Нервно-мышечный переход
| Нервно-мышечный переход | |
|---|---|
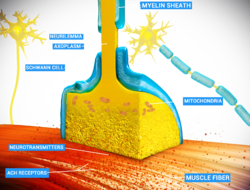 В нервно-мышечном соединении нервное волокно способно передавать сигнал мышечным волокнам, высвобождая АХ (и другие вещества), вызывая сокращение мышц. | |
 Схематическое изображение нервно-мышечного соединения | |
| Подробности | |
| Идентификаторы | |
| латинский | нервно-мышечный синапсис; нервно-мышечное соединение |
| МеШ | D009469 |
| ТД | Х2.00.06.1.02001 |
| ФМА | 61803 |
| Анатомическая терминология | |

Нервно -мышечное соединение (или мионевральное соединение ) — это химический синапс между двигательным нейроном и мышечным волокном . [1]
Он позволяет мотонейрону передавать сигнал мышечным волокнам, вызывая сокращение мышц .
Мышцам необходима иннервация, чтобы функционировать и даже просто поддерживать мышечный тонус , избегая атрофии . В нервно-мышечной системе нервы центральной и периферической нервной системы связаны и работают вместе с мышцами. [2] Синаптическая передача в нервно-мышечном соединении начинается, когда потенциал действия достигает пресинаптического окончания мотонейрона, который активирует потенциалзависимые кальциевые каналы , позволяя ионам кальция проникать в нейрон. Ионы кальция связываются с сенсорными белками ( синаптотагминами ) на синаптических везикулах, вызывая слияние везикул с клеточной мембраной и последующее высвобождение нейротрансмиттера из мотонейрона в синаптическую щель . У позвоночных мотонейроны выделяют ацетилхолин (АХ), небольшую молекулу нейромедиатора, которая диффундирует через синаптическую щель и связывается с никотиновыми рецепторами ацетилхолина (нАХР) на клеточной мембране мышечного волокна, также известной как сарколемма . нАХР являются ионотропными рецепторами, то есть они служат лиганд -управляемыми ионными каналами . Связывание АХ с рецептором может деполяризовать мышечное волокно, вызывая каскад, который в конечном итоге приводит к мышечному сокращению.
Заболевания нервно-мышечных соединений могут иметь генетическое и аутоиммунное происхождение. Генетические нарушения, такие как врожденный миастенический синдром , могут возникать из-за мутированных структурных белков, входящих в состав нервно-мышечного соединения, тогда как аутоиммунные заболевания, такие как миастения , возникают, когда антитела вырабатываются против никотиновых рецепторов ацетилхолина на сарколемме.
Структура и функции [ править ]
передача Квантовая
В нервно-мышечном соединении пресинаптические моторные аксоны заканчиваются на расстоянии 30 нанометров от клеточной мембраны или сарколеммы мышечного волокна. Сарколемма в месте соединения имеет впячивания, называемые постпереходными складками, которые увеличивают площадь ее поверхности, обращенной к синаптической щели. [3] Эти постпереходные складки образуют моторную концевую пластинку, которая усеяна никотиновыми рецепторами ацетилхолина (нАХР) с плотностью 10 000 рецепторов/мкм. 2 . [4] Пресинаптические аксоны оканчиваются выпуклостями, называемыми терминальными бутонами (или пресинаптическими окончаниями), которые выступают в сторону постсинаптических складок сарколеммы. У лягушки каждое окончание двигательного нерва содержит около 300 000 пузырьков со средним диаметром 0,05 микрометра. Везикулы содержат ацетилхолин. Некоторые из этих пузырьков собраны в группы по пятьдесят штук и расположены в активных зонах рядом с нервной мембраной. Активные зоны находятся на расстоянии около 1 микрометра друг от друга.Щель размером 30 нанометров между нервным окончанием и замыкательной пластинкой содержит сеть ацетилхолинэстеразы (АХЭ) с плотностью 2600 молекул фермента/мкм. 2 , удерживаемый структурными белками дистрофином и рапсином . Также присутствует рецепторный тирозинкиназный белок MuSK , сигнальный белок, участвующий в развитии нервно-мышечного соединения, который также удерживается на месте рапсином. [3]
