Никотиновый рецептор ацетилхолина


Никотиновые рецепторы ацетилхолина , или nAChR , представляют собой рецепторные полипептиды , которые реагируют на нейромедиатор ацетилхолин . Никотиновые рецепторы также реагируют на такие лекарства, как агонист никотин . Они обнаружены в центральной и периферической нервной системе, мышцах и многих других тканях многих организмов. В нервно-мышечном соединении они являются первичными рецепторами в мышцах, обеспечивающими двигательную нервно-мышечную связь, которая контролирует сокращение мышц. В периферической нервной системе : (1) они передают исходящие сигналы от пресинаптических к постсинаптическим клеткам симпатической и парасимпатической нервной системы и (2) являются рецепторами скелетных мышц, которые получают ацетилхолин, высвобождаемый для подачи сигнала о мышечном сокращении. В иммунной системе нАХР регулируют воспалительные процессы и передают сигналы через отдельные внутриклеточные пути. [1] У насекомых холинергическая система ограничена центральной нервной системой . [2]
Никотиновые рецепторы считаются холинергическими рецепторами , поскольку они реагируют на ацетилхолин. Никотиновые рецепторы получили свое название от никотина , который не стимулирует мускариновые рецепторы ацетилхолина , а вместо этого избирательно связывается с никотиновыми рецепторами. [3] [4] [5] Мускариновый рецептор ацетилхолина также получил свое название от химического вещества, которое избирательно присоединяется к этому рецептору — мускарина . [6] Сам ацетилхолин связывается как с мускариновыми, так и с никотиновыми рецепторами ацетилхолина. [7]
Являясь ионотропными рецепторами, нАХР напрямую связаны с ионными каналами. Новые данные свидетельствуют о том, что эти рецепторы в некоторых случаях могут также использовать вторичные мессенджеры (как это делают метаботропные рецепторы ). [8] Никотиновые рецепторы ацетилхолина являются наиболее изученными из ионотропных рецепторов. [3]
Поскольку никотиновые рецепторы помогают передавать исходящие сигналы для симпатической и парасимпатической систем, антагонисты никотиновых рецепторов, такие как гексаметоний, препятствуют передаче этих сигналов. Так, например, антагонисты никотиновых рецепторов нарушают барорефлекс. [9] который обычно корректирует изменения артериального давления путем симпатической и парасимпатической стимуляции сердца.
Структура
[ редактировать ]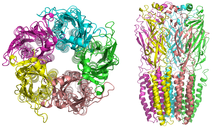
Никотиновые рецепторы с молекулярной массой 290 кДа . [10] состоят из пяти субъединиц, расположенных симметрично вокруг центральной поры . [3] Каждая субъединица состоит из четырех трансмембранных доменов, N- и C-концы которых расположены внеклеточно. Они обладают сходством с ГАМК А рецепторами , рецепторами глицина и рецепторами серотонина 3 типа (которые все являются ионотропными рецепторами) или характерными белками Cys-петли . [11]
У позвоночных никотиновые рецепторы подразделяются на два подтипа в зависимости от их основных мест экспрессии: никотиновые рецепторы мышечного типа и никотиновые рецепторы нейронального типа . В рецепторах мышечного типа, обнаруженных в нервно-мышечном соединении, рецепторы представляют собой либо эмбриональную форму, состоящую из субъединиц α 1 , β 1 , γ и δ в соотношении 2:1:1:1 ((α 1 ) 2 β 1 γδ), или взрослая форма, состоящая из субъединиц α 1 , β 1 , δ и ε в соотношении 2:1:1:1 ((α 1 ) 2 β 1 δε). [3] [4] [5] [12] Подтипы нейронов представляют собой различные гомомерные (все субъединицы одного типа) или гетеромерные (по крайней мере, один α и один β) комбинации двенадцати различных субъединиц никотиновых рецепторов: α 2 -α 10 и β 2 -β 4 . Примеры подтипов нейронов включают: (α 4 ) 3 (β 2 ) 2 , (α 4 ) 2 (β 2 ) 3 , (α 3 ) 2 (β 4 ) 3 , α 4 α 6 β 3 (β 2 ) 2 , (α 7 ) 5 и многие другие. В рецепторах как мышечного, так и нейронального типа субъединицы очень похожи друг на друга, особенно в гидрофобных областях. [13]
Ряд исследований электронной микроскопии и рентгеновской кристаллографии предоставили структурную информацию с очень высоким разрешением для мышечных и нейрональных нАХР и их связывающих доменов. [10] [14] [15] [16]
Связывание
[ редактировать ]Как и во всех лиганд-управляемых ионных каналах, открытие поры канала нАХР требует связывания химического мессенджера. Для обозначения молекул, связывающих рецепторы, используются несколько разных терминов, таких как лиганд , агонист или передатчик. Помимо эндогенного агониста ацетилхолина , агонисты нАХР включают никотин , эпибатидин и холин . Никотиновые антагонисты, блокирующие рецептор, включают мекамиламин, дигидро-β-эритроидин и гексаметоний . [13]
В нАХР мышечного типа сайты связывания ацетилхолина расположены на границе раздела субъединиц α и ε или δ. В нейрональных нАХР сайт связывания расположен на границе раздела субъединиц α и β или между двумя субъединицами α в случае α7 - рецепторов. Сайт связывания расположен во внеклеточном домене рядом с N-концом . [4] [17] Когда агонист связывается с сайтом, все присутствующие субъединицы претерпевают конформационные изменения и канал открывается. [18] и открывается пора диаметром около 0,65 нм. [4]
Открытие канала
[ редактировать ]Никотиновые АХР могут существовать в различных взаимопревращаемых конформационных состояниях. Связывание агониста стабилизирует открытое и десенсибилизированное состояния. В нормальных физиологических условиях для открытия рецептору необходимо ровно две молекулы АХ. [19] Открытие канала позволяет положительно заряженным ионам перемещаться по нему; в частности, натрий поступает в клетку, а калий выходит. Чистый поток положительно заряженных ионов направлен внутрь.
