клетка Гольджи
| клетка Гольджи | |
|---|---|
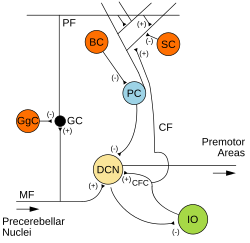 Микросхема мозжечка. Возбуждающие синапсы обозначаются знаком (+), тормозные – знаком (-). МФ: Мшистое волокно . DCN: Глубокие ядра мозжечка . Я: Плохие оливки . CF: Восхождение по волокну . GC: Гранульная клетка . PF: Параллельное волокно . ПК: клетка Пуркинье . GgC: клетка Гольджи. СК: Звездчатая клетка . БК: Ячейка-корзина . | |
| Подробности | |
| Расположение | Зернистый мозжечка слой |
| Идентификаторы | |
| НейроЛекс ID | нифекст_129 |
| Анатомические термины нейроанатомии | |
В нейробиологии обнаруженными клетки Гольджи являются наиболее распространенными интернейронами , в зернистом слое мозжечка тормозными . [1] Клетки Гольджи можно найти в зернистом слое на различных слоях. [2] Клетка Гольджи необходима для контроля активности зернистого слоя. [3] Впервые они были идентифицированы как тормозные в 1964 году. [4] Это также был первый пример тормозной обратной связи , в которой тормозной интернейрон был идентифицирован анатомически.Клетки Гольджи производят широкое латеральное торможение, выходящее за пределы афферентного синаптического поля, и ингибируют гранулярные клетки посредством тормозных петель прямой и обратной связи. [4] Эти клетки образуют синапсы на дендритах гранулярных клеток и униполярных щеточных клетках . Они получают возбуждающий сигнал от мшистых волокон , также образующих синапсы на гранулярных клетках , и параллельных волокон , которые представляют собой длинные аксоны гранулярных клеток. Таким образом, эта схема позволяет осуществлять прямое и обратное ингибирование гранулярных клеток.
Соединения
[ редактировать ]Сеть мозжечка содержит большое количество связей между клетками Гольджи. [5] Основной синапс, образуемый этими клетками, представляет собой синапс на мшистое волокно – возбуждающий синапс гранулярных клеток в клубочке . Клубочек состоит из мшистого окончания волокна, дендритов гранулярных клеток и окончания Гольджи и окружен глиальной оболочкой. [3] Клетка Гольджи действует путем изменения мшисто-волокнистого синапса клеток-зерен. По имеющимся данным, гранулярные клетки в основном соединяются с клеткой Гольджи через параллельные волокна, при этом определенную роль могут играть и синапсы. Было показано, что лазящие волокна соединяются с клетками Гольджи, повторно входя в более высокий зернистый слой через тонкие коллатеральные ветви, которые достигают немного ниже клеток Пуркинье. [6] Три различных типа тормозных интернейронов — корзинчатые и звездчатые клетки, которые находятся в молекулярном слое, и клетки Гольджи, которые находятся в зернистом слое, — запускаются параллельными волокнами и контролируют активность клеток Пуркинье. [7]
В диапазоне тета-частот клетки Гольджи демонстрируют кардиостимуляцию, резонанс, сброс фазы и обратное возбуждение. Эти характеристики, вероятно, влияют на их поведение. In vivo демонстрирует беспорядочное, спонтанное биение, регулируемое сенсорными сигналами, и внезапные тихие паузы между взрывными реакциями для акцентирования стимула. Более того, взаимодействие клеток Гольджи в сети дает представление о том, как эти нейроны могут контролировать пространственно-временную организацию активности мозжечка. Оказывается, клетки Гольджи могут влиять как на временную динамику, так и на географическое распределение информации, передаваемой по сети мозжечка. Клетки Гольджи также контролируют выработку синапсом мшисто-волокнистых клеток долгосрочной синаптической пластичности. Таким образом, начинает обретать форму идея о том, что клетки Гольджи играют решающую роль в контроле активности сети гранулярных слоев, что имеет важное значение для вычислений мозжечка. [8]
