Анатомия мозжечка
| Мозжечок | |
|---|---|
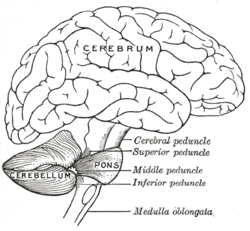 Рисунок человеческого мозга, показывающий мозжечок и мост. | |
 Вертикальное поперечное сечение мозжечка человека по средней линии, показывающее складчатый рисунок коры и внутренние структуры. | |
| Подробности | |
| Часть | Метэнцефалон |
| Артерия | СКА , АИКА , ПИКА |
| вена | Высший , низший |
| Идентификаторы | |
| НейроЛекс ID | бирнлекс_1489 |
| ТА98 | A14.1.07.001 |
| ТА2 | 5788 |
| Анатомические термины нейроанатомии | |
Анатомию мозжечка можно рассматривать на трех уровнях. На уровне общей анатомии мозжечок , под которым расположено белое вещество, несколько глубоких ядер , состоит из плотно свернутого и смятого слоя коры встроенных в белое вещество, и заполненный жидкостью желудочек посередине. [1] На промежуточном уровне мозжечок и его вспомогательные структуры могут быть разбиты на несколько сотен или тысяч независимо функционирующих модулей или отсеков, известных как микрозоны . [1] На микроскопическом уровне каждый модуль состоит из одного и того же небольшого набора нейронных элементов, расположенных с весьма стереотипной геометрией. [2]
Общая анатомия
[ редактировать ]Мозжечок расположен у основания мозга, над ним располагается большая масса коры головного мозга часть ствола мозга, называемая мостом , а перед ним — . Он отделен от вышележащего головного мозга слоем прочной твердой мозговой оболочки ; все его связи с другими частями мозга проходят через мост. Анатомы относят мозжечок к части промежуточного мозга , в который входит также мост; метэнцефалон, в свою очередь, является верхней частью ромбенцефалона или «заднего мозга». Как и кора головного мозга, мозжечок разделен на два полушария; он также содержит узкую срединную зону, называемую червем . Ряд крупных складок традиционно используется для разделения общей структуры на десять более мелких долек .
Из-за большого количества крошечных гранулярных клеток мозжечок содержит больше нейронов , чем весь остальной мозг вместе взятый, но он занимает лишь 10% от общего объема мозга. [3] Мозжечок получает около 200 миллионов входных волокон; напротив, зрительный нерв состоит всего из миллиона волокон.
Необычный внешний вид мозжечка скрывает тот факт, что основная часть структуры состоит из очень плотно сложенного слоя серого вещества — коры мозжечка . Было подсчитано, что если бы кору мозжечка человека можно было полностью развернуть, то образовался бы слой нервной ткани длиной около 1 метра и шириной 10 сантиметров — общая площадь поверхности 500–1000 квадратных см. [4] все упаковано в объем 100-150 куб.см. [5] [6] Под серым веществом коры находится белое вещество , состоящее в основном из миелинизированных нервных волокон, идущих к коре и от нее. В белом веществе мозжечка, которое иногда называют древом жизни (Древом жизни) из-за его разветвленного древовидного вида, находятся четыре глубоких ядра мозжечка .
Мозжечок можно разделить по трем различным критериям: грубоанатомическому, филогенетическому и функциональному.
Общие анатомические подразделения
[ редактировать ]При общем осмотре в мозжечке можно различить три доли: флокулонодулярную долю , переднюю долю (ростральную по отношению к «первичной щели») и заднюю долю (дорсальную по отношению к «первичной щели»). Последние два можно разделить на срединный червь мозжечка и боковые полушария мозжечка.