НАХР представляет собой неселективный катионный канал, а это означает, что через него могут проходить несколько различных положительно заряженных ионов. [3] Он проницаем для Na + и К + , с некоторыми комбинациями субъединиц, которые также проницаемы для Ca 2+ . [4] [20] [21] Количество натрия и калия, которые каналы пропускают через свои поры (их проводимость ), варьируется от 50 до 110 пСм , причем проводимость зависит от состава конкретной субъединицы, а также от проникающего иона. [22]
Многие нейрональные нАХР могут влиять на высвобождение других нейротрансмиттеров. [5] Канал обычно открывается быстро и имеет тенденцию оставаться открытым до тех пор , пока агонист не диффундирует , что обычно занимает около 1 миллисекунды . [4] AChR могут спонтанно открываться без связанных лигандов или могут спонтанно закрываться со связанными лигандами, а мутации в канале могут изменить вероятность любого события. [23] [18] Следовательно, связывание ACh изменяет вероятность открытия пор, которая увеличивается по мере связывания большего количества ACh.
nAChR не способен связывать ACh при связывании с любым из змеиного яда α-нейротоксинов . Эти α- нейротоксины прочно и нековалентно связываются с нАХР скелетных мышц и нейронов, блокируя тем самым действие АХ на постсинаптическую мембрану, ингибируя поток ионов и приводя к параличу и смерти. nAChR содержит два сайта связывания нейротоксинов змеиного яда. Прогресс в обнаружении динамики связывающего действия этих сайтов оказался трудным, хотя недавние исследования с использованием нормального режима динамики [24] помогли предсказать природу механизмов связывания змеиных токсинов и АХ с нАХР. Эти исследования показали, что закручивающееся движение, вызванное связыванием ACh, вероятно, ответственно за открытие пор, и что одной или двух молекул α-бунгаротоксина (или другого длинноцепочечного α-нейротоксина) достаточно, чтобы остановить это движение. Токсины, по-видимому, сцепляют соседние субъединицы рецептора, подавляя поворот и, следовательно, открывающее движение. [25]
Эффекты
[ редактировать ]Активация рецепторов никотином изменяет состояние нейронов посредством двух основных механизмов. С одной стороны, движение катионов вызывает деполяризацию плазматической мембраны (что приводит к возникновению возбуждающего постсинаптического потенциала в нейронах ), что приводит к активации потенциалзависимых ионных каналов . С другой стороны, поступление кальция прямо или косвенно действует на различные внутриклеточные каскады . Это приводит, например, к регуляции активности некоторых генов или высвобождению нейротрансмиттеров . [ нужна ссылка ]
Регулирование
[ редактировать ]Десенсибилизация
[ редактировать ]Лиганд-связанная десенсибилизация рецепторов была впервые охарактеризована Кацем и Теслеффом в никотиновом рецепторе ацетилхолина. [26]
Длительное или неоднократное воздействие раздражителя часто приводит к снижению реакции этого рецептора на раздражитель, что называется десенсибилизацией. Функция нАХР может модулироваться фосфорилированием. [27] путем активации вторичных мессенджер-зависимых протеинкиназ. ПКА [26] и ПКС , [28] а также тирозинкиназы, [29] Было показано, что фосфорилирует нАХР, что приводит к его десенсибилизации. Сообщалось, что после длительного воздействия агониста на рецептор сам агонист вызывает индуцированные агонистом конформационные изменения рецептора, что приводит к десенсибилизации рецептора. [30]
Десенсибилизированные рецепторы могут вернуться в пролонгированное открытое состояние, когда агонист связывается в присутствии положительного аллостерического модулятора, например PNU-120,596 . [31] Кроме того, есть данные, указывающие на то, что определенные молекулы шаперонов оказывают регуляторное воздействие на эти рецепторы. [32]
Роли
[ редактировать ]Субъединицы никотиновых рецепторов принадлежат к мультигенному семейству (16 членов у человека), и сборка комбинаций субъединиц приводит к образованию большого количества различных рецепторов (более подробную информацию см. в базе данных Ligand-Gated Ion Channel ). Эти рецепторы с весьма изменчивыми кинетическими , электрофизиологическими и фармакологическими свойствами по-разному реагируют на никотин при очень разных эффективных концентрациях. Это функциональное разнообразие позволяет им принимать участие в двух основных типах нейротрансмиссии. Классическая синаптическая передача (проводная передача) предполагает высвобождение высоких концентраций нейромедиатора, действующего на непосредственно соседние рецепторы. Напротив, паракринная передача (объемная передача) включает нейротрансмиттеры, высвобождаемые окончаниями аксонов , которые затем диффундируют через внеклеточную среду, пока не достигнут своих рецепторов, которые могут находиться на расстоянии. [33] Никотиновые рецепторы также можно найти в разных синаптических местах; например, мышечный никотиновый рецептор всегда функционирует постсинаптически. Нейрональные формы рецептора могут быть обнаружены как постсинаптически (участвуют в классической нейротрансмиссии), так и пресинаптически. [34] где они могут влиять на высвобождение нескольких нейротрансмиттеров.