Глутаматергические стимулы являются основными возбуждающими факторами для клеток Гольджи. Текущие исследования показывают, что NMDA [9] рецепторы и АМРА [10] рецепторы участвуют в реле мшистых волокон-клеток Гольджи. [11] Функции цепи клеток Гольджи, по-видимому, также регулируются метаботропными рецепторами глутамата. Клетки Гольджи обладают рецепторами mGluR2. [12] и когда эти рецепторы активированы, внутренний выпрямительный ток К усиливается, способствуя подавлению активности клеток Гольджи после периода интенсивной передачи между гранулярными клетками и клетками Гольджи. [13] Этот mGluR2-зависимый процесс может облегчить прохождение длительных всплесков по мшистым волокнистым клеточным маршрутам. [14]
Нейромедиаторы
[ редактировать ]Клетки Гольджи преимущественно используют ГАМК и глицин в качестве нейромедиаторов, хотя в зависимости от их мишени одна клетка Гольджи избирательно облегчает ГАМКергическую или глицинергическую передачу. Когда клетки Гольджи активированы, гранулярные клетки и UBC проявляют ГАМКергические и глицинергические токи соответственно. Это происходит главным образом потому, что клетки Гольджи высвобождают ГАМК и глицин в каждом отдельном бутоне, что является результатом экспрессии постсинаптических рецепторов, специфичных для данного типа клеток, и/или их доставки в синапс. [15] На основе электронной микроскопии [16] и электрофизиологические данные, [17] считалось, что интернейроны молекулярного слоя (звездчатые и корзинчатые клетки) образуют ГАМКергические синапсы на дендритах клеток Гольджи. Однако более поздние исследования ставят под сомнение существование этих функциональных синапсов. [18] Вместо этого локальные интернейроны обеспечивают лишь небольшую часть тормозных сигналов клеткам Гольджи в молекулярном слое, и большая часть этих входных сигналов высвобождает только ГАМК. Скорее, клетки Гольджи иннервируются ГАМКергическими клетками в DCN, что указывает на то, что клетки Гольджи получают большой входной сигнал торможения по обратной связи от глубоких ядер мозжечка для модуляции локальных тормозных сетей. [19] Базальный уровень ГАМК вызывает постсинаптическую утечку проводимости путем тонической активации альфа-6-содержащих рецепторов ГАМК-А на гранулярных клетках. [1] [2] [10] Эти рецепторы с высоким сродством расположены как синаптически, так и экстрасинаптически на гранулярной клетке. Синаптические рецепторы опосредуют фазовое сокращение длительностью около 20–30 мс, тогда как экстрасинаптические рецепторы опосредуют тоническое торможение длительностью около 200 мс и активируются за счет перетекания синапса. [9]
Кроме того, ГАМК действует на рецепторы ГАМК-В, которые расположены пресинаптически на окончаниях мшистых волокон. Они ингибируют вызванные мшистыми волокнами EPSC гранулярных клеток в зависимости от температуры и частоты. При высокой частоте импульсации мха (10 Гц) действие ГАМК на пресинаптические рецепторы ГАМК-В на вызванных ВПСК не наблюдается. Однако при низком (1 Гц) воздействии ГАМК действительно оказывает влияние на ВПСК, опосредованное через эти пресинаптические рецепторы ГАМК-В.