 |  |
Филогенетические и функциональные подразделения
[ редактировать ]Мозжечок также можно разделить на три части на основе как филогенетических критериев (эволюционный возраст каждой части), так и функциональных критериев (входящие и исходящие связи, которые имеет каждая часть, и роль, которую играет в нормальной функции мозжечка). От филогенетически древнейшей до новейшей можно выделить три части:
| Функциональное наименование ( филогенетическое наименование) | Анатомические части | Роль |
| Вестибулоцеребеллум (архомозжечок) | Флоккулонодулярная доля (и непосредственно прилегающий червь) | Вестибулоцеребеллум регулирует равновесие и движения глаз. Он получает вестибулярную информацию как от полукружных каналов , так и от вестибулярных ядер и отправляет волокна обратно к медиальным и латеральным вестибулярным ядрам. Он также получает визуальную информацию от верхних холмов и от зрительной коры (последняя через ядра моста , образуя кортико-понто-мозжечковый путь). Поражения вестибулоцеребеллума вызывают нарушения равновесия и походки . Есть еще одна небольшая область, известная как двустворчатая долька . |
| Спиноцеребеллум (Палеоцеребеллум) | Червь и промежуточные части полушарий («паравермис») | Спинноцеребеллум регулирует движения тела и конечностей. Он получает проприоцептивные сигналы от дорсальных столбов спинного мозга (включая спинно-мозжечковый тракт ) и тройничного нерва , а также от зрительной и слуховой систем. Он посылает волокна к глубоким ядрам мозжечка (включая фастигиальное ядро ), которые, в свою очередь, проецируются как в кору головного мозга (через средний мозг и таламус ), так и в ствол мозга (через ретикулярную формацию в мосту и вестибулярные ядра в продолговатом мозге ), таким образом обеспечение модуляции нисходящих двигательных систем. Спинно-мозжечок содержит сенсорные карты, поскольку получает данные о положении различных частей тела в пространстве: в частности, червь получает волокна от туловища и проксимальных отделов конечностей, а промежуточные отделы полушарий получают волокна от дистальных отделов конечностей. . Спинноцеребеллум способен вырабатывать проприоцептивные сигналы, чтобы предвидеть будущее положение части тела во время движения в режиме «упреждения». |
| Цереброцеребеллум (Neocerebellum, Pontocerebellum) | Боковые части полушарий | Неоцеребеллум участвует в планировании движения и оценивает сенсорную информацию для действий. Он получает входные данные исключительно от коры головного мозга (особенно теменной доли ) через ядра моста (в мосту , образующие кортико-понто-мозжечковые пути) и зубчатое ядро (в мозжечке ) и отправляет волокна преимущественно в вентролатеральный таламус (в в свою очередь связан с двигательными областями премоторной коры и первичной двигательной областью коры головного мозга) и с красным ядром (в свою очередь связанным с нижним оливковым ядром , которое связано с полушариями мозжечка). Неоцеребеллум участвует в планировании предстоящего движения. [7] и имеет чисто когнитивные функции. |
Многое из того, что известно о функциях мозжечка, связано с тщательным документированием эффектов очаговых поражений у пациентов, перенесших травмы или заболевания, или в результате исследований поражений на животных.
Клеточная анатомия
[ редактировать ]Как более подробно объясняется в разделе «Функции» , мозжечок отличается от большинства других областей мозга тем, что поток нервных сигналов через него почти полностью однонаправлен: между его нейрональными элементами практически нет обратных связей. Таким образом, наиболее логичный способ описания клеточной структуры — начать с входов и проследить последовательность соединений до выходов.
Глубокие ядра
[ редактировать ]Четыре глубоких ядра мозжечка — это зубчатое , эмболиформное , шаровидное и фастигиевое ядра, и они действуют как основные центры связи, отправляя и получая информацию в определенные части мозга и из них. Кроме того, эти ядра получают как тормозящие, так и возбуждающие сигналы из других частей мозга, которые, в свою очередь, влияют на исходящие сигналы ядер. [8] (Шарообразное и эмболиформное ядра составляют вставочное ядро ).
Кортикальные слои
[ редактировать ]

Цитоархитектура организация ) ( клеточная мозжечка очень однородна, соединения организованы в грубый трехмерный массив перпендикулярных элементов цепи . Такое организационное единообразие делает нервные схемы относительно простыми для изучения.
