Змея
Эту статью может потребовать очистки Википедии , чтобы она соответствовала стандартам качества . Конкретная проблема заключается в следующем: нецитируемый текст, чрезмерный вес и плохая структура. ( июнь 2024 г. ) |
| Змеи | |
|---|---|
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Заказ: | Чешуйчатый |
| Клэйд : | Офидия |
| Подотряд: | Змеи Линней , 1758 г. |
| Инфраструктурные заказы | |
| |
 | |
| Примерное мировое распространение змей всех видов. | |




Змеи — удлиненные, конечностей рептилии подотряда ˈ Serpentes ( / s ɜːr лишенные p ɛ n t iː z / ). [2] Как и все другие чешуйчатые , змеи являются экзотермическими , амниотами позвоночными покрытыми перекрывающимися чешуйками . У многих видов змей черепа имеют на несколько больше суставов, чем у их предков- ящериц , что позволяет им заглатывать добычу, размер которой намного превышает размер их головы ( краниальный кинез ). Чтобы разместить свое узкое тело, парные органы змей (например, почки) располагаются один перед другим, а не рядом, и у большинства из них есть только одно функциональное легкое . У некоторых видов сохраняется тазовый пояс с парой рудиментарных когтей по обе стороны от клоаки . Ящерицы независимо развивали удлиненные тела без конечностей или со значительно уменьшенными конечностями, по крайней мере, двадцать пять раз в результате конвергентной эволюции , что привело к появлению множества линий безногих ящериц . [3] Они напоминают змей, но некоторые распространенные группы безногих ящериц имеют веки и наружные уши, которых нет у змей, хотя это правило не является универсальным (см. Amphisbaenia , Dibamidae и Pygopodidae ).
Живые змеи встречаются на всех континентах, кроме Антарктиды, и на большинстве небольших участков суши; исключения включают некоторые крупные острова, такие как Ирландия, Исландия, Гренландия и острова Новой Зеландии, а также множество мелких островов Атлантического и центральной части Тихого океана. [4] Кроме того, морские змеи широко распространены в Индийском и Тихом океанах. около тридцати семейств В настоящее время известно , включающих около 520 родов и около 3900 видов . [5] Их размер варьируется от крошечной барбадосской змеи длиной 10,4 см (4,1 дюйма). [6] до сетчатого питона длиной 6,95 метра (22,8 фута). [7] Ископаемый вид Titanoboa cerrejonensis имел длину 12,8 метра (42 фута). [8] Считается, что змеи произошли от роющих или водных ящериц, возможно, в юрский период, причем самые ранние известные окаменелости датируются периодом от 143 до 167 млн лет назад. [9] [10] Разнообразие современных змей появилось в эпоху палеоцена ( ок. 66–56 млн лет назад, после мел-палеогенового вымирания ). Самые старые сохранившиеся описания змей можно найти в Бруклинском папирусе .
Большинство видов змей неядовиты, а те, у кого есть яд, используют его в первую очередь для убийства и подчинения добычи, а не для самообороны. Некоторые из них обладают ядом, достаточно сильным, чтобы причинить людям болезненные травмы или смерть. Неядовитые змеи либо заглатывают добычу живьем, либо убивают ее сжиманием .