Клетки Гольджи необходимы для сложной координации движений, поскольку это исследование показывает, что «Аблация мозжечковых клеток Гольджи нарушает синаптическую интеграцию, включающую ингибирование ГАМК и активацию рецептора NMDA в координации движений», проведенную Ватанабэ Д., Инокавой Х. и др. Более того, такие составные движения зависят от синаптической интеграции, которая возникает в результате активации рецептора NMDA гранулярных клеток и ГАМК-опосредованного ингибирования. [7] В конце концов, клетки Гольджи используют расширенное аксональное сплетение, чтобы блокировать широкие поля гранулярных клеток. Фундаментальные вопросы о том, лежит ли дендритный процессинг в основе предсказанной теорией пластичности, зависящей от времени спайков (STDP), и как синаптические входы управляют генерацией спайков клеток Гольджи, остаются без ответа. [20] Интересно отметить, что дендриты экспрессируют разнообразный набор ионных каналов Ca, Na и K. [21] это может влиять на дендритные вычисления, в то время как синапсы мшистое волокно-клетка Гольджи экспрессируют каналы NMDA, которые необходимы для синаптической пластичности. [22] Прогнозирование потенциальных взаимосвязей между этими многочисленными активными функциями является сложной задачей и требует тщательного вычислительного исследования синаптической интеграции и электрогенной архитектуры нейрона. [23]
Гольджи тип I
[ редактировать ]Клеточные тела нейронов Гольджи типа I имеют средние и крупные размеры. [24] Нейрон Гольджи I типа , имеет длинный аксон который начинается в сером веществе центральной нервной системы и может простираться оттуда.Их клеточные тела были в основном многополярными, но иногда они могли иметь треугольную форму и не иметь каких-либо придатков или шипов. Они имели от трех до десяти основных дендритов. На этих дендритах не было придатков и шипиков. Дендриты клеток имели обильное ветвление. [25]
Эти нейроны имеют тафтинговый и лучистый паттерн ветвления в своих дендритах, по сравнению с тафтинговым паттерном, расходящийся паттерн ветвления был более распространенным. [26] В этих клетках обычно присутствует плотность дендритных деревьев, но количество и диаметр первичных дендритов крайне неравномерны. Вне тела клетки видны от трех до одиннадцати дендритов. Прежде чем расщепиться на третичные ветви, он быстро дает начало более тонким вторичным дендритам. [27]
Он также известен как проекционный нейрон. К ним относятся нейроны, образующие периферические нервы и длинные пути головного и спинного мозга. [11] с размерами сомат обычно от 20 до 40 мкм. [27] Нейроны Гольджи II, напротив, определяются как имеющие короткие аксоны или вообще не имеющие аксонов. Это различие было введено нейроанатомом-новатором Камилло Гольджи на основании внешнего вида под микроскопом нейронов, окрашенных красителем Гольджи изобретенным им . Сантьяго Рамон-и-Кахаль предположил, что у более развитых животных больше нейронов Гольджи типа II по сравнению с нейронами Гольджи типа I. Эти нейроны Гольджи типа II имеют звездообразный вид и находятся в коре головного мозга, мозжечка и сетчатке. [28]
Гольджи тип II
[ редактировать ]Тела клеток этих нейронов были яйцевидными, сфероидальными или мультиполярными. [27] Нейрон Гольджи типа II либо не имеет аксона , либо имеет короткий аксон , который не дает ответвлений из серого вещества центральной нервной системы. [12] Дендриты Гольджи типа II имеют примерно симметричные синаптические связи и имеют бледный, асимметричный и часто массивный профиль, содержащий огромные плеоморфные везикулы. Синаптические окончания аксона Гольджи типа II могут напоминать дендритные окончания, однако многие аксональные окончания, по-видимому, имеют более узкие профили с меньшими и более плоскими пузырьками. [29] Их средний диаметр варьировал от 12 до 30 мкм, в среднем 22,2 мкм (5,8 ± n = 120). [27] По сравнению с нейронами Гольджи типа I, нейроны Гольджи типа II имеют большее соотношение ядра и цитоплазмы (N/C). [27] По сравнению с нейронами Гольджи типа I, дендриты этих нейронов имеют значительно меньше тафтинговых дендритов. Два из десяти основных дендритов выступали из тела клетки и давали небольшое количество ветвей. [27] Нейрон Гольджи типа II генерирует дендро-дендритные связи с основным нейроном в терминальных агрегатах, называемых синаптическими гнездами. Афферентные аксоны, идущие от слуховой коры и восходящие от задних холмиков, образуют синаптические связи с обоими видами нейронов. [30]
Клетки Гольджи типа II могут быть возбуждающими или тормозными интернейронами, или они могут быть и тем, и другим. Клетки Гольджи типа II функционируют как тормозные интернейроны, которые могут создавать паттерны реакции, которые делают первичные нейроны более чувствительными к началу стимулов и к временным изменениям афферентной входной информации. Клетки Гольджи типа II, являющиеся возбуждающими интернейронами, обладают способностью генерировать постепенные или непрерывные паттерны ответов, которые имеют тенденцию удлинять определенные последовательности сигналов. В каждом сценарии кортикальный анализ местоположения звуков и временных паттернов зависит от синаптических взаимодействий между клетками Гольджи типа II, чтобы определить пространственные и временные особенности кодирования стимулов. [31]
См. также
[ редактировать ]Список различных типов клеток в организме взрослого человека
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Брикли С.Г., Калл-Кэнди С.Г., Фаррант М. (декабрь 1996 г.). «Развитие тонической формы синаптического торможения в гранулярных клетках мозжечка крысы в результате стойкой активации ГАМКА-рецепторов» . Журнал физиологии . 497 (Часть 3) (Часть 3): 753–759. doi : 10.1113/jphysicalol.1996.sp021806 . ПМК 1160971 . ПМИД 9003560 .
