вызвано травмой или нарушением кровоснабжения, известным как инсульт . Мозг подвержен дегенеративным расстройствам , таким как болезнь Паркинсона , деменция, включая болезнь Альцгеймера , и рассеянный склероз . Считается, что психические заболевания , включая шизофрению и клиническую депрессию , связаны с дисфункциями головного мозга. Мозг также может быть местом возникновения опухолей , как доброкачественных , так и злокачественных ; они в основном происходят из других участков тела .
Изучением анатомии мозга занимается нейроанатомия , а изучением его функций — нейробиология . Для изучения мозга используются многочисленные методы. Образцы других животных, которые можно исследовать под микроскопом , традиционно дают много информации. Технологии медицинской визуализации , такие как функциональная нейровизуализация и записи электроэнцефалографии (ЭЭГ), важны для изучения мозга. История болезни людей с черепно-мозговой травмой позволила лучше понять функцию каждой части мозга. Неврологические исследования значительно расширились, и исследования продолжаются.
В культуре философия разума на протяжении веков пыталась решить вопрос о природе сознания и проблеме разума и тела . Псевдонаука веке френология . попыталась локализовать свойства личности в областях коры головного мозга в 19 В научной фантастике трансплантация мозга представлена в таких рассказах, как « Мозг Донована» 1942 года .
Структура [ править ]

Общая анатомия [ править ]
Мозг взрослого человека весит в среднем около 1,2–1,4 кг (2,6–3,1 фунта), что составляет около 2% от общей массы тела. [2] [3] объемом около 1260 см. 3 у мужчин и 1130 см. 3 у женщин. [4] Существуют значительные индивидуальные различия, [4] стандартный референсный диапазон для мужчин составляет 1180–1620 г (2,60–3,57 фунта). [5] а для женщин 1030–1400 г (2,27–3,09 фунта). [6]
Головной мозг , состоящий из полушарий головного мозга , образует самую большую часть головного мозга и покрывает другие структуры мозга. [7] область полушарий, кора головного мозга , представляет собой серое вещество , состоящее из корковых слоев нейронов Внешняя . Каждое полушарие разделено на четыре основные доли – лобную долю , теменную долю , височную долю и затылочную долю . [8] Некоторые источники включают еще три доли: центральную долю , лимбическую долю и островковую долю . [9] Центральная доля включает в себя прецентральную извилину и постцентральную извилину и включена в нее, поскольку выполняет особую функциональную роль. [9] [10]
Ствол мозга , напоминающий стебель, прикрепляется к головному мозгу и выходит из него в начале области среднего мозга . Ствол мозга включает средний мозг, мост и продолговатый мозг . За стволом мозга находится мозжечок ( лат . «маленький мозг» ). [7]
Головной мозг, ствол мозга, мозжечок и спинной мозг покрыты четырьмя [11] оболочки, называемые мозговыми оболочками . Мембраны представляют собой прочную твердую мозговую оболочку ; средняя паутинная оболочка и более нежная внутренняя мягкая мозговая оболочка . Между паутинной и мягкой мозговой оболочкой находится субарахноидальное пространство и субарахноидальные цистерны , содержащие спинномозговую жидкость . [12] Наружной мембраной коры головного мозга является базальная мембрана мягкой мозговой оболочки, называемая ограничивающей глией , и она является важной частью гематоэнцефалического барьера . [13] Живой мозг очень мягкий и имеет гелеобразную консистенцию, похожую на мягкий тофу. [14] Корковые слои нейронов составляют большую часть серого вещества головного мозга , тогда как более глубокие подкорковые области миелинизированных аксонов составляют белое вещество . [7] Белое вещество головного мозга составляет около половины всего объема мозга. [15]

Cerebrum[editмозг
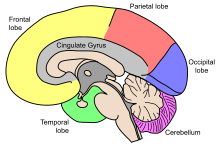
Головной мозг — самая большая часть головного мозга, разделенная на почти симметричные левое и правое полушария глубокой бороздой — продольной щелью . [16] Асимметрия между долями отмечается в виде петалий . [17] Полушария соединены пятью спайками , охватывающими продольную щель, самая крупная из них — мозолистое тело . [7] Каждое полушарие условно делится на четыре основные доли ; лобная доля , теменная доля , височная доля и затылочная доля , названные в честь костей черепа , которые над ними лежат. [8] Каждая доля связана с одной или двумя специализированными функциями, хотя между ними есть некоторое функциональное перекрытие. [18] Поверхность головного мозга сложена на гребни ( извилины ) и бороздки ( борозды ), многие из которых называются, обычно в зависимости от их положения, например, лобная извилина лобной доли или центральная борозда, разделяющая центральные области полушарий. . Существует множество небольших вариаций вторичных и третичных складок. [19]
Внешняя часть головного мозга — это кора головного мозга , состоящая из серого вещества, расположенного слоями. Он имеет толщину от 2 до 4 миллиметров (от 0,079 до 0,157 дюйма) и глубоко сложен, что придает извилистый вид. [20] Под корой находится белое вещество головного мозга . Самой большой частью коры головного мозга является неокортекс , имеющий шесть слоев нейронов. Остальная часть коры представляет собой аллокортекс , имеющий три или четыре слоя. [7]
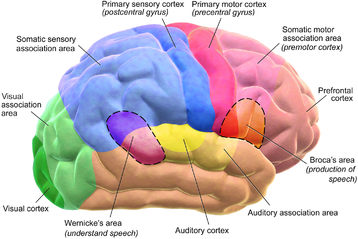
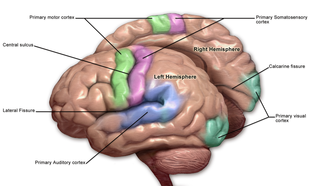
Кора разделена примерно на пятьдесят различных функциональных областей, известных как зоны Бродмана . Эти области отчетливо различаются, если смотреть под микроскопом . [21] Кора разделена на две основные функциональные области — моторную кору и сенсорную кору . [22] Первичная моторная кора , которая посылает аксоны вниз к мотонейронам ствола головного мозга и спинного мозга, занимает заднюю часть лобной доли, непосредственно перед соматосенсорной областью. Первичные сенсорные области сигналы от сенсорных нервов и трактов через релейные ядра таламуса получают . К первичным сенсорным областям относятся зрительная кора затылочной доли , слуховая кора в частях височной доли и островковой коры , соматосенсорная кора в теменной доле . Остальные части коры называются ассоциативными областями . Эти области получают информацию от сенсорных областей и нижних отделов мозга и участвуют в сложных когнитивных процессах восприятия , принятия мышления и решений . [23] Основные функции лобной доли — контроль внимания , абстрактного мышления, поведения, решения задач, а также физических реакций и личности. [24] [25] Затылочная доля - самая маленькая доля; его основные функции — зрительный прием, зрительно-пространственная обработка, движение и распознавание цвета . [24] [25] В доле имеется меньшая затылочная долька, известная как кунеус . Височная доля контролирует слуховую и зрительную память , язык , а также некоторые слух и речь. [24]
Головной мозг содержит желудочки , в которых вырабатывается и циркулирует спинномозговая жидкость. Ниже мозолистого тела находится прозрачная перегородка , мембрана, разделяющая боковые желудочки . Под боковыми желудочками находится таламус , а впереди и ниже — гипоталамус . Гипоталамус ведет к гипофизу . В задней части таламуса находится ствол мозга. [26]
Базальные ганглии , также называемые базальными ядрами, представляют собой набор структур глубоко внутри полушарий, участвующих в регуляции поведения и движений. [27] Самый крупный компонент — полосатое тело , другие — бледный шар , черная субстанция и субталамическое ядро . [27] Полосатое полосатое тело делится на вентральное полосатое тело и дорсальное полосатое тело, подразделения которых основаны на функциях и связях. Вентральное полосатое тело состоит из прилежащего ядра и обонятельного бугорка , тогда как дорсальное полосатое тело состоит из хвостатого ядра и скорлупы . Спутник и бледный шар отделены от боковых желудочков и таламуса внутренней капсулой , тогда как хвостатое ядро простирается вокруг боковых желудочков и примыкает к ним с внешней стороны. [28] В самой глубокой части латеральной борозды между островковой корой и полосатым телом находится тонкий слой нейронов, называемый клауструмом . [29]
Ниже и впереди полосатого тела находится ряд базальных структур переднего мозга . К ним относятся базальное ядро , диагональная полоска Брока , безымянная субстанция и медиальное перегородочное ядро . структуры играют важную роль в выработке нейромедиатора ацетилхолина Эти , который затем широко распределяется по всему мозгу. Базальный отдел переднего мозга, в частности базальное ядро, считается основным холинергическим продуктом центральной нервной системы, направляемым в полосатое тело и неокортекс. [30]
Cerebellum[editмозжечок
Мозжечок делится на переднюю долю , заднюю долю и флокулонодулярную долю . [31] Передняя и задняя доли соединены посередине червем . [32] По сравнению с корой головного мозга мозжечок имеет гораздо более тонкую внешнюю кору, изрезанную узкими бороздками и многочисленными изогнутыми поперечными щелями. [32] Если смотреть снизу, между двумя долями находится третья доля, флоккулонодулярная доля. [33] Мозжечок лежит в задней части полости черепа , под затылочными долями, и отделен от них наметом мозжечка , слоем волокон. [34]
Он связан со стволом мозга тремя парами нервных путей, называемых ножками мозжечка . Высшая пара соединяется со средним мозгом; средняя пара соединяется с продолговатым мозгом, а нижняя пара соединяется с мостом. [32] Мозжечок состоит из внутреннего мозгового вещества белого вещества и внешней коры из богато складчатого серого вещества. [34] Передняя и задняя доли мозжечка, по-видимому, играют роль в координации и сглаживании сложных двигательных движений, а флокулонодулярная доля - в поддержании равновесия. [35] хотя существуют споры относительно его когнитивных, поведенческих и двигательных функций. [36]
Ствол мозга [ править ]
Ствол мозга лежит под головным мозгом и состоит из среднего мозга , моста и продолговатого мозга . Он лежит в задней части черепа , опираясь на часть основания, известную как скат , и заканчивается у большого затылочного отверстия , большого отверстия в затылочной кости . Ствол мозга продолжается ниже спинного мозга . [37] защищен позвоночником .
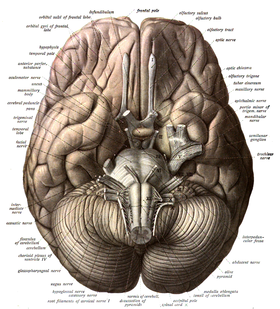
Десять из двенадцати пар черепно-мозговых нервов [а] выходят непосредственно из ствола мозга. [37] Ствол мозга также содержит множество ядер черепных нервов и ядер , периферических нервов а также ядер, участвующих в регуляции многих важнейших процессов, включая дыхание , контроль движений глаз и равновесия. [38] [37] Ретикулярная формация , сеть ядер нечеткой формации, присутствует внутри и по всей длине ствола мозга. [37] Через ствол мозга проходит множество нервных путей , передающих информацию в кору головного мозга и обратно к остальным частям тела. [37]
Микроанатомия [ править ]
Человеческий мозг в основном состоит из нейронов , глиальных клеток , нервных стволовых клеток и кровеносных сосудов . Типы нейронов включают вставочные нейроны , пирамидные клетки, включая клетки Бетца , мотонейроны ( верхние и нижние мотонейроны ) и клетки Пуркинье мозжечка . Клетки Беца — самые крупные клетки (по размеру клеточного тела) нервной системы. [39] По оценкам, мозг взрослого человека содержит 86±8 миллиардов нейронов и примерно такое же количество (85±10 миллиардов) ненейрональных клеток. [40] Из этих нейронов 16 миллиардов (19%) расположены в коре головного мозга, а 69 миллиардов (80%) — в мозжечке. [3] [40]
Типами глиальных клеток являются астроциты (включая глию Бергмана ), олигодендроциты , эпендимальные клетки (включая танициты ), радиальные глиальные клетки , микроглия и подтип клеток-предшественников олигодендроцитов . Астроциты — самые крупные из глиальных клеток. Это звездчатые клетки со множеством отростков, исходящих от их клеточных тел . Некоторые из этих процессов заканчиваются периваскулярными окончаниями на стенках капилляров . [41] коры Ограниченная глия состоит из отростков ножек астроцитов, которые частично служат для содержания клеток головного мозга. [13]
Тучные клетки — это лейкоциты , которые взаимодействуют с нейроиммунной системой головного мозга. [42] Тучные клетки центральной нервной системы присутствуют во многих структурах, включая мозговые оболочки; [42] они опосредуют нейроиммунные реакции при воспалительных состояниях и помогают поддерживать гематоэнцефалический барьер, особенно в тех областях мозга, где барьер отсутствует. [42] [43] Тучные клетки выполняют одни и те же общие функции в организме и центральной нервной системе, такие как воздействие или регулирование аллергических реакций, врожденного и адаптивного иммунитета , аутоиммунитета и воспаления . [42] Тучные клетки служат основной эффекторной клеткой , посредством которой патогены могут влиять на биохимическую передачу сигналов, происходящую между желудочно-кишечным трактом и центральной нервной системой . [44] [45]
около 400 генов Показано, что специфичны для мозга. Во всех нейронах ELAVL3 экспрессируется NRGN и REEP2 , а в пирамидных клетках также экспрессируются . GAD1 , необходимый для биосинтеза нейромедиатора ГАМК , экспрессируется в интернейронах. Белки, экспрессируемые в глиальных клетках, включают маркеры астроцитов GFAP и S100B, тогда как основной белок миелина и фактор транскрипции OLIG2 экспрессируются в олигодендроцитах. [46]
Цереброспинальная жидкость [ править ]

Цереброспинальная жидкость — прозрачная бесцветная трансклеточная жидкость , циркулирующая вокруг головного мозга в субарахноидальном пространстве , в желудочковой системе и в центральном канале спинного мозга. Он также заполняет некоторые пробелы в субарахноидальном пространстве, известные как субарахноидальные цистерны . [47] Четыре желудочка (два боковых , третий и четвертый) содержат сосудистое сплетение , вырабатывающее спинномозговую жидкость. [48] Третий желудочек расположен по средней линии и соединяется с боковыми желудочками. [47] Единственный проток , церебральный водопровод между мостом и мозжечком, соединяет третий желудочек с четвертым желудочком. [49] Три отдельных отверстия, среднее и два боковых , отводят спинномозговую жидкость из четвертого желудочка в большую цистерну , одну из главных цистерн. Отсюда спинномозговая жидкость циркулирует вокруг головного и спинного мозга в субарахноидальном пространстве, между паутинной и мягкой мозговой оболочкой. [47] В любой момент времени имеется около 150 мл спинномозговой жидкости, большая часть которой находится в субарахноидальном пространстве. Он постоянно регенерируется и усваивается и заменяется примерно раз в 5–6 часов. [47]
. глимфатическая система Описана [50] [51] [52] как лимфодренажная система головного мозга. Глимфатический путь, охватывающий весь мозг, включает пути оттока спинномозговой жидкости и менингеальных лимфатических сосудов , которые связаны с синусами твердой мозговой оболочки и проходят рядом с кровеносными сосудами головного мозга. [53] [54] Путь отводит интерстициальную жидкость из ткани головного мозга. [54]
Кровоснабжение [ править ]


Внутренние сонные артерии снабжают кровью, насыщенной кислородом , переднюю часть мозга, а позвоночные артерии снабжают кровью заднюю часть мозга. [55] Эти два кровообращения соединяются в Виллизиев круг , кольцо связанных артерий, которое лежит в межножковой цистерне между средним мозгом и мостом. [56]
Внутренние сонные артерии являются ветвями общих сонных артерий . Они проникают в череп через сонный канал , проходят через кавернозный синус и попадают в субарахноидальное пространство . [57] Затем они входят в виллизиев круг с двумя ветвями, от которых выходят передние мозговые артерии . Эти ветви идут вперед, а затем вверх по продольной щели и снабжают кровью переднюю и срединную части мозга. [58] Одна или несколько мелких передних соединительных артерий присоединяются к двум передним мозговым артериям вскоре после того, как они образуют ветви. [58] Внутренние сонные артерии продолжаются вперед как средние мозговые артерии . Они проходят вбок вдоль клиновидной кости , глазницы затем вверх через кору островка , где возникают конечные ветви. Средние мозговые артерии отдают ветви по всей своей длине. [57]
Позвоночные артерии отходят от левой и правой подключичных артерий . Они проходят вверх через поперечные отверстия , которые представляют собой пространства в шейных позвонках . Каждая сторона входит в полость черепа через большое затылочное отверстие вдоль соответствующей стороны продолговатого мозга. [57] Они отдают одну из трех ветвей мозжечка . Позвоночные артерии соединяются перед средней частью продолговатого мозга, образуя более крупную базилярную артерию , которая дает несколько ветвей, кровоснабжающих продолговатый мозг и мост, а также две другие переднюю и верхнюю мозжечковые ветви . [59] Наконец, базилярная артерия разделяется на две задние мозговые артерии . Они направляются наружу, вокруг верхних ножек мозжечка и вдоль верхушки намета мозжечка, откуда отдают ветви для кровоснабжения височной и затылочной долей. [59] Каждая задняя мозговая артерия отдает небольшую заднюю соединительную артерию, которая соединяется с внутренними сонными артериями.