Кора мозжечка состоит из трех слоев; от внешнего слоя к внутреннему — это молекулярный слой, слой Пуркинье и зернистый слой. Функция коры мозжечка по существу заключается в модуляции информации, проходящей через глубокие ядра. Микросхема мозжечка схематически представлена на рис. 5. Мшистые и лазающие волокна переносят сенсомоторную информацию в глубокие ядра, которые, в свою очередь, передают ее различным премоторным областям, регулируя тем самым усиление и время двигательных действий. Мшистые и лазающие волокна также передают эту информацию в кору мозжечка, которая выполняет различные вычисления, что приводит к регуляции активности клеток Пуркинье. Нейроны Пуркинье возвращаются в глубокие ядра через мощный тормозной синапс . Этот синапс регулирует степень, в которой мшистые и лазающие волокна активируют глубокие ядра и, таким образом, контролируют конечный эффект мозжечка на двигательную функцию. Было показано, что синаптическая сила почти каждого синапса в коре мозжечка подвергается синаптическая пластичность . Это позволяет схемам коры мозжечка постоянно корректировать и точно настраивать работу мозжечка, формируя основу некоторых типов двигательного обучения и координации. Каждый слой коры мозжечка содержит различные типы клеток, составляющие эту схему.
Молекулярный слой
[ редактировать ]Этот внешний слой коры мозжечка содержит два типа тормозных интернейронов : звездчатые и корзинчатые клетки . Он также содержит дендритные ветви нейронов Пуркинье и параллельные волокна из гранулярных клеток. И звездчатые, и корзинчатые клетки образуют ГАМКергические синапсы на дендритах клеток Пуркинье.
Слой Пуркинье
[ редактировать ]Средний слой содержит только один тип клеточного тела — большую клетку Пуркинье . Клетки Пуркинье являются первичными интегративными нейронами коры мозжечка и обеспечивают его единственный выходной сигнал. Дендриты клеток Пуркинье представляют собой большие ветви с сотнями шиповатых ветвей, доходящих до молекулярного слоя (рис. 6). Эти дендритные беседки плоские — почти все они лежат в плоскостях — с соседними беседками Пуркинье в параллельных плоскостях. Каждое параллельное волокно из ячеек-зерен проходит через эти оправки ортогонально , как проволока, проходящая через множество слоев. Нейроны Пуркинье являются ГАМКергическими (то есть имеют тормозные синапсы) с нейронами глубоких мозжечковых и вестибулярных ядер ствола мозга. Каждая клетка Пуркинье получает возбуждающий сигнал от 100 000 до 200 000 параллельных волокон. Говорят, что параллельные волокна ответственны за простой (все или ничего, амплитуда инвариантна) выброс клеток Пуркинье.
Клетки Пуркинье также получают информацию от нижнего оливкового ядра через лазающие волокна . Хорошей мнемоникой для этого взаимодействия является фраза «залезть на другую оливковое дерево», учитывая, что волокна для лазания берут начало от контралатеральной нижней оливы. В отличие от более чем 100 000 входных сигналов от параллельных волокон, каждая клетка Пуркинье получает входные данные ровно от одного восходящего волокна; но это единственное волокно «взбирается» на дендриты клетки Пуркинье, обвивая их и образуя при этом большое количество синапсов. Чистый входной сигнал настолько силен, что одиночный потенциал действия восходящего волокна способен вызвать «сложный спайк» в клетке Пуркинье: вспышку из нескольких спайков подряд с уменьшающейся амплитудой. [9] за которым следует пауза, во время которой подавляются простые всплески.
Прямо под слоем Пуркинье находятся клетки Лугаро, чьи очень длинные дендриты проходят вдоль границы между слоем Пуркинье и зернистым слоем.
Зернистый слой
[ редактировать ]Самый внутренний слой содержит клеточные тела трех типов клеток: многочисленные и крошечные клетки-зерна , немного более крупные униполярные клетки-щеточки. [10] и гораздо более крупные клетки Гольджи . Мшистые волокна входят в зернистый слой из основного места своего происхождения — ядер моста. Эти волокна образуют возбуждающие синапсы с клетками-зернами и клетками глубоких ядер мозжечка. Клетки-зерна посылают свои Т-образные аксоны, известные как параллельные волокна , вверх в поверхностный молекулярный слой, где они образуют сотни тысяч синапсов с дендритами клеток Пуркинье . Человеческий мозжечок содержит порядка 60–80 миллиардов гранулярных клеток, что делает этот тип клеток безусловно самым многочисленным нейроном в головном мозге (примерно 70% всех нейронов головного и спинного мозга вместе взятых). Клетки Гольджи обеспечивают тормозящую обратную связь с гранулярными клетками, образуя с ними синапс и проецируя аксон в молекулярный слой.