- ^ Перейти обратно: а б Тиа С., Ван Дж. Ф., Котчабхакди Н., Вичини С. (июнь 1996 г.). «Изменения в развитии тормозных синаптических токов в гранулярных нейронах мозжечка: роль альфа-6-субъединицы рецептора ГАМК (А)» . Журнал неврологии . 16 (11): 3630–3640. doi : 10.1523/JNEUROSCI.16-11-03630.1996 . ПМК 6578841 . ПМИД 8642407 .
- ^ Перейти обратно: а б Якаб Р.Л., Хамори Дж. (1988). «Количественная морфология и синаптология клубочков мозжечка у крыс». Анатомия и эмбриология . 179 (1): 81–88. дои : 10.1007/BF00305102 . ПМИД 3213958 . S2CID 22651721 .
- ^ Перейти обратно: а б Экклс Дж., Ллинас Р., Сасаки К. (декабрь 1964 г.). «Торможение клеток Гольджи в коре мозжечка». Природа . 204 (4965): 1265–1266. Бибкод : 1964Natur.204.1265E . дои : 10.1038/2041265a0 . ПМИД 14254404 . S2CID 4187138 .
- ^ Палай С.Л., Чан-Палай В (1974). Кора мозжечка . дои : 10.1007/978-3-642-65581-4 . ISBN 978-3-642-65583-8 . S2CID 31651518 .
- ^ Шинода Ю., Сугихара И., Ву Х.С., Сугиучи Ю. (2000). «Вся траектория одиночных лазающих и мшистых волокон в ядрах и коре мозжечка». Модули мозжечка: Молекулы, морфология и функции . Прогресс в исследованиях мозга. Том. 124. Эльзевир. стр. 173–186. дои : 10.1016/s0079-6123(00)24015-6 . ISBN 978-0-444-50108-0 . ПМИД 10943124 .
- ^ Перейти обратно: а б Ватанабэ Д., Инокава Х., Хасимото К., Сузуки Н., Кано М., Сигэмото Р. и др. (октябрь 1998 г.). «Аблация клеток Гольджи мозжечка нарушает синаптическую интеграцию, включая ингибирование ГАМК и активацию рецептора NMDA в координации движений». Клетка . 95 (1): 17–27. дои : 10.1016/s0092-8674(00)81779-1 . hdl : 2433/181271 . ПМИД 9778244 .
- ^ Д'Анджело Э (июль 2008 г.). «Критическая роль клеток Гольджи в регуляции пространственно-временной интеграции и пластичности на входной стадии мозжечка» . Границы в неврологии . 2 (1): 35–46. дои : 10.3389/нейро.01.008.2008 . ПМК 2570065 . ПМИД 18982105 .
- ^ Перейти обратно: а б Нуссер З., Зигхарт В., Сомоджи П. (март 1998 г.). «Сегрегация различных ГАМК-рецепторов на синаптических и экстрасинаптических мембранах зернистых клеток мозжечка» (аннотация) . Журнал неврологии . 18 (5): 1693–1703. doi : 10.1523/JNEUROSCI.18-05-01693.1998 . ПМК 6792611 . ПМИД 9464994 .