пресинаптического нейрона Примерно раз в секунду в состоянии покоя один из синаптических пузырьков случайно сливается с клеточной мембраной в процессе, опосредованном белками SNARE . Слияние приводит к опорожнению содержимого пузырька (7000–10 000 молекул ацетилхолина) в синаптическую щель — процесс, известный как экзоцитоз . [5] Следовательно, экзоцитоз высвобождает ацетилхолин пакетами, которые называются квантами. Квант ацетилхолина диффундирует через сеть ацетилхолинэстеразы, где высокая локальная концентрация медиатора занимает все места связывания фермента на своем пути. Ацетилхолин, достигающий концевой пластинки, активирует около 2000 рецепторов ацетилхолина, открывая их ионные каналы, что позволяет ионам натрия перемещаться в концевую пластинку, вызывая деполяризацию ~ 0,5 мВ, известную как миниатюрный потенциал концевой пластинки (MEPP). К моменту высвобождения ацетилхолина из рецепторов ацетилхолинэстераза разрушила связанный с ним АХ, что занимает около 0,16 мс, и, следовательно, может разрушить АХ, высвободившийся из рецепторов. [ нужна ссылка ]
При стимуляции двигательного нерва между поступлением нервного импульса в окончания двигательного нерва и первым ответом замыкательной пластинки происходит задержка всего 0,5–0,8 мс. [6] Прибытие потенциала действия двигательного нерва на пресинаптическое окончание нейрона открывает потенциал-зависимые кальциевые каналы и Ca 2+ Ионы пресинаптического нейрона переходят из внеклеточной жидкости в цитозоль . Этот приток Ca 2+ заставляет несколько сотен нейротрансмиттеры содержащих везикул, , сливаться с клеточной мембраной пресинаптического нейрона посредством белков SNARE , высвобождая их кванты ацетилхолина путем экзоцитоза. Деполяризация концевой пластинки высвобождаемым ацетилхолином называется потенциалом концевой пластинки (EPP). EPP достигается, когда ACh связывается с никотиновыми рецепторами ацетилхолина (nAChR) на концевой пластинке мотора и вызывает приток ионов натрия. Этот приток ионов натрия генерирует ПКП (деполяризацию) и запускает потенциал действия, который перемещается вдоль сарколеммы в мышечные волокна через Т-трубочки (поперечные канальцы) посредством потенциалзависимых натриевых каналов. [7] Проведение потенциалов действия по Т-трубочкам стимулирует открытие потенциалзависимых Ca. 2+ каналы, механически связанные с Ca 2+ каналы высвобождения в саркоплазматическом ретикулуме. [8] Калифорния 2+ затем диффундирует из саркоплазматического ретикулума к миофибриллам, способствуя стимулированию сокращений. Таким образом, потенциал концевой пластинки отвечает за создание потенциала действия в мышечном волокне, который запускает мышечное сокращение. Передача от нерва к мышце происходит так быстро, потому что каждый квант ацетилхолина достигает концевой пластинки в миллимолярной концентрации, достаточно высокой, чтобы соединиться с рецептором с низким сродством, который затем быстро высвобождает связанный передатчик. [ нужна ссылка ]
рецепторы Ацетилхолиновые

- Рецептор, связанный с ионным каналом
- Ионы
- Лиганд (например, ацетилхолин )
Ацетилхолин представляет собой нейромедиатор, синтезируемый из пищевого холина и ацетил-КоА (АКоА), и участвует в стимуляции мышечной ткани у позвоночных , а также у некоторых беспозвоночных животных. У позвоночных подтип рецептора ацетилхолина, который обнаруживается в нервно-мышечных соединениях скелетных мышц, представляет собой никотиновый рецептор ацетилхолина (nAChR), который представляет собой лиганд-управляемый ионный канал . Каждая субъединица этого рецептора имеет характерную «цис-петлю», состоящую из остатка цистеина, за которым следуют 13 аминокислотных остатков и еще один остаток цистеина. Два остатка цистеина образуют дисульфидную связь , в результате чего образуется рецептор «цис-петли», способный связывать ацетилхолин и другие лиганды. Эти рецепторы цис-петли встречаются только у эукариот , но прокариоты обладают рецепторами АХ со схожими свойствами. [4] Не все виды используют холинергические нервно-мышечные соединения; например, раки и плодовые мухи имеют глутаматергическое нервно-мышечное соединение. [3]
АХР в скелетных нервно-мышечных соединениях образуют гетеропентамеры, состоящие из двух субъединиц α, одной β, одной ɛ и одной δ. [9] Когда одиночный лиганд ACh связывается с одной из α-субъединиц рецептора ACh, он вызывает конформационные изменения на границе раздела со второй α-субъединицей AChR. Это конформационное изменение приводит к увеличению сродства второй α-субъединицы ко второму лиганду ACh. Таким образом, AChR демонстрируют сигмоидальную кривую диссоциации из-за этого кооперативного связывания . [4] Наличие неактивной промежуточной структуры рецептора с одинарным лигандом удерживает АХ в синапсе , который в противном случае мог бы быть потерян в результате гидролиза или диффузии холинэстеразы . Сохранение этих лигандов АХ в синапсе может вызвать продолжительный постсинаптический ответ. [10]
