Змеиный яд

Змеиный яд — высокотоксичная слюна. [1] содержат зоотоксины , способствующие обездвиживанию и перевариванию добычи . Это также обеспечивает защиту от угроз. Змеиный яд обычно вводится через уникальные клыки во время укуса , хотя некоторые виды также способны выплевывать яд . [2]
Ядовитые железы , выделяющие зоотоксины, представляют собой модификацию околоушных слюнных желез , встречающихся у других позвоночных , и обычно расположены по обе стороны головы, ниже и позади глаза и заключены в мышечную оболочку. Яд хранится в больших железах, называемых альвеолами, а затем по протоку попадает к основанию желобчатых или трубчатых клыков, через которые он выбрасывается. [3] [4]
Яд содержит более 20 различных соединений, в основном белков и полипептидов . [3] [5] Сложная смесь белков, ферментов и других веществ обладает токсическими и смертоносными свойствами. [2] Яд служит для обездвиживания добычи. [6] Ферменты яда играют важную роль в переваривании добычи. [4] и различные другие вещества ответственны за важные, но несмертельные биологические эффекты. [2] Некоторые белки змеиного яда оказывают весьма специфическое воздействие на различные биологические функции, включая свертывание крови, регуляцию артериального давления и передачу нервных или мышечных импульсов. Эти яды были изучены и разработаны для использования в качестве фармакологических или диагностических инструментов и даже лекарств. [2] [5]
Химия
[ редактировать ]Белки составляют 90–95% сухой массы яда и отвечают почти за все его биологические эффекты. [5] [7] Сотни, даже тысячи белков, обнаруженных в яде, включают токсины, в частности нейротоксины , а также нетоксичные белки (которые также обладают фармакологическими свойствами) и множество ферментов, особенно гидролитических . [2] Ферменты ( молекулярная масса 13-150 кДа) составляют 80-90% ядов гадюк и 25-70% ядов элапид , в том числе пищеварительные гидролазы , оксидаза L-аминокислот , фосфолипазы , тромбин -подобный прокоагулянт и калликреин -подобный сериновые протеазы и металлопротеиназы (геморрагины), повреждающие эндотелий сосудов . Полипептидные токсины (молекулярная масса 5–10 кДа) включают цитотоксины , кардиотоксины и постсинаптические нейротоксины (такие как α-бунгаротоксин и α-кобратоксин ), которые связываются с рецепторами ацетилхолина в нервно-мышечных соединениях. К соединениям с низкой молекулярной массой (до 1,5 кДа) относятся металлы, пептиды, липиды, нуклеозиды , углеводы, амины и олигопептиды , которые ингибируют ангиотензинпревращающий фермент (АПФ) и потенцируют брадикинин (БПП). Меж- и внутривидовая изменчивость химического состава ядов носит географический и онтогенный характер. [3] Фосфодиэстераза влияет на сердечную систему жертвы, главным образом, для снижения кровяного давления . Фосфолипаза А2 вызывает гемолиз лизируя фосфолипидные клеточные мембраны эритроцитов , . [8] аминокислот оксидазы и протеазы Для пищеварения используются . Оксидаза аминокислот также запускает некоторые другие ферменты и отвечает за желтый цвет яда некоторых видов. Гиалуронидаза увеличивает проницаемость тканей, ускоряя всасывание других ферментов в ткани. Яды некоторых змей содержат фасцикулины , например, мамбы ( Dendroaspis ), которые ингибируют холинэстеразу , заставляя жертву терять контроль над мышцами. [9]
| Тип | Имя | Источник |
|---|---|---|
| Оксидоредуктазы | лактатдегидрогеназа | Элапиды |
| Оксидаза L-аминокислот | Все виды | |
| Каталаза | Все виды | |
| Трансферазы | Аланинаминотрансфераза | |
| Гидролазы | Фосфолипаза А2 | Все виды |
| Лизофосфолипаза | Elapidae, Viperidae | |
| Ацетилхолинэстераза | Элапиды | |
| Щелочная фосфатаза | Ботропс атрокс | |
| Кислая фосфатаза | Дейнагистродон острый | |
| 5'-нуклеотидаза | Все виды | |
| фосфодиэстераза | Все виды | |
| Дезоксирибонуклеаза | Все виды | |
| Рибонуклеаза 1 | Все виды | |
| Аденозинтрифосфатаза | Все виды | |
| Амилаза | Все виды | |
| Гиалуронидаза | Все виды | |
| НАД-нуклеотидаза | Все виды | |
| Кининогеназа | Гадюки | |
| фактора X Активатор | Viperidae, Crotalinae | |
| Гепариназа | Кроталины | |
| α-фибриногеназа | Viperidae, Crotalinae | |
| β-фибриногеназа | Viperidae, Crotalinae | |
| α-β-фибриногеназа | Битис Габоника | |
| Фибринолитический фермент | Кроталины | |
| Активатор протромбина | Кроталины | |
| Коллагеназа | Гадюки | |
| Эластаза | Гадюки | |
| Лиазы | Глюкозаминатаммиаклиаза |
Змеиные токсины сильно различаются по своим функциям. Двумя широкими классами токсинов, обнаруженных в ядах змей, являются нейротоксины (в основном обнаруженные у элапид) и гемотоксины (в основном обнаруженные у гадюк). Однако встречаются и исключения — яд черношейной плюющейся кобры ( Naja nigricollis ), элапида, состоит в основном из цитотоксинов , тогда как яд гремучей змеи Мохаве ( Crotalus scutulatus ), гадюки, преимущественно нейротоксичен. И элапиды, и гадюки могут переносить множество других типов токсинов.
| α-нейротоксины | α-бунгаротоксин , α-токсин, эрабутоксин , кобратоксин |
|---|---|
| β-нейротоксины ( PLA2 ) | β-бунгаротоксин , нотексин, аммодитоксин , кротоксин , тайпоксин |
| κ-нейротоксины | Каппа-бунгаротоксин |
| Дендротоксины ( Куниц ) | Дендротоксин , токсины I и К; возможно, β-бунгаротоксина цепь B |
| Кардиотоксины | Naja nigricollis Y-токсин , кардиотоксин III (также известный как цитотоксины) |
| Миотоксины | Миотоксин -а, кротамин |
| Сарафотоксины | Сарафотоксины a, b и c |
| Геморрагины (металлопротеазы) | Мукролизин , Атролизины , Акутолизины и др. [11] |
| Гемотоксины (сериновая протеаза) | Веномбин А |
Токсины
[ редактировать ]В этом разделе отсутствует информация о том, как эволюционные/структурные классификации соответствуют функциональным классификациям. ( август 2021 г. ) |
Нейротоксины
[ редактировать ]
Начало нового нервного импульса происходит следующим образом:
- Обмен ионов (заряженных атомов) через мембрану нервной клетки посылает деполяризующий ток к концу нервной клетки (концу клетки).
- Когда деполяризующий ток достигает окончания нервных клеток, нейромедиатор ацетилхолин (АХ), который содержится в везикулах , высвобождается в пространство между двумя нервами (синапс). Он движется через синапс к постсинаптическим рецепторам.
- АХ связывается с рецепторами и передает сигнал клетке-мишени, которая через короткое время разрушается ацетилхолинэстеразой.
Фасцикулины
[ редактировать ]- Эти токсины атакуют холинергические нейроны (те, которые используют АХ в качестве передатчика), разрушая ацетилхолинэстеразу (АХЭ). Следовательно, АХ не может расщепляться и остается в рецепторе. Это вызывает тетанию (непроизвольное сокращение мышц), что может привести к смерти. Токсины получили название фасцикулины, поскольку после инъекции мышам они вызывают тяжелые, генерализованные и длительные (5-7 часов) фасцикуляции (быстрые мышечные сокращения).