- ^ Перейти обратно: а б Уолл М.Ю., Усович М.М. (март 1997 г.). «Развитие потенциал-зависимых и независимых спонтанных токов, опосредованных рецептором ГАМКАА, в гранулярных клетках постнатального мозжечка крыс». Европейский журнал неврологии . 9 (3): 533–548. дои : 10.1111/j.1460-9568.1997.tb01630.x . ПМИД 9104595 . S2CID 39603115 .
- ^ Перейти обратно: а б «Определение нейрона Гольджи типа I» . Словарь.com . 2008 год . Проверено 25 декабря 2008 г.
- ^ Перейти обратно: а б «Определение нейрона Гольджи типа II» . Словарь.com . Проверено 15 августа 2019 г.
- ^ Ватанабэ Д., Наканиси С. (август 2003 г.). «mGluR2 постсинаптически воспринимает входы гранулярных клеток в синапсах клеток Гольджи» . Нейрон . 39 (5): 821–829. дои : 10.1016/S0896-6273(03)00530-0 . ПМИД 12948448 .
- ^ Аренц А., Сильвер Р.А., Шефер А.Т., Маргри Т.В. (август 2008 г.). «Вклад одиночных синапсов в сенсорное представление in vivo» . Наука . 321 (5891): 977–980. Бибкод : 2008Sci...321..977A . дои : 10.1126/science.1158391 . ПМЦ 2771362 . ПМИД 18703744 .
- ^ Дюге ГП, Дюмулен А, Триллер А, Дьедонне С (июль 2005 г.). «Целево-зависимое использование совместно высвобождаемых тормозных передатчиков в центральных синапсах» . Журнал неврологии . 25 (28): 6490–6498. doi : 10.1523/jneurosci.1500-05.2005 . ПМЦ 6725433 . ПМИД 16014710 .
- ^ Чан-Палай В., Палай С.Л., Биллингс-Гальярди С.М. (ноябрь 1974 г.). «Клетки Мейнерта в зрительной коре приматов». Журнал нейроцитологии . 3 (5): 631–658. дои : 10.1007/bf01097628 . ПМИД 4142639 . S2CID 25165435 .
- ^ Дюмулен А, Триллер А, Дьедонне С (август 2001 г.). «Кинетика ИПСК в идентифицированных ГАМКергических и смешанных ГАМКергических и глицинергических синапсах на клетках Гольджи мозжечка» . Журнал неврологии . 21 (16): 6045–6057. doi : 10.1523/jneurosci.21-16-06045.2001 . ПМК 6763194 . ПМИД 11487628 .
- ^ Эйр, доктор медицинских наук, Нуссер З. (май 2016 г.). «Только незначительная часть тормозящих воздействий на мозжечковые клетки Гольджи происходит от локальных ГАМКергических клеток» . эНейро . 3 (2): ЭНЕВРО.0055–16.2016. дои : 10.1523/euro.0055-16.2016 . ПМЦ 4876488 . ПМИД 27257627 .
- ^ Анкри Л., Хуссон З., Пьетрайтис К., Провилл Р., Лена С., Яром Ю., Дьедонне С., Уусисаари М.Ю. (22 апреля 2015 г.). «Ответ автора: новый тормозной нуклеокортикальный контур контролирует активность клеток Гольджи мозжечка» . электронная жизнь . дои : 10.7554/elife.06262.015 .
- ^ Гарридо Х.А., Люке Н.Р., Толу С., Д'Анджело Э. (август 2016 г.). «Пластичность, зависящая от времени всплеска колебаний, позволяет распознавать множественные перекрывающиеся образы в тормозных промежуточных нейронных сетях». Международный журнал нейронных систем . 26 (5): 1650020. doi : 10.1142/S0129065716500209 . ПМИД 27079422 . S2CID 9779306 .
- ^ Рудольф С., Халл С., Регер В.Г. (ноябрь 2015 г.). «Активные дендриты и дифференциальное распределение кальциевых каналов обеспечивают функциональную компартментализацию клеток Гольджи» . Журнал неврологии . 35 (47): 15492–15504. doi : 10.1523/JNEUROSCI.3132-15.2015 . ПМЦ 4659820 . ПМИД 26609148 .