Развитие [ править ]
Для развития нервно-мышечного соединения необходима передача сигналов как от окончания мотонейрона, так и от центральной области мышечной клетки. Во время развития мышечные клетки вырабатывают рецепторы ацетилхолина (AChR) и экспрессируют их в центральных областях в процессе, называемом препаттерном. Считается, что агрин гепарина , протеогликан , и киназа MuSK помогают стабилизировать накопление AChR в центральных областях миоцита. MuSK представляет собой рецепторную тирозинкиназу , что означает, что она индуцирует клеточную передачу сигналов путем связывания молекул фосфата с собственными участками, такими как тирозины , и с другими мишенями в цитоплазме . [11] При активации лигандом агрином MuSK передает сигналы через два белка, называемые « Dok-7 » и « rapsyn », индуцируя «кластеризацию» рецепторов ацетилхолина. [12] Высвобождение АХ развивающимися мотонейронами создает постсинаптические потенциалы в мышечной клетке, что положительно усиливает локализацию и стабилизацию развивающегося нервно-мышечного соединения. [13]
Эти результаты были частично продемонстрированы в ходе « нокаутных » исследований на мышах. У мышей с дефицитом агрина или MuSK нервно-мышечный синапс не формируется. Кроме того, мыши с дефицитом Dok-7 не образовывали ни кластеры рецепторов ацетилхолина, ни нервно-мышечные синапсы. [14]
Развитие нервно-мышечных соединений изучается преимущественно на модельных организмах, например на грызунах. было создано полностью человеческое нервно-мышечное соединение . Кроме того, в 2015 году in vitro с использованием эмбриональных стволовых клеток человека и стволовых клеток соматических мышц [15] В этой модели пресинаптические мотонейроны активируются оптогенетикой , и в ответ синаптически связанные мышечные волокна подергиваются при световой стимуляции.
Методы исследования [ править ]
Хосе дель Кастильо и Бернард Кац использовали ионофорез, чтобы определить расположение и плотность никотиновых рецепторов ацетилхолина (нАХР) в нервно-мышечных соединениях. С помощью этого метода микроэлектрод помещался внутрь моторной концевой пластинки мышечного волокна, а микропипетка, наполненная ацетилхолином (АХ), помещалась непосредственно перед концевой пластинкой в синаптической щели. На кончик микропипетки подавалось положительное напряжение, что вызывало выброс положительно заряженных молекул АХ из пипетки. Эти лиганды проникли в пространство, представляющее синаптическую щель, и связались с AChR. Внутриклеточный микроэлектрод контролировал амплитуду деполяризации . моторной концевой пластинки в ответ на связывание АХ с никотиновыми (ионотропными) рецепторами Кац и дель Кастильо показали, что амплитуда деполяризации ( возбуждающий постсинаптический потенциал ) зависит от близости микропипетки, высвобождающей ионы АХ, к концевой пластинке. Чем дальше микропипетка находилась от концевой пластинки двигателя, тем меньше была деполяризация в мышечном волокне. Это позволило исследователям определить, что никотиновые рецепторы локализованы на концевой пластинке мотора и имеют высокую плотность. [3] [4]
Токсины также используются для определения местоположения рецепторов ацетилхолина в нервно-мышечных соединениях. α-Бунгаротоксин — это токсин, обнаруженный у змей Bungarus multicinctus , который действует как антагонист ACh и необратимо связывается с AChR. Путем связывания поддающихся анализу ферментов, таких как пероксидаза хрена (HRP) или флуоресцентных белков, таких как зеленый флуоресцентный белок (GFP), с α-бунгаротоксином, AChR можно визуализировать и количественно оценить. [3]
поражающие нервно-мышечные Токсины , соединения
Нервно-паралитические газы [ править ]
Нервно-паралитические газы связываются и фосфорилируют АХЭ, эффективно деактивируя их. Накопление АХ в синаптической щели приводит к постоянному сокращению мышечных клеток, что приводит к тяжелым осложнениям, таким как паралич и смерть в течение нескольких минут после воздействия.