- Пример змей: встречается главным образом в яде мамб ( виды Dendroaspis ) и некоторых гремучих змей ( вид Crotalus ).
Дендротоксины
[ редактировать ]- Дендротоксины ингибируют нейротрансмиссию, блокируя обмен положительных и отрицательных ионов через мембрану нейрона, что приводит к прекращению нервного импульса, тем самым парализуя нервы.
- Пример змеи: мамба
α-нейротоксины
[ редактировать ]- Альфа-нейротоксины представляют собой большую группу; идентифицировано и секвенировано более 100 постсинаптических нейротоксинов. [12] α-нейротоксины атакуют никотиновые ацетилхолиновые рецепторы холинергических нейронов. Они имитируют форму молекулы ацетилхолина и поэтому встраиваются в рецепторы, где блокируют поток АХ, что приводит к ощущению онемения и паралича.
- Примеры змей: королевская кобра ( Ophiophagus hannah ) (известная как ханнатоксин, содержащий α-нейротоксины), [13] морские змеи (Hydrophiinae) (известные как эрабутоксин), многополосный крайт ( Bungarus multicinctu s) (известный как α-бунгаротоксин ) и кобры ( Naja spp.) (известные как кобратоксин )
Цитотоксины
[ редактировать ]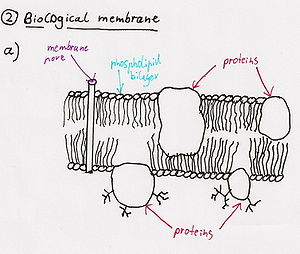

Фосфолипазы
[ редактировать ]- Фосфолипаза — это фермент, который превращает молекулу фосфолипида в лизофосфолипид (мыло) → новая молекула притягивает и связывает жир и разрывает клеточные мембраны. Фосфолипаза А2 — один из специфических типов фосфолипаз, обнаруженных в змеином яде.
- Пример змеи: Окинавский хабу ( Trimeresurus flavoviridis ).
Кардиотоксины/цитотоксины
[ редактировать ]- Кардиотоксины – это компоненты, которые особенно токсичны для сердца. Они связываются с определенными участками на поверхности мышечных клеток и вызывают деполяризацию → токсин предотвращает сокращение мышц. Эти токсины могут привести к тому, что сердце начнет биться нерегулярно или перестанет биться, что приведет к смерти. Примером может служить трехпалый кардиотоксин III китайской кобры , представитель семейства коротких трехпалых ( ИнтерПро : IPR003572 ).
- Пример змей: мамбы и некоторые наджа . виды
Гемотоксины
[ редактировать ]- Гемотоксины вызывают гемолиз, разрушение эритроцитов или индуцируют свертывание крови (свертывание крови, например мукроцетин ). Общее семейство гемотоксинов включает металлопротеиназы змеиного яда, такие как мукролизин . [11] [14]
- Примеры змей: большинство гадюк и многие виды кобр : Тропическая гремучая змея Crotalus durissus вырабатывает конвульксин , коагулянт. [15]
Миотоксины
[ редактировать ]
Миотоксины — это небольшие основные пептиды , обнаруженные у гремучей змеи. [16] [17] и ящерица (например, мексиканская ящерица из бисера ) [18] яды. Это связано с неферментативным механизмом, который приводит к тяжелому скелетных мышц некрозу . Эти пептиды действуют очень быстро, вызывая мгновенный паралич, предотвращающий добычи побег , и, в конечном итоге, смерть из-за паралича диафрагмы .
Первым миотоксином, который был идентифицирован и выделен, был кротамин , открытый в 1950-х годах бразильским ученым Хосе Моурой Гонсалвесом из яда тропической южноамериканской гремучей змеи Crotalus durissus terrificus . Его биологическое действие, молекулярная структура и ген, ответственный за его синтез, были выяснены за последние два десятилетия.
Определение токсичности яда (LD 50 )
[ редактировать ]В этом разделе отсутствует информация о том, почему разведение альбумина работает лучше. ( август 2021 г. ) |
Токсичность змеиного яда оценивается с помощью токсикологического теста, называемого средней летальной дозой , летальная доза 50% (сокращенно ЛД 50 ), который определяет концентрацию токсина, необходимую для уничтожения половины членов тестируемой популяции. Эффективность яда дикой змеи значительно варьируется из-за различных влияний, таких как биофизическая среда , физиологический статус, экологические переменные , генетические вариации ( адаптивные или случайные), а также другие молекулярные и экологические эволюционные факторы. [ нужна ссылка ] Это справедливо даже для представителей одного вида. Такая изменчивость меньше в неволе в лабораторных условиях, хотя ее невозможно устранить. Однако исследования по определению эффективности змеиного яда должны быть разработаны так, чтобы свести к минимуму изменчивость.
С этой целью было разработано несколько методик. Один из подходов заключается в использовании 0,1% бычьего сывороточного альбумина (также известного как «фракция V» в процессе Кона ) в качестве разбавителя при определении значений LD 50 . Это приводит к более точному и последовательному определению LD 50 , чем при использовании 0,1% физиологического раствора в качестве разбавителя. Например, фракция V дает около 95% очищенного альбумина (высушенный сырой яд). Солевой раствор в качестве разбавителя постоянно дает сильно различающиеся результаты LD 50 практически для всех ядовитых змей. Это приводит к непредсказуемому изменению чистоты осадка (35-60%). [19] Фракция V структурно стабильна, поскольку имеет семнадцать дисульфидных связей ; он уникален тем, что имеет самую высокую растворимость и самую низкую изоэлектрическую точку среди основных белков плазмы. Это делает его последней фракцией, которая выпадает в осадок из раствора. Альбумин бычьей сыворотки находится во фракции V. Осаждение альбумина осуществляется путем снижения pH до 4,8, близкого к pH белков, и поддержания концентрации этанола на уровне 40% при концентрации белка 1%. Таким образом, в пятой фракции остается только 1% исходной плазмы. [20]
Когда конечной целью обработки плазмы является получение очищенного компонента плазмы для инъекций или переливания , этот компонент плазмы должен быть очень чистым. Первый практический крупномасштабный метод фракционирования плазмы крови был разработан Эдвином Дж. Коном во время Второй мировой войны . он известен как процесс Кона (или метод Кона). Этот процесс также известен как холодное фракционирование этанола, поскольку он включает постепенное увеличение концентрации в растворе этанола при 5 °C и 3 °C. [21] Процесс Кона использует различия в свойствах белков плазмы, в частности, высокую растворимость и низкий пи альбумина. По мере поэтапного увеличения концентрации этанола от 0 до 40% pH снижается от нейтрального (pH ~ 7) примерно до 4,8, что близко к pI альбумина. [21] На каждом этапе белки осаждаются из раствора и удаляются. Конечный осадок представляет собой очищенный альбумин. Существует несколько вариантов этого процесса, включая адаптированный метод Ничмана и Кистлера, который использует меньше шагов и заменяет центрифугирование и замораживание в массе фильтрацией и диафильтрацией. [21] [22] Некоторые новые методы очистки альбумина добавляют дополнительные этапы очистки к процессу Кона и его вариантам. Хроматографическая обработка альбумина возникла в 1980-х годах, однако широкое распространение она получила лишь позднее из-за нехватки крупномасштабного хроматографического оборудования. Методы, включающие хроматографию, обычно начинаются с криообедненной плазмы, подвергаемой замене буфера посредством диафильтрации или буферно-обменной хроматографии, чтобы подготовить плазму для следующих ионообменной хроматографии стадий . После ионного обмена обычно происходят этапы очистки и замены буфера. [21]