Этимология
Английское слово « змея» происходит от древнеанглийского snaca , которое само по себе происходит от протогерманского * snak-an- ( ср. германское Schnake «кольцевая змея», шведское snok «травяная змея»), от протоиндоевропейского корня * (s)nēg. -o- 'ползти, чтобы ползти', что также дает подкрадываться, а также санскритское nagá 'змея'. [11] Слово вытеснило гадюку , поскольку гадюка сузила свое значение, хотя в древнеанглийском языке næddre было общим словом, обозначающим змею. [12] Другой термин, змей , происходит от французского языка, в конечном итоге от индоевропейского * serp — «ползти», [13] что также дало древнегреческое ἕρπω ( hérpō ) «я ползаю» и санскритское sarpá «змея». [14]
Таксономия
Все современные змеи сгруппированы в подотряде Serpentes в таксономии Линнея , входящем в отряд Чешуйчатые , хотя их точное размещение внутри чешуйчатых остается спорным. [15]
Два инфраотряда Serpentes — это Alethinophidia и Scolecophidia . [15] Такое разделение основано на морфологических характеристиках и сходстве последовательностей митохондриальной ДНК . Алетинофидии иногда разделяются на Henophidia и Caenophidia , причем последние состоят из «колубридных» змей ( колобриды , гадюки , элапиды , гидрофииды и атрактаспиды ) и акрохордид, в то время как другие семейства алетинофидий включают Henophidia. [16] не сохранилось сегодня Хотя Madtsoiidae , семейство гигантских, примитивных, похожих на питона змей, , оно существовало примерно 50 000 лет назад в Австралии и было представлено такими родами, как Wonambi . [17]
Недавние молекулярные исследования подтверждают монофилию клад Cylindrophis современных змей, сколекофидий, тифлопидов + аномалепидид, алетинофидий, основных алетинофидий, уропельтид ( , Anomochilus , uropeltines), макростоматанов, буоидов, боидов, питонидов и ценофидий. [18]
Семьи
| Инфраотряд алетинофидии 25 семейств. | |||||
| Семья [5] | Автор таксона [5] | Роды [5] | Разновидность [5] | Общее имя | Географический диапазон [19] |
|---|---|---|---|---|---|
| Акрохордиды | Бонапарт , 1831 г. | 1 | 3 | Бородавки змеи | Западная Индия и Шри-Ланка через тропическую Юго-Восточную Азию до Филиппин, на юг через индонезийско-малайзийскую группу островов до Тимора, на восток через Новую Гвинею до северного побережья Австралии до острова Муссау , архипелага Бисмарка и острова Гуадалканал на Соломоновых островах. |
| Анилииды | Штейнегер , 1907 год. | 1 | 1 | Ложная коралловая змея | Тропическая Южная Америка. |
| Аномохилиды | Кундалл, Уоллах, 1993 г. | 1 | 3 | Карликовые трубочные змеи | Западная Малайзия и на индонезийском острове Суматра . |
| Атрактаспидиды | Гюнтер , 1858 г. | 12 | 72 | Роющие аспиды | Африка и Ближний Восток |
| Боиды | Грей , 1825 г. | 14 | 61 | Хороший | Северная, Центральная и Южная Америка, Карибский бассейн, Юго-Восточная Европа и Малая Азия, Северная, Центральная и Восточная Африка, Мадагаскар и остров Реюньон , Аравийский полуостров, Центральная и Юго-Западная Азия, Индия и Шри-Ланка, Молуккские острова и Новая Гвинея вплоть до Меланезии. и Самоа. |
| Больерииды | Хоффстеттер , 1946 год. | 2 | 2 | Двучелюстные змеи | Маврикий . |
| Colubridae | Опал , 1811 г. | 258 [5] | 2055 [5] | Типичные змеи | Распространен на всех континентах, кроме Антарктиды. [20] |
| Циклокориды | Вейнелл и Браун , 2017 г. | 5 | 8 | Циклокориды | Филиппины |