Субъединицы
[ редактировать ]Идентифицировано 17 субъединиц нАХР позвоночных, которые разделены на субъединицы мышечного и нейронального типа. Хотя субъединица/ген α8 присутствует у видов птиц, таких как курица, она не присутствует у человека или млекопитающих. [35]
Субъединицы nAChR были разделены на четыре подсемейства (I–IV) на основании сходства белковых последовательностей. [36] Кроме того, подсемейство III было разделено на три типа.
| нейронального типа | Мышечный тип | ||||
| я | II | III | IV | ||
|---|---|---|---|---|---|
| а9 , а10 | а7 , а8 | 1 | 2 | 3 | а1 , б1 , г , в , е |
| а2 , а3 , а4 , а6 | б2 , б4 | б3 , а5 | |||
- α-гены: CHRNA1 (мышечный), CHRNA2 (нейрональный), CHRNA3 , CHRNA4 , CHRNA5 , CHRNA6 , CHRNA7 , CHRNA8 , CHRNA9 , CHRNA10 .
- β-гены: CHRNB1 (мышечный), CHRNB2 (нейронный), CHRNB3 , CHRNB4.
- Другие гены: CHRND (дельта), CHRNE (эпсилон), CHRNG (гамма).
Нейрональные nAChRs представляют собой трансмембранные белки , которые образуют пентамерные структуры , собранные из семейства субъединиц, состоящих из α 2 –α 10 и β 2 –β 4 . [37] Эти субъединицы были обнаружены с середины 1980-х по начало 1990-х годов, когда кДНК для нескольких субъединиц нАХР были клонированы из мозга крыс и кур, что привело к идентификации одиннадцати различных генов (двенадцать у кур), которые кодируют нейрональные субъединицы нАХР; Идентифицированные гены-субъединицы были названы α 2 –α 10 (α 8 обнаружен только у кур) и β 2 –β 4 . [38] Также было обнаружено, что различные комбинации субъединиц могут образовывать функциональные нАХР, которые могут активироваться ацетилхолином и никотином , а различные комбинации субъединиц создают подтипы нАХР с разнообразными функциональными и фармакологическими свойствами. [39] При экспрессии по отдельности α7 , α8 , α9 и α10 способны образовывать функциональные рецепторы, но другие субъединицы α требуют присутствия субъединиц β для формирования функциональных рецепторов. [37] Было обнаружено, что у млекопитающих субъединицы nAchR кодируются 17 генами, из них девять генов, кодирующих α-субъединицы, и три, кодирующие β-субъединицы, экспрессируются в мозге. β 2 nAChR, содержащие субъединицу (β 2 nAChR) и α 7 nAChR, широко экспрессируются в головном мозге, тогда как другие субъединицы nAChR имеют более ограниченную экспрессию. [40] Пентамерная сборка нАХР подвергается воздействию субъединиц, которые продуцируются в различных типах клеток, например, в легких человека, где эпителиальные и мышечные пентамеры существенно различаются. [41]
CHRNA5/A3/B4
[ редактировать ]Важный кластер генов nAchR (CHRNA5/A3/B4) содержит гены, кодирующие субъединицы α 5 , α3 и β 4 . Генетические исследования выявили однонуклеотидные полиморфизмы (SNP) в хромосомном локусе, кодирующем эти три гена nAChR, как факторы риска никотиновой зависимости , рака легких , хронической обструктивной болезни легких , алкоголизма и заболеваний периферических артерий . [37] [42] Гены субъединицы nAChR CHRNA5/A3/B4 обнаружены плотным кластером в хромосомной области 15q24–25. Субъединицы nAChR, кодируемые этим локусом, образуют преобладающие подтипы никотиновых рецепторов, экспрессируемые в периферической нервной системе (ПНС) и других ключевых участках центральной нервной системы (ЦНС) , таких как медиальная хабенула , структура между лимбической частью переднего мозга и средним мозгом, участвующая в основных процессах. пути холинергической системы. [37] Дальнейшие исследования генов CHRNA5/A3/B4 показали, что «нейрональные» гены nAChR также экспрессируются в ненейрональных клетках, где они участвуют в различных фундаментальных процессах, таких как воспаление. [43] Гены CHRNA5/A3/B4 совместно экспрессируются во многих типах клеток, а транскрипционная активность промоторных областей трех генов регулируется многими из одних и тех же факторов транскрипции, демонстрируя, что их кластеризация может отражать контроль экспрессии генов. [37]
CHRNA6/CHRNB3
[ редактировать ]CHRNB3 и CHRNA6 также сгруппированы в кластер генов, расположенный на 8p11. [42] Многочисленные исследования показали, что SNPS в CHRNB3–CHRNA6 связаны с никотиновой зависимостью и курением, например, два SNP в CHRNB3: rs6474413 и rs10958726. [42] Генетические вариации в этом регионе также влияют на склонность к употреблению наркотиков, включая кокаин и алкоголь. [44] Никотиновые рецепторы, содержащие вентральной области покрышки субъединицы α6 или β3, экспрессируемые в областях мозга, особенно в и черной субстанции , важны для поведения, связанного с употреблением наркотиков, из-за их роли в высвобождении дофамина . [45] Генетические вариации в этих генах могут изменять чувствительность к наркотикам различными способами, включая изменение аминокислотной структуры белка или вызывать изменения в регуляции транскрипции и трансляции. [44]