Ботулинический токсин [ править ]
Ботулинический токсин (также известный как ботулинический нейротоксин и коммерчески продаваемый под торговым названием «Ботокс») ингибирует высвобождение ацетилхолина в нервно-мышечных соединениях, воздействуя на белки SNARE. [3] Этот токсин проникает в нервное окончание в процессе эндоцитоза и впоследствии расщепляет белки SNARE, предотвращая слияние везикул ACh с внутриклеточной мембраной. Это вызывает преходящий вялый паралич и химическую денервацию, локализованную в пораженной поперечно-полосатой мышце. Ингибирование высвобождения АХ начинается примерно через две недели после инъекции. Через три месяца после возникновения торможения активность нейронов начинает частично восстанавливаться, а через шесть месяцев восстанавливается полная функция нейронов. [16]
Столбнячный токсин [ править ]
Столбнячный токсин, также известный как тетаноспазмин, представляет собой мощный нейротоксин, вырабатываемый Clostridium tetani и вызывающий болезненное состояние — столбняк. ЛД 50 этого токсина составляет примерно 1 нг/кг, что делает его вторым после ботулинического токсина D как самого смертоносного токсина в мире. Он действует очень похоже на ботулинический нейротоксин, прикрепляясь и эндоцитируя пресинаптическое нервное окончание и взаимодействуя с белками SNARE. Он отличается от ботулинического нейротоксина по нескольким признакам, наиболее очевидно по своему конечному состоянию, при котором тетаноспазмин вызывает спастический паралич в отличие от вялого паралича, наблюдаемого при использовании ботулинического нейротоксина.
Латротоксин [ править ]
Латротоксин (α-латротоксин), обнаруженный в яде пауков-вдов, также влияет на нервно-мышечные соединения, вызывая высвобождение ацетилхолина из пресинаптических клеток. Механизмы действия включают связывание с рецепторами пресинаптических клеток, активацию пути IP3/DAG , высвобождение кальция из внутриклеточных хранилищ и образование пор, что приводит к непосредственному притоку ионов кальция. Любой механизм вызывает повышение уровня кальция в пресинаптических клетках, что затем приводит к высвобождению синаптических везикул ацетилхолина. Латротоксин вызывает боль, сокращение мышц и, если его не лечить, потенциально паралич и смерть.