- ^ Чезана Э., Пьетрайтис К., Бидоре К., Исопе П., Д'Анджело Э., Дьедонне С., Форти Л. (июль 2013 г.). «Возбуждающие синапсы восходящих аксонов гранулярных клеток на клетках Гольджи реализуют мощную цепь обратной связи в зернистом слое мозжечка» . Журнал неврологии . 33 (30): 12430–12446. doi : 10.1523/JNEUROSCI.4897-11.2013 . ПМК 6618671 . ПМИД 23884948 .
- ^ Масоли С., Оттавиани А., Казали С., Д'Анджело Э. (декабрь 2020 г.). Кунц Х. (ред.). «Модели мозжечковых клеток Гольджи предсказывают дендритную обработку и механизмы синаптической пластичности» . PLOS Вычислительная биология . 16 (12): e1007937. Бибкод : 2020PLSCB..16E7937M . дои : 10.1371/journal.pcbi.1007937 . ПМЦ 7837495 . ПМИД 33378395 .
- ^ Аль-Хусейн С.М., Албостанджи С.А., Мустафа А.Г., Закут С. (сентябрь 2021 г.). «Типы нейрональных клеток в переднем вентральном таламическом ядре верблюда». Анатомическая запись . 304 (9): 2044–2049. дои : 10.1002/ar.24592 . hdl : 10576/17786 . ПМИД 33554482 .
- ^ Аль-Хусейн С.М., Албостанджи С.А., Мустафа А.Г., Закут С. (сентябрь 2021 г.). «Типы нейрональных клеток в переднем вентральном таламическом ядре верблюда». Анатомическая запись . 304 (9): 2044–2049. дои : 10.1002/ar.24592 . hdl : 10576/17786 . ПМИД 33554482 .
- ^ Аль-Хусейн С.М., Албостанджи С.А., Мустафа А.Г., Закут С. (сентябрь 2021 г.). «Типы нейрональных клеток в переднем вентральном таламическом ядре верблюда». Анатомическая запись . 304 (9): 2044–2049. дои : 10.1002/ar.24592 . hdl : 10576/17786 . ПМИД 33554482 .
- ^ Перейти обратно: а б с д и ж Аль-Хусейн Бани Хани С.М., Эль-Дваири QA, Батаине З.М., Аль-Хайдари М.С., Аль-Алами Дж. (май 2008 г.). «Нейроны Гольджи типа I и Гольджи типа II в вентральном переднем ядре таламуса взрослого человека: морфологические особенности и количественный анализ». Клеточная и молекулярная нейробиология . 28 (3): 411–416. дои : 10.1007/s10571-007-9248-8 . ПМИД 18264756 . S2CID 19916699 .
- ^ Даулинг Дж. Э. (2001). Нейроны и сети: введение в поведенческую нейронауку . Издательство Гарвардского университета. п. 46. ИСБН 978-0-674-00462-7 .
- ^ Морест Д.К. (июль 1975 г.). «Синаптические взаимоотношения клеток Гольджи типа II в медиальном коленчатом теле кошки». Журнал сравнительной неврологии . 162 (2): 157–193. дои : 10.1002/cne.901620202 . ПМИД 1150917 . S2CID 11379762 .
- ^ Морест Д.К. (июль 1975 г.). «Синаптические взаимоотношения клеток Гольджи типа II в медиальном коленчатом теле кошки». Журнал сравнительной неврологии . 162 (2): 157–193. дои : 10.1002/cne.901620202 . ПМИД 1150917 . S2CID 11379762 .
- ^ Морест Д.К. (июль 1975 г.). «Синаптические взаимоотношения клеток Гольджи типа II в медиальном коленчатом теле кошки». Журнал сравнительной неврологии . 162 (2): 157–193. дои : 10.1002/cne.901620202 . ПМИД 1150917 . S2CID 11379762 .
Внешние ссылки
[ редактировать ]- Поиск NIF - Клетка Гольджи, архивировано 4 марта 2016 г. в Wayback Machine через Neuroscience Information Framework.
- Поиск NIF - Клетка Гольджи II [ постоянная мертвая ссылка ] через информационную структуру нейронауки