Однако хроматографические методы начали внедряться в 1980-х годах. [ нужна ссылка ] Развитие продолжалось между моментом, когда фракционирование Кона начало появляться в 1946 году, и появлением хроматографии в 1983 году. В 1962 году процесс Кистлера и Нишмана был создан как побочный продукт процесса Кона. В 1990-х годах были созданы процессы Zenalb и CSL Albumex, которые включали хроматографию с некоторыми вариациями. Общий подход к использованию хроматографии для фракционирования плазмы на альбумин заключается в следующем: выделение супернатанта I, делипидирование, анионообменная хроматография , катионообменная хроматография и гель-фильтрационная хроматография. Извлеченный очищенный материал смешивают с комбинациями октаноата натрия и N-ацетилтриптофаната натрия, а затем подвергают процедурам вирусной инактивации, включая пастеризацию при 60 °C. Это более эффективная альтернатива, чем процесс Кона, потому что:
- нужна была плавная автоматизация и относительно недорогой завод,
- легче стерилизовать оборудование и поддерживать хорошую производственную среду
- хроматографические процессы менее повреждают белок альбумин
- может быть достигнут более успешный результат по альбумину. [ нужна ссылка ]
По сравнению с процессом Кона чистота альбумина увеличилась примерно с 95% до 98% при использовании хроматографии, а выход увеличился примерно с 65% до 85%. Небольшое процентное увеличение имеет значение в отношении таких чувствительных измерений, как чистота. Большой недостаток связан с экономикой. Хотя метод оказался эффективным, приобретение необходимого оборудования было затруднено. Необходима большая техника, и в течение длительного времени отсутствие оборудования ограничивало его широкое использование. [ нужна ссылка ]
Эволюция
[ редактировать ]Яд появился среди всех токсикофер только один раз , около 170 миллионов лет назад, а затем превратился в огромное разнообразие ядов, наблюдаемое сегодня. [23] Первоначальный яд токсикоферанов представлял собой очень простой набор белков, собранных в паре желез. Впоследствии этот набор белков эволюционировал независимо в различных линиях токсикоферан, включая Serpentes , Anguimorpha и Iguania . [24] Некоторые линии змей с тех пор утратили способность производить яд, часто из-за изменения диеты или изменения тактики хищничества. [23] Кроме того, сила и состав яда изменились из-за изменений в добыче некоторых видов змей. Например, яд мраморной морской змеи ( Aipysurus eydouxii ) стал значительно менее токсичным после того, как рацион этого вида изменился с рыбы на строго рыбью икру. [23] Считается, что эволюция яда ответственна за огромное распространение змей по всему земному шару. [23] [25]
Механизмом эволюции в большинстве случаев было дупликация генов в тканях, не связанных с ядом. [24] Ранее существовавшие белки слюны являются вероятными предками большинства генов ядовитых токсинов. [26] Экспрессия нового белка в ядовитой железе последовала за дупликацией. [24] Затем продолжился естественный отбор адаптивных признаков по модели рождения и смерти, где за дупликацией следует функциональная диверсификация, приводящая к созданию структурно родственных белков, имеющих несколько разные функции. [23] [24] [27] Изучение эволюции яда было приоритетом для ученых с точки зрения научных исследований из-за медицинской значимости змеиного яда, с точки зрения создания противоядия и исследований рака. Очень полезно знать больше о составе яда и о том, как он потенциально может развиваться. Были тщательно изучены три основных фактора, влияющих на эволюцию яда: хищники змей, устойчивые к змеиному яду, добыча, участвующая в эволюционной гонке вооружений со змеями, и специфические диеты, влияющие на внутривидовую эволюцию яда. [23] [28] Яды продолжают развиваться как специфические токсины и модифицируются, чтобы нацеливаться на конкретную добычу, а у некоторых видов токсины различаются в зависимости от диеты. [29] [30]
Быструю эволюцию яда также можно объяснить гонкой вооружений между молекулами, нацеленными на яд, у устойчивых хищников, таких как опоссум , и змеиным ядом, который нацелен на эти молекулы. Ученые провели эксперименты на опоссумах и обнаружили, что многочисленные исследования показали замену на молчащие замены в гене фактора фон Виллебранда ( vWf ), который кодирует гемостатический белок крови, нацеленный на яд. Считается, что эти замены ослабляют связь между vWf и лигандом токсичного змеиного яда (ботроцетином), что изменяет суммарный заряд и гидрофобность. Эти результаты имеют важное значение для эволюции яда, поскольку это первое свидетельство быстрой эволюции молекулы, нацеленной на яд. Это показывает, что эволюционная гонка вооружений может происходить в оборонительных целях. Альтернативные гипотезы предполагают, что эволюция яда обусловлена трофической адаптацией, тогда как эти ученые полагают, что в этом случае отбор будет происходить по признакам, которые помогают выживанию добычи с точки зрения эволюции яда, а не успеха хищничества. Некоторые другие хищники гадюки (мангусты и ежи) демонстрируют тот же тип взаимоотношений между змеями, что помогает поддержать гипотезу о том, что яд имеет очень сильную защитную роль наряду с трофической ролью. Это, в свою очередь, подтверждает идею о том, что хищничество змей может быть гонкой вооружений, приводящей к эволюции змеиного яда. [31]
Некоторые из различных адаптаций, возникающих в результате этого процесса, включают яд, более токсичный для конкретной добычи в нескольких линиях. [30] [32] [33] белки, которые предварительно переваривают добычу, [34] а также метод выслеживания добычи после укуса. [35] Хотя функция яда эволюционировала и стала специфичной для класса добычи (например, особые коагуляционные эффекты), [36] развитие широких токсикологических эффектов (например, нейротоксичности или коагулотоксичности), по-видимому, не сильно зависит от типа добычи. [37] Когда-то считалось, что наличие пищеварительных ферментов в змеином яде является адаптацией, способствующей пищеварению. Однако исследования западной гремучей змеи ( Crotalus atrox ), змеи с высокопротеолитическим ядом , показывают, что яд не влияет на время, необходимое для прохождения пищи через кишечник . [38] Эти различные адаптации яда также привели к серьезным спорам об определении яда и ядовитых змей. [23]
Инъекция
[ редактировать ]Змеи
[ редактировать ]У гадюк , имеющих наиболее развитый аппарат для доставки яда, ядовитая железа очень велика и окружена жевательной или височной мышцей , состоящей из двух полос: верхняя выходит из-за глаза, нижняя - отходит от железы. к нижней челюсти. Проток переносит яд из железы в клык. У гадюк и элапид эта борозда полностью закрыта, образуя подкожную игольчатую трубку. У других видов бороздки не прикрыты или прикрыты лишь частично. От переднего конца железы проток проходит ниже глаза и над верхнечелюстной костью к базальному отверстию ядовитого клыка, покрытого толстой складкой слизистой оболочки . С помощью подвижной верхнечелюстной кости, шарнирно прикрепленной к предлобной кости и соединенной с поперечной костью, которая выдвигается вперед мышцами, приводящимися в действие при открывании рта, клык поднимается и яд выбрасывается через дистальное отверстие. Когда змея кусает, челюсти закрываются, а мышцы, окружающие железу, сокращаются, в результате чего яд выбрасывается через клыки.
Элапиды
[ редактировать ]У протероглифных элапидов клыки трубчатые, но короткие и не обладают подвижностью, наблюдаемой у гадюк.
Колубриды
[ редактировать ]Опистоглифные колубриды имеют увеличенные зубы с бороздками, расположенные на заднем конце верхней челюсти , где небольшая задняя часть верхней губной или слюнной железы выделяет яд.