| Цилиндрофииды | Фитцингер , 1843 г. | 1 | 14 | Азиатские трубочные змеи | Шри-Ланка на восток через Мьянму, Таиланд, Камбоджу, Вьетнам и Малайский архипелаг до островов Ару у юго-западного побережья Новой Гвинеи. Также встречается на юге Китая (Фуцзянь, Гонконг и на острове Хайнань) и в Лаосе. |
| Элапиды | Бойе , 1827 г. | 55 | 389 | Элапиды | На суше, по всему миру в тропических и субтропических регионах, кроме Европы. Морские змеи встречаются в Индийском океане и Тихом океане. [21] |
| Гомалопсиды | Бонапарт , 1845 г. | 28 | 53 | Гомалопсиды | Юго-Восточная Азия и север Австралии. |
| Лампрофииды | Фитцингер , 1843 г. | 16 | 89 | Лампрофииды (ранее включали Atracaspididae, Psammophiidae и некоторые другие семейства) | Африка (включая Сейшельские острова) |
| локсоцемиды | Коуп , 1861 г. | 1 | 1 | Мексиканская роющая змея | Вдоль Тихого океана от Мексики на юг до Коста-Рики. |
| Микрелапиды | Дас и др ., 2023 г. | 1 | 4 | Двуглавые змеи | Восточная Африка и Левант |
| Парейды | Ромер, 1956 год. | 3 | 20 | Змеи, питающиеся улитками | Юго-Восточная Азия и острова Зондского шельфа (Суматра, Борнео, Ява и окружающие их более мелкие острова). |
| Просимниды | Келли, Баркер, Виллет и Бродли, 2009 г. | 1 | 16 | Лопатоносые змеи | Африка к югу от Сахары |
| Псаммодинастиды | Дас и др ., 2024 г. | 1 | 2 | Ложные гадюки | Тропическая Азия |
| Псаммофииды | Буржуа , 1968 год. | 8 | 55 | Псаммофииды | Африка (включая Мадагаскар), Азия и южная Европа. |
| Псевдаспидиды | Коуп , 1893 г. | 3 | 4 | Псевдаспидиды | В основном страны Африки к югу от Сахары; два вида в Юго-Восточной Азии |
| Псевдоксирофииды | Даулинг , 1975 год. | 22 | 89 | Псевдоксирофииды | В основном Мадагаскар и Коморские острова; 5 видов в Африке к югу от Сахары, 1 на Сокотре. |
| Pythonidae | Фитцингер , 1826 г. | 8 | 40 | Питоны | Африка к югу от Сахары, Индия, Мьянма, южный Китай, Юго-Восточная Азия и от юго-востока Филиппин через Индонезию до Новой Гвинеи и Австралии. |
| Тропидофииды | Бронгерсма , 1951 год. | 2 | 34 | хороший гном | Вест-Индия; также Панама и северо-запад Южной Америки, а также северо-запад и юго-восток Бразилии. |
| Уропелтиды | Мюллер , 1832 г. | 8 | 55 | Щитчатохвостые змеи | Южная Индия и Шри-Ланка. |
| Гадюки | Опал , 1811 г. | 35 | 341 | Змеи | Америка, Африка и Евразия к востоку от линии Уоллеса . |
| Ксенодермиды | Коуп , 1900 год. | 6 | 18 | Дракон и змеи необычной чешуи | Южная и Юго-Восточная Азия, а также острова Зондского шельфа (Суматра, Борнео, Ява и окружающие их более мелкие острова). |
| Ксенопельтиды | Бонапарт , 1845 г. | 1 | 2 | Солнечные змеи | Юго-Восточная Азия от Андаманских и Никобарских островов , на восток через Мьянму до южного Китая, Таиланда, Лаоса, Камбоджи, Вьетнама, Малайского полуострова и Ост-Индии до Сулавеси , а также Филиппин. |
| Ксенофидииды | Уоллах и Гюнтер, 1998 г. | 1 | 2 | Змеи с хребтовой челюстью | Борнео и полуостров Малайзия. |
| Инфраотряд сколекофидиев 5 семейств. | |||||
| Семья [5] | Автор таксона [5] | Роды [5] | Разновидность [5] | Общее имя | Географический диапазон [19] |
|---|---|---|---|---|---|
| Аномалепиды | Тейлор , 1939 год. | 4 | 18 | Примитивные слепые змеи | От юга Центральной Америки до северо-запада Южной Америки. Разрозненные популяции на северо-востоке и юго-востоке Южной Америки. |
| Герропилиды | Видал, Винн, Доннеллан и Хеджес, 2010 г. | 2 | 18 | Индо-малайские слепые змеи | Южная и Юго-Восточная Азия, включая Шри-Ланку, Филиппины и Новую Гвинею. |