Дренаж крови [ править ]
Церебральные вены отводят дезоксигенированную кровь из мозга. Мозг имеет две основные сети вен : внешнюю, или поверхностную , на поверхности головного мозга, имеющую три ветви, и внутреннюю сеть . Эти две сети сообщаются через анастомозирующие (соединяющиеся) вены. [60] Вены головного мозга впадают в более крупные полости дуральных венозных синусов, обычно расположенных между твердой мозговой оболочкой и оболочкой черепа. [61] Кровь от мозжечка и среднего мозга оттекает в большую мозговую вену . Кровь из продолговатого мозга и моста ствола мозга имеет различный характер оттока: либо в спинномозговые вены , либо в соседние мозговые вены. [60]
Кровь в глубокой части мозга оттекает через венозное сплетение в кавернозный синус спереди, верхний и нижний каменистые синусы по бокам и нижний сагиттальный синус сзади. [61] Кровь оттекает из наружного мозга в большой верхний сагиттальный синус , который находится на средней линии верхней части мозга. Кровь отсюда соединяется с кровью из прямого синуса при впадении синусов . [61]
Кровь отсюда оттекает в левый и правый поперечные синусы . [61] Затем они стекают в сигмовидные синусы , куда поступает кровь из кавернозного синуса, а также верхних и нижних каменистых синусов. Сигмовидная впадает в крупные внутренние яремные вены . [61] [60]
Гематоэнцефалический барьер [ править ]
Более крупные артерии головного мозга снабжают кровью более мелкие капилляры . Эти мельчайшие кровеносные сосуды головного мозга выстланы клетками, соединенными плотными соединениями , поэтому жидкости не просачиваются и не вытекают в той же степени, как в других капиллярах; это создает гематоэнцефалический барьер . [43] Перициты играют важную роль в формировании плотных контактов. [62] Барьер менее проницаем для более крупных молекул, но все же проницаем для воды, углекислого газа, кислорода и большинства жирорастворимых веществ (включая анестетики и алкоголь). [43] Гематоэнцефалический барьер отсутствует в околожелудочковых органах (которые представляют собой структуры головного мозга, которым может потребоваться реагировать на изменения в жидкостях организма), таких как шишковидная железа , область пострема и некоторые области гипоталамуса . [43] Существует аналогичный барьер гемато-спинномозговой жидкости , который служит той же цели, что и гематоэнцефалический барьер, но облегчает транспорт различных веществ в мозг из-за различных структурных характеристик между двумя барьерными системами. [43] [63]
Развитие [ править ]
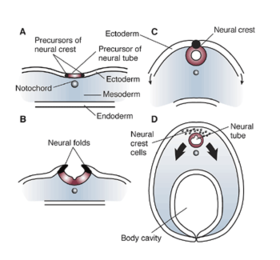

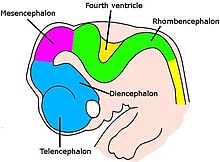
В начале третьей недели развития эмбриональная нервной эктодерма образует утолщенную полоску, называемую пластинкой . [64] К четвертой неделе развития нервная пластинка расширяется, образуя широкий головной конец, менее широкую среднюю часть и узкий хвостовой конец. Эти опухоли известны как первичные мозговые везикулы и представляют собой зачатки переднего мозга (передний мозг), среднего мозга (мезенцефалон) и заднего мозга (ромбенцефалон). [65] [66]
Клетки нервного гребня (происходящие из эктодермы) заселяют боковые края пластинки в области нервных складок . На четвертой неделе — на стадии нейруляции — нервные складки смыкаются, образуя нервную трубку , объединяя клетки нервного гребня в нервном гребне . [67] Нервный гребень проходит по всей длине трубки с краниальными клетками нервного гребня на головном конце и каудальными клетками нервного гребня на хвосте. Клетки отделяются от гребня и мигрируют краниокаудальной волной (от головы к хвосту) внутри трубки. [67] Клетки головного конца дают начало головному мозгу, а клетки каудального конца дают начало спинному мозгу. [68]
По мере роста трубка изгибается , образуя у головы полушария головного мозга серповидной формы. Полушария головного мозга впервые появляются на 32-й день. [69] В начале четвертой недели головная часть резко наклоняется вперед, образуя головной изгиб . [67] Эта согнутая часть становится передним мозгом (прозэнцефалоном); примыкающая изогнутая часть становится средним мозгом (средний мозг), а часть, каудальная к изгибу, становится задним мозгом (ромбэнцефалон). Эти области формируются в виде отеков, известных как три первичных мозговых пузырька . На пятой неделе развития пять вторичных мозговых пузырьков . сформировались [70] Передний мозг разделяется на два пузыря — передний промежуточный мозг и задний промежуточный мозг . Конечный мозг дает начало коре головного мозга, базальным ганглиям и родственным структурам. Промежуточный мозг дает начало таламусу и гипоталамусу. Задний мозг также разделяется на две области — метэнцефалон и продолговатый мозг . Метэнцефалон дает начало мозжечку и мосту. Продолговатый мозг дает начало продолговатому мозгу. [71] Также на пятой неделе мозг делится на повторяющиеся сегменты, называемые нейромерами . [65] [72] В заднем мозге они известны как ромбомеры . [73]
Характерной чертой мозга является корковая складчатость, известная как гирификация . В течение чуть более пяти месяцев пренатального развития кора головного мозга гладкая. К сроку беременности 24 недели становится очевидной морщинистая морфология, показывающая трещины, которые начинают выделять доли мозга. [74] Почему кора имеет морщины и складки, до конца не понятно, но гирификация связана с интеллектом и неврологическими расстройствами , и ряд теорий гирификации . был предложен [74] Эти теории включают теории, основанные на механическом выпучивании , [75] [18] аксональное напряжение , [76] и дифференциальное тангенциальное расширение . [75] Ясно то, что гирификация — это не случайный процесс, а скорее сложный процесс, предопределенный развитием, который генерирует узоры складок, одинаковые у отдельных особей и большинства видов. [75] [77]
Первой бороздой, которая появляется на четвертом месяце, является боковая мозговая ямка. [69] Расширяющийся каудальный конец полушария должен изогнуться вперед, чтобы вписаться в ограниченное пространство. Это покрывает ямку и превращает ее в гораздо более глубокий гребень, известный как латеральная борозда , и этим выделяется височная доля. [69] К шестому месяцу формируются другие борозды, разграничивающие лобную, теменную и затылочную доли. [69] Ген, присутствующий в геноме человека ( ARHGAP11B ), может играть важную роль в гирификации и энцефализации. [78]
 Мозг человеческого эмбриона на сроке 4,5 недели: видна внутренняя часть переднего мозга.
Мозг человеческого эмбриона на сроке 4,5 недели: видна внутренняя часть переднего мозга.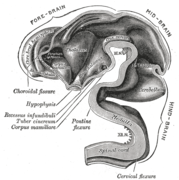 Внутренняя часть мозга на 5 неделе
Внутренняя часть мозга на 5 неделе Мозг осмотрен по средней линии через 3 месяца.
Мозг осмотрен по средней линии через 3 месяца.



Функция [ править ]
Управление двигателем [ править ]
Лобная доля участвует в рассуждении, управлении моторикой, эмоциях и речи. Он содержит моторную кору , которая участвует в планировании и координации движений; префронтальная кора , отвечающая за когнитивные функции более высокого уровня; и зона Брока , которая необходима для языкового производства. [79] мозга Двигательная система отвечает за создание и контроль движений. [80] Генерируемые движения передаются от мозга через нервы к мотонейронам тела, которые контролируют действие мышц . Корково -спинномозговой путь передает движения от головного мозга через спинной мозг к туловищу и конечностям. [81] Черепно -мозговые нервы осуществляют движения, связанные с глазами, ртом и лицом.
Общие движения, такие как передвижение и движения рук и ног, генерируются в моторной коре , разделенной на три части: первичная моторная кора , расположенная в прецентральной извилине и имеющая участки, предназначенные для движения различных частей тела. Эти движения поддерживаются и регулируются двумя другими областями, расположенными впереди первичной моторной коры: премоторной областью и дополнительной двигательной областью . [82] Руки и рот имеют гораздо большую площадь, чем другие части тела, что обеспечивает более тонкие движения; это было визуализировано в моторном гомункулусе . [82] Импульсы, генерируемые моторной корой, проходят по кортикоспинальному тракту вдоль передней части продолговатого мозга и пересекаются ( перекрещиваются ) у медуллярных пирамид . Затем они перемещаются вниз по спинному мозгу , при этом большая часть из них связывается с интернейронами , которые, в свою очередь, соединяются с нижними мотонейронами серого вещества , которые затем передают импульс к движению самим мышцам. [81] Мозжечок и базальные ганглии играют роль в тонких, сложных и скоординированных движениях мышц. [83] Связи между корой и базальными ганглиями контролируют мышечный тонус, позу и начало движений и называются экстрапирамидной системой . [84]
Сенсорика [ править ]


Сенсорная нервная система участвует в приеме и обработке сенсорной информации . Эта информация поступает через черепные нервы, через тракты спинного мозга и непосредственно в центры головного мозга, контактирующие с кровью. [85] Мозг также получает и интерпретирует информацию от особых органов чувств : зрения , обоняния , слуха и вкуса . смешанные двигательные и сенсорные сигналы . Также интегрированы [85]
От кожи мозг получает информацию о тонком прикосновении , давлении , боли , вибрации и температуре . От суставов мозг получает информацию о положении суставов . [86] Сенсорная кора находится рядом с моторной корой и, как и моторная кора, имеет области, связанные с ощущениями от разных частей тела. Ощущение, собираемое сенсорным рецептором на коже, преобразуется в нервный сигнал, который передается ряду нейронов через тракты спинного мозга. Путь дорсальный столб – медиальная петля содержит информацию о тонком прикосновении, вибрации и положении суставов. Волокна проводящих путей проходят вверх по задней части спинного мозга к задней части продолговатого мозга, где они соединяются с нейронами второго порядка , которые немедленно направляют волокна через срединную линию . Затем эти волокна направляются вверх в вентробазальный комплекс таламуса, где соединяются с нейронами третьего порядка , которые направляют волокна вверх к сенсорной коре. [86] Спиноталамический тракт несет информацию о боли, температуре и грубом прикосновении. Волокна этого пути проходят вверх по спинному мозгу и соединяются с нейронами второго порядка в ретикулярной формации ствола головного мозга, отвечающих за боль и температуру, а также заканчиваются в вентробазальном комплексе таламуса при грубом прикосновении. [87]
Зрение создается светом, попадающим на сетчатку глаза. Фоторецепторы сетчатки преобразуют сенсорный световой стимул в электрический нервный сигнал , который отправляется в зрительную кору затылочной доли. Зрительные сигналы покидают сетчатку через зрительные нервы .Волокна зрительного нерва от носовых половин сетчатки переходят на противоположные стороны, соединяясь с волокнами височных половин противоположных сетчаток, образуя зрительные тракты .Расположение оптики глаз и зрительных путей означает, что зрение из левого поля зрения воспринимается правой половиной каждой сетчатки, обрабатывается правой зрительной корой, и наоборот. Волокна зрительного тракта достигают мозга в латеральном коленчатом ядре и проходят через оптическое излучение, чтобы достичь зрительной коры. [88]
Слух и равновесие формируются во внутреннем ухе . Звук вызывает вибрации косточек , которые в конечном итоге доходят до органа слуха , а изменение баланса приводит к движению жидкостей во внутреннем ухе . Это создает нервный сигнал, который проходит через преддверно-улитковый нерв . Отсюда он проходит к ядрам улитки , верхнему оливковому ядру , медиальному коленчатому ядру и, наконец, слуховой лучистости к слуховой коре . [89]
Обоняние . генерируется клетками эпителия слизистой обонятельной оболочки полости носа рецепторными Эта информация проходит через обонятельный нерв , который проходит в череп через относительно проницаемую часть . Этот нерв передает информацию в нервную схему обонятельной луковицы, откуда информация передается в обонятельную кору . [90] [91] Вкус генерируется рецепторами языка и передается по лицевому и языкоглоточному нервам в одиночное ядро ствола мозга. Некоторая вкусовая информация также передается из глотки в эту область через блуждающий нерв . Отсюда информация передается через таламус во вкусовую кору . [92]
Регламент [ править ]
Вегетативные функции мозга включают регуляцию или ритмический контроль частоты сердечных сокращений и частоты дыхания , а также поддержание гомеостаза .
На кровяное давление и частоту сердечных сокращений влияет сосудодвигательный центр продолговатого мозга, что приводит к некоторому сужению артерий и вен в состоянии покоя. Это происходит путем воздействия на симпатическую и парасимпатическую нервную систему через блуждающий нерв . [93] Информация об артериальном давлении генерируется барорецепторами в тельцах дуги аорты и передается в мозг по афферентным волокнам блуждающего нерва. Информация об изменении давления в каротидном синусе поступает от сонных тел, расположенных вблизи сонной артерии , и передается через нерв, соединяющийся с языкоглоточным нервом . Эта информация доходит до одиночного ядра продолговатого мозга. Сигналы отсюда воздействуют на вазомоторный центр, соответствующим образом регулируя сужение вен и артерий. [94]
Мозг контролирует частоту дыхания , главным образом, с помощью дыхательных центров в продолговатом мозге и мосте. [95] Дыхательные центры контролируют дыхание , генерируя двигательные сигналы, которые передаются по спинному мозгу по диафрагмальному нерву к диафрагме и другим дыхательным мышцам . Это смешанный нерв , который передает сенсорную информацию обратно в центры. Выделяют четыре дыхательных центра, три с более четко выраженной функцией и апнейстический центр с менее четкой функцией. В продолговатом мозге дорсальная дыхательная группа вызывает желание вдохнуть и получает сенсорную информацию непосредственно от тела. Также в продолговатом мозге вентральная дыхательная группа влияет на выдох при нагрузке. В мосту пневмотаксический центр влияет на продолжительность каждого вдоха. [95] и апнейстический центр , по-видимому, влияет на вдох. Дыхательные центры непосредственно воспринимают углекислый газ и pH крови . Информация об уровне кислорода , углекислого газа и pH крови также воспринимается на стенках артерий в периферических хеморецепторах аортальных и сонных тел. Эта информация передается через блуждающий и языкоглоточный нервы в дыхательные центры. Высокий уровень углекислого газа, кислый pH или низкий уровень кислорода стимулируют дыхательные центры. [95] На желание вдохнуть также влияют рецепторы растяжения легких в легких, которые при активации предотвращают чрезмерное раздувание легких, передавая информацию в дыхательные центры через блуждающий нерв. [95]
Гипоталамус участвует в промежуточном мозге в регуляции многих функций организма. Функции включают нейроэндокринную регуляцию, регуляцию циркадного ритма , контроль вегетативной нервной системы , а также регуляцию потребления жидкости и пищи. Циркадный ритм контролируется двумя основными группами клеток гипоталамуса. Передний гипоталамус включает супрахиазматическое ядро и вентролатеральное преоптическое ядро , которое посредством циклов экспрессии генов генерирует примерно 24-часовой циркадный ритм . В циркадные сутки ультрадианным ритмом режим сна контролируется . Сон является важнейшим требованием для тела и мозга и позволяет закрыть и дать отдых системам организма. Есть также данные, которые позволяют предположить, что ежедневные накопления токсинов в мозге удаляются во время сна. [96] Во время бодрствования мозг потребляет пятую часть общей потребности организма в энергии. Сон обязательно уменьшает это использование и дает время для восстановления дающего энергию АТФ . Последствия лишения сна показывают абсолютную потребность во сне. [97]
Латеральный гипоталамус содержит орексинергические нейроны, которые контролируют аппетит и возбуждение через свои проекции на восходящую ретикулярную активирующую систему . [98] [99] Гипоталамус контролирует гипофиз посредством высвобождения пептидов, таких как окситоцин и вазопрессин , а также дофамина в срединное возвышение . Посредством вегетативных проекций гипоталамус участвует в регуляции таких функций, как артериальное давление, частота сердечных сокращений, дыхание, потоотделение и другие гомеостатические механизмы. [100] Гипоталамус также играет роль в терморегуляции и при стимуляции иммунной системой способен вызывать лихорадку . На гипоталамус влияют почки: когда кровяное давление падает, ренин, выделяемый почками, стимулирует потребность пить. Гипоталамус также регулирует потребление пищи посредством вегетативных сигналов и высвобождение гормонов пищеварительной системой. [101]
Язык [ править ]
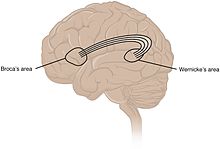
Традиционно считалось, что языковые функции локализованы в зоне Вернике и зоне Брока . [102] в настоящее время общепризнано, что более широкая сеть корковых областей способствует речевым функциям. [103] [104] [105]
Исследование того, как язык представляется, обрабатывается и усваивается мозгом, называется нейролингвистикой и представляет собой крупную междисциплинарную область, основанную на когнитивной нейробиологии , когнитивной лингвистике и психолингвистике . [106]
Латерализация [ править ]
Головной мозг имеет контралатеральную организацию : каждое полушарие мозга взаимодействует преимущественно с одной половиной тела: левое полушарие мозга взаимодействует с правой стороной тела, и наоборот. Предполагается, что это вызвано осевым поворотом в процессе развития . [107] Двигательные связи от головного мозга к спинному мозгу и сенсорные связи от спинного мозга к головному мозгу пересекают ствол мозга. Визуальный ввод подчиняется более сложному правилу: зрительные нервы двух глаз соединяются в точке, называемой перекрестом зрительных нервов , и половина волокон каждого нерва разделяется, чтобы присоединиться к другому. [108] В результате соединения из левой половины сетчатки обоих глаз идут в левое полушарие мозга, тогда как соединения из правой половины сетчатки идут в правое полушарие мозга. [109] Поскольку каждая половина сетчатки получает свет, исходящий из противоположной половины поля зрения, функциональное следствие состоит в том, что зрительная информация из левого полушария мира поступает в правую часть мозга, и наоборот. [110] Таким образом, правое полушарие мозга получает соматосенсорную информацию от левой стороны тела и зрительную информацию от левой части поля зрения. [111] [112]
Левое и правое полушария мозга кажутся симметричными, но функционируют асимметрично. [113] Например, аналогом моторной области левого полушария, контролирующей правую руку, является область правого полушария, контролирующая левую руку. Однако есть несколько важных исключений, касающихся языка и пространственного познания. Левая лобная доля является доминирующей в речевом развитии. Если ключевая речевая область в левом полушарии повреждена, жертва может потерять способность говорить и понимать. [113] тогда как эквивалентное повреждение правого полушария приведет лишь к незначительному ухудшению языковых навыков.