Механика прикуса
[ редактировать ]
Несколько родов, в том числе азиатские коралловые змеи ( Calliophis ), роющие аспиды ( Atractaspis ) и ночные гадюки ( Causus ), отличаются исключительно длинными ядовитыми железами, простирающимися вдоль каждой стороны тела, а в некоторых случаях простирающимися назад до самого конца тела. сердце. Вместо мышц височной области, служащих для выдавливания яда в проток, это действие выполняют мышцы боковых сторон тела.
Среди змей наблюдаются значительные различия в кусающем поведении. При укусе змеи-гадюки часто наносят быстрый удар, выделяя яд, когда клыки проникают в кожу, а затем сразу же выпускают. Альтернативно, как и в случае с реакцией на кормление, некоторые гадюки (например, Lachesis ) кусают и держатся. Протероглиф опистоглиф или . может сомкнуть челюсти и сильно кусать или жевать в течение значительного времени
Различия в длине клыков у разных ядовитых змей, вероятно, связаны с эволюцией разных стратегий нанесения ударов. [39] Кроме того, было показано, что клыки разных видов ядовитых змей имеют разные размеры и форму в зависимости от биомеханических свойств змеиной добычи. [40]
Механика плевания
[ редактировать ]Плюющиеся кобры родов Naja и Hemachatus при раздражении или угрозе могут выбрасывать струи или брызги яда на расстояние от 1,2 метра (4 фута) до 2,4 метра (8 футов). Клыки этих змей были модифицированы для того, чтобы они могли плеваться; внутри клыков канал изгибается на 90° к нижней передней части клыка. Плеваки могут плюнуть несколько раз и при этом нанести смертельный укус.
Плевок – это всего лишь защитная реакция. Змеи, как правило, целятся в глаза предполагаемой угрозы. Прямое попадание может вызвать временный шок и слепоту из-за воспаления роговицы . и конъюнктивы тяжелого Хотя обычно никаких серьезных симптомов не возникает, если яд немедленно смыть большим количеством воды, слепота может стать постоянной, если ее не лечить. Кратковременный контакт с кожей не представляет непосредственной опасности, но открытые раны могут быть переносчиками отравления.
Физиологические эффекты
[ редактировать ]Эта статья нуждается в дополнительных цитатах для проверки . ( август 2021 г. ) |
Четыре различных типа яда действуют на организм по-разному:
- Протеолитический яд разрушает молекулярное окружение, в том числе и в месте укуса.
- Гемотоксический яд действует на сердечно-сосудистую систему, в том числе на сердце и кровь.
- Нейротоксический яд действует на нервную систему, в том числе на головной мозг.
- Цитотоксический яд оказывает локализованное действие в месте укуса.
Протероглифные змеи
[ редактировать ]Действие яда протероглифных змей ( морских змей , крайтов , мамб , черных змей , тигровых змей , смертоносных змей ) преимущественно воздействует на нервную систему , при этом быстро возникает паралич дыхания за счет попадания яда в центральный нервный механизм, который контролирует дыхание; Боль и местный отек, возникающие после укуса, обычно не являются сильными. Укус всех протероглифических элапидов, даже самых маленьких и нежных, таких как коралловые змеи , насколько известно, смертелен для человека. Однако сохранились некоторые слабоядовитые элапиды, например хохлатые змеи ( Parasuta ), хохлатые змеи ( Vermicella ) и др.
Змеи
[ редактировать ]Яд гадюки ( гадюки Рассела , чешуйчатой гадюки , бушмастера и гремучей змеи ) действует больше на сосудистую систему, вызывая свертывание крови и свертывание легочных артерий; его действие на нервную систему невелико, ни одна отдельная группа нервных клеток не выделяется, и влияние на дыхание не столь прямое; влияние на кровообращение объясняет большую депрессию, которая является симптомом отравления гадюкой. Боль в ране сильная, за ней быстро следует отек и изменение цвета. Симптомы, вызываемые укусом европейской гадюки, описаны Мартином и Лэмбом: [41]
Укус немедленно сопровождается местной болью жгучего характера; конечность вскоре опухает и обесцвечивается, и через один-три часа сильная прострация, сопровождающаяся рвотой и часто поносом наступает . Обычно наблюдается холодный липкий пот. Пульс становится крайне слабым, легкая одышка может наблюдаться и беспокойство. В тяжелых случаях, встречающихся преимущественно у детей, пульс может стать незаметным, а конечности похолодеть; больной может впасть в кому . Через двенадцать-двадцать четыре часа эти тяжелые конституциональные симптомы обычно проходят; но тем временем отек и изменение цвета сильно распространились. Конечность становится флегмонизированной и иногда нагнаивается. Выздоровление в течение нескольких дней обычно наступает внезапно, но смерть может наступить в результате тяжелой депрессии или побочных эффектов нагноения . В последней главе настоящего Введения упоминается, что случаи смерти как у взрослых, так и у детей нередки в некоторых частях континента.
Гадюки сильно различаются между собой по токсичности ядов. Некоторые из них, такие как индийская гадюка Рассела ( Daboia russelli ) и чешуйчатая гадюка ( E. carinatus ); американские гремучие змеи ( Crotalus вид ), бушмастеры (вид Lachesis ) и копьеголовки ( Bothrops вид ); а африканские змеи ( Bitis spp.), ночные змеи ( Causus spp.) и рогатые гадюки ( Cerastes spp.) вызывают летальные исходы, если не приступить к немедленному лечению. Укус более крупных европейских гадюк может быть очень опасным и может привести к летальному исходу, особенно у детей, по крайней мере, в более жарких частях континента; в то время как маленькая луговая гадюка ( Vipera ursinii ), которая почти никогда не кусает, если с ней грубо обращаться, по-видимому, не обладает очень опасным ядом и, хотя и очень распространена в некоторых частях Австрии и Венгрии , неизвестно, когда-либо вызывала серьезная авария.
Опистоглифные колубриды
[ редактировать ]Биологи давно знали, что у некоторых змей есть задние клыки, «низшие» механизмы инъекции яда, которые могут обездвижить добычу; хотя до 1957 года было зарегистрировано несколько смертельных случаев, возможность того, что такие змеи были смертельными для человека, казалась весьма маловероятной. Смерть двух выдающихся герпетологов, Роберта Мертенса и Карла Шмидта , от укусов африканских колубрид изменила эту оценку, а недавние события показали, что некоторые другие виды заднезубых змей имеют яды, потенциально смертельные для крупных позвоночных.
Яды бумсланга ( Dispholidus typus ) и ветчатой змеи ( Thelotornis spp.) токсичны для клеток крови и разжижают кровь (гемотоксичные, геморрагические). Ранние симптомы включают головные боли, тошноту, диарею, летаргию, умственную дезориентацию, синяки и кровотечения в этом месте и во всех отверстиях тела. Обескровливание — основная причина смерти от такого укуса.
Яд бумсланга является самым сильным из всех заднезубых змей в мире по LD 50 . Хотя ее яд может быть более сильным, чем у некоторых гадюк и элапидов, она вызывает меньше смертельных исходов из-за различных факторов (например, эффективность клыков не высока по сравнению со многими другими змеями, доставляемая доза яда мала, а бумсланги, как правило, менее опасны). агрессивен по сравнению с другими ядовитыми змеями, такими как кобры и мамбы). Симптомы укуса этих змей включают тошноту и внутреннее кровотечение, а человек может умереть от кровоизлияния в мозг и дыхательного коллапса .
Аглифные змеи
[ редактировать ]Опыты, проведенные с секретом околоушной железы Rhabdophis показали и Zamenis, , что даже аглифные змеи не совсем лишены яда, и позволяют сделать вывод, что физиологическое различие между так называемыми безвредными и ядовитыми змеями заключается лишь в степени, всего лишь степени. так же существуют различные этапы превращения обычной околоушной железы в ядовитую железу или твердого зуба в трубчатый или желобчатый клык.