Змеиный яд [ править ]
Змеиные яды действуют как токсины на нервно-мышечные соединения и могут вызывать слабость и паралич . Яды могут действовать как пресинаптические, так и постсинаптические нейротоксины. [17]
Пресинаптические нейротоксины, широко известные как β-нейротоксины, поражают пресинаптические области нервно-мышечного соединения. Большинство этих нейротоксинов действуют путем ингибирования высвобождения нейротрансмиттеров, таких как ацетилхолин, в синапс между нейронами. Однако известно, что некоторые из этих токсинов усиливают высвобождение нейромедиаторов. Те, которые ингибируют высвобождение нейромедиаторов, создают нервно-мышечную блокаду , которая не позволяет сигнальным молекулам достичь своих постсинаптических рецепторов-мишеней. При этом жертва укуса змеи страдает от глубокой слабости. Такие нейротоксины плохо реагируют на противоядия. После одного часа введения этих токсинов, в том числе нотексина и тайпоксина , на многих пораженных нервных окончаниях проявляются признаки необратимого физического повреждения, в результате чего они теряют какие-либо синаптические пузырьки . [17]
Постсинаптические нейротоксины, также известные как α-нейротоксины, действуют противоположно пресинаптическим нейротоксинам, связываясь с постсинаптическими рецепторами ацетилхолина. Это предотвращает взаимодействие между ацетилхолином, высвобождаемым пресинаптическим терминалем, и рецепторами постсинаптической клетки. Фактически, открытие натриевых каналов, связанных с этими рецепторами ацетилхолина, запрещается, что приводит к нервно-мышечной блокаде, аналогичной эффектам, наблюдаемым из-за пресинаптических нейротоксинов. Это вызывает паралич мышц, задействованных в пораженных соединениях. В отличие от пресинаптических нейротоксинов, постсинаптические токсины легче поддаются воздействию противоядов, которые ускоряют диссоциацию токсина от рецепторов, что в конечном итоге приводит к обращению паралича. Эти нейротоксины экспериментально и качественно помогают в изучении плотности и оборота рецепторов ацетилхолина , а также в исследованиях, наблюдающих за направлением антител к пораженным рецепторам ацетилхолина у пациентов с диагнозом миастения гравис . [17]
Болезни [ править ]
Любое заболевание , которое нарушает синаптическую передачу между двигательным нейроном и мышечной клеткой, классифицируется под общим термином нервно-мышечных заболеваний . Эти расстройства могут быть наследственными или приобретенными и могут различаться по степени тяжести и смертности. В целом, большинство этих расстройств, как правило, вызваны мутациями или аутоиммунными нарушениями. Аутоиммунные нарушения, в случае нервно-мышечных заболеваний, как правило, опосредованы гуморально , опосредованы В-клетками и приводят к неправильному образованию антител против мотонейрона или белка мышечных волокон, которые мешают синаптической передаче или передаче сигналов.
Аутоиммунный [ править ]
Миастения гравис [ править ]
Миастения гравис — это аутоиммунное заболевание, при котором организм вырабатывает антитела либо против рецептора ацетилхолина (AchR) (в 80% случаев), либо против постсинаптической специфической для мышц киназы (MuSK) (0–10% случаев). При серонегативной миастении гравис белок 4, родственный рецептору липопротеинов низкой плотности , подвергается воздействию IgG1 , который действует как конкурентный ингибитор своего лиганда, предотвращая связывание лиганда со своим рецептором. Неизвестно, будет ли серонегативная миастения реагировать на стандартную терапию. [18]
Неонатальный MG [ править ]
Неонатальная МГ — аутоиммунное заболевание, которым страдает каждый восьмой ребенок, рожденный от матерей, у которых диагностирована миастения гравис (МГ). МГ может передаваться от матери к плоду путем перемещения антител АХР через плаценту . Признаки этого заболевания при рождении включают слабость, которая реагирует на антихолинэстеразные препараты, а также акинезию плода или отсутствие движений плода. Эта форма заболевания носит транзиторный характер и длится около трех месяцев. Однако в некоторых случаях неонатальный МГ может привести к другим последствиям для здоровья, таким как артрогрипоз и даже гибель плода. Считается, что эти состояния возникают, когда материнские антитела к AChR направляются на AChR плода , и могут сохраняться до 33-й недели беременности , когда субъединица γ AChR заменяется субъединицей ε. [19] [20]
Ламберта- Миастенический Итона синдром