| Лептотифлопиды | Штейнегер , 1892 г. | 13 | 139 | Тонкие слепые змеи | Африка, Западная Азия от Турции до северо-западной Индии, на острове Сокотра , от юго-запада США на юг через Мексику и от Центральной до Южной Америки, хотя и не в высоких Андах . В Тихоокеанском регионе Южной Америки они встречаются на юге до южного побережья Перу, а на атлантической стороне - до Уругвая и Аргентины. В Карибском бассейне они встречаются на Багамах, Эспаньоле и Малых Антильских островах . |
| Тифлопиды | Я так понимаю , 1820 г. | 18 | 266 | Типичные слепые змеи | Большинство тропических и многие субтропические регионы мира, особенно в Африке, на Мадагаскаре, в Азии, на островах Тихого океана, в тропической Америке и в юго-восточной Европе. |
| Ксенотифлопиды | Видаль, Венс, Бранч и Хеджес, 2010 г. | 1 | 1 | Круглоносая слепая змея | Северный Мадагаскар. |
Безногие ящерицы
Хотя змеи — это рептилии без конечностей, произошедшие от ящериц (и сгруппированные с ними), существует множество других видов ящериц, которые потеряли свои конечности независимо, но внешне внешне похожи на змей. К ним относятся медленный червь , стеклянная змея и земноводные . [22]
Эволюция
| Филогенетический обзор современных змей на семейном уровне. | |||
|
Летопись окаменелостей змей относительно скудна, поскольку скелеты змей обычно маленькие и хрупкие, поэтому окаменелость встречается редко. Окаменелости, легко идентифицируемые как змеи (хотя часто сохраняющие задние конечности), впервые появляются в летописи окаменелостей в меловой период. [24] Самые ранние известные окаменелости настоящих змей (члены кронной группы Serpentes) происходят от морских симолиофиид , самым старым из которых является позднемеловой ( сеноманский возраст) Haasiophis terrasanctus с Западного берега реки Иордан . [1] возраст от 112 до 94 миллионов лет. [25]
На основании геномного анализа установлено, что змеи произошли от ящериц . [23] Этот вывод также подтверждается данными сравнительной анатомии и летописью окаменелостей. [26] : 11 [27] [23]
Питоны и удавы — примитивные группы современных змей — имеют рудиментарные задние конечности: крошечные когтистые пальцы, известные как анальные шпоры , которые используются для захвата во время спаривания. [26] : 11 [28] Семейства Leptotyphlopidae и Typhlopidae также обладают остатками тазового пояса, которые при видимом виде выглядят как роговые выступы.
Передние конечности отсутствуют у всех известных змей. Это вызвано эволюцией их Hox-генов , контролирующих морфогенез конечностей . Осевой скелет общего предка змей, как и большинство других четвероногих, имел региональную специализацию, состоящую из шейных (шеи), грудных (грудных), поясничных (нижней части спины), крестцовых (тазовых) и хвостовых (хвостовых) позвонков. На ранних этапах эволюции змей экспрессия гена Hox в осевом скелете, ответственная за развитие грудной клетки, стала доминантной. В результате все позвонки перед зачатками задних конечностей (если они есть) имеют одинаковую грудную идентичность (за исключением атланта , оси и 1–3 шейных позвонков). Другими словами, большая часть скелета змеи представляет собой чрезвычайно вытянутую грудную клетку. Ребра находятся исключительно на грудных позвонках. Число шейных, поясничных и тазовых позвонков сильно уменьшено (имеются только 2–10 поясничных и тазовых позвонков), а от хвостовых позвонков остался лишь короткий хвост. Тем не менее, хвост все еще достаточно длинный, чтобы иметь важное применение у многих видов, а у некоторых водных и древесных видов он видоизменился.