Существенная часть нынешнего понимания взаимодействия между двумя полушариями возникла в результате изучения « пациентов с расщепленным мозгом » — людей, перенесших хирургическое пересечение мозолистого тела в попытке уменьшить тяжесть эпилептических припадков. [114] Эти пациенты не демонстрируют необычного поведения, которое сразу бросается в глаза, но в некоторых случаях могут вести себя почти как два разных человека в одном теле: правая рука выполняет действие, а левая отменяет его. [114] [115] Эти пациенты, если кратко показать изображение справа от точки зрительной фиксации, способны описать его словесно, но когда изображение показано слева, не могут его описать, но могут дать указание. левой рукой характер изображенного предмета. [115] [116]
Эмоции [ править ]
Эмоции обычно определяются как двухэтапные многокомпонентные процессы, включающие извлечение , за которым следуют психологические чувства, оценка, выражение, вегетативные реакции и тенденции к действию. [117] Попытки локализовать основные эмоции в определенных областях мозга вызвали споры; Некоторые исследования не обнаружили доказательств наличия определенных мест, соответствующих эмоциям, но вместо этого обнаружили схемы, участвующие в общих эмоциональных процессах. Миндалевидное тело , орбитофронтальная кора , средняя и передняя островковая кора и латеральная префронтальная кора , по-видимому, участвуют в генерации эмоций, в то время как более слабые доказательства были обнаружены для вентральной покрышки , вентрального бледного тела и прилежащего ядра в стимулирующей значимости . [118] Другие, однако, обнаружили доказательства активации определенных областей, таких как базальные ганглии при счастье, подмозолистая поясная кора при печали и миндалевидное тело при страхе. [119]
Познание [ править ]
Мозг отвечает за познание , [120] [121] который функционирует посредством многочисленных процессов и исполнительных функций . [121] [122] [123] Исполнительные функции включают в себя способность фильтровать информацию и отключать ненужные стимулы с помощью контроля внимания и когнитивного торможения , способность обрабатывать и манипулировать информацией, хранящейся в рабочей памяти , способность думать о нескольких концепциях одновременно и переключать задачи с когнитивной гибкостью , способность подавлять импульсы и доминантные реакции с помощью тормозящего контроля , а также способность определять актуальность информации или целесообразность действия. [122] [123] Исполнительные функции более высокого порядка требуют одновременного использования нескольких основных исполнительных функций и включают планирование , поиск и подвижный интеллект (т. е. рассуждение и решение проблем ). [123]
Префронтальная кора играет важную роль в реализации управляющих функций. [121] [123] [124] Планирование включает активацию дорсолатеральной префронтальной коры (ДЛПФК), передней поясной извилины , угловой префронтальной коры, правой префронтальной коры и супрамаргинальной извилины . [124] Манипулирование рабочей памятью затрагивает DLPFC, нижнюю лобную извилину и области теменной коры . [121] [124] Тормозной контроль задействует несколько областей префронтальной коры, а также хвостатое ядро и субталамическое ядро . [123] [124] [125]
Физиология [ править ]
Нейротрансмиссия [ править ]
Мозговая деятельность становится возможной благодаря взаимосвязям нейронов , которые связаны друг с другом для достижения своих целей. [126] Нейрон состоит из тела клетки , аксона и дендритов . Дендриты часто представляют собой обширные ветви, которые получают информацию в виде сигналов от окончаний аксонов других нейронов. Полученные сигналы могут заставить нейрон инициировать потенциал действия (электрохимический сигнал или нервный импульс), который передается по его аксону к окончанию аксона, чтобы соединиться с дендритами или с телом клетки другого нейрона. Потенциал действия инициируется в начальном сегменте аксона, который содержит специализированный комплекс белков. [127] Когда потенциал действия достигает окончания аксона, он вызывает высвобождение нейромедиатора в синапсе , который распространяет сигнал, который действует на клетку-мишень. [128] Эти химические нейротрансмиттеры включают дофамин , серотонин , ГАМК , глутамат и ацетилхолин . [129] ГАМК является основным тормозным нейромедиатором в головном мозге, а глутамат — основным возбуждающим нейромедиатором. [130] Нейроны соединяются в синапсах, образуя нейронные пути , нейронные цепи и большие сложные сетевые системы, такие как сеть значимости и сеть режима по умолчанию , а активность между ними управляется процессом нейротрансмиссии .
Метаболизм [ править ]
Мозг потребляет до 20% энергии, используемой организмом человека, больше, чем любой другой орган. [131] У людей глюкоза в крови является основным источником энергии для большинства клеток и имеет решающее значение для нормального функционирования ряда тканей, включая мозг. [132] Человеческий мозг потребляет около 60% глюкозы в крови у голодающих и малоподвижных людей. [132] мозга Метаболизм обычно зависит от глюкозы в крови в качестве источника энергии, но во время низкого уровня глюкозы (например, при голодании , упражнениях на выносливость или ограниченном потреблении углеводов ) мозг использует кетоновые тела в качестве топлива с меньшей потребностью в глюкозе. Мозг также может использовать лактат во время физических упражнений . [133] Мозг хранит глюкозу в форме гликогена , хотя и в значительно меньших количествах, чем в печени или скелетных мышцах . [134] Длинноцепочечные жирные кислоты не могут проникнуть через гематоэнцефалический барьер , но печень может расщеплять их с образованием кетоновых тел. Однако жирные кислоты с короткой цепью (например, масляная кислота , пропионовая кислота и уксусная кислота ) и жирные кислоты со средней длиной цепи , октановая кислота и гептановая кислота , могут преодолевать гематоэнцефалический барьер и метаболизироваться клетками головного мозга . [135] [136] [137]
Хотя человеческий мозг составляет всего 2% массы тела, он получает 15% сердечного выброса, 20% общего потребления кислорода организмом и 25% общего использования глюкозы организмом . [138] Мозг в основном использует глюкозу для получения энергии, а лишение глюкозы, как это может произойти при гипогликемии , может привести к потере сознания. [139] Энергопотребление мозга не сильно меняется со временем, но активные области коры потребляют несколько больше энергии, чем неактивные, что составляет основу функциональных нейровизуализации методов ПЭТ и фМРТ . [140] Эти методы обеспечивают трехмерное изображение метаболической активности. [141] Предварительное исследование показало, что метаболические потребности мозга у людей достигают пика примерно в пятилетнем возрасте. [142]
Функция сна до конца не изучена; однако есть свидетельства того, что сон усиливает выведение продуктов метаболизма, некоторые из которых потенциально нейротоксичны , из мозга, а также может способствовать восстановлению. [52] [143] [144] Имеющиеся данные свидетельствуют о том, что увеличение выведения метаболических отходов во время сна происходит за счет улучшения функционирования глимфатической системы . [52] Сон также может влиять на когнитивные функции, ослабляя ненужные связи. [145]
Исследования [ править ]
Мозг до конца не изучен, и исследования продолжаются. [146] Нейробиологи вместе с исследователями смежных дисциплин изучают, как работает человеческий мозг. Границы между неврологией , неврологией и другими дисциплинами, такими как психиатрия, исчезли, поскольку все они находятся под влиянием фундаментальных исследований в области неврологии.
Нейронаучные исследования значительно расширились. Считается, что « Десятилетие мозга », инициатива правительства Соединенных Штатов в 1990-х годах, во многом ознаменовало этот рост исследований. [147] а в 2013 году последовала инициатива BRAIN . [148] Проект Human Connectome — это пятилетнее исследование, начатое в 2009 году с целью анализа анатомических и функциональных связей частей мозга, которое предоставило много данных. [146]
Новым этапом исследований может стать моделирование активности мозга. [149]
Методы [ править ]
Информация о структуре и функциях человеческого мозга поступает с помощью различных экспериментальных методов, в том числе на животных и человеке. Информация о травмах головного мозга и инсульте предоставила информацию о функции частей мозга и последствиях повреждения головного мозга . Нейровизуализация используется для визуализации мозга и записи мозговой активности. Электрофизиология используется для измерения, записи и мониторинга электрической активности коры головного мозга. Измерения могут касаться потенциалов локальных полей корковых областей или активности отдельного нейрона. Электроэнцефалограмма неинвазивно может регистрировать электрическую активность коры головного мозга с помощью электродов, помещаемых на кожу головы . [150] [151]
Инвазивные меры включают электрокортикографию , при которой используются электроды, помещаемые непосредственно на открытую поверхность мозга. Этот метод используется при картировании корковой стимуляции , используемом при изучении взаимосвязи между областями коры и их системной функцией. [152] Используя микроэлектроды гораздо меньшего размера , можно производить единичные записи одного нейрона, которые обеспечивают высокое пространственное разрешение и высокое временное разрешение . Это позволило связать активность мозга с поведением и создать карты нейронов. [153]
Развитие церебральных органоидов открыло пути изучения роста мозга и коры головного мозга, а также понимания развития заболеваний, что открывает новые возможности для терапевтического применения. [154] [155]
Изображение [ править ]

Методы функциональной нейровизуализации показывают изменения в активности мозга, связанные с функцией определенных областей мозга. Одним из методов является функциональная магнитно-резонансная томография (фМРТ), которая имеет преимущества перед более ранними методами ОФЭКТ и ПЭТ, поскольку не требует использования радиоактивных материалов и обеспечивает более высокое разрешение. [156] Другой метод — функциональная спектроскопия ближнего инфракрасного диапазона . Эти методы основаны на гемодинамической реакции , которая показывает изменения в активности мозга в зависимости от изменений в кровотоке , что полезно при сопоставлении функций с областями мозга . [157] ФМРТ в состоянии покоя рассматривает взаимодействие областей мозга, пока мозг не выполняет конкретную задачу. [158] Это также используется для отображения сети в режиме по умолчанию .
Любой электрический ток создает магнитное поле; нейронные колебания индуцируют слабые магнитные поля, а при функциональной магнитоэнцефалографии производимый ток может показать локализованную функцию мозга в высоком разрешении. [159] Трактография использует МРТ и анализ изображений для создания трехмерных изображений нервных путей головного мозга. Коннектограммы дают графическое представление нейронных связей головного мозга. [160]
Различия в структуре мозга можно измерить при некоторых расстройствах, особенно при шизофрении и деменции . Различные биологические подходы с использованием визуализации позволили лучше понять, например, расстройства депрессии и обсессивно-компульсивного расстройства . Ключевым источником информации о функции областей мозга являются последствия их повреждения. [161]
Достижения в области нейровизуализации позволили получить объективное представление о психических расстройствах, что привело к более быстрой диагностике, более точному прогнозу и лучшему мониторингу. [162]
генов Экспрессия белков и
Биоинформатика — это область исследований, которая включает в себя создание и развитие баз данных, а также вычислительных и статистических методов, которые можно использовать в исследованиях человеческого мозга, особенно в области экспрессии генов и белков . Биоинформатика и исследования в области геномики и функциональной геномики породили потребность в аннотации ДНК , технологии транскриптома , позволяющей идентифицировать гены , их расположение и функции. [163] [164] [165] GeneCards — это крупная база данных.
По состоянию на 2017 год [update] чуть менее 20 000 генов, кодирующих белки . у человека экспрессируется [163] и около 400 из этих генов специфичны для мозга. [166] [167] Полученные данные об экспрессии генов в мозге стимулировали дальнейшие исследования ряда заболеваний. Например, длительное употребление алкоголя показало изменение экспрессии генов в мозге и специфические изменения типов клеток, которые могут быть связаны с расстройством, вызванным употреблением алкоголя . [168] Эти изменения были отмечены в синаптическом транскриптоме префронтальной коры и рассматриваются как фактор, вызывающий склонность к алкогольной зависимости, а также к злоупотреблению другими психоактивными веществами . [169]
Другие аналогичные исследования также показали доказательства синаптических изменений и их утраты в стареющем мозге . Изменения в экспрессии генов изменяют уровни белков в различных нервных путях, и было показано, что это проявляется в дисфункции или потере синаптических контактов. Было замечено, что эта дисфункция затрагивает многие структуры головного мозга и оказывает заметное влияние на тормозные нейроны, что приводит к снижению уровня нейротрансмиссии и последующему когнитивному упадку и заболеванию. [170] [171]
Клиническое значение
Травма [ править ]
Травма головного мозга может проявляться по-разному. Черепно-мозговая травма , например, полученная при занятиях контактными видами спорта , после падения , дорожно-транспортного происшествия или несчастного случая на производстве , может быть связана как с непосредственными, так и с долгосрочными проблемами. Непосредственные проблемы могут включать кровоизлияние в мозг , которое может сдавить ткань мозга или повредить его кровоснабжение. Может возникнуть ушиб головного мозга. Синяки могут вызвать обширное повреждение нервных путей, что может привести к диффузному повреждению аксонов . [172] , Перелом черепа травма определенной области, глухота и сотрясение мозга также являются возможными непосредственными последствиями. Помимо места травмы, может быть поражена и противоположная сторона мозга, что называется контр- травмой. Более долгосрочные проблемы, которые могут возникнуть, включают посттравматическое стрессовое расстройство и гидроцефалию . Хроническая травматическая энцефалопатия может развиться после множественных травм головы . [173]
Болезнь [ править ]
Нейродегенеративные заболевания приводят к прогрессирующему повреждению различных частей мозга и ухудшаются с возрастом . Общие примеры включают деменцию, такую как болезнь Альцгеймера , алкогольную деменцию или сосудистую деменцию ; болезнь Паркинсона ; и другие более редкие инфекционные, генетические или метаболические причины, такие как болезнь Хантингтона , заболевания двигательных нейронов , ВИЧ-деменция , деменция, связанная с сифилисом, и болезнь Вильсона . Нейродегенеративные заболевания могут поражать различные части мозга и влиять на движение, память и познание. [174]
Церебральный атеросклероз – это атеросклероз , поражающий мозг. результате накопления бляшек холестериновых Это заболевание возникает в в крупных артериях головного мозга и может быть от легкой до значительной степени. В серьезных случаях артерии могут сузиться настолько, что уменьшится кровоток. Он способствует развитию деменции и имеет сходство белков с белками, обнаруженными при болезни Альцгеймера. [175]
Мозг, хотя и защищен гематоэнцефалическим барьером, может поражаться инфекциями, включая вирусы , бактерии и грибки . Инфекция может поразить мозговые оболочки ( менингит ), вещество головного мозга ( энцефалит ) или внутри вещества головного мозга (например, абсцесс мозга ). [176] Редкие прионные заболевания , включая болезнь Крейтцфельдта-Якоба и ее вариант , а также куру, также могут поражать мозг. [176]
Опухоли [ править ]
Опухоли головного мозга могут быть доброкачественными или раковыми . Большинство злокачественных опухолей возникают из других частей тела , чаще всего из легких , молочной железы и кожи . [177] Рак ткани головного мозга также может возникать и возникать из любой ткани внутри и вокруг мозга. Менингиома , рак мозговых оболочек головного мозга, встречается чаще, чем рак ткани головного мозга. [177] Рак головного мозга может вызывать симптомы, связанные с их размером или положением, включая головную боль и тошноту, или постепенное развитие очаговых симптомов, таких как постепенное затруднение зрения, глотания, разговора или изменение настроения. [177] Рак обычно исследуют с помощью компьютерной томографии и МРТ. Для выяснения причины рака и оценки типа и стадии рака можно использовать множество других тестов, включая анализы крови и люмбальную пункцию. [177] Кортикостероид дексаметазон ткани головного мозга часто назначают для уменьшения отека вокруг опухоли. Можно рассмотреть возможность хирургического вмешательства, однако, учитывая сложную природу многих опухолей или в зависимости от стадии или типа опухоли, лучевая терапия или химиотерапия могут считаться более подходящими. [177]
Психические расстройства [ править ]
Известно, что психические расстройства , такие как депрессия , шизофрения , биполярное расстройство , посттравматическое стрессовое расстройство , синдром дефицита внимания с гиперактивностью , обсессивно-компульсивное расстройство , синдром Туретта и зависимость , связаны с функционированием мозга. [125] [129] [178] Лечение психических расстройств может включать психотерапию , психиатрию , социальное вмешательство личности по восстановлению и работу или когнитивно-поведенческую терапию ; основные проблемы и связанные с ними прогнозы значительно различаются у разных людей. [179]
Эпилепсия [ править ]
эпилептические припадки связаны с аномальной электрической активностью. Считается, что [180] Судорожная активность может проявляться в виде отсутствия сознания , очаговых явлений, таких как движения конечностей или затруднения речи, или носить генерализованный характер. [180] Эпилептический статус – это приступ или серия приступов, которые не прекратились в течение 5 минут. [181] Приступы имеют большое количество причин, однако многие припадки происходят без установления точной причины. У человека с эпилепсией факторами риска дальнейших приступов могут быть бессонница, употребление наркотиков и алкоголя, а также стресс. Судороги можно оценить с помощью анализов крови , ЭЭГ и различных методов медицинской визуализации на основе истории болезни и результатов медицинского обследования . [180] Помимо лечения основной причины и снижения воздействия факторов риска, противосудорожные препараты могут сыграть роль в предотвращении дальнейших судорог. [180]
Врожденный [ править ]
Некоторые заболевания головного мозга, такие как болезнь Тея-Сакса , [182] являются врожденными и связаны с генетическими и хромосомными мутациями. [183] Редкая группа врожденных заболеваний головного мозга, известная как лиссэнцефалия, характеризуется отсутствием или неадекватностью кортикальных складок. [184] Нормальное развитие мозга во время беременности может быть нарушено из-за дефицита питательных веществ . [185] тератогены , [186] инфекционные заболевания , [187] и употреблением легких наркотиков , включая алкоголь (что может привести к расстройствам алкогольного спектра плода ). [185] [188] Большинство церебральных артериовенозных мальформаций являются врожденными, эти запутанные сети кровеносных сосудов могут оставаться без симптомов, но в худшем случае могут разорваться и вызвать внутричерепное кровоизлияние . [189]