Использование змеиного яда для лечения болезней
[ редактировать ]Учитывая, что змеиный яд содержит множество биологически активных ингредиентов, некоторые из них могут быть полезны для лечения заболеваний. [42]
Например, было обнаружено, что фосфолипазы типа А2 (PLA2) тунисских гадюк Cerastes cerastes и Macrovipera lebetina обладают противоопухолевой активностью. [43] Сообщалось также о противораковой активности других соединений змеиного яда. [44] [45] PLA2 гидролизуют фосфолипиды и, таким образом, могут действовать на поверхности бактериальных клеток, обеспечивая новую противомикробную (антибиотическую) активность. [46]
Обезболивающее ( обезболивающее ) действие многих белков змеиного яда известно давно. [47] [48] Однако основная проблема заключается в том, как доставить белок в нервные клетки: белки обычно неприменимы в виде таблеток.
Иммунитет
[ редактировать ]Среди змей
[ редактировать ]Вопрос о том, обладают ли отдельные змеи иммунитетом к собственному яду, еще окончательно не решен, хотя известен пример кобры, которая отравила сама себя, что привело к большому абсцессу, требующему хирургического вмешательства, но не продемонстрировало никаких других эффектов, которые могли бы иметь место. доказано, что он быстро смертелен для видов-жертв или людей. [49] Кроме того, некоторые безобидные виды, такие как североамериканская королевская змея ( Lampropeltis getula ) и центрально- и южноамериканская mussurana ( Clelia spp.), устойчивы к яду кроталинов , которые часто встречаются в одних и тех же районах и которые они способны подавлять и питаться. Куриная змея ( Spilotes pullatus ) — враг фер-де-ланса ( Botrops caribbaeus ) на Сент-Люсии, и в их столкновениях куриная змея неизменно выходит победителем. Повторные эксперименты показали, что европейская уж ( Natrix natrix ) не подвержена воздействию укуса европейской гадюки ( Vipera berus ) и европейского жереха ( Vipera aspis ), поскольку в крови безобидной змеи присутствует токсических веществ, выделяемых околоушными и губными железами, аналогичных яду этих гадюк. Несколько североамериканских видов крысиных змей, а также королевские змеи оказались невосприимчивы или обладают высокой устойчивостью к яду гремучих змей. Говорят, что королевская кобра, которая охотится на кобр, невосприимчива к их яду.
Среди других животных
[ редактировать ]( Известно, что еж Erinaceidae), мангуст (Herpestidae), медоед ( Mellivora capensis ) и опоссум невосприимчивы к дозе змеиного яда. [ нужна ссылка ] Недавно было обнаружено, что медоед и домашняя свинья конвергентно развили аминокислотные замены в никотиновом рецепторе ацетилхолина, которые, как известно, придают ежам устойчивость к альфа-нейротоксинам. [50] Можно ли свинью иммунитетом, пока неясно, хотя ранние исследования показывают эндогенную устойчивость у свиней, протестированных против нейротоксинов. считать [51] жировой слой свиньи Хотя подкожный может защитить ее от змеиного яда, большинство ядов легко проходят через слои сосудистого жира, поэтому это вряд ли будет способствовать ее способности противостоять яду. Садовая соня ( Eliomys quercinus ) недавно была добавлена в список животных, устойчивых к яду гадюки. Некоторые популяции калифорнийского суслика ( Otospermophilus beechei ) во взрослом состоянии, по крайней мере, частично невосприимчивы к яду гремучей змеи.
Среди людей
[ редактировать ]Приобретение человеком иммунитета против змеиного яда является древним (около 60 г. н. э., племя Псилли ). Исследования по разработке вакцин, которые обеспечат иммунитет, продолжаются. Билл Хааст , владелец и директор Серпентария Майами, на протяжении большей части своей взрослой жизни вводил себе змеиный яд, пытаясь выработать иммунитет к широкому спектру ядовитых змей. Эта практика известна как митридатизм . Хааст дожил до 100 лет и пережил 172 укуса змей. Он сдал свою кровь для лечения пациентов, пострадавших от укуса змеи, когда подходящего противоядия не было в наличии. Более 20 человек, подвергшихся такому лечению, выздоровели. [52] [53] [54] Исследователь-любитель Тим Фриде также позволяет ядовитым змеям кусать себя в надежде на разработку вакцины против змеиного яда, и по состоянию на январь 2016 года он пережил более 160 укусов разных видов. [55]
Традиционные методы лечения
[ редактировать ]По оценкам Всемирной организации здравоохранения , 80% населения мира зависит от традиционной медицины в удовлетворении своих основных медицинских потребностей. [56] Методы традиционного лечения змеиных укусов, хотя и имеют сомнительную эффективность и, возможно, даже вредны, тем не менее актуальны.
Из растений, используемых для лечения змеиных укусов в Тринидаде и Тобаго, готовят настойки на спирте или оливковом масле и хранят в фляжках из-под рома, называемых змеиными бутылками, которые содержат несколько различных растений и/или насекомых. Используемые растения включают виноградную лозу, называемую обезьяньей лестницей ( Bauhinia cumanensis или Bauhinia excisa , Fabaceae), которую растирают и прикладывают к укусу. Альтернативно, настойку готовят из кусочка виноградной лозы и хранят в бутылке со змеей. Другие используемые растения включают корень циновки ( Aristolochia Rugosa ), кошачий коготь ( Pithecellobim unguis-cati ), табак ( Nicotiana tabacum ), змеиный куст ( Barleria lupulina ), семена оби ( Cola nitida ) и корень дикого гри-гри ( Acrocomia aculeata ). В некоторых змеиных бутылках также содержатся гусеницы ( Battus Polydamas , Papilionidae), поедающие листья деревьев ( Aristolochia trilobata ). Лекарства от змей для экстренной помощи получают, пережевывая трехдюймовый кусок корня буа канот ( Cecropia peltata ) и вводя этот раствор из пережеванного корня укушенному субъекту (обычно охотничьей собаке). Это распространенное растение Латинской Америки и Карибского бассейна, что делает его подходящим средством экстренной помощи. Еще одно используемое местное растение — марди гра ( Renealmia alpinia ) (ягоды), которые растирают вместе с соком дикого тростника ( Costus scaber ) и дают укушенным. Быстрые решения включают применение жевательного табака из сигарет, сигар или трубок. [57] Раньше считалось полезным делать надрезы вокруг прокола или высасывать яд, но сейчас этот курс лечения настоятельно не рекомендуется из-за риска самоотравления через порезы ножом или порезы во рту (присоски от укуса змеи). можно использовать наборы, но отсасывание редко приносит какую-либо измеримую пользу). [58] [59]
Серотерапия
[ редактировать ]Серотерапия с использованием противоядия является распространенным в настоящее время методом лечения и была описана еще в 1913 году. [примечание 1] И адаптивный иммунитет, и серотерапия специфичны для данного типа змей; Яд с идентичным физиологическим действием не подвергается перекрестной нейтрализации. Boulenger 1913 описывает следующие случаи:
Европеец в Австралии, который стал невосприимчивым к яду смертельной австралийской тигровой змеи ( Notechis scutatus ), безнаказанно манипулировал этими змеями и находился под впечатлением, что его иммунитет распространяется и на другие виды, когда его укусил равнинный медноголовый змей ( Austrelaps). superbus ), родственный элапину, умер на следующий день.
В Индии было обнаружено , что сыворотка, приготовленная из яда моноклельной кобры Naja kaouthia, не оказывает воздействия на яд двух видов крайтов ( Bungarus ), гадюки Рассела ( Daboia russelli ), чешуйчатой гадюки ( Echis carinatus ) и Папская гадюка ( Trimeresurus popeiorum ). Сыворотка гадюки Рассела не действует на яды колобрина, а также ядов Echis и Trimeresurus .