Миастенический синдром Ламберта-Итона (LEMS) — аутоиммунное заболевание, поражающее пресинаптическую часть нервно-мышечного соединения. Это редкое заболевание характеризуется уникальной триадой симптомов: слабостью проксимальных мышц, вегетативной дисфункцией и арефлексией. [21] Слабость проксимальных мышц является продуктом патогенных аутоантител, направленных против потенциалзависимых кальциевых каналов P/Q-типа, что, в свою очередь, приводит к снижению высвобождения ацетилхолина из окончаний двигательных нервов в пресинаптических клетках. Примеры вегетативной дисфункции, вызванной LEMS, включают эректильную дисфункцию у мужчин, запор и, чаще всего, сухость во рту . Менее распространенные нарушения включают сухость глаз и изменение потоотделения . Арефлексия — это состояние, при котором сухожильные рефлексы снижены и могут временно исчезнуть после периода физических упражнений. [22]
У 50–60% пациентов с диагнозом LEMS также имеется сопутствующая опухоль , которая обычно представляет собой мелкоклеточную карциному легких (МРЛ). Этот тип опухоли также экспрессирует потенциалзависимые кальциевые каналы . [22] Часто LEMS также возникает наряду с миастенией. [21]
Лечение LEMS состоит из использования 3,4-диаминопиридина в качестве первой меры, который служит для увеличения сложного потенциала действия мышц, а также мышечной силы за счет удлинения времени, в течение которого потенциалзависимые кальциевые каналы остаются открытыми после блокирования потенциалзависимых калиевых каналов. В США лечение 3,4-диаминопиридином для пациентов с LEMS, отвечающих критериям, доступно бесплатно в рамках программы расширенного доступа. [23] [24] Дальнейшее лечение включает применение преднизолона и азатиоприна в том случае, если 3,4-диаминопиридин не помогает в лечении. [22]
Нейромиотония [ править ]
Нейромиотония (НМТ), также известная как синдром Исаака, отличается от многих других заболеваний, возникающих в нервно-мышечных соединениях. Вместо того, чтобы вызывать мышечную слабость, НМТ приводит к гипервозбуждению двигательных нервов. НМТ вызывает это гипервозбуждение, вызывая более длительную деполяризацию за счет подавления потенциалзависимых калиевых каналов , что вызывает большее высвобождение нейротрансмиттеров и повторяющиеся импульсы. Это увеличение скорости стрельбы приводит к более активной передаче и, как следствие, к большей мышечной активности у пострадавшего. Также считается, что НМТ имеет аутоиммунное происхождение из-за его связи с аутоиммунными симптомами у больного. [19]
Генетический [ править ]
синдромы Врожденные миастенические
Врожденные миастенические синдромы (ВМС) по своим функциям очень похожи как на МГ, так и на LEMS, но основное различие между ВМС и этими заболеваниями заключается в том, что ВМС имеет генетическое происхождение. В частности, эти синдромы представляют собой заболевания, возникающие из-за мутаций, обычно рецессивных , в 1 из по меньшей мере 10 генов, которые влияют на пресинаптические, синаптические и постсинаптические белки нервно-мышечного соединения. Такие мутации обычно возникают в ε-субъединице АХР, [19] тем самым влияя на кинетику и экспрессию самого рецептора. Замены или делеции одиночных нуклеотидов могут привести к потере функции субъединицы. Другие мутации , например, затрагивающие ацетилхолинэстеразу и ацетилтрансферазу , также могут вызывать экспрессию CMS, причем последняя специфически связана с эпизодическим апноэ . [25] Эти синдромы могут проявляться в разное время жизни человека. Они могут возникнуть во время внутриутробной фазы, вызывая акинезию плода , или в перинатальном периоде, во время которого могут наблюдаться определенные состояния, такие как артрогрипоз , птоз , гипотония , офтальмоплегия , а также затруднения при кормлении или дыхании. Они также могут активироваться в подростковом или взрослом возрасте, вызывая у человека развитие синдрома медленных каналов. [19]
Лечение определенных подтипов CMS (постсинаптическая CMS с быстрым каналом) [26] [27] аналогично лечению других нервно-мышечных заболеваний. 3,4-Диаминопиридин , препарат первой линии для лечения LEMS, находится в стадии разработки в качестве орфанного препарата для лечения CMS. [28] в США и бесплатно доступны пациентам, имеющим право на участие в программе расширенного доступа. [23] [24]
См. также [ править ]
Внешние ссылки [ править ]
- Гистологическое изображение: 21501lca - Система обучения гистологии в Бостонском университете.
Дальнейшее чтение [ править ]
- Кандел, ER ; Шварц Дж. Х.; Джесселл ТМ. (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 0-8385-7701-6 .
- Николлс, Дж.Г.; А.Р. Мартин; Б.Г. Уоллес; П. А. Фукс (2001). От нейрона к мозгу (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 0-87893-439-1 .
- Энгель, АГ (2004). Миология (3-е изд.). Нью-Йорк: McGraw Hill Professional. ISBN 0-07-137180-Х .