Связь с корой головного мозга
[ редактировать ]Потенциалы локального поля неокортекса и мозжечка когерентно колеблются с частотой (6–40 Гц) у бодрствующих животных. [11] Похоже, что они находятся под контролем коры головного мозга. [12] Этот результат будет опосредован путем от нейронов слоев 5/6 в неокортексе через этот проект либо к мосту, либо к нижней оливе. Если через мост он будет идти к мшистым волокнам, которые синапсируют с гранулами, а нейронам Гольджи с клетками-зернами, то они будут нацелены на нейроны Пуркинье через их возбуждающие параллельные волокна. Если это нижняя олива, то он пойдет через входы возбуждающих лазающих волокон к нейронам Пуркинье. [12] Они возвращают этот выходной сигнал обратно в кору головного мозга через вентролатеральный таламус, завершая петлю.
Кортикопонтоцеребеллярный путь — самый крупный путь, связанный с мозжечком. Возникая в коре головного мозга, эти волокна сначала оканчиваются ипсилатерально в ядрах моста . Затем волокна перекрещиваются и образуют среднюю ножку мозжечка, оканчивающуюся в коре мозжечка в виде мшистых волокон. Этот путь передает сигналы, которые информируют мозжечок о текущем и предстоящем движении. Это помогает постоянной корректировке двигательной активности. [13]
Начало движения передается в мозжечок по кортикоретуло-мозжечковому пути. Эти синапсы ипсилатерально образуют ретикулярную формацию , затем через нижнюю и среднюю ножки в червь мозжечка . [13]
Моторная кора и соматосенсорная кора проецируются на ипсилатеральные нижние и добавочные ядра оливы, образуя затем оливо-мозжечковый тракт . Кортико-оливарные волокна образуют билатеральные синапсы в нижнем оливковом ядре . Порядок сохраняется в проекциях оливо-мозжечкового тракта на «карты тела» в контралатеральной коре мозжечка. В экспериментах на животных в условиях покоя группы оливарных нейронов разряжаются синхронно с частотой от 5 до 10 Гц (импульсов/с). В коре мозжечка ответ клеток Пуркинье принимает форму сложных спайков. [14]
Мозжечок посылает свои проекции обратно в кору головного мозга через мозжечково-таламический тракт .
Латеральное расширение мозжечка, или неоцеребеллум, может быть связано с когнитивными функциями и анатомически связано с латеральной префронтальной корой . Наибольшую активность он проявляет во время речи с односторонним преобладанием, что соответствует возможной связи (через таламус) с двигательной речевой областью. [14]
Когда поражения возникают в ассоциативных областях, связанных с мозжечком кортикопонтоцеребеллярными волокнами, может возникнуть когнитивно-аффективный синдром. Это приводит к когнитивным дефектам в виде снижения способности рассуждать, невнимательности, грамматических ошибок в речи, плохого пространственного чувства и неоднородной потери памяти. [14]
Кровоснабжение
[ редактировать ]

К мозжечку кровоснабжают три артерии (рис. 7): верхняя мозжечковая артерия (SCA), передняя нижняя мозжечковая артерия (AICA) и задняя нижняя мозжечковая артерия (PICA).