Инсульт [ править ]

Инсульт гибель – это уменьшение кровоснабжения определенной области мозга, вызывающее клеток и повреждение головного мозга . Это может привести к широкому спектру симптомов , включая « БЫСТРЫЕ » симптомы, такие как опущение лица, слабость рук и трудности с речью (в том числе с речью , подбором слов или формированием предложений ). [190] Симптомы связаны с функцией пораженного участка мозга и могут указывать на вероятное место и причину инсульта. Трудности с движением, речью или зрением обычно связаны с головным мозгом, тогда как дисбаланс , двоение в глазах , головокружение и симптомы, поражающие более чем одну сторону тела, обычно связаны со стволом мозга или мозжечком. [191]
Большинство инсультов возникают в результате потери кровоснабжения, обычно из-за эмболии , разрыва жировой бляшки, вызывающего тромб , или сужения мелких артерий . Инсульт может также возникнуть в результате кровоизлияния в мозг . [192] Транзиторные ишемические атаки (ТИА) — это инсульты, при которых симптомы проходят в течение 24 часов. [192] Расследование инсульта будет включать медицинское обследование (включая неврологическое обследование ) и сбор анамнеза с упором на продолжительность симптомов и факторы риска (включая высокое кровяное давление , фибрилляцию предсердий и курение ). [193] Дальнейшие исследования необходимы у более молодых пациентов. [194] ЭКГ ; и биотелеметрия быть проведены Для выявления мерцательной аритмии могут УЗИ ; позволяет сужение сонных артерий выявить Эхокардиограмму можно использовать для поиска тромбов в сердце, заболеваний сердечных клапанов или наличия открытого овального окна . [194] обычно проводятся анализы крови В рамках обследования , включая тесты на диабет и липидный профиль . [194]
Некоторые методы лечения инсульта критичны по времени. К ним относятся растворение тромба или хирургическое удаление тромба при ишемических инсультах и декомпрессия при геморрагических инсультах . [195] [196] Поскольку инсульт имеет решающее значение во времени, [197] больницы и даже догоспитальная помощь при инсульте включает в себя ускоренные исследования – обычно компьютерную томографию для выявления геморрагического инсульта и КТ- или МР-ангиограмму для оценки артерий, кровоснабжающих головной мозг. [194] МРТ-сканирование , которое не так широко доступно, может более точно продемонстрировать пораженный участок мозга, особенно при ишемическом инсульте. [194]
Перенесший инсульт человек может быть госпитализирован в инсультное отделение , и лечение может быть направлено на предотвращение будущих инсультов, включая постоянную антикоагулянтную терапию (например, аспирин или клопидогрел ), антигипертензивные средства и гиполипидемические препараты . [195] включающая Многопрофильная команда, логопедов , физиотерапевтов , эрготерапевтов и психологов , играет большую роль в поддержке человека, перенесшего инсульт, и его реабилитации . [198] [194] Инсульт в анамнезе увеличивает риск развития деменции примерно на 70%, а недавний инсульт увеличивает риск примерно на 120%. [199]
Смерть мозга [ править ]
Смерть мозга означает необратимую полную потерю функций мозга. [200] [201] Это характеризуется комой , потерей рефлексов и апноэ . [200] однако заявление о смерти мозга варьируется в зависимости от географического положения и не всегда принимается. [201] В некоторых странах также существует определенный синдром смерти ствола мозга . [202] Заявление о смерти мозга может иметь серьезные последствия, поскольку это заявление, согласно принципу медицинской бесполезности , будет связано с прекращением жизнеобеспечения. [203] и поскольку у людей со смертью мозга часто есть органы, пригодные для донорства органов . [201] [204] Этот процесс часто осложняется плохим общением с семьями пациентов. [205]
При подозрении на смерть мозга необходимо исключить обратимые дифференциальные диагнозы , такие как электролитное, неврологическое и лекарственное когнитивное подавление. [200] [203] Проверка рефлексов [б] может помочь в принятии решения, как и отсутствие реакции и дыхания. [203] Клинические наблюдения, включая полное отсутствие реакции, известный диагноз и данные нейровизуализации , могут сыграть роль в решении констатировать смерть мозга. [200]
и Общество культура
Нейроантропология – это исследование взаимосвязи между культурой и мозгом. В нем исследуется, как мозг порождает культуру и как культура влияет на развитие мозга. [206] Культурные различия и их связь с развитием и структурой мозга исследуются в разных областях. [207]
Разум [ править ]

Философия разума изучает такие вопросы, как проблема понимания сознания и проблема разума и тела . Отношения между мозгом и разумом представляют собой серьезную проблему как с философской, так и с научной точки зрения. Это связано с трудностью объяснения того, как умственная деятельность, такая как мысли и эмоции, может осуществляться физическими структурами, такими как нейроны и синапсы , или любым другим типом физического механизма. Эту трудность выразил Готфрид Лейбниц в аналогии, известной как « Мельница Лейбница» :
Приходится признать, что восприятие и то, что от него зависит, необъяснимы механическими принципами, т. е. фигурами и движениями. Вообразив, что существует машина, конструкция которой позволяет ей думать, чувствовать и воспринимать, можно было бы представить ее увеличенной, сохраняя при этом те же пропорции, чтобы можно было войти в нее, как в ветряную мельницу. Если предположить это, то, посещая его, мы должны обнаружить только части, подталкивающие друг друга, и никогда не найти ничего, чем можно было бы объяснить восприятие.
- - Лейбниц, Монадология [209]
Сомнение в возможности механистического объяснения мышления привело Рене Декарта , а вместе с ним и большинство других философов, к дуализму : вере в то, что разум в некоторой степени независим от мозга. [210] Однако всегда существовали сильные аргументы в противоположном направлении. Существуют четкие эмпирические доказательства того, что физические манипуляции с мозгом или его травмы (например, наркотиками или повреждениями соответственно) могут влиять на разум сильным и интимным образом. [211] [212] В XIX веке случай с Финеасом Гейджем , железнодорожным рабочим, который был ранен толстым железным стержнем, проходящим через его мозг, убедил как исследователей, так и общественность в том, что когнитивные функции локализованы в мозге. [208] Следуя этой линии мышления, большой объем эмпирических данных о тесной связи между активностью мозга и умственной деятельностью привел к тому, что большинство нейробиологов и современных философов стали материалистами , полагая, что психические явления в конечном итоге являются результатом физических явлений или могут быть сведены к ним. [213]
Размер мозга [ править ]
человека Размер мозга и интеллект не сильно связаны. [214] Исследования, как правило, указывают на корреляцию от небольшой до умеренной (в среднем от 0,3 до 0,4) между объемом мозга и IQ . [215] Наиболее устойчивые ассоциации наблюдаются в лобной, височной и теменной долях, гиппокампе и мозжечке, но они объясняют лишь относительно небольшую дисперсию IQ, который сам по себе имеет лишь частичное отношение к общему интеллекту и реальному состоянию. мировое исполнение. [216] [217]
У других животных, в том числе китов и слонов, мозг больше, чем у человека. Однако, если принять во внимание соотношение массы мозга и тела , человеческий мозг почти в два раза больше, чем у афалины , и в три раза больше, чем у шимпанзе . Однако высокое соотношение само по себе не свидетельствует об интеллекте: у очень мелких животных оно высокое, а у землеройки самый высокий коэффициент среди всех млекопитающих. [218]
В популярной культуре [ править ]

Более ранние представления об относительной важности различных органов человеческого тела иногда подчеркивали сердце. [219] Современные западные популярные концепции, напротив, уделяют все больше внимания мозгу . [220]
Исследования опровергли некоторые распространенные заблуждения о мозге . К ним относятся как древние, так и современные мифы. Неправда (например), что нейроны не заменяются после двухлетнего возраста; ни то, что нормальные люди используют только десять процентов мозга . [221] Популярная культура также слишком упростила латерализацию мозга , предполагая, что функции полностью специфичны для той или иной стороны мозга. Акио Мори придумал термин « игровой мозг » для обозначения ненадежно подтвержденной теории о том, что длительное времяпровождение за видеоиграми наносит вред префронтальной области мозга и ухудшает выражение эмоций и творческие способности. [222]
Исторически, особенно в начале 19-го века, мозг фигурировал в массовой культуре через френологию — псевдонауку , которая приписывала атрибуты личности различным областям коры головного мозга. Кора головного мозга остается важной в популярной культуре, о чем говорится в книгах и сатире. [223] [224]
Человеческий мозг может фигурировать в научной фантастике с такими темами, как трансплантация мозга и киборги (существа с характеристиками, похожими на частично искусственный мозг ). [225] Научно-фантастическая книга 1942 года (трижды адаптированная для кино) « Мозг Донована» рассказывает историю об изолированном мозге , сохраняемом живым in vitro , постепенно принимая на себя личность главного героя книги. [226]
История [ править ]
Ранняя история [ править ]

Папирус Эдвина Смита , древнеегипетский медицинский трактат, написанный в 17 веке до нашей эры, содержит самое раннее зарегистрированное упоминание о мозге. Иероглиф « мозг», встречающийся в этом папирусе восемь раз, описывает симптомы, диагностику и прогноз двух травм головы. В папирусе упоминаются внешняя поверхность мозга, последствия травм (включая судороги и афазию ), мозговые оболочки и спинномозговая жидкость. [227] [228]
В пятом веке до нашей эры Алкмеон из Кротона в Великой Греции впервые считал мозг вместилищем разума . [228] Также в пятом веке до нашей эры в Афинах неизвестный автор « О священной болезни », медицинского трактата, который является частью « Корпуса Гиппократа» и традиционно приписывался Гиппократу , полагал, что мозг является местом обитания интеллекта. Аристотель в своей биологии первоначально считал сердце вместилищем разума и рассматривал мозг как механизм охлаждения крови. Он рассуждал, что люди более рациональны, чем звери, потому что, среди прочего, у них больший мозг, способный охладить их вспыльчивость. [229] Аристотель описал мозговые оболочки и различал головной мозг и мозжечок. [230]
Герофил Халкидонский в четвертом и третьем веках до нашей эры различал головной мозг и мозжечок и дал первое четкое описание желудочков ; и вместе с экспериментировали с Эрасистратом Кеосским живым мозгом. Их труды сейчас большей частью утеряны, а об их достижениях мы знаем преимущественно из вторичных источников. Некоторые из их открытий пришлось заново открыть через тысячелетие после их смерти. [228] Врач-анатом Гален во втором веке нашей эры, во времена Римской империи , препарировал мозг овец, обезьян, собак и свиней. Он пришел к выводу, что, поскольку мозжечок плотнее мозга, он должен управлять мышцами , а поскольку головной мозг мягок, то именно в нем и обрабатываются чувства. Гален далее предположил, что мозг функционирует за счет движения духов животных через желудочки. [228] [229]
Ренессанс [ править ]


В 1316 году Мондино де Луцци положила «Анатомия» начало современному изучению анатомии мозга. [231] Никколо Масса в 1536 году обнаружил, что желудочки заполнены жидкостью. [232] Архангело Пикколомини из Рима был первым, кто различал головной мозг и кору головного мозга. [233] В 1543 году Андреас Везалий опубликовал семитомный труд « О человеческом телесном фабрике» . [233] [234] [235] Седьмая книга охватывала мозг и глаза, содержала подробные изображения желудочков, черепных нервов, гипофиза , мозговых оболочек, структур глаза , кровоснабжения головного и спинного мозга, а также изображения периферических нервов. [236] Везалий отверг распространенное мнение о том, что желудочки отвечают за работу мозга, утверждая, что многие животные имеют желудочковую систему, похожую на человеческую, но не имеют настоящего интеллекта. [233]
Рене Декарт предложил теорию дуализма , чтобы решить проблему отношения мозга к разуму. Он предположил, что шишковидная железа — это место, где разум взаимодействует с телом, служащее вместилищем души и связующим звеном, через которое духи животных переходят из крови в мозг. [232] Этот дуализм, вероятно, послужил стимулом для более поздних анатомов к дальнейшему исследованию взаимосвязи между анатомическими и функциональными аспектами анатомии мозга. [237]
Томас Уиллис считается вторым пионером в изучении неврологии и науки о мозге. Он написал Cerebri Anatome ( лат . Анатомия мозга ). [с] в 1664 г., за ним последовали «Церебральная патология» в 1667 г. В них он описал строение мозжечка, желудочков, полушарий головного мозга, ствола мозга и черепных нервов, изучил его кровоснабжение; и предполагаемые функции, связанные с различными областями мозга. [233] Круг Уиллиса был назван в честь его исследований кровоснабжения головного мозга, и он первым употребил слово «неврология». [238] Уиллис удалил мозг из тела при его исследовании и отверг широко распространенное мнение о том, что кора состоит только из кровеносных сосудов, а также мнение последних двух тысячелетий о том, что кора играет лишь второстепенную роль. [233]
В середине XIX века Эмиль дю Буа-Реймон и Герман фон Гельмгольц смогли с помощью гальванометра показать, что электрические импульсы передаются с измеримой скоростью по нервам, опровергнув точку зрения своего учителя Иоганна Петера Мюллера о том, что нервный импульс является жизненно важной функцией. это невозможно было измерить. [239] [240] [241] Ричард Кейтон в 1875 году продемонстрировал электрические импульсы в полушариях головного мозга кроликов и обезьян. [242] В 1820-х годах Жан-Пьер Флуранс впервые применил экспериментальный метод повреждения определенных частей мозга животных, описывающий влияние на движение и поведение. [243]
Современный период [ править ]


Исследования мозга стали более сложными с использованием микроскопа и разработкой окрашивания серебром метода Камилло Гольджи в 1880-х годах. Это позволило показать сложные структуры отдельных нейронов. [244] Это было использовано Сантьяго Рамоном-и-Кахалем и привело к формированию доктрины нейронов , революционной на тот момент гипотезы о том, что нейрон является функциональной единицей мозга. Он использовал микроскопию, чтобы обнаружить многие типы клеток, и предложил функции клеток, которые он видел. [244] За это Гольджи и Кахаля считают основателями нейробиологии двадцатого века , оба получили Нобелевскую премию 1906 года за свои исследования и открытия в этой области. [244]
Чарльз Шеррингтон опубликовал в 1906 году свою влиятельную работу «Интегративное действие нервной системы», в которой исследуются функции рефлексов, эволюционное развитие нервной системы, функциональная специализация мозга, а также расположение и клеточные функции центральной нервной системы. [245] В 1942 году он ввёл термин « зачарованный ткацкий станок» как метафору мозга. Джон Фаркуар Фултон основал Журнал нейрофизиологии и опубликовал первый всеобъемлющий учебник по физиологии нервной системы в 1938 году. [246] Нейронаука в двадцатом веке стала признаваться как отдельная единая академическая дисциплина, а Дэвид Риоч , Фрэнсис О. Шмитт и Стивен Каффлер сыграли решающую роль в становлении этой области. [247] Риоч инициировал интеграцию фундаментальных анатомических и физиологических исследований с клинической психиатрией в Армейском исследовательском институте Уолтера Рида , начиная с 1950-х годов. [248] В тот же период Шмитт учредил Программу нейробиологических исследований — межуниверситетскую и международную организацию, объединяющую биологию, медицину, психологию и поведенческие науки. Само слово «нейронаука» возникло из этой программы. [249]
Поль Брока связал области мозга с определенными функциями, в частности речью в зоне Брока , после работы с пациентами с повреждением головного мозга. [250] Джон Хьюлингс Джексон описал функцию моторной коры , наблюдая за развитием эпилептических припадков по всему телу. Карл Вернике описал область, связанную с пониманием и производством языка. Корбиниан Бродманн разделил области мозга в зависимости от внешнего вида клеток. [250] К 1950 году Шеррингтон, Папес и Маклин определили многие функции ствола мозга и лимбической системы. [251] [252] Способность мозга к реорганизации и изменению с возрастом, а также признанный критический период развития были приписаны нейропластичности , впервые разработанной Маргарет Кеннард , которая экспериментировала на обезьянах в 1930-40-х годах. [253]
Харви Кушинг (1869–1939) признан первым опытным нейрохирургом . в мире [254] В 1937 году Уолтер Денди начал практику сосудистой нейрохирургии , выполнив первое хирургическое клипирование внутричерепной аневризмы . [255]
Сравнительная анатомия [ править ]
Человеческий мозг обладает многими свойствами, общими для мозга всех позвоночных . [256] Многие из его особенностей являются общими для мозга всех млекопитающих . [257] в первую очередь шестислойная кора головного мозга и набор связанных с ней структур, [258] включая гиппокамп и миндалевидное тело . [259] Кора головного мозга у человека пропорционально больше, чем у многих других млекопитающих. [260] У людей больше ассоциативной коры, сенсорных и моторных частей, чем у более мелких млекопитающих, таких как крыса и кошка. [261]
Как и мозг приматов , человеческий мозг имеет гораздо большую кору головного мозга (пропорционально размеру тела), чем у большинства млекопитающих. [259] и высокоразвитая зрительная система. [262] [263]
Как мозг гоминида , человеческий мозг значительно увеличен даже по сравнению с мозгом типичной обезьяны. Последовательность эволюции человека от австралопитека (четыре миллиона лет назад) до человека разумного (современного человека) была отмечена устойчивым увеличением размера мозга. [264] [265] Увеличение размера мозга привело к изменению размера и формы черепа. [266] примерно от 600 см 3 у Homo habilis в среднем около 1520 см. 3 у Homo neanderthalensis . [267] Различия в ДНК , экспрессии генов и взаимодействиях генов и окружающей среды помогают объяснить различия между функциями мозга человека и других приматов. [268]
См. также [ править ]
- Очертание человеческого мозга
- Очерк нейробиологии
- Церебральная атрофия
- Кортикальная распространяющаяся депрессия
- Эволюция человеческого интеллекта
- Крупномасштабные мозговые сети
- Поверхностные вены головного мозга
Ссылки [ править ]
- ^ « Энцефало -этимология» . Интернет-словарь этимологии . Архивировано из оригинала 2 октября 2017 года . Проверено 24 октября 2015 г.