В Бразилии сыворотка, приготовленная из яда копьеголовых ( вид Bothrops ), не действует на яд гремучей змеи ( вид Crotalus ).
Лечение противоядием от змеиного укуса должно соответствовать типу произошедшего отравления. В Америке доступны поливалентные противоядия, эффективные против укусов большинства гадюк. Крофаб — это противоядие, разработанное для лечения укусов североамериканских гадюк. [60] Они не эффективны против отравления коралловых змей , для чего требуется специальное противоядие к их нейротоксичному яду. Ситуация еще более сложная в таких странах, как Индия, с ее богатой смесью гадюк (Viperidae) и высоко нейротоксичных кобр и крайтов Elapidae.
Примечания
[ редактировать ]- ^ Этот раздел основан на книге Змеи Европы» Г. А. Буленджера « 1913 года, которая сейчас находится в общественном достоянии в Соединенных Штатах (и, возможно, в других странах). Из-за своего возраста текст этой статьи не обязательно следует рассматривать как отражает современные знания о змеином яде
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Исследование яда рептилий» . Австралийский парк рептилий. Архивировано из оригинала 2 февраля 2010 года . Проверено 21 декабря 2010 г.
- ^ Jump up to: а б с д и ж Баушо Р. (1994). Змеи: естественная история . Нью-Йорк, штат Нью-Йорк, США: Sterling Publishing Co., Inc., стр. 194–209 . ISBN 978-1-4027-3181-5 .
- ^ Jump up to: а б с Холлидей А., Крейг Т., ред. (2002). Светлячок Энциклопедия рептилий и земноводных . Торонто, Канада: Firefly Books Ltd., стр. 202–203 . ISBN 978-1-55297-613-5 .
- ^ Jump up to: а б Боттралл Дж.Л., Мадарас Ф., Бивен К.Д., Веннинг М.Г., Мирчин П.Дж. (сентябрь 2010 г.). «Протеолитическая активность ядов змей Elapid и Viperid и ее влияние на пищеварение» . Журнал исследований ядов . 1 (3): 18–28. ПМК 3086185 . ПМИД 21544178 .
- ^ Jump up to: а б с д Оливейра, Ана Л.; Вьегас, Матильда Ф.; да Силва, Сауло Л.; Соареш, Андреймар М.; Рамос, Мария Дж.; Фернандес, Педро А. (июль 2022 г.). «Химия змеиного яда и его лечебный потенциал» . Обзоры природы Химия . 6 (7): 451–469. дои : 10.1038/s41570-022-00393-7 . ISSN 2397-3358 . ПМЦ 9185726 . ПМИД 35702592 .
- ^ Мэттисон С (2007). Новая энциклопедия змей . Нью-Джерси, США (впервые опубликовано в Великобритании): Princeton University Press (Принстон и Оксфорд), впервые опубликовано в Блэндфорде. п. 117. ИСБН 978-0-691-13295-2 .
- ^ Кардозо К.К., Да Силва М.Дж., Коста Г.Г., Торрес Т.Т., Дель Бем Л.Е., Видал Р.О. и др. (октябрь 2010 г.). «Транскриптомный анализ экспрессии генов в ядовитой железе змеи Bothrops alternatus (urutu)» . БМК Геномика . 11 (1): 605. дои : 10.1186/1471-2164-11-605 . ПМК 3017861 . ПМИД 20977763 .
- ^ Кондреа Э., Деврис А., Магер Дж. (февраль 1964 г.). «Гемолиз и расщепление фосфолипидов эритроцитов человека змеиными ядами». Biochimica et Biophysical Acta (BBA) — Специализированная секция по липидам и смежным темам . 84 (1): 60–73. дои : 10.1016/0926-6542(64)90101-5 . ПМИД 14124757 .

- ^ Родригес-Итурральде Д., Сильвейра Р., Барбейто Л., Дахас Ф. (1983). «Фасцикулин, мощный антихолинэстеразный полипептид из яда Dendroaspis angusticeps». Нейрохимия Интернэшнл . 5 (3): 267–74. дои : 10.1016/0197-0186(83)90028-1 . ПМИД 20487949 . S2CID 8952817 .
- ^ Макесси, Стивен П. (1 мая 2021 г.). Макесси, Стивен П. (ред.). Справочник по ядам и токсинам рептилий (2-е изд.). Второе издание. | Бока-Ратон: CRC Press, 2021.: CRC Press. дои : 10.1201/9780429054204 . ISBN 978-0-429-05420-4 .
{{cite book}}: CS1 maint: местоположение ( ссылка ) - ^ Jump up to: а б «Ключевое слово: Геморрагический токсин KW-1200» . ЮниПрот . Проверено 1 июня 2019 г.
- ^ Ходжсон У.К., Викрамаратна Дж.К. (сентябрь 2002 г.). «Нейромышечная активность змеиных ядов in vitro» . Клиническая и экспериментальная фармакология и физиология . 29 (9): 807–14. дои : 10.1046/j.1440-1681.2002.03740.x . ПМИД 12165047 . S2CID 20158638 .
- ^ Хэ YY, Ли WH, Чжан Ю (сентябрь 2004 г.). «Клонирование и очистка альфа-нейротоксинов королевской кобры (Ophiophagus hannah)». Токсикон . 44 (3): 295–303. doi : 10.1016/j.токсикон.2004.06.003 . ПМИД 15302536 .
- ^ Бернардони Х.Л., Соуза Л.Ф., Вермелингер Л.С., Лопес А.С., Пресото БК, Серрано С.М., Зингали РБ, Моура-да-Силва А.М. (14 октября 2014 г.). «Функциональная изменчивость металлопротеиназ змеиного яда: адаптивные преимущества при нацеливании на различную добычу и последствия для отравления человека» . ПЛОС ОДИН . 9 (10): e109651. Бибкод : 2014PLoSO...9j9651B . дои : 10.1371/journal.pone.0109651 . ПМК 4196926 . ПМИД 25313513 .
- ^ Херманс С., Виттевронгель С., Тис С., Сметерст П.А., Ван Гит С., Фресон К. (август 2009 г.). «Сложная гетерозиготная мутация гликопротеина VI у пациента с нарушением свертываемости крови» . Журнал тромбозов и гемостаза . 7 (8): 1356–63. дои : 10.1111/j.1538-7836.2009.03520.x . ПМИД 19552682 . S2CID 205728095 .

- ^ Гриффин PR, Aird SD (ноябрь 1990 г.). «Новый малый миотоксин из яда степной гремучей змеи (Crotalus viridis viridis)» . Письма ФЭБС . 274 (1–2): 43–47. дои : 10.1016/0014-5793(90)81325-I . ПМИД 2253781 . S2CID 45019479 .
- ^ Самедзима Ю., Аоки Ю., Мебс Д. (1991). «Аминокислотная последовательность миотоксина из яда восточной гремучей змеи (Crotalus adamanteus)». Токсикон . 29 (4–5): 461–468. дои : 10.1016/0041-0101(91)90020-р . ПМИД 1862521 .
- ^ Уиттингтон CM, Папенфусс AT, Бансал П., Торрес AM, Вонг ES, Дикин JE и др. (июнь 2008 г.). «Дефензины и конвергентная эволюция генов яда утконоса и рептилий» . Геномные исследования . 18 (6): 986–994. дои : 10.1101/гр.7149808 . ПМК 2413166 . ПМИД 18463304 .