CHRNA4/CHRNB2
[ редактировать ]Другие хорошо изученные гены nAChR включают CHRNA4 и CHRNB2, которые ассоциированы с генами аутосомно-доминантной ночной лобной эпилепсии (ADNFLE) . [42] [46] Обе субъединицы нАХР присутствуют в головном мозге, и возникновение мутаций в этих двух субъединицах вызывает генерализованный тип эпилепсии. Примеры включают инсерционную мутацию CHRNA4 776ins3, которая связана с ночными судорогами и психическими расстройствами, а также мутацию CHRNB2 I312M, которая, по-видимому, вызывает не только эпилепсию, но и очень специфические когнитивные нарушения, такие как дефицит обучения и памяти. [46] [47] Между этими двумя генами существуют естественные генетические вариации, и анализ однонуклеотидных полиморфизмов (SNP) и других модификаций генов показывает более высокую изменчивость в гене CHRNA4, чем в гене CHRNB2, подразумевая, что nAChR β 2 , белок, кодируемый CHRNB2, связывает с большим количеством субъединиц, чем α 4 . Сообщалось также, что CHRNA2 является третьим кандидатом на ночные судороги в лобных долях. [42] [46]
CHRNA7
[ редактировать ]В нескольких исследованиях сообщалось о связи между CHRNA7 и эндофенотипами психических расстройств и никотиновой зависимостью, что способствует значительной клинической значимости α7 и проводимым по нему исследованиям. [46] CHRNA7 был одним из первых генов, которые считались связанными с шизофренией . Исследования выявили несколько полиморфизмов промотора CHRNA7, которые снижают транскрипционную активность генов, что связано с шизофренией, что согласуется с обнаружением сниженных уровней a7 nAChR в мозге пациентов с шизофренией. [46] оба подтипа nAChRs, α 4 β 2 и α 7 , значительно снижены в посмертных исследованиях людей с шизофренией. Было обнаружено, что [48] Кроме того, уровень курения значительно выше среди больных шизофренией, а это означает, что курение никотина может быть формой самолечения. [49]
Известные вариации
[ редактировать ]Никотиновые рецепторы представляют собой пентамеры этих субъединиц; т.е. каждый рецептор содержит пять субъединиц. Таким образом, существует огромный потенциал вариаций этих субъединиц, некоторые из которых встречаются чаще, чем другие. Наиболее широко выраженные подтипы включают (α 1 ) 2 β 1 δε (взрослый мышечный тип), (α 3 ) 2 (β 4 ) 3 (ганглиозный тип), (α 4 ) 2 (β 2 ) 3 (ЦНС-тип). тип) и (α 7 ) 5 (другой тип ЦНС). [50] Далее следует сравнение:
| Рецепторного типа | Расположение | Эффект; функции | Никотиновые агонисты | Никотиновые антагонисты |
|---|---|---|---|---|
| Тип мышц : (а 1 ) 2 б 1 нет [50] или (а 1 ) 2 б 1 дг | Нервно-мышечный переход | ВПСП , главным образом за счет увеличения Na + и К + проницаемость |
| |
| Ганглионного типа : (а 3 ) 2 (б 4 ) 3 | вегетативные ганглии | ВПСП , главным образом за счет увеличения Na + и К + проницаемость | ||
| Гетеромерный ЦНС-тип: (а 4 ) 2 (б 2 ) 3 | Мозг | Пост- и пресинаптическое возбуждение . [50] главным образом за счет увеличения Na + и К + проницаемость. Основной подтип, участвующий в повышении внимания и вознаграждающих эффектах никотина, а также в патофизиологии никотиновой зависимости. [52] [53] [54] | ||
| Дальнейший тип ЦНС: (а 3 ) 2 (б 4 ) 3 | Мозг | Пост- и пресинаптическое возбуждение | ||
| Гомомерный ЦНС-тип: (а 7 ) 5 | Мозг | Пост- и пресинаптическое возбуждение . [50] главным образом за счет увеличения Na + , К + и Ca 2+ проницаемость. Основной подтип, участвующий в некоторых когнитивных эффектах никотина. [55] Более того, активация (α 7 ) 5 может улучшить реакцию нейрососудистой связи при нейродегенеративных заболеваниях. [56] и нейрогенез при ишемическом инсульте. [57] Также участвует в проангиогенном эффекте никотина и ускоряет прогрессирование хронической болезни почек у курильщиков. [58] [59] [60] |
См. также
[ редактировать ]- Мускариновый агонист
- Мускариновый антагонист
- TDBzхолин
- Миастения гравис
- Врожденный миастенический синдром
- Адренергический
- Холинергический
- Рецептор ацетилхолина (холинергический рецептор)
Ссылки
[ редактировать ]- ^ Лу Б., Кван К., Левин Ю.А., Олофссон П.С., Ян Х., Ли Дж. и др. (август 2014 г.). «Передача сигналов никотинового ацетилхолинового рецептора α7 ингибирует активацию воспаления, предотвращая высвобождение митохондриальной ДНК» . Молекулярная медицина . 20 (1): 350–8. дои : 10.2119/molmed.2013.00117 . ПМЦ 4153835 . ПМИД 24849809 .
- ^ Ямамото I (1999). «Никотин в никотиноиды: с 1962 по 1997 год». Никотиноидные инсектициды и никотиновый рецептор ацетилхолина . стр. 3–27. дои : 10.1007/978-4-431-67933-2_1 . ISBN 978-4-431-68011-6 .
- ^ Перейти обратно: а б с д и ж г час я дж к Первс Д., Августин Г.Дж., Фитцпатрик Д., Холл У.К., ЛаМантия А.С., Макнамара Дж.О., Уайт Л.Е. (2008). Нейронаука (4-е изд.). Синауэр Ассошиэйтс. стр. 122 –6. ISBN 978-0-87893-697-7 .