Многие современные группы змей возникли в палеоцене , наряду с адаптивной радиацией млекопитающих после вымирания (нептичьих) динозавров . Расширение лугов в Северной Америке также привело к взрывной радиации среди змей. [29] Раньше змеи были второстепенным компонентом фауны Северной Америки, но в миоцене число видов и их распространенность резко возросли с первыми появлениями гадюк и элапид в Северной Америке и значительным разнообразием Colubridae (включая происхождение многих современные роды, такие как Nerodia , Lampropeltis , Pituophis и Pantherophis ). [29]
Окаменелости
Существуют ископаемые свидетельства, позволяющие предположить, что змеи произошли от роющих ящериц. [30] в меловой период . [31] Ранний ископаемый родственник змеи, Наджаш рионегрина , был двуногим роющим животным с крестцом и был полностью наземным . [32] Наджаш , живший 95 миллионов лет назад, также имел череп с некоторыми особенностями, типичными для ящериц, но у него развились некоторые подвижные суставы черепа, которые определяют гибкий череп у большинства современных змей. Этот вид не имел никакого сходства с современными роющими слепыми змеями, которых часто рассматривают как наиболее примитивную группу современных форм. [33] Одним из сохранившихся аналогов этих предполагаемых предков является безухий варан Лантанот с Борнео (хотя он также ведет полуводный образ жизни ). [34] Подземные виды развили тела, обтекаемые для рытья нор, и в конечном итоге лишились конечностей. [34] Согласно этой гипотезе, такие особенности, как прозрачные , сросшиеся веки ( бриллы ) и потеря наружных ушей, развились, чтобы справиться с трудностями, связанными с окаменением , такими как царапины на роговице и грязь в ушах. [31] [34] Известно, что у некоторых примитивных змей были задние конечности, но их тазовые кости не имели прямого соединения с позвонками. К ним относятся ископаемые виды, такие как Haasiophis , Pachyrhachis и Eupodophis , которые немного старше Наджаша . [28]
Эта гипотеза была подкреплена в 2015 году открытием в Бразилии окаменелости четвероногой змеи возрастом 113 миллионов лет, получившей название Tetrapodophis amplectus . У него много змееподобных черт, он приспособлен к рытью нор, а его желудок указывает на то, что он охотился на других животных. [35] В настоящее время неясно, является ли Tetrapodophis змеей или другим видом из отряда чешуйчатых , поскольку змееподобное тело эволюционировало независимо по крайней мере 26 раз. Tetrapodophis не имеет характерных змеиных черт позвоночника и черепа. [36] [37] Исследование, проведенное в 2021 году, относит это животное к группе вымерших морских ящериц мелового периода, известных как долихозавры и не имеющих прямого отношения к змеям. [38]
Альтернативная гипотеза, основанная на морфологии , предполагает, что предки змей были связаны с мозазаврами — вымершими водными рептилиями мелового периода — образующими кладу Pythonomorpha . [27] Согласно этой гипотезе, считается, что сросшиеся прозрачные веки змей эволюционировали для борьбы с морскими условиями (потеря воды роговицей в результате осмоса), а наружные уши были потеряны из-за неиспользования в водной среде. В конечном итоге это привело к появлению животного, похожего на сегодняшних морских змей . В позднем меловом периоде змеи вновь заселили землю и продолжили превращаться в сегодняшних змей. Ископаемые останки змей известны из морских отложений раннего позднего мела, что согласуется с этой гипотезой; тем более, что они старше наземного Наджаша рионегрины . Сходное строение черепа, уменьшенные или отсутствующие конечности и другие анатомические особенности, обнаруженные как у мозазавров, так и у змей, приводят к положительной кладистической корреляции, хотя некоторые из этих особенностей являются общими с варанидами. [ нужна ссылка ]