- ^ Броуд А.Дж., Сазерленд С.К., Коултер А.Р. (17 мая 1979 г.). «Смертельность опасного яда австралийских и других змей у мышей» (PDF) . Токсикон . 17 (6): 661–4. дои : 10.1016/0041-0101(79)90245-9 . ПМИД 524395 .
- ^ Розен Ф.С. (31 июля 2003 г.). «Эдвин Дж. Кон и развитие химии белков». Медицинский журнал Новой Англии . 349 (5): 511–512. дои : 10.1056/NEJM200307313490522 .
- ^ Jump up to: а б с д Матейчук П., Даш CH, Гаскойн EW (декабрь 2000 г.). «Производство раствора человеческого альбумина: постоянно развивающийся коллоид» . Британский журнал анестезии . 85 (6): 887–95. дои : 10.1093/бья/85.6.887 . ПМИД 11732525 .
- ^ Бродневич-Проба Т (декабрь 1991 г.). «Фракционирование плазмы человека и влияние новых технологий на использование и качество продуктов, полученных из плазмы». Обзоры крови . 5 (4): 245–57. дои : 10.1016/0268-960x(91)90016-6 . ПМИД 1782484 .
- ^ Jump up to: а б с д и ж г Фрай Б.Г., Кейсвелл Н.Р., Вюстер В., Видал Н., Янг Б., Джексон Т.Н. (сентябрь 2012 г.). «Структурное и функциональное разнообразие системы яда рептилий Toxicofera». Токсикон . 60 (4): 434–48. дои : 10.1016/j.токсикон.2012.02.013 . ПМИД 22446061 .
- ^ Jump up to: а б с д Кейсвелл Н.Р., Вюстер В., Вонк Ф.Дж., Харрисон Р.А., Фрай Б.Г. (апрель 2013 г.). «Сложные коктейли: эволюционная новизна ядов». Тенденции в экологии и эволюции . 28 (4): 219–29. дои : 10.1016/j.tree.2012.10.020 . ПМИД 23219381 .
- ^ Ломонте Б., Фернандес Х., Санс Л., Ангуло Ю., Саса М., Гутьеррес Х.М., Кальвете Х.Дж. (июнь 2014 г.). «Ядовитые змеи Коста-Рики: биологические и медицинские последствия протеомных профилей их ядов, проанализированные с помощью стратегии змеиной ядовитости». Журнал протеомики . 105 : 323–39. дои : 10.1016/j.jprot.2014.02.020 . ПМИД 24576642 .
- ^ Харгривз, Адам Д.; Суэйн, Мартин Т.; Хегарти, Мэтью Дж.; Логан, Даррен В.; Малли, Джон Ф. (август 2014 г.). «Ограничение и набор — дупликация генов, происхождение и эволюция токсинов змеиного яда» . Геномная биология и эволюция . 6 (8): 2088–2095. дои : 10.1093/gbe/evu166 . ISSN 1759-6653 . ПМЦ 4231632 . ПМИД 25079342 .
- ^ Линч VJ (январь 2007 г.). «Изобретение арсенала: адаптивная эволюция и неофункционализация генов фосфолипазы А2 змеиного яда» . Эволюционная биология BMC . 7 (2): 2. дои : 10.1186/1471-2148-7-2 . ПМЦ 1783844 . ПМИД 17233905 .
- ^ Барлоу А., Пук CE, Харрисон Р.А., Вюстер В. (июль 2009 г.). «Совместная эволюция диеты и активности яда, специфичной для добычи, подтверждает роль отбора в эволюции змеиного яда» . Слушания. Биологические науки . 276 (1666): 2443–2449. дои : 10.1098/rspb.2009.0048 . ПМК 2690460 . ПМИД 19364745 .
- ^ Пахари С., Бикфорд Д., Фрай Б.Г., Кини Р.М. (сентябрь 2007 г.). «Схема экспрессии генов трехпальцевого токсина и фосфолипазы А2 в ядовитых железах двух морских змей, Lapemis curtus и Acalyptophis peronii: сравнение эволюции этих токсинов у наземных змей, морских крайтов и морских змей» . Эволюционная биология BMC . 7 : 175. дои : 10.1186/1471-2148-7-175 . ПМК 2174459 . ПМИД 17900344 .
- ^ Jump up to: а б Барлоу А., Пук CE, Харрисон Р.А., Вюстер В. (июль 2009 г.). «Совместная эволюция диеты и активности яда, специфичной для добычи, подтверждает роль отбора в эволюции змеиного яда» . Труды: Биологические науки . 276 (1666): 2443–9. дои : 10.1098/rspb.2009.0048 . JSTOR 30244073 . ПМК 2690460 . ПМИД 19364745 .
- ^ Янса С.А., Восс РС (22 июня 2011 г.). «Адаптивная эволюция белка vWF, нацеленного на яд, у опоссумов, поедающих гадюк» . ПЛОС ОДИН . 6 (6): e20997. Бибкод : 2011PLoSO...620997J . дои : 10.1371/journal.pone.0020997 . ПМК 3120824 . ПМИД 21731638 .
- ^ Кальвете Дж.Дж., Гезеллу П., Пайва О., Матинахо Т., Гассемпур А., Гударзи Х., Краус Ф., Санс Л., Уильямс DJ (июль 2012 г.). «Змеиная ядовитость двух малоизвестных Hydrophiinae: сравнительная протеомика ядов наземных Toxicocalamus longissimus и морских Hydrophis cyanocinctus». Журнал протеомики . 75 (13): 4091–101. дои : 10.1016/j.jprot.2012.05.026 . ПМИД 22643073 .
- ^ Ли М., Фрай Б.Г., Кини Р.М. (январь 2005 г.). «Диета, состоящая только из яиц: ее влияние на изменения профиля токсинов и экологию мраморной морской змеи (Aipysurus eydouxii)». Журнал молекулярной эволюции . 60 (1): 81–9. Бибкод : 2005JMolE..60...81L . дои : 10.1007/s00239-004-0138-0 . ПМИД 15696370 . S2CID 17572816 .
- ^ Макесси С.П. (июль 2010 г.). «Эволюционные тенденции в составе яда западных гремучих змей (Crotalus viridis sensu lato): токсичность и смягчающие вещества». Токсикон . 55 (8): 1463–74. doi : 10.1016/j.токсикон.2010.02.028 . ПМИД 20227433 .
- ^ Савиола А.Дж., Чизар Д., Буш С., Макесси С.П. (март 2013 г.). «Молекулярные основы перемещения добычи у змей-гадюк» . БМК Биология . 11 (1): 20. дои : 10.1186/1741-7007-11-20 . ПМЦ 3635877 . ПМИД 23452837 .
- ^ Янгман, Николас Дж; Ллинас, Джошуа; Хаворт, Марк; Джиллетт, Эмбер; Джонс, Ли; Уокер, Эндрю А; Фрай, Брайан Дж. (июнь 2022 г.). «Распутывание взаимодействий между гадюками Bitis и их добычей с использованием коагулотоксичности против плазмы различных позвоночных» . Токсикон . 216 : 37–44. doi : 10.1016/j.токсикон.2022.06.012 . ISSN 1879-3150 . ПМИД 35780972 . S2CID 250188352 .
- ^ Дэвис, Эмма-Луиза; Арбакл, Кевин (декабрь 2019 г.). «Коэволюция токсичной деятельности и диеты змеиного яда: свидетельства того, что экологический обобщение способствует токсикологическому разнообразию» . Токсины . 11 (12): 711. doi : 10.3390/toxins11120711 . ISSN 2072-6651 . ПМК 6950196 . ПМИД 31817769 .
- ^ МакКью, доктор медицинских наук (октябрь 2007 г.). «Отравление жертвой не улучшает пищеварение у западных гремучих змей с ромбовидной спиной (Crotalus atrox)». Журнал экспериментальной зоологии, часть A. 307 (10): 568–77. дои : 10.1002/jez.411 . ПМИД 17671964 .