SCA ответвляется от латеральной части основной артерии, чуть ниже ее бифуркации в заднюю мозговую артерию . Здесь он огибает сзади мост (к которому он также снабжает кровью), прежде чем достичь мозжечка. SCA снабжает кровью большую часть коры мозжечка, ядра мозжечка и верхние ножки мозжечка. [15]
AICA ответвляется от латеральной части основной артерии, чуть выше места соединения позвоночных артерий. От своего начала он разветвляется вдоль нижней части моста у мостомозжечкового угла, прежде чем достичь мозжечка. Эта артерия снабжает кровью переднюю часть нижнего мозжечка, среднюю ножку мозжечка, а также лицевой ( CN VII) и преддверно-улитковый нервы (CN VIII). Обструкция AICA может вызвать парез , паралич и потерю чувствительности лица; это также может привести к ухудшению слуха . Более того, это может вызвать инфаркт мостомозжечкового угла. Это может привести к гиперакузии (дисфункции стременной мышцы , иннервируемой CN VII ) и головокружению вестибулярного полукружного канала, (неправильная интерпретация ускорения эндолимфы вызванного изменением CN VIII ).
PICA ответвляется от латеральной части позвоночных артерий чуть ниже места их соединения с основной артерией. Прежде чем достичь нижней поверхности мозжечка, ЗНСА посылает ветви в продолговатый мозг, снабжая кровью несколько ядер черепных нервов . В мозжечке PICA снабжает кровью заднюю нижнюю часть мозжечка, нижнюю ножку мозжечка, двойное ядро , блуждающее двигательное ядро, спинномозговое тройничное ядро, одиночное ядро и вестибулокохлеарные ядра.
Вариации среди позвоночных
[ редактировать ]У разных видов позвоночных существуют значительные различия в размерах и форме мозжечка. Обычно он самый крупный у хрящевых и костных рыб , птиц и млекопитающих, но несколько мельче у рептилий. Большие парные и извитые доли, обнаруженные у людей, типичны для млекопитающих, но в других группах мозжечок обычно представляет собой одну срединную долю и либо гладкий, либо лишь слегка бороздчатый. У млекопитающих неоцеребеллум составляет большую часть мозжечка по массе, но у других позвоночных это обычно спиноцеребеллум. [16]
У земноводных , миног и миксин мозжечок развит слабо; в последних двух группах он едва отличим от ствола мозга. Хотя в этих группах присутствует спино-мозжечок, первичными структурами являются небольшие парные ядра, соответствующие вестибуло-мозжечку. [16]
Цветоносы
[ редактировать ]Мозжечок следует общей схеме групп из трех, обнаруженной в анатомии. [17] с тремя основными входными и выходными ножками мозжечка (пучками волокон). Это верхняя (brachium conjunctivum), средняя (brachium pontis) и нижняя (рестиформное и юктарестиформное тела ) ножки мозжечка.
| Цветонос | Описание |
| Начальство | есть некоторые афферентные волокна Хотя из переднего спинно-мозжечкового пути , которые передаются к передней доле мозжечка через эту ножку, большинство волокон являются эфферентными. Таким образом, верхняя ножка мозжечка является основным выходным путем мозжечка. Большинство эфферентных волокон берут начало в зубчатом ядре , которое, в свою очередь, проецируется на различные среднего мозга, структуры включая красное ядро , вентральное латеральное/вентральное переднее ядро таламуса и продолговатый мозг . Дентаторуброталамокортикальный ) (зубчатое ядро > красное ядро > таламус > премоторная кора и мозжечково-таламокортикальный (мозжечок > таламус > премоторная кора) пути являются двумя основными путями, которые проходят через эту ножку и играют важную роль в планировании движений . |
| Середина | Он полностью состоит из афферентных волокон, берущих начало в ядрах моста и являющихся частью массивного кортико-понто-мозжечкового тракта (кора головного мозга > мост > мозжечок). Эти волокна спускаются от сенсорных и моторных областей неокортекса головного мозга и делают среднюю ножку мозжечка самой крупной из трех ножок мозжечка. |
| Низший | Он несет множество типов входных и выходных волокон, которые в основном связаны с интеграцией проприоцептивной сенсорной информации с двигательными вестибулярными функциями, такими как поддержание баланса и позы. Проприоцептивная информация от тела передается в мозжечок через дорсальный спинно-мозжечковый тракт . Этот тракт проходит через нижнюю ножку мозжечка и синапсы в палеоцеребеллуме. Вестибулярная информация проецируется на архицеребеллум. Лазящие волокна нижней оливы проходят через нижнюю ножку мозжечка. Эта ножка также передает информацию непосредственно от клеток Пуркинье к вестибулярным ядрам дорсального ствола мозга, расположенным на стыке моста и продолговатого мозга . |
Существует три источника входных сигналов в мозжечок: две категории, состоящие из мшистых и лазающих волокон соответственно. Мшистые волокна могут происходить из ядер моста, которые представляют собой скопления нейронов, расположенных в мосту, которые несут информацию из контрлатеральной коры головного мозга. Они также могут возникать в спинно-мозжечковом тракте, начало которого находится в ипсилатеральном отделе спинного мозга . Большая часть выходного сигнала мозжечка первоначально попадает в синапсы в глубокие ядра мозжечка, а затем выходит через три ножки. Наиболее заметным исключением является прямое ингибирование вестибулярных ядер клетками Пуркинье.