- ^ Перейти обратно: а б с д и ж Сигел Г.Дж., Агранов Б.В., Фишер С.К., Альберс Р.В., Улер, доктор медицинских наук (1999). «Физиология и фармакология рецепторов ГАМК» . Базовая нейрохимия: молекулярные, клеточные и медицинские аспекты (6-е изд.). Американское общество нейрохимии . Проверено 1 октября 2008 г.
{{cite book}}:|website=игнорируется ( помогите ) - ^ Перейти обратно: а б с Итье V, Бертран Д. (август 2001 г.). «Нейрональные никотиновые рецепторы: от структуры белка к функции» . Письма ФЭБС . 504 (3): 118–25. Бибкод : 2001FEBSL.504..118I . дои : 10.1016/s0014-5793(01)02702-8 . ПМИД 11532443 .
- ^ Исии М., Курачи Ю. (1 октября 2006 г.). «Мускариновые ацетилхолиновые рецепторы». Текущий фармацевтический дизайн . 12 (28): 3573–81. дои : 10.2174/138161206778522056 . ПМИД 17073660 .
- ^ Лотт Э.Л., Джонс Э.Б. (2020). «Холинергическая токсичность». СтатПерлс . Издательство StatPearls. ПМИД 30969605 .
- ^ Каббани Н., Нордман Дж.К., Коргиат Б.А., Велтри Д.П., Шеху А., Сеймур В.А., Адамс DJ (декабрь 2013 г.). «Связаны ли никотиновые рецепторы ацетилхолина с G-белками?». Биоэссе . 35 (12): 1025–34. doi : 10.1002/bies.201300082 . ПМИД 24185813 . S2CID 9441100 .
- ^ Хендерсон К.Г., Унгар А. (апрель 1978 г.). «Влияние холинергических антагонистов на симпатическую ганглионарную передачу вазомоторных рефлексов от каротидных барорецепторов и хеморецепторов собаки» . Журнал физиологии . 277 (1): 379–385. doi : 10.1113/jphysicalol.1978.sp012278 . ПМЦ 1282395 . ПМИД 206690 .
- ^ Перейти обратно: а б Анвин Н. (март 2005 г.). «Уточненная структура никотинового рецептора ацетилхолина с разрешением 4А». Журнал молекулярной биологии . 346 (4): 967–89. дои : 10.1016/j.jmb.2004.12.031 . ПМИД 15701510 .
- ^ Касио М. (май 2004 г.). «Структура и функция глицинового рецептора и родственных ему никотиноидных рецепторов» . Журнал биологической химии . 279 (19): 19383–6. дои : 10.1074/jbc.R300035200 . ПМИД 15023997 .
- ^ Гиниатуллин Р., Нистри А., Якел Дж.Л. (июль 2005 г.). «Десенсибилизация никотиновых АХ-рецепторов: формирование холинергической сигнализации». Тенденции в нейронауках . 28 (7): 371–8. дои : 10.1016/j.tins.2005.04.009 . ПМИД 15979501 . S2CID 19114228 .
- ^ Перейти обратно: а б Матера, Карло; Папотто, Клаудио; Далланоче, Клелия; Де Амичи, Марко (август 2023 г.). «Достижения в области малых молекул селективных лигандов для гетеромерных никотиновых рецепторов ацетилхолина» . Фармакологические исследования . 194 : 106813. doi : 10.1016/j.phrs.2023.106813 . hdl : 2434/978688 . ПМИД 37302724 .
- ^ Брейц К., ван Дейк В.Дж., Клаассен Р.В., Шуурманс М., ван дер Ост Дж., Смит А.Б., Сиксма Т.К. (май 2001 г.). «Кристаллическая структура ACh-связывающего белка обнаруживает лиганд-связывающий домен никотиновых рецепторов». Природа . 411 (6835): 269–76. Бибкод : 2001Natur.411..269B . дои : 10.1038/35077011 . ПМИД 11357122 . S2CID 4415937 .
- ^ Зуридакис М., Гиастас П., Заркадас Э., Хрони-Царту Д., Брегестовский П., Цартос С.Дж. (ноябрь 2014 г.). «Кристаллические структуры свободных и связанных с антагонистами состояний внеклеточного домена никотинового рецептора α9 человека». Структурная и молекулярная биология природы . 21 (11): 976–80. дои : 10.1038/nsmb.2900 . ПМИД 25282151 . S2CID 30096256 .
- ^ Моралес-Перес CL, Новиелло CM, Хиббс RE (октябрь 2016 г.). «Рентгеновская структура никотинового рецептора α4β2 человека» . Природа . 538 (7625): 411–415. Бибкод : 2016Natur.538..411M . дои : 10.1038/nature19785 . ПМК 5161573 . ПМИД 27698419 .
- ^ Сквайр Л. (2003). Фундаментальная нейронаука (2-е изд.). Амстердам: Акад. Нажимать. п. 1426. ИСБН 978-0-12-660303-3 .
- ^ Перейти обратно: а б Колкухун Д., Сивилотти Л.Г. (июнь 2004 г.). «Функция и структура глициновых рецепторов и некоторых их родственников». Тенденции в нейронауках . 27 (6): 337–44. CiteSeerX 10.1.1.385.3809 . дои : 10.1016/j.tins.2004.04.010 . ПМИД 15165738 . S2CID 19008547 .