- ^ Родитель, А.; Карпентер, МБ (1995). «Ч. 1». Нейроанатомия человека Карпентера . Уильямс и Уилкинс. ISBN 978-0-683-06752-1 .
- ↑ Перейти обратно: Перейти обратно: а б Бигос, КЛ; Харири, А.; Вайнбергер, Д. (2015). Нейровизуализационная генетика: принципы и практика . Издательство Оксфордского университета . п. 157. ИСБН 978-0-19-992022-8 .
- ↑ Перейти обратно: Перейти обратно: а б Косгроув, КП; Мазуре, СМ; Стейли, Дж. К. (2007). «Развитие знаний о половых различиях в структуре, функциях и химии мозга» . Биологическая психиатрия . 62 (8): 847–855. doi : 10.1016/j.biopsych.2007.03.001 . ПМК 2711771 . ПМИД 17544382 .
- ^ Молина, Д. Кимберли; ДиМайо, Винсент Дж. М. (2012). «Нормальный вес органов у мужчин». Американский журнал судебной медицины и патологии . 33 (4): 368–372. дои : 10.1097/PAF.0b013e31823d29ad . ISSN 0195-7910 . ПМИД 22182984 . S2CID 32174574 .
- ^ Молина, Д. Кимберли; ДиМайо, Винсент Дж. М. (2015). «Нормальный вес органов у женщин». Американский журнал судебной медицины и патологии . 36 (3): 182–187. дои : 10.1097/PAF.0000000000000175 . ISSN 0195-7910 . ПМИД 26108038 . S2CID 25319215 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Анатомия Грея 2008 , стр. 227–9.
- ↑ Перейти обратно: Перейти обратно: а б Анатомия Грея 2008 , стр. 335–7.
- ↑ Перейти обратно: Перейти обратно: а б Рибас, ГК (2010). «Мозговые борозды и извилины» . Нейрохирургический фокус . 28 (2): 7. doi : 10.3171/2009.11.FOCUS09245 . ПМИД 20121437 .
- ^ Фригери, Т.; Пальоли, Э.; Де Оливейра, Э.; Ротон-младший, Алабама (2015). «Микрохирургическая анатомия центральной доли». Журнал нейрохирургии . 122 (3): 483–98. дои : 10.3171/2014.11.JNS14315 . ПМИД 25555079 .
- ^ Мёллгорд, Кьельд; Бейнлих, Феликс Р.М.; Куск, Питер; Миякоши, Лео М.; Делле, Кристина; Пла, Вирджиния; Хаугланд, Натали Л.; Эсмаил, Тина; Расмуссен, Мартин К.; Гомолка, Рышард С.; Мори, Юки; Недергаард, Майкен (2023). «Мезотелий делит субарахноидальное пространство на функциональные отсеки» . Наука . 379 (6627): 84–88. Бибкод : 2023Sci...379...84M . doi : 10.1126/science.adc8810 . ПМИД 36603070 . S2CID 255440992 .
- Медицинский центр Рочестерского университета (5 января 2023 г.). «Недавно открытая анатомия защищает и контролирует мозг» . Медицинский Экспресс . Архивировано из оригинала 7 января 2023 года . Проверено 15 февраля 2023 г.
- ^ Первес 2012 , с. 724.
- ↑ Перейти обратно: Перейти обратно: а б Чиполла, MJ (1 января 2009 г.). «Анатомия и ультраструктура» . Церебральное кровообращение . Морган и Клейпул Науки о жизни. Архивировано из оригинала 1 октября 2017 года на NCBI Bookshelf.
- ^ «Взгляд на мозг глазами хирурга» . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . Свежий воздух. 10 мая 2006 г. Архивировано из оригинала 7 ноября 2017 г.
- ^ Сампайо-Баптиста, К; Йохансен-Берг, Х. (20 декабря 2017 г.). «Пластичность белого вещества в мозгу взрослого человека» . Нейрон . 96 (6): 1239–1251. дои : 10.1016/j.neuron.2017.11.026 . ПМК 5766826 . ПМИД 29268094 .
- ^ Дэйви, Г. (2011). Прикладная психология . Джон Уайли и сыновья . п. 153. ИСБН 978-1-4443-3121-9 .
- ^ Арсава, EY; Арсава, Э.М.; Огуз, КК; Топчуоглу, Массачусетс (2019). «Затылочные петалии как прогностический признак доминирования поперечного синуса». Неврологические исследования . 41 (4): 306–311. дои : 10.1080/01616412.2018.1560643 . ПМИД 30601110 . S2CID 58546404 .
- ↑ Перейти обратно: Перейти обратно: а б Акерман, С. (1992). Открытие мозга . Вашингтон, округ Колумбия: Издательство Национальной академии. стр. 22–25 . ISBN 978-0-309-04529-2 .
- ^ Ларсен 2001 , стр. 455–456.
- ^ Кандел, скорая помощь; Шварц, Дж. Х.; Джессел ТМ (2000). Принципы нейронауки . МакГроу-Хилл Профессионал. п. 324 . ISBN 978-0-8385-7701-1 .
- ^ Гайтон и Холл 2011 , с. 574.
- ^ Гайтон и Холл 2011 , с. 667.
- ^ Принципы анатомии и физиологии, 12-е издание - Тортора, с. 519.
- ↑ Перейти обратно: Перейти обратно: а б с Фреберг, Л. (2009). Открытие биологической психологии . Cengage Обучение . стр. 44–46. ISBN 978-0-547-17779-3 .
- ↑ Перейти обратно: Перейти обратно: а б Колб, Б.; Уишоу, И. (2009). Основы нейропсихологии человека . Макмиллан . стр. 73–75. ISBN 978-0-7167-9586-5 .
- ^ Покок 2006 , с. 64.
- ↑ Перейти обратно: Перейти обратно: а б Первес 2012 , с. 399.
- ^ Анатомия Грея 2008 , стр. 325–6.
- ^ Голл, Ю.; Атлан, Г.; Ситри, А. (август 2015 г.). «Внимание: клауструм». Тенденции в нейронауках . 38 (8): 486–95. дои : 10.1016/j.tins.2015.05.006 . ПМИД 26116988 . S2CID 38353825 .
- ^ Гоард, М.; Дэн, Ю. (4 октября 2009 г.). «Активация базального отдела переднего мозга усиливает корковое кодирование природных сцен» . Природная неврология . 12 (11): 1444–1449. дои : 10.1038/nn.2402 . ПМЦ 3576925 . ПМИД 19801988 .
- ^ Гайтон и Холл 2011 , с. 699.
- ↑ Перейти обратно: Перейти обратно: а б с Анатомия Грея 2008 , с. 298.
- ^ Неттер, Ф. (2014). Атлас анатомии человека, включая интерактивные вспомогательные материалы и руководства для консультаций для студентов (6-е изд.). Филадельфия, Пенсильвания: WB Saunders Co., с. 114. ИСБН 978-1-4557-0418-7 .
- ↑ Перейти обратно: Перейти обратно: а б Анатомия Грея 2008 , с. 297.
- ^ Гайтон и Холл 2011 , стр. 698–9.
- ^ Сквайр 2013 , стр. 761–763.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Анатомия Грея 2008 , с. 275.
- ^ Гайтон и Холл 2011 , с. 691.
- ^ Первес 2012 , с. 377.
- ↑ Перейти обратно: Перейти обратно: а б Азеведо, Ф.; и др. (10 апреля 2009 г.). «Равное количество нейрональных и ненейрональных клеток делает человеческий мозг мозгом примата, увеличенным в изометрическом масштабе». Журнал сравнительной неврологии . 513 (5): 532–541. дои : 10.1002/cne.21974 . ПМИД 19226510 . S2CID 5200449 .
Несмотря на распространенные цитаты о том, что человеческий мозг содержит 100 миллиардов нейронов и в десять раз больше глиальных клеток, абсолютное количество нейронов и глиальных клеток в человеческом мозге остается неизвестным. Здесь мы определяем эти числа с помощью изотропного фракционатора и сравниваем их с ожидаемыми значениями для примата размером с человека. Мы обнаружили, что мозг взрослого мужчины содержит в среднем 86,1 ± 8,1 миллиарда NeuN-положительных клеток («нейронов») и 84,6 ± 9,8 миллиардов NeuN-отрицательных («ненейрональных») клеток.
- ^ Павел, Фиала; Иржи, Валента (1 января 2013 г.). Центральная нервная система . Каролинум Пресс. стр. 79. ISBN 978-80-246-2067-1 .
- ↑ Перейти обратно: Перейти обратно: а б с д Полизоидис, С.; Колеца, Т.; Панайотиду, С.; Ашкан, К.; Теохаридес, TC (2015). «Тучные клетки при менингиомах и воспалении головного мозга» . Журнал нейровоспаления . 12 (1): 170. дои : 10.1186/s12974-015-0388-3 . ПМЦ 4573939 . ПМИД 26377554 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Гайтон и Холл, 2011 , стр. 748–749.
- ^ Будзинский, Дж; Клопоцка, М. (2014). «Ось мозг-кишечник в патогенезе инфекции Helicobacter pylori» . Мир Дж. Гастроэнтерол . 20 (18): 5212–25. дои : 10.3748/wjg.v20.i18.5212 . ПМК 4017036 . ПМИД 24833851 .
- ^ Каработти, М.; Сирокко, А.; Маселли, Массачусетс; Севери, К. (2015). «Ось кишечник-мозг: взаимодействие между кишечной микробиотой, центральной и кишечной нервной системой» . Энн Гастроэнтерол . 28 (2): 203–209. ПМЦ 4367209 . ПМИД 25830558 .
- ^ Шёстедт, Эвелина; Фагерберг, Линн; Халльстрем, Бьорн М.; Хэггмарк, Анна; Мициос, Николай; Нильссон, Питер; Понтен, Фредрик; Хёкфельт, Томас; Улен, Матиас (15 июня 2015 г.). «Определение протеома головного мозга человека с использованием транскриптомики и профилирования на основе антител с акцентом на кору головного мозга» . ПЛОС ОДИН . 10 (6): e0130028. Бибкод : 2015PLoSO..1030028S . дои : 10.1371/journal.pone.0130028 . ISSN 1932-6203 . ПМЦ 4468152 . ПМИД 26076492 .
- ↑ Перейти обратно: Перейти обратно: а б с д Анатомия Грея 2008 , стр. 242–244.
- ^ Первес 2012 , с. 742.
- ^ Анатомия Грея 2008 , с. 243.
- ^ Илифф, Джей-Джей; Недергаард, М. (июнь 2013 г.). «Есть ли церебральная лимфатическая система?» . Гладить . 44 (6 Приложение 1): С93-5. дои : 10.1161/СТРОКЕАХА.112.678698 . ПМК 3699410 . ПМИД 23709744 .
- ^ Гайяр, Ф. «Глимфатический путь» . Radiopaedia.org . Архивировано из оригинала 30 октября 2017 года.
- ↑ Перейти обратно: Перейти обратно: а б с Бацински А., Сюй М., Ван В., Ху Дж. (ноябрь 2017 г.). «Параваскулярный путь клиренса мозговых отходов: современное понимание, значение и противоречия» . Границы нейроанатомии . 11 : 101. дои : 10.3389/fnana.2017.00101 . ПМК 5681909 . ПМИД 29163074 .
Параваскулярный путь, также известный как «глимфатический» путь, представляет собой недавно описанную систему удаления отходов в мозге. Согласно этой модели спинномозговая жидкость (СМЖ) поступает в параваскулярные пространства, окружающие пенетрирующие артерии головного мозга, смешивается с интерстициальной жидкостью (ИСЖ) и растворенными веществами в паренхиме и выходит по параваскулярным пространствам дренирующих вен. ... Помимо клиренса Aβ, глифатическая система может участвовать в удалении других интерстициальных растворенных веществ и метаболитов. Измеряя концентрацию лактата в мозге и шейных лимфатических узлах бодрствующих и спящих мышей, Lundgaard et al. (2017) продемонстрировали, что лактат может выходить из ЦНС параваскулярным путем. В их анализе использовалась обоснованная гипотеза о том, что глифатическая функция усиливается во время сна (Xie et al., 2013; Lee et al., 2015; Liu et al., 2017).
- ^ Диссинг-Олесен, Л.; Хонг, С.; Стивенс, Б. (август 2015 г.). «Новые лимфатические сосуды мозга истощают старые представления» . Электронная биомедицина . 2 (8): 776–7. дои : 10.1016/j.ebiom.2015.08.019 . ПМЦ 4563157 . ПМИД 26425672 .
- ↑ Перейти обратно: Перейти обратно: а б Вс, БЛ; Ван, Л.Х.; Ян, Т; Солнце, JY; Мао, LL; Ян, МФ; Юань, Х; Колвин, РА; Ян, XY (апрель 2018 г.). «Лимфодренажная система головного мозга: новая мишень для лечения неврологических заболеваний». Прогресс нейробиологии . 163–164: 118–143. doi : 10.1016/j.pneurobio.2017.08.007 . ПМИД 28903061 . S2CID 6290040 .
- ^ Анатомия Грея 2008 , с. 247.
- ^ Анатомия Грея 2008 , стр. 251–2.
- ↑ Перейти обратно: Перейти обратно: а б с Анатомия Грея 2008 , с. 250.
- ↑ Перейти обратно: Перейти обратно: а б Анатомия Грея 2008 , с. 248.
- ↑ Перейти обратно: Перейти обратно: а б Анатомия Грея 2008 , с. 251.
- ↑ Перейти обратно: Перейти обратно: а б с Анатомия Грея 2008 , стр. 254–6.
- ↑ Перейти обратно: Перейти обратно: а б с д и Elsevier's, 2007 , стр. 311–4.
- ^ Дейнман, Р.; Чжоу, Л.; Кебеде, А.А.; Баррес, бакалавр наук (25 ноября 2010 г.). «Перициты необходимы для целостности гематоэнцефалического барьера во время эмбриогенеза» . Природа . 468 (7323): 562–6. Бибкод : 2010Natur.468..562D . дои : 10.1038/nature09513 . ПМК 3241506 . ПМИД 20944625 .
- ^ Латерра, Дж.; Держите, Р.; Бетц, Луизиана; и др. (1999). «Барьер кровь-спинномозговая жидкость» . Базовая нейрохимия: молекулярные, клеточные и медицинские аспекты (6-е изд.). Филадельфия: Липпинкотт-Рэйвен.
- ^ Сэдлер, Т. (2010). Медицинская эмбриология Лангмана (11-е изд.). Филадельфия: Липпинкотт Уильямс и Уилкинс. п. 293. ИСБН 978-0-7817-9069-7 .
- ↑ Перейти обратно: Перейти обратно: а б Ларсен 2001 , стр. 419.
- ^ Чжоу, И; Сон, Хунцзюнь; Мин, Го-Ли (28 июля 2023 г.). «Генетика развития мозга человека» . Обзоры природы. Генетика . 25 (1): 26–45. дои : 10.1038/s41576-023-00626-5 . ISSN 1471-0064 . ПМК 10926850. ПМИД 37507490 . S2CID 260286574 .
- ↑ Перейти обратно: Перейти обратно: а б с Ларсен 2001 , стр. 85–88.
- ^ Первес 2012 , стр. 480–482.
- ↑ Перейти обратно: Перейти обратно: а б с д Ларсен 2001 , стр. 445–446.
- ^ «ОпенСтакс CNX» . cnx.org . Архивировано из оригинала 5 мая 2015 года . Проверено 5 мая 2015 г.
- ^ Ларсен 2001 , стр. 85–87.
- ^ Первес 2012 , стр. 481–484.
- ^ Первс, Дейл; Августин, Джордж Дж; Фитцпатрик, Дэвид; Кац, Лоуренс С; ЛаМантия, Энтони-Самуэль; Макнамара, Джеймс О; Уильямс, С. Марк, ред. (2001). «Ромбомеры» . Нейронаука (2-е изд.). Пэлгрейв Макмиллан. ISBN 978-0-87893-742-4 .
- ↑ Перейти обратно: Перейти обратно: а б Чен, X. (2012). Механическая самосборка: наука и приложения . Springer Science & Business Media . стр. 188–189. ISBN 978-1-4614-4562-3 .
- ↑ Перейти обратно: Перейти обратно: а б с Ронан, Л; Воэтс, Н; Руа, С; Александр-Блох, А; Хаф, М; Маккей, К; Кроу, Ти Джей; Джеймс, А; Гидд, Дж. Н.; Флетчер, ПК (август 2014 г.). «Дифференциальное тангенциальное расширение как механизм корковой гирификации» . Кора головного мозга . 24 (8): 2219–28. дои : 10.1093/cercor/bht082 . ПМК 4089386 . ПМИД 23542881 .