Генетические исследования последних лет показали, что змеи не так тесно связаны с варанами, как когда-то считалось, и, следовательно, не с мозазаврами, предполагаемыми предками в водном сценарии их эволюции. Однако больше доказательств связывают мозазавров со змеями, чем с варанидами. Фрагментированные останки, обнаруженные в юрском и раннем меловом периоде, указывают на более глубокие летописи окаменелостей этих групп, которые потенциально могут опровергнуть любую гипотезу. [39] [40]



 Eupodophis descouensi Задняя нога
Eupodophis descouensi Задняя нога




Генетическая основа эволюции змей
И окаменелости, и филогенетические исследования показывают, что змеи произошли от ящериц , поэтому возник вопрос, какие генетические изменения привели к потере конечностей у предков змей. Потеря конечностей на самом деле очень распространена среди современных рептилий и случалась десятки раз у сцинков , ангвидов и других ящериц. [41]
В 2016 году два исследования показали, что потеря конечностей у змей связана с мутациями ДНК в регуляторной последовательности зоны поляризационной активности (ZRS), регуляторной области гена звукового ежа , который критически необходим для развития конечностей. У более продвинутых змей нет остатков конечностей, но у базальных змей, таких как питоны и удавы, действительно есть следы сильно редуцированных рудиментарных задних конечностей. У эмбрионов питона даже есть полностью развитые зачатки задних конечностей, но их дальнейшее развитие останавливается из-за мутаций ДНК в ZRS. [42] [43] [44] [45]
Распределение
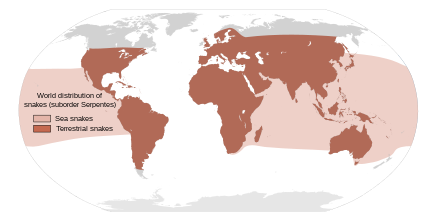
Существует около 3900 видов змей. [46] простирается на север до Полярного круга в Скандинавии и на юг через Австралию. [27] Змей можно найти на всех континентах, кроме Антарктиды, а также в море и на высоте до 16 000 футов (4900 м) в Гималайских горах Азии. [27] [47] : 143 Есть множество островов, на которых змеи отсутствуют, например, Ирландия , Исландия и Новая Зеландия. [4] [47] (хотя северные воды Новой Зеландии нечасто посещают желтобрюхая морская змея и полосатый морской крайт ). [48]
Биология

Размер
Ныне вымерший Titanoboa cerrejonensis имел длину 12,8 м (42 фута). [8] Для сравнения, самые крупные из ныне живущих змей — сетчатый питон длиной около 6,95 м (22,8 фута). [7] и зеленая анаконда длиной около 5,21 м (17,1 фута) и считающаяся самой тяжелой змеей на Земле (97,5 кг (215 фунтов)). [49]
На другом конце шкалы находится самая маленькая из ныне живущих змей — Leptotyphlops carlae , длина которой составляет около 10,4 см (4,1 дюйма). [6] Большинство змей — довольно маленькие животные, около 1 м (3,3 фута) в длину. [50]
Восприятие
Некоторые из наиболее высокоразвитых сенсорных систем обнаружены у Crotalidae, или гадюк, — гремучих змей и их сородичей. У гадюк есть все органы чувств других змей, а также дополнительные вспомогательные средства. Ямка относится к специальным чувствительным к инфракрасному излучению рецепторам, расположенным по обе стороны головы, между ноздрями и глазами. На самом деле ямка выглядит как дополнительная пара ноздрей. Все змеи обладают способностью ощущать тепло с помощью сенсорных и тепловых рецепторов, как и другие животные; однако у ямных гадюк характерна сильно развитая ямка. Каждая ямка состоит из ямочной полости и внутренней полости, более крупная из них лежит сразу за и обычно ниже уровня ноздри и открывается вперед. За этой большей полостью находится более тонкая, едва заметная; полости соединены внутри, разделены только мембраной с нервами, которые необычайно приспособлены к обнаружению изменений температуры между ними. Как и в случае с перекрывающимися полями зрения человеческих глаз, обращенная вперед ямка с обеих сторон лица вместе создает поле зрения: гадюка может различать объекты и их окружение, а также точно оценивать расстояние между объектами и собой. . Способность гадюки чувствовать тепло настолько велика, что она может реагировать на разницу всего в одну треть градуса по Фаренгейту. У других змей, чувствительных к инфракрасному излучению, есть несколько небольших губных ямок, выстилающих верхнюю губу, чуть ниже ноздрей. [51]