- ^ Эйдли диджей (1998). Физиология возбудимых клеток (4-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-0521574150 . ОСЛК 38067558 . [ нужна страница ]
- ^ Бекер Ф., Вебер М., Финк Р.Х., Адамс DJ (сентябрь 2003 г.). «Активация мускариновых и никотиновых рецепторов АХ дифференциально мобилизует Ca2+ во внутрисердечных ганглиозных нейронах крысы». Журнал нейрофизиологии . 90 (3): 1956–64. дои : 10.1152/jn.01079.2002 . ПМИД 12761283 . S2CID 8684707 .
- ^ Вебер М., Мотин Л., Гал С., Бекер Ф., Финк Р.Х., Адамс DJ (январь 2005 г.). «Внутривенные анестетики ингибируют токи, опосредованные никотиновыми ацетилхолиновыми рецепторами, и переходные процессы Ca2+ во внутрисердечных ганглиозных нейронах крысы» . Британский журнал фармакологии . 144 (1): 98–107. дои : 10.1038/sj.bjp.0705942 . ПМК 1575970 . ПМИД 15644873 .
- ^ Мишина М., Такай Т., Имото К., Нода М., Такахаши Т., Нума С. и др. (май 1986 г.). «Молекулярное различие между фетальными и взрослыми формами мышечного рецептора ацетилхолина». Природа . 321 (6068): 406–11. Бибкод : 1986Natur.321..406M . дои : 10.1038/321406a0 . ПМИД 2423878 . S2CID 4356336 .
- ^ Эйнав Т., Филлипс Р. (апрель 2017 г.). «Анализ Монода-Ваймана-Чанге мутантов лиганд-зависимых ионных каналов» . Журнал физической химии Б. 121 (15): 3813–3824. arXiv : 1701.06122 . Бибкод : 2017arXiv170106122E . дои : 10.1021/acs.jpcb.6b12672 . ПМЦ 5551692 . ПМИД 28134524 .
- ^ Левитт М., Сандер С., Стерн П.С. (февраль 1985 г.). «Динамика белка в нормальном режиме: ингибитор трипсина, крамбин, рибонуклеаза и лизоцим». Журнал молекулярной биологии . 181 (3): 423–47. дои : 10.1016/0022-2836(85)90230-х . ПМИД 2580101 .
- ^ Самсон А.О., Левитт М. (апрель 2008 г.). «Механизм ингибирования рецептора ацетилхолина альфа-нейротоксинами, выявленный с помощью динамики в нормальном режиме» . Биохимия . 47 (13): 4065–70. дои : 10.1021/bi702272j . ПМК 2750825 . ПМИД 18327915 .
- ^ Перейти обратно: а б Питчфорд С., Дэй Дж.В., Гордон А., Мокли-Розен Д. (ноябрь 1992 г.). «Десенситизация никотиновых рецепторов ацетилхолина регулируется индуцированным активацией внеклеточным накоплением аденозина» . Журнал неврологии . 12 (11): 4540–4. doi : 10.1523/JNEUROSCI.12-11-04540.1992 . ПМК 6576003 . ПМИД 1331363 .
- ^ Хуганир Р.Л., Грингард П. (февраль 1983 г.). «цАМФ-зависимая протеинкиназа фосфорилирует никотиновый ацетилхолиновый рецептор» . Труды Национальной академии наук Соединенных Штатов Америки . 80 (4): 1130–4. Бибкод : 1983PNAS...80.1130H . дои : 10.1073/pnas.80.4.1130 . ПМЦ 393542 . ПМИД 6302672 .
- ^ Сафран А., Саги-Айзенберг Р., Нейман Д., Фукс С. (август 1987 г.). «Фосфорилирование рецептора ацетилхолина протеинкиназой C и идентификация сайта фосфорилирования внутри дельта-субъединицы рецептора» . Журнал биологической химии . 262 (22): 10506–10. дои : 10.1016/S0021-9258(18)60990-1 . ПМИД 3038884 .
- ^ Хопфилд Дж. Ф., Танк Д. В., Грингард П., Хуганир Р. Л. (декабрь 1988 г.). «Функциональная модуляция никотинового рецептора ацетилхолина путем фосфорилирования тирозина». Природа . 336 (6200): 677–80. Бибкод : 1988Natur.336..677H . дои : 10.1038/336677a0 . ПМИД 3200319 . S2CID 4239105 .
- ^ Баррантес FJ (сентябрь 1978 г.). «Агонист-опосредованные изменения рецептора ацетилхолина в его мембранной среде». Журнал молекулярной биологии . 124 (1): 1–26. дои : 10.1016/0022-2836(78)90144-4 . ПМИД 712829 .
- ^ Херст Р.С., Хайос М., Раггенбасс М., Уолл Т.М., Хигдон Н.Р., Лоусон Дж.А. и др. (апрель 2005 г.). «Новый положительный аллостерический модулятор альфа7-нейронного никотинового ацетилхолинового рецептора: характеристика in vitro и in vivo» . Журнал неврологии . 25 (17): 4396–405. doi : 10.1523/JNEUROSCI.5269-04.2005 . ПМК 6725110 . ПМИД 15858066 .
- ^ Садиг-Этегад С., Мажди А., Талеби М., Махмуди Дж., Бабри С. (май 2015 г.). «Регуляция никотиновых рецепторов ацетилхолина при болезни Альцгеймера: возможная роль шаперонов». Европейский журнал фармакологии . 755 : 34–41. дои : 10.1016/j.ejphar.2015.02.047 . ПМИД 25771456 . S2CID 31929001 .
- ^ Пиччиотто М.Р., Хигли М.Дж., Майнер Ю.С. (октябрь 2012 г.). «Ацетилхолин как нейромодулятор: холинергическая передача сигналов формирует функцию и поведение нервной системы» . Нейрон . 76 (1): 116–29. дои : 10.1016/j.neuron.2012.08.036 . ПМЦ 3466476 . ПМИД 23040810 .