- ^ Ван Эссен, округ Колумбия (23 января 1997 г.). «Теория морфогенеза и компактной проводки в центральной нервной системе, основанная на напряжении». Природа . 385 (6614): 313–8. Бибкод : 1997Natur.385..313E . дои : 10.1038/385313a0 . ПМИД 9002514 . S2CID 4355025 .
- ^ Боррелл, В. (24 января 2018 г.). «Как клетки сгибают кору головного мозга» . Журнал неврологии . 38 (4): 776–783. doi : 10.1523/JNEUROSCI.1106-17.2017 . ПМК 6596235 . ПМИД 29367288 .
- ^ Флорио, М.; и др. (27 марта 2015 г.). «Человеческий ген ARHGAP11B способствует амплификации базальных предшественников и расширению неокортекса» . Наука . 347 (6229): 1465–70. Бибкод : 2015Sci...347.1465F . дои : 10.1126/science.aaa1975 . ПМИД 25721503 . S2CID 34506325 .
- ^ «Части мозга | Введение в психологию» . Courses.lumenlearning.com . Проверено 20 сентября 2019 г.
- ^ Гайтон и Холл 2011 , с. 685.
- ↑ Перейти обратно: Перейти обратно: а б Гайтон и Холл 2011 , с. 687.
- ↑ Перейти обратно: Перейти обратно: а б Гайтон и Холл 2011 , с. 686.
- ^ Гайтон и Холл 2011 , стр. 698, 708.
- ^ Дэвидсона, 2010 , с. 1139.
- ↑ Перейти обратно: Перейти обратно: а б Хеллиер, Дж. (2014). Мозг, нервная система и их болезни [3 тома] . АВС-КЛИО . стр. 300–303. ISBN 978-1-61069-338-7 .
- ↑ Перейти обратно: Перейти обратно: а б Гайтон и Холл, 2011 , стр. 571–576.
- ^ Гайтон и Холл 2011 , стр. 573–574.
- ^ Гайтон и Холл 2011 , стр. 623–631.
- ^ Гайтон и Холл 2011 , стр. 739–740.
- ^ Покок 2006 , стр. 138–139.
- ^ Сквайр 2013 , стр. 525–526.
- ^ Гайтон и Холл 2011 , стр. 647–648.
- ^ Гайтон и Холл 2011 , стр. 202–203.
- ^ Гайтон и Холл 2011 , стр. 205–208.
- ↑ Перейти обратно: Перейти обратно: а б с д Гайтон и Холл, 2011 , стр. 505–509.
- ^ «Основы работы мозга: понимание сна | Национальный институт неврологических расстройств и инсульта» . www.ninds.nih.gov . Архивировано из оригинала 22 декабря 2017 года.
- ^ Гайтон и Холл 2011 , с. 723.
- ^ Дэвис, Дж. Ф.; Чой, Д.Л.; Бенуа, Южная Каролина (2011). «24. Поведение и питание орексигенных гипоталамических пептидов - 24,5 орексин» . В Приди, VR; Уотсон, Р.Р.; Мартин, ЧР (ред.). Справочник по поведению, еде и питанию . Спрингер. стр. 361–362. ISBN 978-0-387-92271-3 .
- ^ Сквайр 2013 , с. 800.
- ^ Сквайр 2013 , с. 803.
- ^ Сквайр 2013 , с. 805.
- ^ Гайтон и Холл 2011 , стр. 720–2.
- ^ Поппель, Д .; Эммори, К .; Хикок, Г.; Пюлкканен, Л. (10 октября 2012 г.). «На пути к новой нейробиологии языка» . Журнал неврологии . 32 (41): 14125–14131. doi : 10.1523/JNEUROSCI.3244-12.2012 . ПМК 3495005 . ПМИД 23055482 .
- ^ Хикок, Дж. (сентябрь 2009 г.). «Функциональная нейроанатомия языка» . Обзоры физики жизни . 6 (3): 121–143. Бибкод : 2009PhLRv...6..121H . дои : 10.1016/j.plrev.2009.06.001 . ПМК 2747108 . ПМИД 20161054 .
- ^ Федоренко Е.; Канвишер, Н. (2009). «Нейровизуализация языка: почему не появилась более ясная картина?» . Язык и лингвистический компас . 3 (4): 839–865. дои : 10.1111/j.1749-818x.2009.00143.x . S2CID 2833893 .
- ^ Дамасио, Х. (2001). «Нейральная основа речевых расстройств». В Чапи, Роберта (ред.). Стратегии языкового вмешательства при афазии и связанных с ней нейрогенных коммуникативных расстройствах (4-е изд.). Липпинкотт Уильямс и Уилкинс. стр. 18–36. ISBN 978-0-7817-2133-2 . OCLC 45952164 .
- ^ де Люссане, MHE; Оссе, JWM (2012). «Наследственное осевое скручивание объясняет контралатеральную переднюю часть и перекрест зрительных нервов у позвоночных». Биология животных . 62 (2): 193–216. arXiv : 1003.1872 . дои : 10.1163/157075611X617102 . S2CID 7399128 .
- ^ Хеллиер, Дж. (2014). Мозг, нервная система и их болезни [3 тома] . АВС-КЛИО . п. 1135. ИСБН 978-1-61069-338-7 .
- ^ Колб, Б.; Уишоу, IQ (2013). Введение в мозг и поведение . Макмиллан Высшее образование . п. 296. ИСБН 978-1-4641-3960-4 .
- ^ Бернтсон, Г.; Качиоппо, Дж. (2009). Справочник по нейронаукам для поведенческих наук, том 1 . Джон Уайли и сыновья . п. 145. ИСБН 978-0-470-08355-0 .
- ^ Шервуд, Л. (2012). Физиология человека: от клеток к системам . Cengage Обучение . п. 181. ИСБН 978-1-133-70853-7 .
- ^ Калат, Дж (2015). Биологическая психология . Cengage Обучение . п. 425. ИСБН 978-1-305-46529-9 .
- ↑ Перейти обратно: Перейти обратно: а б Коуин, Южная Каролина; Доти, С.Б. (2007). Тканевая механика . Springer Science & Business Media . п. 4. ISBN 978-0-387-49985-7 .
- ↑ Перейти обратно: Перейти обратно: а б Моррис, CG; Майсто, А.А. (2011). Понимание психологии . Прентис Холл . п. 56. ИСБН 978-0-205-76906-3 .
- ↑ Перейти обратно: Перейти обратно: а б Колб, Б.; Уишоу, IQ (2013). Введение в мозг и поведение (вкладыш) . Макмиллан Высшее образование . стр. 524–549. ISBN 978-1-4641-3960-4 .
- ^ Шактер, Д.Л.; Гилберт, DT; Вегнер, DM (2009). Знакомство с психологией . Макмиллан . п. 80. ИСБН 978-1-4292-1821-4 .
- ^ Сандер, Дэвид (2013). Армони, Дж.; Вийемье, Патрик (ред.). Кембриджский справочник по аффективной нейробиологии человека . Кембридж: Кембриджский университет. Нажимать. п. 16. ISBN 978-0-521-17155-7 .
- ^ Линдквист, Калифорния; Вейгер, Т.Д.; Кобер, Х; Блисс-Моро, Э; Барретт, Л.Ф. (23 мая 2012 г.). «Мозговая основа эмоций: метааналитический обзор» . Поведенческие и мозговые науки . 35 (3): 121–143. дои : 10.1017/S0140525X11000446 . ПМЦ 4329228 . ПМИД 22617651 .
- ^ Фан, КЛ; Вейджер, Тор; Тейлор, Сан-Франциско; Либерзон, л (1 июня 2002 г.). «Функциональная нейроанатомия эмоций: метаанализ исследований активации эмоций с помощью ПЭТ и фМРТ». НейроИмидж . 16 (2): 331–348. дои : 10.1006/нимг.2002.1087 . ПМИД 12030820 . S2CID 7150871 .
- ^ Маленка, РЦ; Нестлер, Э.Дж.; Хайман, SE (2009). "Предисловие". В Сидоре, А; Браун, Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. п. xiii. ISBN 978-0-07-148127-4 .
- ↑ Перейти обратно: Перейти обратно: а б с д Маленка Р.Ц., Нестлер Э.Дж., Хайман С.Е., Хольцман Д.М. (2015). «Глава 14: Высшие когнитивные функции и поведенческий контроль». Молекулярная нейрофармакология: фонд клинической неврологии (3-е изд.). Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-182770-6 .
- ↑ Перейти обратно: Перейти обратно: а б Маленка Р.Ц., Нестлер Э.Дж., Хайман С.Е., Хольцман Д.М. (2015). «Глава 6: Широко распространенные системы: моноамины, ацетилхолин и орексин». Молекулярная нейрофармакология: фонд клинической неврологии (3-е изд.). Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-182770-6 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Даймонд, А (2013). «Исполнительные функции» . Ежегодный обзор психологии . 64 : 135–168. doi : 10.1146/annurev-psych-113011-143750 . ПМК 4084861 . ПМИД 23020641 .
Рисунок 4. Исполнительные функции и связанные с ними термины. Архивировано 9 мая 2018 г. на Wayback Machine. - ↑ Перейти обратно: Перейти обратно: а б с д Хён, Джей Си; Вейандт, LL; Свентоски, А. (2014). «Глава 2: Физиология исполнительного функционирования» . В Гольдштейне, С.; Наглиери, Дж. (ред.). Справочник исполнительного функционирования . Нью-Йорк: Спрингер. стр. 13–23. ISBN 978-1-4614-8106-5 .
- ↑ Перейти обратно: Перейти обратно: а б Маленка Р.Ц., Нестлер Э.Дж., Хайман С.Е., Хольцман Д.М. (2015). «Глава 14: Высшие когнитивные функции и поведенческий контроль». Молекулярная нейрофармакология: фонд клинической неврологии (3-е изд.). Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-182770-6 .
В условиях, когда преобладающие реакции имеют тенденцию доминировать в поведении, например, при наркозависимости, когда сигналы о наркотике могут вызвать поиск наркотика (глава 16), или при синдроме дефицита внимания и гиперактивности (СДВГ; описан ниже), могут возникнуть серьезные негативные последствия. ... СДВГ можно представить как расстройство исполнительной функции; в частности, СДВГ характеризуется снижением способности проявлять и поддерживать когнитивный контроль над поведением. По сравнению со здоровыми людьми, у людей с СДВГ снижена способность подавлять неадекватные доминантные реакции на раздражители (нарушенное торможение ответов) и уменьшенная способность подавлять ответы на нерелевантные раздражители (нарушенное подавление помех). ... Функциональная нейровизуализация у людей демонстрирует активацию префронтальной коры и хвостатого ядра (часть дорсального полосатого тела) при выполнении задач, требующих тормозного контроля поведения. ... Ранние результаты структурной МРТ показывают, что кора головного мозга на большей части головного мозга у субъектов с СДВГ тоньше по сравнению с контрольной группой того же возраста, включая области префронтальной коры, участвующие в рабочей памяти и внимании.
- ^ Покок 2006 , с. 68.
- ^ Кларк, Б.Д.; Гольдберг, Э.М.; Руди, Б. (декабрь 2009 г.). «Электрогенная настройка начального сегмента аксона» . Нейробиолог . 15 (6): 651–68. дои : 10.1177/1073858409341973 . ПМЦ 2951114 . ПМИД 20007821 .
- ^ Покок 2006 , стр. 70–74.
- ↑ Перейти обратно: Перейти обратно: а б «НИМЗ »Основы мозга» . www.nimh.nih.gov . Архивировано из оригинала 26 марта 2017 года . Проверено 26 марта 2017 г.
- ^ Первс, Дейл (2011). Нейронаука (5-е изд.). Сандерленд, Массачусетс: Синауэр. п. 139. ИСБН 978-0-87893-695-3 .
- ^ Сваминатан, Н. (29 апреля 2008 г.). «Почему мозгу нужно так много энергии?» . Научный американец . Архивировано из оригинала 27 января 2014 года . Проверено 19 ноября 2010 г.
- ↑ Перейти обратно: Перейти обратно: а б Вассерман Д.Х. (январь 2009 г.). «Четыре грамма глюкозы» . Американский журнал физиологии. Эндокринология и обмен веществ . 296 (1): Е11–21. дои : 10.1152/ajpendo.90563.2008 . ПМК 2636990 . ПМИД 18840763 .
В крови человека массой 70 кг циркулирует четыре грамма глюкозы. Эта глюкоза имеет решающее значение для нормального функционирования многих типов клеток. В соответствии с важностью этих 4 г глюкозы существует сложная система контроля для поддержания постоянного уровня глюкозы в крови. Наше внимание было сосредоточено на механизмах, с помощью которых регулируется поток глюкозы из печени в кровь и из крови в скелетные мышцы. ... Мозг потребляет ~60% глюкозы в крови человека, ведущего сидячий образ жизни и голодающего. ...Количество глюкозы в крови сохраняется за счет резервуаров гликогена (рис. 2). У людей после абсорбции около 100 г гликогена содержится в печени и около 400 г гликогена в мышцах. Окисление углеводов работающими мышцами может увеличиться примерно в 10 раз при физической нагрузке, но через 1 час уровень глюкозы в крови поддерживается на уровне примерно 4 г. ... Сейчас точно известно, что и инсулин, и физические упражнения вызывают транслокацию GLUT4 в плазматическую мембрану. За исключением фундаментального процесса транслокации GLUT4, [поглощение глюкозы мышцами (MGU)] контролируется по-разному с помощью упражнений и инсулина. Стимулируемая сокращением внутриклеточная передача сигналов (52, 80) и MGU (34, 75, 77, 88, 91, 98) не зависят от инсулина. Более того, судьба глюкозы, экстрагированной из крови, различается в зависимости от физической нагрузки и инсулина (91, 105). По этим причинам барьеры для потока глюкозы из крови в мышцы должны быть определены независимо для этих двух регуляторов MGU.
- ^ Квисторф, Б; Сечер, Н; Ван Лисхаут, Дж. (24 июля 2008 г.). «Лактат питает человеческий мозг во время тренировок» . Журнал ФАСЭБ . 22 (10): 3443–3449. дои : 10.1096/fj.08-106104 . ПМИД 18653766 . S2CID 15394163 .
- ^ Обель, Л.Ф.; Мюллер, М.С.; Стены, AB; Зикманн, HM; Бак, ЛК; Ваагепетерсен, HS; Шусбо, А. (2012). «Мозговой гликоген – новые взгляды на его метаболическую функцию и регуляцию на субклеточном уровне» . Границы нейроэнергетики . 4 :3. дои : 10.3389/fnene.2012.00003 . ПМК 3291878 . ПМИД 22403540 .
- ^ Марин-Валенсия, И.; и др. (февраль 2013 г.). «Гептаноат как нервное топливо: энергетические предшественники нейромедиаторов в нормальном мозге и мозге с дефицитом транспортера глюкозы I (G1D)» . Журнал церебрального кровотока и метаболизма . 33 (2): 175–82. дои : 10.1038/jcbfm.2012.151 . ПМК 3564188 . ПМИД 23072752 .
- ^ Цудзи, А. (2005). «Малогомолекулярный перенос лекарств через гематоэнцефалический барьер через транспортные системы, опосредованные переносчиками» . НейроРкс . 2 (1): 54–62. дои : 10.1602/neurorx.2.1.54 . ПМК 539320 . ПМИД 15717057 .
Поглощение вальпроевой кислоты снижалось в присутствии жирных кислот со средней длиной цепи, таких как гексаноат, октаноат и деканоат, но не пропионата или бутирата, что указывает на то, что вальпроевая кислота попадает в мозг через систему транспорта жирных кислот со средней длиной цепи. , а не жирные кислоты с короткой цепью. ... На основании этих сообщений считается, что вальпроевая кислота транспортируется в двух направлениях между кровью и мозгом через ГЭБ с помощью двух различных механизмов: переносчиков, чувствительных к монокарбоновой кислоте, и переносчиков, чувствительных к среднецепочечным жирным кислотам, для оттока и поглощения соответственно.
- ^ Виджай, Н.; Моррис, Мэн (2014). «Роль переносчиков монокарбоксилатов в доставке лекарств в мозг» . Курс. Фарм. Дес . 20 (10): 1487–98. дои : 10.2174/13816128113199990462 . ПМК 4084603 . ПМИД 23789956 .
Известно, что переносчики монокарбоксилатов (MCT) опосредуют транспорт монокарбоксилатов с короткой цепью, таких как лактат, пируват и бутират. ... MCT1 и MCT4 также связаны с транспортом короткоцепочечных жирных кислот, таких как ацетат и формиат, которые затем метаболизируются в астроцитах [78].
- ^ Кларк, Д.Д.; Соколов. Л. (1999). Сигел, Дж.Дж.; Агранов, Б.В.; Альберс, RW; Фишер, СК; Улер, доктор медицины (ред.). Базовая нейрохимия: молекулярные, клеточные и медицинские аспекты . Филадельфия: Липпинкотт. стр. 637–670. ISBN 978-0-397-51820-3 .
- ^ Мрсуля, Б.Б. (2012). Патофизиология церебрального энергетического обмена . Springer Science & Business Media . стр. 2–3. ISBN 978-1-4684-3348-7 .
- ^ Райхл, М.; Гуснард, Д.А. (2002). «Оценка энергетического баланса мозга» . Учеб. Натл. акад. наук. США . 99 (16): 10237–10239. Бибкод : 2002PNAS...9910237R . дои : 10.1073/pnas.172399499 . ПМЦ 124895 . ПМИД 12149485 .