Разработка
[ редактировать ]На ранних стадиях эмбрионального развития мозг начинает формироваться в трех отдельных сегментах: переднем мозге , среднем мозге и ромбовидном мозге . Ромбовидный мозг — самый каудальный (к хвосту) сегмент эмбрионального мозга; именно из этого сегмента развивается мозжечок. Вдоль эмбрионального ромбэнцефалического сегмента развиваются восемь вздутий, называемых ромбомерами . Мозжечок возникает из двух ромбомеров, расположенных в крыльной пластинке нервной трубки , структуры, которая в конечном итоге формирует головной и спинной мозг. Конкретными ромбомерами, из которых формируется мозжечок, являются ромбомер 1 (Rh.1) каудально (около хвоста) и «перешеек» рострально (около передней части). [18]
Считается, что две основные области дают начало нейронам, составляющим мозжечок. Первая область — желудочковая зона крыши четвертого желудочка . Эта область производит клетки Пуркинье и глубокие ядерные нейроны мозжечка. Эти клетки являются первичными выходными нейронами коры мозжечка и мозжечка. Вторая зародышевая зона (место рождения клеток) известна как ромбическая губа, затем к 27-й неделе эмбрионального развития человека нейроны перемещаются во внешний зернистый слой . Этот слой клеток, расположенный на внешней стороне мозжечка, производит гранулярные нейроны. Нейроны-гранулы мигрируют из этого внешнего слоя, чтобы сформировать внутренний слой, известный как внутренний слой гранул. [19] В зрелом мозжечке внешний зернистый слой перестает существовать, и во внутреннем зернистом слое остаются только гранулярные клетки. мозжечка Белое вещество может быть третьей зародышевой зоной мозжечка; однако его функция как зародышевой зоны противоречива.
Дополнительные изображения
[ редактировать ]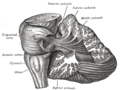 Вскрытие, показывающее проекционные волокна мозжечка.
Вскрытие, показывающее проекционные волокна мозжечка.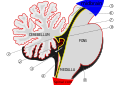 Схема крыши четвертого желудочка. Стрела находится в отверстии Мадженди.
Схема крыши четвертого желудочка. Стрела находится в отверстии Мадженди. Среднесагиттальный вид человеческого мозга
Среднесагиттальный вид человеческого мозга Вид человеческого мозжечка спереди, с цифрами, обозначающими основные ориентиры.
Вид человеческого мозжечка спереди, с цифрами, обозначающими основные ориентиры.




Ссылки
[ редактировать ]- ^ Перейти обратно: а б Книерим, Джеймс. «Глава 5: Мозжечок» . Neuroscience Online: Электронный учебник по нейронаукам .
- ^ Фриде, Рейнхард Л. (1 марта 1973 г.). «Датирование развития мозжечка человека». Акта Нейропатологика . 23 (1): 48–58. дои : 10.1007/BF00689004 . ISSN 1432-0533 . ПМИД 4698523 . S2CID 5387374 .
- ^ Мозг сверху вниз
- ^ Лю, Вэньцзяо; Ву, Йе; Хюинь, Кхой Мин; Ахмад, Сахар; Да, Пью-Тиан (2024). «Мультимодальный субмиллиметровый МРТ-атлас мозжечка человека» . Научные отчеты . 14 (1): 5622. doi : 10.1038/s41598-024-55412-y . ISSN 2045-2322 . ПМЦ 10920891 . ПМИД 38453991 .