- ^ Воннакотт С. (февраль 1997 г.). «Пресинаптические никотиновые АХ-рецепторы». Тенденции в нейронауках . 20 (2): 92–8. дои : 10.1016/S0166-2236(96)10073-4 . ПМИД 9023878 . S2CID 42215860 .
- ^ Грэм А., Корт Дж.А., Мартин-Руис С.М., Ярос Э., Перри Р., Волсен С.Г. и др. (2002). «Иммуногистохимическая локализация субъединиц никотиновых рецепторов ацетилхолина в мозжечке человека». Нейронаука . 113 (3): 493–507. дои : 10.1016/S0306-4522(02)00223-3 . ПМИД 12150770 . S2CID 39839166 .
- ^ Ле Новер Н., Changeux JP (февраль 1995 г.). «Молекулярная эволюция никотинового рецептора ацетилхолина: пример семейства мультигенов в возбудимых клетках». Журнал молекулярной эволюции . 40 (2): 155–72. Бибкод : 1995JMolE..40..155L . дои : 10.1007/BF00167110 . ПМИД 7699721 . S2CID 2040912 .
- ^ Перейти обратно: а б с д и Импрого М.Р., Скофилд, доктор медицинских наук, Таппер А.Р., Гарднер П.Д. (октябрь 2010 г.). «Кластер генов никотинового ацетилхолинового рецептора CHRNA5/A3/B4: двойная роль в никотиновой зависимости и раке легких» . Прогресс нейробиологии . 92 (2): 212–26. doi : 10.1016/j.pneurobio.2010.05.003 . ПМЦ 2939268 . ПМИД 20685379 .
- ^ Таммимяки А., Хортон В.Дж., Ститцель Дж.А. (октябрь 2011 г.). «Последние достижения в области генных манипуляций и биологии никотиновых рецепторов ацетилхолина» . Биохимическая фармакология . 82 (8): 808–19. дои : 10.1016/j.bcp.2011.06.014 . ПМК 3162071 . ПМИД 21704022 .
- ^ Грэм А., Корт Дж.А., Мартин-Руис С.М., Ярос Э., Перри Р., Волсен С.Г. и др. (сентябрь 2002 г.). «Иммуногистохимическая локализация субъединиц никотиновых рецепторов ацетилхолина в мозжечке человека». Нейронаука . 113 (3): 493–507. дои : 10.1016/S0306-4522(02)00223-3 . ПМИД 12150770 . S2CID 39839166 .
- ^ Changeux JP (июнь 2010 г.). «Никотиновая зависимость и никотиновые рецепторы: уроки генетически модифицированных мышей». Обзоры природы. Нейронаука . 11 (6): 389–401. дои : 10.1038/nrn2849 . ПМИД 20485364 . S2CID 661315 .
- ^ Диабасана, З; Перотин, Дж. М.; Бельгачеми, Р; Ансель, Дж; Мулетт, П; Делепин, Дж; Госсет, П; Маскос, У; Полетт, М; Десли, Дж; Дормой, В. (2020). «Атлас субъединиц никотиновых рецепторов в легких взрослого человека» . Межд. Дж. Мол. Наука . 21 (20): 7446. doi : 10.3390/ijms21207446 . ПМЦ 7588933 . ПМИД 33050277 .
- ^ Перейти обратно: а б с д и Гринбаум Л., Лерер Б. (октябрь 2009 г.). «Дифференциальный вклад генетических вариаций в множественных никотиновых холинергических рецепторах мозга в никотиновую зависимость: недавний прогресс и возникающие открытые вопросы» . Молекулярная психиатрия . 14 (10): 912–45. дои : 10.1038/mp.2009.59 . ПМИД 19564872 .
- ^ Гаринг Л.К., Роджерс С.В. (январь 2006 г.). «Нейрональная экспрессия никотинового ацетилхолинового рецептора и его функция на ненейрональных клетках» . Журнал AAPS . 7 (4): Е885-94. дои : 10.1208/aapsj070486 . ПМК 2750958 . ПМИД 16594641 .
- ^ Перейти обратно: а б Каменс Х.М., Корли Р.П., Ричмонд П.А., Дарлингтон Т.М., Доуэлл Р., Хопфер С.Дж. и др. (сентябрь 2016 г.). «Доказательства связи между низкочастотными вариантами CHRNA6/CHRNB3 и антисоциальной лекарственной зависимостью» . Генетика поведения . 46 (5): 693–704. дои : 10.1007/s10519-016-9792-4 . ПМЦ 4975622 . ПМИД 27085880 .
- ^ Грэди С.Р., Салминен О., Лаверти Д.С., Уайтакер П., Макинтош Дж.М., Коллинз А.С., Маркс М.Дж. (октябрь 2007 г.). «Подтипы никотиновых ацетилхолиновых рецепторов на дофаминергических окончаниях полосатого тела мыши» . Биохимическая фармакология . 74 (8): 1235–46. дои : 10.1016/j.bcp.2007.07.032 . ПМЦ 2735219 . ПМИД 17825262 .
- ^ Перейти обратно: а б с д и Стейнлайн ОК, Бертран Д. (ноябрь 2008 г.). «Нейрональные никотиновые ацетилхолиновые рецепторы: от генетического анализа к неврологическим заболеваниям». Биохимическая фармакология . 76 (10): 1175–83. дои : 10.1016/j.bcp.2008.07.012 . ПМИД 18691557 .