Змея выслеживает свою добычу с помощью обоняния, собирая частицы в воздухе своим раздвоенным языком , а затем передавая их в сошниково-носовой орган или орган Якобсона во рту для исследования. [51] Вилка языка обеспечивает своего рода направленное обоняние и вкус одновременно. [51] Язык змеи постоянно находится в движении, отбирая частицы из воздуха, земли и воды, анализируя найденные химические вещества и определяя присутствие добычи или хищников в местной среде. У водных змей, таких как анаконда , язык эффективно функционирует под водой. [51]
Чтобы уловить частицы в воздухе, язык высовывается. Подобно руке, принимающей что-то на себя, вилка языка одновременно обеспечивает своего рода чувство направления. Змеи обладают хорошим обонянием, но это чувство значительно усиливается в окне особого органа — органа Якобсона. Когда язык втягивается обратно в рот, его раздвоенный кончик вдавливается в полости органа Якобсона. Находясь в определенной точке, язык и орган Якобсона работают согласованно, выполняя анализ вкуса и запаха. Сам орган дает змее экстрасенсорный канал. В буквальном смысле змея чувствует вкус окрестностей, способная скользить по информационным комнатам, как будто двери открыты. [52] [ нужна страница ] [51]

Вплоть до середины 20 века считалось, что змеи не слышат. [53] [54] На самом деле у змей есть две отдельные и совершенно независимые системы. Одна из этих систем, соматическая, включает передачу частот через вентральные рецепторы кожи через позвоночник. Другая система включает в себя вибрации, которые передаются через ослабленные легкие змеи в мозг через черепной нерв. Чувствительность змеи к вибрации чрезвычайно высока. [53] [51] [54] В тихой комнате змея может услышать тихую речь. [52] [ нужна страница ]
Зрение змей сильно различается у разных видов. Некоторые из них обладают острым зрением, а другие способны отличать только свет от темноты, но важной тенденцией является то, что зрительное восприятие змей достаточно адекватно, чтобы отслеживать движения. [55] Как правило, зрение лучше всего у древесных змей и самое слабое у роющих змей. Некоторые обладают бинокулярным зрением , при котором оба глаза способны фокусироваться на одной и той же точке, примером тому является азиатская виноградная змея . Большинство змей фокусируются, перемещая хрусталик вперед и назад относительно сетчатки . У дневных змей зрачки круглые, а у многих ночных змей зрачки щелевые. Большинство видов обладают тремя зрительными пигментами и, вероятно, способны видеть два основных цвета при дневном свете. Кольчатая морская змея и род Helicops, похоже, восстановили большую часть своего цветового зрения в результате адаптации к морской среде, в которой они живут. [56] [57] Был сделан вывод, что последние общие предки всех змей имели зрение, чувствительное к ультрафиолетовому излучению, но у большинства змей, охота которых зависит от зрения при дневном свете, развились линзы, которые действуют как солнцезащитные очки и фильтруют ультрафиолетовый свет, что, вероятно, также обостряет их зрение. зрение за счет улучшения контраста . [58] [59]
Кожа
Кожа змеи покрыта чешуей . Вопреки популярному представлению о том, что змеи склизкие (из-за того, что змей можно спутать с червями ), змеиная кожа имеет гладкую сухую текстуру. Большинство змей используют для передвижения специальные чешуйки на брюшке, позволяющие им цепляться за поверхности. Чешуя тела может быть гладкой, килевидной или зернистой. Веки змеи представляют собой прозрачные «очковые» чешуйки, также известные как брилле , которые остаются постоянно закрытыми.