- ^ Брокховен С., Дю Плесси А (август 2017 г.). «Эволюция змеиных клыков утратила свою остроту? Новые идеи с точки зрения строительной механики» . Письма по биологии . 13 (8): 20170293. doi : 10.1098/rsbl.2017.0293 . ПМК 5582107 . ПМИД 28768797 .
- ^ Клерен С.Г., Хокинг Д.П., Эванс А.Р. (июнь 2021 г.). «Эволюция клыков ядовитых змей: адаптация трехмерной формы зубов к биомеханическим свойствам добычи» . Эволюция; Международный журнал органической эволюции . 75 (6): 1377–1394. дои : 10.1111/evo.14239 . ПМИД 33904594 . S2CID 233411378 .
- ^ Мартин С.Дж., Лэмб Дж. (1907). «Змеиный яд и змеиный укус». В Allbutt TC, Rolleston ND (ред.). Система медицины . Лондон: Макмиллан. стр. 783–821.
- ^ Макклири Р.Дж., Кини Р.М. (февраль 2013 г.). «Неферментативные белки из змеиных ядов: золотая жила фармакологических инструментов и лекарств». Токсикон . 62 : 56–74. дои : 10.1016/j.токсикон.2012.09.008 . ПМИД 23058997 .
- ^ Зуари-Кессентини Р., Сраири-Абид Н., Базаа А., Эль Айеб М., Луис Дж., Марракчи Н. (2013). «Противоопухолевый потенциал ядов тунисских змей, секретируемых фосфолипазами А2» . БиоМед Исследования Интернэшнл . 2013 : 1–9. дои : 10.1155/2013/391389 . ПМЦ 3581298 . ПМИД 23509718 .
- ^ Вьяс В.К., Брамбхатт К., Бхатт Х., Пармар У. (февраль 2013 г.). «Терапевтический потенциал змеиного яда в терапии рака: современные перспективы» . Азиатско-Тихоокеанский журнал тропической биомедицины . 3 (2): 156–62. дои : 10.1016/S2221-1691(13)60042-8 . ПМЦ 3627178 . ПМИД 23593597 .
- ^ Джайн Д., Кумар С. (2012). «Змеиный яд: мощное противораковое средство» . Азиатско-Тихоокеанский журнал профилактики рака . 13 (10): 4855–60. дои : 10.7314/apjcp.2012.13.10.4855 . ПМИД 23244070 .
- ^ де Оливейра-младший, НГ, и Сильва Кардосо, МХ, Франко ОЛ (декабрь 2013 г.). «Змеиные яды: привлекательные противомикробные белковые соединения для терапевтических целей» . Клеточные и молекулярные науки о жизни . 70 (24): 4645–58. дои : 10.1007/s00018-013-1345-x . ПМЦ 11113393 . ПМИД 23657358 . S2CID 15127065 .
- ^ Вульф CJ (январь 2013 г.). «Боль: морфин, метаболиты, мамбы и мутации». «Ланцет». Неврология . 12 (1): 18–20. дои : 10.1016/S1474-4422(12)70287-9 . ПМИД 23237896 . S2CID 8697382 .
- ^ Осипов А, Уткин Ю (декабрь 2012 г.). «Влияние полипептидов змеиного яда на центральную нервную систему». Агенты центральной нервной системы в медицинской химии . 12 (4): 315–28. дои : 10.2174/187152412803760618 . ПМИД 23270323 . S2CID 36274766 .
- ^ «Стерильный хвостовой абсцесс у Naja annulifera - случай самоотравления» . Архивировано из оригинала 27 октября 2004 года . Проверено 2 апреля 2009 г.
- ^ Драбек Д., Янса С. (2015). «Почему медоеду все равно: независимая эволюция устойчивости к трехпальцевым токсинам в никотиновом рецепторе ацетилхолина». Токсикон . 99 : 68–72. дои : 10.1016/j.токсикон.2015.03.007 . ПМИД 25796346 .
- ^ Грассе Э., Зоутендиканда А., Шаафсма А. (1935). «Исследование токсических и антигенных свойств ядов южноафриканских змей с особым упором на поливалентность южноафриканского противовенена». Пер. Р. Сок. Троп. Мед. Гигиена . 28 (6): 601–612. дои : 10.1016/S0035-9203(35)90031-1 .
- ^ «Прощание с этими знаменитыми флоридцами» . Флоридский тренд . 19 декабря 2011 года . Проверено 2 апреля 2012 г.
- ^ Розенберг С. (21 июня 2011 г.). «Билл Хааст умирает в возрасте 100 лет; змеи были очарованием знаменитостей Южной Флориды» . Лос-Анджелес Таймс . Проверено 16 октября 2012 г.
- ^ Шудель М. (18 июня 2011 г.). «Билл Хааст умирает в 100 лет: человек-змея из Флориды предоставил яд для сыворотки от змеиного укуса» . Вашингтон Пост . Проверено 16 октября 2012 г.
- ^ «Человек заставил смертельных змей укусить себя 160 раз в поисках человеческого противоядия | Америка | Новости | The Independent» . Independent.co.uk . 21 января 2016 года . Проверено 7 июля 2016 г.
- ^ Хиремат В.Т., Таранат ТК (февраль 2010 г.). «Традиционная фитотерапия от укусов змей племенами района Читрадурга, Карнатака, Индия» . Этноботанические листовки . 14 (2): 120–125.
- ^ Зетелиус М., Балик М.Дж. (март 1982 г.). «Современная медицина и шаманский ритуал: случай положительного синергетического ответа при лечении укуса змеи» (PDF) . Журнал этнофармакологии . 5 (2): 181–5. дои : 10.1016/0378-8741(82)90042-3 . ПМИД 7057657 .
- ^ «Лечение змеиных укусов» . Ces.ncsu.edu . Проверено 16 октября 2012 г.
- ^ «CDC — Ядовитые змеи — Тема безопасности и гигиены труда NIOSH» . CDC.gov. 1 июля 2016 года . Проверено 7 июля 2016 г.
- ^ http://www.savagelabs.com/Products/CroFab/Home/crofab_frame.htm. Архивировано 3 марта 2016 г. на Wayback Machine. Ссылка на PDF-файл с полной информацией о назначении, получено 12.11.12.
Дальнейшее чтение
[ редактировать ]- Йонассен I, Коллинз Дж. Ф., Хиггинс Д. Г. (август 1995 г.). «Обнаружение гибких закономерностей в невыровненных белковых последовательностях» . Белковая наука . 4 (8): 1587–95. дои : 10.1002/pro.5560040817 . ПМК 2143188 . ПМИД 8520485 .
- Шоу IC (2007). «Глава 19: Змеиные токсины». В Уоринге Р.Х., Стивентоне ГБ, Митчелле С.С. (ред.). Молекулы смерти (второе изд.). Ривер Эдж, Нью-Джерси: Издательство Имперского колледжа. стр. 329–344 . ISBN 978-1-86094-815-2 .
Внешние ссылки
[ редактировать ]- Обзор разнообразия и эволюции змеиных клыков.
- Змеиные яды UMich Ориентация белков в мембранах семейств/суперсемейства-55 - Расчетная ориентация фосфолипаз А2 змеиного яда и миотоксинов в липидном бислое.
- LD50 для большинства токсичных ядов.
- Австралийское отделение исследования ядов - общий источник информации о ядовитых существах Австралии.
- biomedcentral.com — Лекарственные и этноветеринарные средства охотников в Тринидаде .
- Reptilis.net - Как действует яд.
- Snakevenom.net - Сушка и хранение змеиного яда.