- ^ Бертран Д., Элмсли Ф., Хьюз Э., Траунс Дж., Сандер Т., Бертран С., Стейнлайн ОК (декабрь 2005 г.). «Мутация CHRNB2 I312M связана с эпилепсией и выраженным дефицитом памяти». Нейробиология болезней . 20 (3): 799–804. дои : 10.1016/j.nbd.2005.05.013 . ПМИД 15964197 . S2CID 29811931 .
- ^ Бриз Ч.Р., Ли М.Дж., Адамс К.Э., Салливан Б., Логель Дж., Гиллен К.М. и др. (октябрь 2000 г.). «Аномальная регуляция никотиновых рецепторов с высоким сродством у больных шизофренией» . Нейропсихофармакология . 23 (4): 351–64. дои : 10.1016/S0893-133X(00)00121-4 . ПМИД 10989262 .
- ^ Маклин С.Л., Грейсон Б., Идрис Н.Ф., Лесаж А.С., Пембертон DJ, Маки С., Нил Дж.К. (апрель 2011 г.). «Активация никотиновых рецепторов α7 улучшает вызванный фенциклидином дефицит когнитивных задач у крыс: значение для терапии когнитивной дисфункции при шизофрении». Европейская нейропсихофармакология . 21 (4): 333–43. дои : 10.1016/j.euroneuro.2010.06.003 . hdl : 10454/8464 . ПМИД 20630711 . S2CID 41306366 .
- ^ Перейти обратно: а б с д Позвонил HP (2003). Фармакология (5-е изд.). Эдинбург: Черчилль Ливингстон. ISBN 978-0-443-07145-4 . [ нужна страница ]
- ^ Перейти обратно: а б Neurosci.pharm - MBC 3320 Ацетилхолин. Архивировано 27 декабря 2007 г. в Wayback Machine.
- ^ Сартер М. (август 2015 г.). «Поведенчески-когнитивные цели для улучшения холинергического действия» . Современное мнение в области поведенческих наук . 4 : 22–26. дои : 10.1016/j.cobeha.2015.01.004 . ПМК 5466806 . ПМИД 28607947 .
- ^ Ву Дж, Гао М, Шен JX, Ши WX, Остер А.М., Гуткин Б.С. (октябрь 2013 г.). «Корковый контроль функции ВТА и влияние на никотиновое вознаграждение». Биохимическая фармакология . 86 (8): 1173–80. дои : 10.1016/j.bcp.2013.07.013 . ПМИД 23933294 .
- ^ «Никотин: Биологическая активность» . Руководство IUPHAR/BPS по фармакологии . Международный союз фундаментальной и клинической фармакологии . Проверено 7 февраля 2016 г.
K заключается в следующем; α 2 β 4 =9900 нМ [5], α 3 β 2 =14 нМ [1], α 3 β 4 =187 нМ [1], α 4 β 2 =1 нМ [4,6]. Из-за гетерогенности каналов nACh мы не отметили первичную мишень препарата для никотина, хотя сообщается, что α 4 β 2 является преобладающим подтипом с высоким сродством в мозге, который опосредует никотиновую зависимость [2-3].
- ^ Левин Э.Д. (май 2012 г.). «α7-никотиновые рецепторы и познание». Текущие цели по борьбе с наркотиками . 13 (5): 602–6. дои : 10.2174/138945012800398937 . ПМИД 22300026 .
- ^ Садиг-Этегад С., Махмуди Дж., Бабри С., Талеби М. (ноябрь 2015 г.). «Влияние активации альфа-7 никотинового ацетилхолинового рецептора на нарушение памяти узнавания, вызванное бета-амилоидом. Возможная роль нейрососудистой функции» . Acta Cirurgica Brasileira . 30 (11): 736–42. дои : 10.1590/S0102-865020150110000003 . ПМИД 26647792 .
- ^ Ван Дж, Лу З, Фу Икс, Чжан Д, Ю Л, Ли Н и др. (май 2017 г.). «Сигнальный путь никотинового рецептора альфа-7 участвует в нейрогенезе, индуцированном ChAT-положительными нейронами в субвентрикулярной зоне» . Трансляционное исследование инсульта . 8 (5): 484–493. дои : 10.1007/s12975-017-0541-7 . ПМК 5704989 . ПМИД 28551702 .
- ^ Ли Дж., Кук Дж.П. (ноябрь 2012 г.). «Никотин и патологический ангиогенез» . Науки о жизни . 91 (21–22): 1058–64. дои : 10.1016/j.lfs.2012.06.032 . ПМЦ 3695741 . ПМИД 22796717 .
- ^ Джайн Дж., Джеймс Э.А. (октябрь 2013 г.). «Передача сигналов никотина и прогрессирование хронической болезни почек у курильщиков» . Биохимическая фармакология . 86 (8): 1215–23. дои : 10.1016/j.bcp.2013.07.014 . ПМЦ 3838879 . ПМИД 23892062 .
- ^ Михалак КБ, Кэрролл Ф.И., Лютье К.В. (сентябрь 2006 г.). «Варениклин является частичным агонистом альфа4бета2 и полным агонистом альфа7-нейронных никотиновых рецепторов». Молекулярная фармакология . 70 (3): 801–5. дои : 10.1124/моль.106.025130 . ПМИД 16766716 . S2CID 14562170 .
Внешние ссылки
[ редактировать ] СМИ, связанные с никотиновыми рецепторами ацетилхолина , на Викискладе?
СМИ, связанные с никотиновыми рецепторами ацетилхолина , на Викискладе? - Расчетное пространственное положение никотинового рецептора ацетилхолина в липидном бислое