- ^ Джанарос, Питер Дж.; Грей, Маркус А.; Оньевуэни, Икечукву; Кричли, Хьюго Д. (2010). «Методы нейровизуализации в поведенческой медицине» . В Стептоу, А. (ред.). Справочник по поведенческой медицине . Springer Science & Business Media . п. 770. дои : 10.1007/978-0-387-09488-5_50 . ISBN 978-0-387-09488-5 .
- ^ Кузава, CW; Чугани, ХТ; Гроссман, Л.И.; Липович Л.; Музик, О.; Хоф, PR; Уайлдман, Делавэр; Шервуд, CC; Леонард, WR; Ланге, Н. (9 сентября 2014 г.). «Метаболические затраты и эволюционные последствия развития человеческого мозга» . Труды Национальной академии наук . 111 (36): 13010–13015. Бибкод : 2014PNAS..11113010K . дои : 10.1073/pnas.1323099111 . ISSN 0027-8424 . ПМК 4246958 . ПМИД 25157149 .
- ^ «Мозг может вымывать токсины во время сна» . Национальные институты здравоохранения . Архивировано из оригинала 20 октября 2013 года . Проверено 25 октября 2013 г.
- ^ Се Л., Кан Х., Сюй Кью, Чен М.Дж., Ляо Ю., Тиягараджан М., О'Доннелл Дж., Кристенсен Д.Д., Николсон С., Илифф Дж.Дж., Такано Т., Дин Р., Недергаард М. (октябрь 2013 г.). «Сон ускоряет выведение метаболитов из мозга взрослого человека» . Наука . 342 (6156): 373–377. Бибкод : 2013Sci...342..373X . дои : 10.1126/science.1241224 . ПМК 3880190 . ПМИД 24136970 .
Таким образом, восстановительная функция сна может быть следствием усиленного удаления потенциально нейротоксичных продуктов жизнедеятельности, накапливающихся в бодрствующей центральной нервной системе.
- ^ Тонони, Джулио; Чирелли, Кьяра (август 2013 г.). «Вероятность обрезки» (PDF) . Научный американец . 309 (2): 34–39. Бибкод : 2013SciAm.309b..34T . doi : 10.1038/scientificamerican0813-34 . ПМИД 23923204 . S2CID 54052089 . Архивировано из оригинала (PDF) 26 декабря 2018 г.
- ↑ Перейти обратно: Перейти обратно: а б Ван Эссен, округ Колумбия; и др. (октябрь 2012 г.). «Проект Human Connectome: перспективы сбора данных» . НейроИмидж . 62 (4): 2222–2231. doi : 10.1016/j.neuroimage.2012.02.018 . ПМК 3606888 . ПМИД 22366334 .
- ^ Джонс, Э.Г .; Менделл, Л.М. (30 апреля 1999 г.). «Оценка десятилетия мозга». Наука . 284 (5415): 739. Бибкод : 1999Sci...284..739J . дои : 10.1126/science.284.5415.739 . ПМИД 10336393 . S2CID 13261978 .
- ^ «Цена инициативы BRAIN в 4,5 миллиарда долларов?» . Наука | АААС . 5 июня 2014 г. Архивировано из оригинала 18 июня 2017 г.
- ^ Фань, Сюэ; Маркрам, Генри (7 мая 2019 г.). «Краткая история симуляционной нейронауки» . Границы нейроинформатики . 13:32 . дои : 10.3389/fninf.2019.00032 . ISSN 1662-5196 . ПМК 6513977 . ПМИД 31133838 .
- ^ Таул, В.Л.; и др. (январь 1993 г.). «Пространственное расположение электродов ЭЭГ: поиск наиболее подходящей сферы относительно кортикальной анатомии». Электроэнцефалография и клиническая нейрофизиология . 86 (1): 1–6. дои : 10.1016/0013-4694(93)90061-у . ПМИД 7678386 .
- ^ Первес 2012 , стр. 632–633.
- ^ Сильверстайн, Дж. (2012). «Картирование моторной и сенсорной коры: исторический взгляд и современное исследование сенсомоторной локализации и прямой кортикальной моторной стимуляции» . Нейродиагностический журнал . 52 (1): 54–68. ПМИД 22558647 . Архивировано из оригинала 17 ноября 2012 года.
- ^ Боро, Т.; Безард, Э.; и др. (2002). «От регистрации отдельных внеклеточных единиц при экспериментальном и человеческом паркинсонизме до разработки функциональной концепции роли базальных ганглиев в двигательном контроле». Прогресс нейробиологии . 66 (4): 265–283. дои : 10.1016/s0301-0082(01)00033-8 . ПМИД 11960681 . S2CID 23389986 .
- ^ Ланкастер, Массачусетс; Реннер, М; Мартин, Калифорния; Венцель, Д; Бикнелл, Л.С.; Херлс, Мэн; Хомфрей, Т; Пеннингер, Дж. М.; Джексон, AP; Кноблих, Ю.А. (19 сентября 2013 г.). «Церебральные органоиды моделируют развитие мозга человека и микроцефалию» . Природа . 501 (7467): 373–9. Бибкод : 2013Natur.501..373L . дои : 10.1038/nature12517 . ПМЦ 3817409 . ПМИД 23995685 .
- ^ Ли, Коннектикут; Бендрием, РМ; Ву, WW; Шен, РФ (20 августа 2017 г.). «3D-органоиды мозга, полученные из плюрипотентных стволовых клеток: многообещающие экспериментальные модели развития мозга и нейродегенеративных расстройств» . Журнал биомедицинской науки . 24 (1): 59. дои : 10.1186/s12929-017-0362-8 . ПМЦ 5563385 . ПМИД 28822354 .
- ^ «Магнитный резонанс, критическое рецензируемое введение; функциональная МРТ» . Европейский форум по магнитному резонансу. Архивировано из оригинала 2 июня 2017 года . Проверено 30 июня 2017 г.
- ^ Бакстон, Р.; Улудаг, К.; Лю, Т. (2004). «Моделирование гемодинамического ответа на активацию мозга». НейроИмидж . 23 : S220–S233. CiteSeerX 10.1.1.329.29 . doi : 10.1016/j.neuroimage.2004.07.013 . ПМИД 15501093 . S2CID 8736954 .
- ^ Бисвал, Б.Б. (15 августа 2012 г.). «ФМРТ в состоянии покоя: личная история». НейроИмидж . 62 (2): 938–44. doi : 10.1016/j.neuroimage.2012.01.090 . ПМИД 22326802 . S2CID 93823 .
- ^ Первес 2012 , с. 20.
- ^ Кейн, РЛ; Парсонс, Т.Д. (2017). Роль технологий в клинической нейропсихологии . Издательство Оксфордского университета . п. 399. ИСБН 978-0-19-023473-7 .
Иримия, Чемберс, Торгерсон и Ван Хорн (2012) представили график первого шага, показывающий, как лучше всего отобразить данные о связях, как показано на рисунке 13.15. Это называется коннектограммой.
- ^ Эндрюс, генеральный директор (2001). Нейропсихология . Психология Пресс. ISBN 978-1-84169-103-9 .
- ^ Лепаж, М. (2010). «Исследования в Центре визуализации мозга» . Университетский институт психического здоровья Дугласа . Архивировано из оригинала 5 марта 2012 года.
- ↑ Перейти обратно: Перейти обратно: а б Стюард, Калифорния; и др. (2017). «Геномная аннотация для клинической геномной диагностики: сильные и слабые стороны» . Геном Мед . 9 (1): 49. дои : 10.1186/s13073-017-0441-1 . ПМЦ 5448149 . ПМИД 28558813 .
- ^ Харроу, Дж.; и др. (сентябрь 2012 г.). «GENCODE: эталонная аннотация генома человека для проекта ENCODE» . Геном Рез . 22 (9): 1760–74. дои : 10.1101/гр.135350.111 . ПМЦ 3431492 . ПМИД 22955987 .
- ^ Гибсон Дж., Муза С.В. (20 апреля 2009 г.). Букварь по геномной науке (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 9780878932368 .
- ^ «Протеом человека в мозге - Атлас белков человека» . www.proteinatlas.org . Архивировано из оригинала 29 сентября 2017 года . Проверено 29 сентября 2017 г.
- ^ Улен, Матиас; Фагерберг, Линн; Халльстрем, Бьорн М.; Линдског, Сесилия; Оксволд, Пер; Мардиноглу, Адиль; Сивертссон, Оса; Кампф, Кэролайн; Шёстедт, Эвелина (23 января 2015 г.). «Тканевая карта протеома человека». Наука . 347 (6220): 1260419. doi : 10.1126/science.1260419 . ISSN 0036-8075 . ПМИД 25613900 . S2CID 802377 .
- ^ Надзиратель, А (2017). «Профиль экспрессии генов в мозгу человека-алкоголика» . Нейрофармакология . 122 : 161–174. doi : 10.1016/j.neuropharm.2017.02.017 . ПМЦ 5479716 . ПМИД 28254370 .
- ^ Фаррис, СП; и др. (2015). «Применение новой геномики к алкогольной зависимости» . Алкоголь . 49 (8): 825–36. дои : 10.1016/j.alcohol.2015.03.001 . ПМК 4586299 . ПМИД 25896098 .
- ^ Розицка, А; Лигуз-Лечнар, М (август 2017 г.). «Пространство, в котором действует старение: сосредоточьтесь на ГАМКергическом синапсе» . Стареющая клетка . 16 (4): 634–643. дои : 10.1111/acel.12605 . ПМК 5506442 . ПМИД 28497576 .
- ^ Флорес, CE; Мендес, П. (2014). «Формирование торможения: структурная пластичность ГАМКергических синапсов, зависящая от активности» . Границы клеточной нейронауки . 8 : 327. дои : 10.3389/fncel.2014.00327 . ПМК 4209871 . ПМИД 25386117 .
- ^ «Черепно-мозговая травма травматическая» . Медциклопедия . ГЭ . Архивировано из оригинала 26 мая 2011 года.
- ^ Даводу, Северная Каролина (9 марта 2017 г.). «Черепно-мозговая травма (ЧМТ) – определение и патофизиология: обзор, эпидемиология, первичная травма» . Медскейп . Архивировано из оригинала 9 апреля 2017 года.
- ^ Дэвидсона, 2010 , стр. 1196–7.
- ^ Винго А.П., Фан В., Дуонг Д.М., Герасимов Э.С., Даммер Э.Б., Лю Й., Хареримана Н.В., Уайт Б., Тамбисетти М., Тронкозо Дж.К., Ким Н., Шнайдер Дж.А., Хаджар И.М., Ла Дж.Дж., Беннетт Д.А., Сейфрид Н.Т., Леви А.И. , Винго ТС (июнь 2020 г.). «Общие протеомные эффекты церебрального атеросклероза и болезни Альцгеймера на мозг человека» . Нат Нейроски . 23 (6): 696–700. дои : 10.1038/s41593-020-0635-5 . ПМК 7269838 . ПМИД 32424284 .
- ↑ Перейти обратно: Перейти обратно: а б Дэвидсона, 2010 , стр. 1205–15.
- ↑ Перейти обратно: Перейти обратно: а б с д и Дэвидсона, 2010 , стр. 1216–7.
- ^ Волков, Северная Дакота; Кооб, Г.Ф.; Маклеллан, AT (январь 2016 г.). «Нейробиологические достижения в модели зависимости от заболеваний головного мозга» . Медицинский журнал Новой Англии . 374 (4): 363–371. дои : 10.1056/NEJMra1511480 . ПМК 6135257 . ПМИД 26816013 .
- ^ Симпсон, Дж. М.; Мориарти, GL (2013). Мультимодальное лечение острых психических заболеваний: руководство по переводу в больницу . Издательство Колумбийского университета . стр. 22–24. ISBN 978-0-231-53609-7 .
- ↑ Перейти обратно: Перейти обратно: а б с д Дэвидсона, 2010 г. , стр. 1172–9.
- ^ «Эпилептический статус» . Фонд эпилепсии .
- ^ Мур, СП (2005). Окончательный обзор Совета по неврологической хирургии . Липпинкотт Уильямс и Уилкинс . п. 112. ИСБН 978-1-4051-0459-3 .
- ^ Пеннингтон, БФ (2008). Диагностика нарушений обучения, второе издание: нейропсихологическая основа . Гилфорд Пресс . стр. 3–10. ISBN 978-1-60623-786-1 .
- ^ Говерт, П.; де Врис, Л.С. (2010). Атлас сонографии головного мозга новорожденных: (CDM 182–183) . Джон Уайли и сыновья . стр. 89–92. ISBN 978-1-898683-56-8 .
- ↑ Перейти обратно: Перейти обратно: а б Перезе, Э.Ф. (2012). Психиатрическая медсестринская практика передовой практики: биопсихологическая основа практики . Ф.А. Дэвис . стр. 82–88. ISBN 978-0-8036-2999-8 .
- ^ Кирни, К.; Трулл, Ти Джей (2016). Аномальная психология и жизнь: многомерный подход . Cengage Обучение . п. 395. ИСБН 978-1-337-09810-6 .
- ^ Стивенсон, ДК; Саншайн, П.; Бенитц, МЫ (2003). Черепно-мозговая травма плода и новорожденного: механизмы, лечение и практические риски . Издательство Кембриджского университета . п. 191. ИСБН 978-0-521-80691-6 .
- ^ Дьюхерст, Джон (2012). Учебник Дьюхерста по акушерству и гинекологии . Джон Уайли и сыновья . п. 43. ИСБН 978-0-470-65457-6 .
- ^ «Артериовенозные мальформации (АВМ) | Национальный институт неврологических расстройств и инсульта» . www.ninds.nih.gov . Проверено 8 февраля 2023 г.
- ^ Харбисон, Дж.; Мэсси, А.; Барнетт, Л.; Ходж, Д.; Форд, Джорджия (июнь 1999 г.). «Протокол скорой помощи при остром инсульте». Ланцет . 353 (9168): 1935. doi : 10.1016/S0140-6736(99)00966-6 . ПМИД 10371574 . S2CID 36692451 .
- ^ Дэвидсона, 2010 , с. 1183.
- ↑ Перейти обратно: Перейти обратно: а б Дэвидсона, 2010 г. , стр. 1180–1.
- ^ Дэвидсон, 2010 , стр. 1181, 1183–1185.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Дэвидсона, 2010 , стр. 1183–1185.
- ↑ Перейти обратно: Перейти обратно: а б Дэвидсона, 2010 г. , стр. 1185–1189.
- ^ Гоял, М.; и др. (апрель 2016 г.). «Эндоваскулярная тромбэктомия после ишемического инсульта крупных сосудов: метаанализ индивидуальных данных пациентов из пяти рандомизированных исследований». Ланцет . 387 (10029): 1723–1731. дои : 10.1016/S0140-6736(16)00163-X . ПМИД 26898852 . S2CID 34799180 .
- ^ Сейвер, Дж. Л. (8 декабря 2005 г.). «Время — это мозг, измеренное количественно» . Гладить . 37 (1): 263–266. дои : 10.1161/01.STR.0000196957.55928.ab . ПМИД 16339467 .
- ^ Уинштейн, CJ; и др. (июнь 2016 г.). «Руководство по реабилитации и восстановлению после инсульта у взрослых» . Гладить . 47 (6): e98–e169. doi : 10.1161/STR.0000000000000098 . ПМИД 27145936 . S2CID 4967333 .
- ^ Кузьма, Эльжбета; Лурида, Илианна; Мур, Сара Ф.; Левин, Дебора А.; Укумунне, Обиоха К.; Ллевеллин, Дэвид Дж. (ноябрь 2018 г.). «Риск инсульта и деменции: систематический обзор и метаанализ» . Болезнь Альцгеймера и деменция . 14 (11): 1416–1426. дои : 10.1016/j.jalz.2018.06.3061 . ISSN 1552-5260 . ПМК 6231970 . ПМИД 30177276 .
- ↑ Перейти обратно: Перейти обратно: а б с д Гойла, АК; Павар, М. (2009). «Диагноз смерть мозга» . Индийский журнал медицины критических состояний . 13 (1): 7–11. дои : 10.4103/0972-5229.53108 . ПМЦ 2772257 . ПМИД 19881172 .
- ↑ Перейти обратно: Перейти обратно: а б с Вейдикс, EFM (8 января 2002 г.). «Смерть мозга во всем мире: общепризнанный факт, но нет глобального консенсуса в диагностических критериях». Неврология . 58 (1): 20–25. дои : 10.1212/wnl.58.1.20 . ПМИД 11781400 . S2CID 219203458 .
- ^ Дханвате, AD (сентябрь 2014 г.). «Смерть ствола мозга: всесторонний обзор с точки зрения Индии» . Индийский журнал медицины критических состояний . 18 (9): 596–605. дои : 10.4103/0972-5229.140151 . ПМК 4166875 . ПМИД 25249744 .
- ↑ Перейти обратно: Перейти обратно: а б с д Дэвидсона, 2010 , с. 1158.
- ^ Дэвидсона, 2010 , с. 200.
- ^ Урден, LD; Стейси, КМ; Лох, Мэн (2013). Приоритеты сестринского ухода в отделениях интенсивной терапии – электронная книга . Elsevier Науки о здоровье . стр. 112–113. ISBN 978-0-323-29414-0 .
- ^ Домингес, Дж. Ф.; Льюис, Эд; Тернер, Р.; Иган, GF (2009). Цзяо, JY (ред.). Мозг в культуре и культура в мозге: обзор основных проблем нейроантропологии . Специальный выпуск: Культурная нейронаука: культурное влияние на функцию мозга. Том. 178. стр. 43–6. дои : 10.1016/S0079-6123(09)17804-4 . ISBN 978-0-444-53361-6 . ПМИД 19874961 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ «Культурная среда влияет на функцию мозга | Psych Central News» . Центральные новости психики . 4 августа 2010 г. Архивировано из оригинала 17 января 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б Макмиллан, Малкольм Б. (2000). Странная слава: Истории Финеаса Гейджа . МТИ Пресс . ISBN 978-0-262-13363-0 .