- ^ Эдвардс Ч.Р., Ньюман С., Бисмарк А. и др. (2008). «Объем мозжечка и состояние моргания при шизофрении» . Психиатрия Рез . 162 (3): 185–194. doi : 10.1016/j.pscychresns.2007.06.001 . ПМК 2366060 . ПМИД 18222655 .
- ^ Хатчинсон С., Ли Л.Х., Гааб Н., Шлауг Г. (2003). «Мозжечковый том музыкантов» . Цереб. Кортекс . 13 (9): 943–9. дои : 10.1093/cercor/13.9.943 . ПМИД 12902393 .
- ^ Кингсли, RE (2000). Краткий текст по неврологии (2-е изд.). Липпинкотт Уильямс и Уилкинс. ISBN 0-683-30460-7 .
- ^ Хартинг, Дж. К. «Глобальный мозжечок '97» . Университета Висконсина Медицинская школа .
- ^ Хойссер, Михаэль; Кларк, Беверли А.; Дэви, Дженни Т. (23 июля 2008 г.). «Происхождение сложного спайка в клетках Пуркинье мозжечка» . Журнал неврологии . 28 (30): 7599–7609. doi : 10.1523/JNEUROSCI.0559-08.2008 . ISSN 0270-6474 . ПМК 2730632 . ПМИД 18650337 .
- ^ Кинни Г.А., Оверстрит Л.С., Слейтер Н.Т. (сентябрь 1997 г.). «Длительное физиологическое удержание глутамата в синаптической щели униполярных клеток щетки мозжечка» (PDF) . J Нейрофизиология . 78 (3): 1320–33. дои : 10.1152/jn.1997.78.3.1320 . ПМИД 9310423 .
- ^ Сотеропулос Д.С., Бейкер С.Н. (2006). «Кортико-мозжечковая когерентность во время точного захвата у обезьяны» . J Нейрофизиология . 95 (2): 1194–206. дои : 10.1152/jn.00935.2005 . ПМИД 16424458 .
- ^ Перейти обратно: а б Рос Х., Сачдев Р.Н., Ю Ю, Сестан Н., Маккормик Д.А. (2009). «Неокортикальные сети охватывают нейронные цепи коры мозжечка» . Журнал неврологии . 29 (33): 10309–20. doi : 10.1523/JNEUROSCI.2327-09.2009 . ПМК 3137973 . ПМИД 19692605 .
- ^ Перейти обратно: а б Гартнер, Лесли П.; Патестас, Мария А. (2009). Учебник нейроанатомии . Уайли-Блэквелл. п. 464. ИСБН 9781405103404 .
- ^ Перейти обратно: а б с Мтуи, Эстомих; Грюнер, Грегори; Докери, Питер (2016). Клиническая нейроанатомия и неврология Фицджеральда (7-е изд.). Эльзевир. стр. 243–252.
- ^ Грей, Генри; Льюис, Уоррен Хармон (1918). Анатомия человеческого тела (20-е изд.). Филадельфия: Леа и Фебигер.
- ^ Перейти обратно: а б Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. п. 531. ИСБН 0-03-910284-Х .
- ^ «Список трех» . www.meddean.luc.edu .
- ^ Мюллер Ф., О'Рахилли Р. (1990). «Человеческий мозг на стадиях 21–23, с особым упором на корковую пластинку головного мозга и развитие мозжечка». Анат Эмбриол (Берл) . 182 (4): 375–400. дои : 10.1007/BF02433497 . ПМИД 2252222 . S2CID 33485509 .
- ^ Смейн, Ричард Дж .; Голдовиц, Дэн (май 1989 г.). «Развитие и гибель клеток внешнего зернистого слоя в мозжечке мыши-ткача: количественное исследование» . Журнал неврологии . 9 (5): 1608–20. doi : 10.1523/JNEUROSCI.09-05-01608.1989 . ПМК 6569844 . ПМИД 2723742 .