Ссылки [ править ]
- ^ Левитан, Ирвин; Качмарек, Леонард (19 августа 2015 г.). «Межклеточная связь». Нейрон: клеточная и молекулярная биология (4-е изд.). Нью-Йорк, штат Нью-Йорк: Издательство Оксфордского университета. стр. 153–328. ISBN 978-0199773893 .
- ^ Ригель, К. (август 2016 г.). «Старение нервно-мышечной системы и саркопения: митохондриальная перспектива» . Дж. Физиол . 594 (16): 4499–4512. дои : 10.1113/JP271212 . ПМЦ 4983621 . ПМИД 26921061 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Николлс, Джон Г.; А. Роберт Мартин; Пол А. Фукс; Дэвид А. Браун; Мэтью Э. Даймонд; Дэвид А. Вайсблат (2012). От нейрона к мозгу (5-е изд.). Сандерленд: Sinauer Associates.
- ↑ Перейти обратно: Перейти обратно: а б с д Синус СМ (июль 2012 г.). «Рецептор ацетилхолина концевой пластинки: структура, механизм, фармакология и заболевания» . Физиол. Преподобный . 92 (3): 1189–234. doi : 10.1152/physrev.00015.2011 . ПМК 3489064 . ПМИД 22811427 .
- ^ Уильям Ван дер Клот; Хорди Молго (1994). «Квантовое высвобождение ацетилхолина в нервно-мышечном соединении позвоночных». Физиол. Преподобный . 74 (4): 900–991. дои : 10.1152/physrev.1994.74.4.899 . ПМИД 7938228 .
- ^ Кац, Бернард (1966). Нерв, мышца и синапс . Нью-Йорк: МакГроу-Хилл. п. 114.
- ^ МакКинли, Майкл; О'Локлин, Валери; Пеннефатер-О'Брайен, Элизабет; Харрис, Рональд (2015). Анатомия человека . Нью-Йорк: Образование Макгроу-Хилл. п. 300. ИСБН 978-0-07-352573-0 .
- ^ Фокс, Стюарт (2016). Физиология человека . Нью-Йорк: Образование Макгроу-Хилл. п. 372. ИСБН 978-0-07-783637-5 .
- ^ анестезия Миллера, 7-е издание.
- ^ Скука М., Мозжимас Дж.В. (1992). «Постсинаптическая потенциация и десенсибилизация рецепторов концевых пластинок позвоночных». Прог. Нейробиол . 38 (1): 19–33. дои : 10.1016/0301-0082(92)90033-Б . ПМИД 1736323 . S2CID 38497982 .
- ^ Валенсуэла Д., Ститт Т., ДиСтефано П., Рохас Э., Мэттссон К., Комптон Д., Нуньес Л., Парк Дж., Старк Дж., Гис Д. (1995). «Рецептор тирозинсиназы, специфичный для скелетных мышц: экспрессия в эмбриональных мышцах, в нервно-мышечных соединениях и после травмы» . Нейрон . 15 (3): 573–84. дои : 10.1016/0896-6273(95)90146-9 . ПМИД 7546737 . S2CID 17575761 .
- ^ Гласс Д., Боуэн Д., Ститт Т., Радзиевски С., Бруно Дж., Райан Т., Гис Д., Шах С., Мэттссон К., Берден С., ДиСтефано П., Валенсуэла Д., ДеКьяра Т., Янкопулос Г. (1996). «Агрин действует через рецепторный комплекс MuSK» . Клетка . 85 (4): 513–23. дои : 10.1016/S0092-8674(00)81252-0 . ПМИД 8653787 . S2CID 14930468 .
- ^ Витземан V (ноябрь 2006 г.). «Развитие нервно-мышечного соединения». Ресурсы клеточных тканей . 326 (2): 263–71. дои : 10.1007/s00441-006-0237-x . hdl : 11858/00-001M-0000-002B-BE74-A . ПМИД 16819627 . S2CID 30829665 .
- ^ Окада К., Иноуэ А., Окада М., Мурата Ю., Какута С., Джигами Т., Кубо С., Сираиси Х., Эгути К., Мотомура М., Акияма Т., Ивакура Ю., Хигучи О., Яманаси Ю. (2006). «Мышечный белок Dok-7 необходим для нервно-мышечного синаптогенеза». Наука . 312 (5781): 1802–5. Бибкод : 2006Sci...312.1802O . дои : 10.1126/science.1127142 . ПМИД 16794080 . S2CID 45730054 .