- ^ Решер, Н. (1992). Монадология Г.В. Лейбница . Психология Пресс. п. 83. ИСБН 978-0-415-07284-7 .
- ^ Харт, WD (1996). Гуттенплан С. (ред.). Спутник философии разума . Блэквелл. стр. 265–267.
- ^ Черчленд, PS (1989). «Гл. 8» . Нейрофилософия . МТИ Пресс. ISBN 978-0-262-53085-9 .
- ^ Селимбейоглу, Аслихан; Парвизи, Дж (2010). «Электрическая стимуляция человеческого мозга: феномены восприятия и поведения, о которых сообщается в старой и новой литературе» . Границы человеческой неврологии . 4 : 46. дои : 10.3389/fnhum.2010.00046 . ПМЦ 2889679 . ПМИД 20577584 .
- ^ Шварц, Дж. Х. Приложение D: Сознание и нейробиология двадцать первого века . В Канделе, скорая помощь; Шварц, Дж. Х.; Джесселл, ТМ (2000). Принципы нейронауки, 4-е издание .
- ^ Лилиенфельд, СО; Линн, С.Дж.; Руссио, Дж.; Байерштейн, Б.Л. (2011). 50 великих мифов популярной психологии: развенчание широко распространенных заблуждений о человеческом поведении . Джон Уайли и сыновья. п. 89. ИСБН 978-1-4443-6074-5 .
- ^ МакДэниел, М. (2005). «Люди с большим мозгом умнее» (PDF) . Интеллект . 33 (4): 337–346. дои : 10.1016/j.intell.2004.11.005 . Архивировано (PDF) из оригинала 6 сентября 2014 г.
- ^ Людерс, Э.; и др. (сентябрь 2008 г.). «Картирование связи между кортикальными извилинами и интеллектом: влияние пола» . Кора головного мозга . 18 (9): 2019–26. дои : 10.1093/cercor/bhm227 . ПМК 2517107 . ПМИД 18089578 .
- ^ Хоппе, К; Стоянович, Дж (2008). «Высокоспособные умы». Научный американский разум . 19 (4): 60–67. doi : 10.1038/scientificamericanmind0808-60 .
- ^ «Тупая беланжери» . Институт генома Вашингтонского университета. Архивировано из оригинала 1 июня 2010 года . Проверено 22 января 2016 г.
- ^ Кэрриер, Мартин; Миттельштрасс, Юрген (1991). философия психологии Разум, мозг, поведение: проблема разума и тела и . Перевод Линдберга, Стивена (переработанное и расширенное английское издание). Берлин: Вальтер де Грюйтер. п. 11. ISBN 9783110128765 . Проверено 22 мая 2021 г.
[...] Аристотелевская точка зрения, что душа обитает прежде всего в сердце [...].
- ^ Кобб, Мэтью (21 апреля 2020 г.). Идея мозга: прошлое и будущее нейронауки . Нью-Йорк: Hachette UK (опубликовано в 2020 г.). ISBN 9781541646865 . Проверено 22 мая 2021 г.
[...] способы, которыми мы думаем о [мозге], стали намного богаче, чем в прошлом, не просто из-за удивительных фактов, которые мы открыли, но, прежде всего, из-за того, как мы их интерпретируем.
- ^ Джарретт, К. (17 ноября 2014 г.). Великие мифы о мозге . Джон Уайли и сыновья. ISBN 978-1-118-31271-1 .
- ^ Филлипс, Хелен (11 июля 2002 г.). «Заявление о «повреждении мозга» видеоигрой подверглось критике» . Новый учёный . Архивировано из оригинала 11 января 2009 года . Проверено 6 февраля 2008 г.
- ^ Попова, Мария (18 августа 2011 г.). « Культура мозга: как нейронаука стала фиксацией поп-культуры» . Атлантика . Архивировано из оригинала 28 июля 2017 года.
- ^ Торнтон, Дэви Джонсон (2011). Мозговая культура. Нейронаука и популярные СМИ . Издательство Университета Рутгерса. ISBN 978-0-8135-5013-8 .
- ↑ Киборги и космос. Архивировано 6 октября 2011 года в Wayback Machine , в журнале «Астронавтика» (сентябрь 1960 г.), Манфредом Э. Клайнсом и Натаном С. Клайном.
- ^ Бергфельдер, Тим (2005). Международные приключения: немецкое популярное кино и европейское совместное производство 1960-х годов . Книги Бергана. п. 129. ИСБН 978-1-57181-538-5 .
- ^ Кандел, ER ; Шварц Дж. Х.; Джесселл ТМ (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-0-8385-7701-1 .
- ↑ Перейти обратно: Перейти обратно: а б с д Гросс, Чарльз Г. (1987). Адельман, Джордж (ред.). Энциклопедия нейробиологии (PDF) (2-е изд.). Бостон: Биркхойзер. стр. 843–847. ISBN 978-0-8176-3335-6 . Архивировано (PDF) из оригинала 5 мая 2013 г.
- ↑ Перейти обратно: Перейти обратно: а б Медведь, МФ; Б.В. Коннорс; М. А. Парадизо (2001). Нейронаука: исследование мозга . Балтимор: Липпинкотт. ISBN 978-0-7817-3944-3 .
- ^ фон Штаден, стр.157
- ^ Суонсон, Ларри В. (12 августа 2014 г.). Нейроанатомическая терминология: лексикон классического происхождения и исторических основ . Издательство Оксфордского университета. ISBN 978-0-19-534062-4 .
- ↑ Перейти обратно: Перейти обратно: а б Лохорст, Герт-Ян (1 января 2016 г.). «Декарт и шишковидная железа» . Стэнфордская энциклопедия философии . Лаборатория метафизических исследований Стэнфордского университета . Проверено 11 марта 2017 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Гросс, Чарльз Г. (1999). Мозг, зрение, память: рассказы из истории нейробиологии (1-е изд. MIT Press pbk.). Кембридж, Массачусетс: Массачусетский технологический институт. стр. 37–51. ISBN 978-0-262-57135-7 .
- ^ Маршалл, Луиза Х.; Магун, Гораций В. (9 марта 2013 г.). Открытия в человеческом мозге: предыстория нейронауки, структура и функции мозга . Springer Science & Business Media. п. 44. ИСБН 978-1-475-74997-7 .
- ^ Хольц, Андерс; Леви, Ричард (20 июля 2010 г.). Травма спинного мозга . Издательство Оксфордского университета. ISBN 978-0-19-970681-5 .
- ^ Тессман, Патрик А.; Суарес, Хосе И. (2002). «Влияние ранней гравюры на развитие нейроанатомии и неврологии». Архив неврологии . 59 (12): 1964–1969. дои : 10.1001/archneur.59.12.1964 . ПМИД 12470188 .
- ^ О'Коннор, Джеймс (2003). «Томас Уиллис и предыстория Cerebri Anatome» . Журнал Королевского медицинского общества . 96 (3): 139–143. дои : 10.1177/014107680309600311 . ПМК 539424 . ПМИД 12612118 .
- ^ ЭМЕРИ, АЛАН (октябрь 2000 г.). «Краткая история неврологии: британский вклад 1660–1910. Под редакцией Ф. КЛИФФОРДА РОУЗА. (стр. 282; иллюстрировано; 25 фунтов стерлингов в мягкой обложке; ISBN 07506 4165 7.) Оксфорд: Баттерворт-Хайнеманн» . Журнал анатомии . 197 (3): 513–518. дои : 10.1046/j.1469-7580.2000.197305131.x . ПМЦ 1468164 .
- ^ Олеско, Кэтрин М.; Холмс, Фредерик Л. (1994). Кахан, Дэвид (ред.). «Эксперимент, количественная оценка и открытие: ранние физиологические исследования Гельмгольца, 1843-50». Герман фон Гельмгольц и основы науки девятнадцатого века. Беркли; Лос-Анджелес; Лондон: Издательство Калифорнийского университета: 50–108.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Саббатини, Ренато М.Э. «Саббатини, RME: Открытие биоэлектричества. Нервная проводимость» . www.cerebromente.org.br . Архивировано из оригинала 26 июня 2017 года . Проверено 10 июня 2017 г.
- ^ Финкельштейн, Габриэль Уорд (2013). Эмиль дю Буа-Реймон: нейробиология, личность и общество в Германии девятнадцатого века . Кембридж, Массачусетс. ISBN 978-1-4619-5032-5 . OCLC 864592470 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Карбовский, Казимеж (14 февраля 2008 г.). «Шестьдесят лет клинической электроэнцефалографии». Европейская неврология . 30 (3): 170–175. дои : 10.1159/000117338 . ПМИД 2192889 .
- ^ Пирс, JMS (17 марта 2009 г.). «Мари-Жан-Пьер Флуранс (1794–1867) и корковая локализация» . Европейская неврология . 61 (5): 311–314. дои : 10.1159/000206858 . ПМИД 19295220 .
- ↑ Перейти обратно: Перейти обратно: а б с Де Карлос, Хуан А.; Боррель, Хосе (август 2007 г.). «Историческое отражение вклада Кахаля и Гольджи в основы нейробиологии». Обзоры исследований мозга . 55 (1): 8–16. дои : 10.1016/j.brainresrev.2007.03.010 . hdl : 10261/62299 . ПМИД 17490748 . S2CID 7266966 .
- ^ Берк, Р.Э. (апрель 2007 г.). «Интегративное действие нервной системы сэра Чарльза Шеррингтона: столетняя оценка» . Мозг . 130 (Часть 4): 887–894. дои : 10.1093/brain/awm022 . ПМИД 17438014 .
- ^ Сквайр, Ларри Р., изд. (1996). История нейробиологии в автобиографии . Вашингтон, округ Колумбия: Общество нейронаук. стр. 475–97. ISBN 978-0-12-660305-7 .
- ^ Коуэн, В.М.; Хартер, Д.Х.; Кандел, ER (2000). «Появление современной нейробиологии: некоторые последствия для неврологии и психиатрии». Ежегодный обзор неврологии . 23 : 345–346. дои : 10.1146/annurev.neuro.23.1.343 . ПМИД 10845068 .
- ^ Брэди, Джозеф В.; Наута, Валле Дж. Х. (22 октября 2013 г.). Принципы, практика и позиции в нейропсихиатрических исследованиях: материалы конференции, состоявшейся в июне 1970 года в Армейском исследовательском институте Уолтера Рида, Вашингтон, округ Колумбия, в честь доктора Дэвида Маккензи Риоча после его выхода на пенсию с поста директора нейропсихиатрического отделения этого института. Институт . Эльзевир. п. VII. ISBN 978-1-4831-5453-4 .
- ^ Адельман, Джордж (15 января 2010 г.). «Программа исследований нейронаук в Массачусетском технологическом институте и начало современной области нейронауки». Журнал истории нейронаук . 19 (1): 15–23. дои : 10.1080/09647040902720651 . ПМИД 20391098 . S2CID 21513317 .
- ↑ Перейти обратно: Перейти обратно: а б Принципы нейронауки, 4-е изд. Эрик Р. Кандел, Джеймс Х. Шварц, Томас М. Джессел, ред. МакГроу-Хилл: Нью-Йорк, штат Нью-Йорк. 2000.
- ^ Папес, JW (февраль 1995 г.). «Предлагаемый механизм эмоций. 1937». Журнал нейропсихиатрии и клинических нейронаук . 7 (1): 103–12. дои : 10.1176/jnp.7.1.103 . ПМИД 7711480 .
- ^ Ламберт, Келли Г. (август 2003 г.). «Жизнь и карьера Пола Маклина». Физиология и поведение . 79 (3): 343–349. дои : 10.1016/S0031-9384(03)00147-1 . ПМИД 12954429 . S2CID 18596574 .
- ^ Чаттерджи, Анджан; Кослетт, Х. Бранч (декабрь 2013 г.). Корни когнитивной нейронауки: поведенческая неврология и нейропсихология . ОУП США. стр. 337–8. ISBN 978-0-19-539554-9 .
- ^ Блисс, Майкл (1 октября 2005 г.). Харви Кушинг: Жизнь в хирургии: Жизнь в хирургии . США: Издательство Оксфордского университета. стр. ix–x. ISBN 978-0-19-534695-4 .
- ^ Кретцер, Р.М.; Кун, Алабама ; Тамарго, Р.Дж. (июнь 2010 г.). «Вклад Уолтера Э. Денди в сосудистую нейрохирургию». Журнал нейрохирургии . 112 (6): 1182–91. дои : 10.3171/2009.7.JNS09737 . ПМИД 20515365 .
- ^ Глис, Пол (2005). Человеческий мозг . Издательство Кембриджского университета. п. 1. ISBN 978-0-521-01781-7 .
- ^ Симпкинс, К. Александр; Симпкинс, Аннеллен М. (2012). Нейронаука для врачей: данные, модели и практика . Springer Science & Business Media . п. 143. ИСБН 978-1-4614-4842-6 .
- ^ Борнштейн, Марк Х.; Лэмб, Майкл Э. (2015). Наука о развитии: учебник для продвинутых пользователей . Психология Пресс . п. 220. ИСБН 978-1-136-28220-1 .
- ↑ Перейти обратно: Перейти обратно: а б Бернштейн, Дуглас (2010). Основы психологии . Cengage Обучение . п. 64. ИСБН 978-0-495-90693-3 .
- ^ Хофман, Мишель А. (27 марта 2014 г.). «Эволюция человеческого мозга: чем больше, тем лучше» . Границы нейроанатомии . 8:15 . дои : 10.3389/fnana.2014.00015 . ПМЦ 3973910 . ПМИД 24723857 .
- ^ Грей, Питер (2002). Психология (4-е изд.). Стоит издательства. ISBN 978-0-7167-5162-5 . OCLC 46640860 .
- ^ Лу, Чжун-Линь; Дошер, Барбара (2013). Визуальная психофизика: от лаборатории к теории . МТИ Пресс . п. 3. ISBN 978-0-262-01945-3 .
- ^ Шарвуд Смит, Майк (2017). Знакомство с языком и познанием . Издательство Кембриджского университета . п. 206. ИСБН 978-1-107-15289-2 .
- ^ Колб, Брайан; Уишоу, Ян К. (2013). Введение в мозг и поведение . Макмиллан Высшее образование . п. 21. ISBN 978-1-4641-3960-4 .
- ^ Ньювенхейс, Рудольф; тен Донкелаар, Ханс Дж.; Николсон, Чарльз (2014). Центральная нервная система позвоночных . Спрингер . стр. 2127. ИСБН 978-3-642-18262-4 .
- ^ Лернер, Ли; Лернер, Бренда Уилмот (2004). Научная энциклопедия Гейла: Фазаны-звезды . Гейл . п. 3759. ИСБН 978-0-7876-7559-2 .
По мере того как положение человека менялось и способ поворота черепа, балансирующего на позвоночнике, мозг расширялся, изменяя форму черепа.
- ^ Бегун, Дэвид Р. (2012). Компаньон палеоантропологии . Джон Уайли и сыновья. п. 388. ИСБН 978-1-118-33237-5 .
- ^ Джонс, Р. (2012). «Нейрогенетика: Что делает человеческий мозг?» . Обзоры природы Неврология . 13 (10): 655. дои : 10.1038/nrn3355 . ПМИД 22992645 . S2CID 44421363 .
Библиография [ править ]
- Колледж, Ники Р.; Уокер, Брайан Р.; Ралстон, Стюарт Х.; Ралстон, ред. (2010). Принципы и медицинская практика Дэвидсона (21-е изд.). Эдинбург: Черчилль Ливингстон/Эльзевир. ISBN 978-0-7020-3085-7 .
- Холл, Джон (2011). Учебник медицинской физиологии Гайтона и Холла (12-е изд.). Филадельфия, Пенсильвания: Сондерс/Эльзевир. ISBN 978-1-4160-4574-8 .
- Ларсен, Уильям Дж. (2001). Эмбриология человека (3-е изд.). Филадельфия, Пенсильвания: Черчилль Ливингстон. ISBN 978-0-443-06583-5 .
- Богарт, Брюс Ян; Орт, Виктория (2007). Интегрированная анатомия и эмбриология Elsevier . Филадельфия, Пенсильвания: Эльзевир Сондерс. ISBN 978-1-4160-3165-9 .
- Покок, Г.; Ричардс, К. (2006). Физиология человека: основы медицины (3-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-856878-0 .
- Первс, Дейл (2012). Нейронаука (5-е изд.). Сандерленд, Массачусетс: Партнеры Sinauer. ISBN 978-0-87893-695-3 .
- Сквайр, Ларри (2013). Фундаментальная нейронаука . Уолтем, Массачусетс: Эльзевир. ISBN 978-0-12-385870-2 .
- Стэндринг, Сьюзен, изд. (2008). Анатомия Грея: анатомические основы клинической практики (40-е изд.). Лондон: Черчилль Ливингстон. ISBN 978-0-8089-2371-8 .
Примечания [ править ]
- ^ В частности глазодвигательный , блоковый нерв , нерв, тройничный нерв , отводящий нерв , лицевой нерв , преддверно-улитковый нерв , языкоглоточный нерв , блуждающий нерв , добавочный нерв и подъязычный нерв . [37]
- ^ Включая вестибулоокулярный рефлекс , роговичный рефлекс , рвотный рефлекс и расширение зрачков в ответ на свет, [203]
- ^ Иллюстрировано архитектором Кристофером Реном. [233]
Внешние ссылки [ править ]
- Факты и цифры о мозге – Washington.edu
- Человеческий мозг – National Geographic