- ^ Стейнбек, Дж.А.; Джайсвал, МК; Колдер, Эл.; Кишиневский, С; Вейсгаупт, А; Тойка, КВ; Гольдштейн, Пенсильвания; Студер, Л. (7 января 2016 г.). «Функциональная связность под оптогенетическим контролем позволяет моделировать нервно-мышечные заболевания человека» . Клеточная стволовая клетка . 18 (1): 134–43. дои : 10.1016/j.stem.2015.10.002 . ПМК 4707991 . ПМИД 26549107 .
- ^ Папапетропулос С., Сингер С. (апрель 2007 г.). «Ботулинический токсин при двигательных расстройствах». Семин Нейрол . 27 (2): 183–94. дои : 10.1055/s-2007-971171 . ПМИД 17390263 .
- ↑ Перейти обратно: Перейти обратно: а б с Льюис Р.Л., Гутманн Л. (июнь 2004 г.). «Змеиные яды и нервно-мышечные соединения». Семин Нейрол . 24 (2): 175–9. дои : 10.1055/s-2004-830904 . ПМИД 15257514 .
- ^ Финстерер Дж., Папич Л., Ауэр-Грумбах М. (октябрь 2011 г.). «Заболевания двигательных нейронов, нервов и нервно-мышечных соединений». Курс. Мнение. Нейрол . 24 (5): 469–74. doi : 10.1097/WCO.0b013e32834a9448 . ПМИД 21825986 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б с д Ньюсом-Дэвис Дж. (июль 2007 г.). «Новое разнообразие нарушений нервно-мышечных соединений» . Акта Миол . 26 (1): 5–10. ПМЦ 2949330 . ПМИД 17915563 .
- ^ Бардхан, М.; Догра, Х.; Саманта, Д. (2021). «Неонатальная миастения Гравис» . СтатПерлз . СтатПерлс. ПМИД 32644361 .
- ↑ Перейти обратно: Перейти обратно: а б Луигетти М., Модони А., Ло Монако М. (октябрь 2012 г.). «Низкочастотная повторяющаяся нервная стимуляция при миастеническом синдроме Ламберта-Итона: особенности декрементального паттерна на основе одноцентрового опыта». Клин Нейрофизиол . 124 (4): 825–6. дои : 10.1016/j.clinph.2012.08.026 . ПМИД 23036181 . S2CID 11396376 .
- ↑ Перейти обратно: Перейти обратно: а б с Титулаер М.Дж., Ланг Б., Вершуурен Дж.Дж. (декабрь 2011 г.). «Миастенический синдром Ламберта-Итона: от клинических характеристик к терапевтическим стратегиям». Ланцет Нейрол . 10 (12): 1098–107. дои : 10.1016/S1474-4422(11)70245-9 . ПМИД 22094130 . S2CID 27421424 .
- ↑ Перейти обратно: Перейти обратно: а б [1] , Пресс-релиз Ассоциации мышечной дистрофии
- ↑ Перейти обратно: Перейти обратно: а б [2] Архивировано 25 июля 2015 г. в Wayback Machine , Отчет о редких заболеваниях.
- ^ Харпер CM (март 2004 г.). «Врожденные миастенические синдромы». Семин Нейрол . 24 (1): 111–23. дои : 10.1055/s-2004-829592 . ПМИД 15229798 .
- ^ Энгель А.Г. и др. (апрель 2015 г.). «Врожденные миастенические синдромы: патогенез, диагностика и лечение» . Ланцет Нейрол . 14 (4): 420–34. дои : 10.1016/S1474-4422(14)70201-7 . ПМК 4520251 . ПМИД 25792100 .
- ^ Энгель А.Г. и др. (2012). «Новые горизонты врожденных миастенических синдромов» . Энн, Нью-Йоркская академия наук . 1275 (1): 1275:54–62. Бибкод : 2012NYASA1275...54E . дои : 10.1111/j.1749-6632.2012.06803.x . ПМЦ 3546605 . ПМИД 23278578 .
- ^ [3] , обозначение FDA для орфанных препаратов.
| Базы данных органов управления : Национальные |
|---|