У змеи кожа модифицирована для ее специализированной формы передвижения. Между внутренним и внешним слоями находится дерма, содержащая все пигменты и клетки, составляющие отличительный рисунок и цвет змеи. Эпидермис, или внешний слой, состоит из вещества, называемого кератином, которое у млекопитающих является тем же основным материалом, из которого формируются ногти, когти и волосы. Кератиновый эпидермис змеи обеспечивает ей броню, необходимую для защиты внутренних органов и уменьшения трения при прохождении по камням. Части этой кератиновой брони более грубые, чем другие. Менее ограниченная часть перекрывает переднюю часть шкалы под ней. Между ними лежит отогнутый соединительный материал, тоже из кератина, также входящий в состав эпидермиса. Этот сложенный назад материал поддается изгибам, когда змея колеблется или поедает предметы, превышающие окружность ее тела. [52] [ нужна страница ]
Сбрасывание чешуи называется шелушением (или, при обычном использовании, линькой или шелушением ). Змеи сбрасывают весь внешний слой кожи целиком. [60] Змеиные чешуйки не являются отдельными, а являются продолжением эпидермиса , поэтому во время каждой линьки они сбрасываются не отдельно, а в виде целого внешнего слоя, подобно носку, выворачиваемому наизнанку. [61]
Змеи имеют большое разнообразие окраски кожи, которая часто связана с поведением, например, с тенденцией бежать от хищников. Змеи, которые подвергаются высокому риску нападения хищников, обычно имеют однотонную окраску или имеют продольные полосы, что дает мало ориентиров для хищников и позволяет змее уйти незамеченной. Обычные змеи обычно применяют активную стратегию охоты, поскольку их образ жизни позволяет им посылать жертве мало информации о движении. Пятнистые змеи обычно используют стратегии, основанные на засадах, вероятно, потому, что это помогает им сливаться с окружающей средой с объектами неправильной формы, такими как палки или камни. Пятнистый рисунок также может помочь змеям сливаться с окружающей средой. [62]
Форма и количество чешуек на голове, спине и брюхе часто являются характерными и используются в таксономических целях. Чешуи названы в основном в зависимости от их положения на теле. У «продвинутых» ( ценофидийских ) змей широкие брюшные чешуи и ряды спинных чешуек соответствуют позвонкам , что позволяет их подсчитывать без необходимости вскрытия .
Линька

Линька (или «шелушение») служит нескольким целям: она позволяет заменить старую, изношенную кожу и может быть синхронизирована с циклами спаривания, как и у других животных. Линька происходит периодически на протяжении всей жизни змеи. Перед каждой линькой змея регулирует свой рацион и ищет надежное укрытие. Непосредственно перед линькой кожа становится серой, а глаза змеи становятся серебристыми. Внутренняя поверхность старой кожи разжижается, в результате чего она отделяется от новой кожи под ней. Через несколько дней глаза проясняются, и змея вылезает из старой кожи, которая трескается. Змея трется своим телом о грубые поверхности, чтобы помочь сбросить старую кожу. Во многих случаях отброшенная кожа отслаивается назад по телу от головы до хвоста целиком, как если бы с книги снимали суперобложку, обнажая новый, более крупный и яркий слой кожи, образовавшийся под ней. [61] [63] Обновление кожи путем линьки предположительно увеличивает массу некоторых животных, например насекомых, но в случае со змеями это оспаривается. [61] [64] Линька кожи может выделять феромоны и оживлять цвет и рисунок кожи, повышая привлекательность партнеров. [65]
Змеи могут линять четыре-пять раз в год, в зависимости от погодных условий, запасов пищи, возраста змеи и других факторов. [52] [ нужна страница ] [63] Теоретически можно идентифицировать змею по слепку кожи, если она практически не повреждена. [61] Мифологические ассоциации змей с символами врачевания и медицины , изображенные на Жезле Асклепия , являются производными от линьки. [66]
Можно попытаться определить пол змеи, если у вида нет явного полового диморфизма, путем подсчета чешуек. Клоаку . исследуют и измеряют по чешуе субкаудальной [67] Подсчет чешуек определяет, является ли змея самцом или самкой, поскольку гемипены исследуемого самца обычно длиннее. [67] [ нужны разъяснения ]
Скелет

Череп змеи отличается от ящерицы по нескольким признакам. У змей челюсти более гибкие, то есть вместо места соединения верхней и нижней челюсти челюсти змеи соединены костным шарниром, который называется квадратной костью . Между двумя половинками нижней челюсти у подбородка имеется эластичная связка, позволяющая осуществить разделение. Это позволяет змее заглатывать пищу большего размера, пропорционально ее размеру, и дольше обходиться без нее, поскольку за одно кормление змеи поглощают относительно больше пищи. [68] Поскольку стороны нижней челюсти могут двигаться независимо друг от друга, змея, опирающаяся челюстью на поверхность, обладает стереослуховым восприятием , которое используется для определения положения добычи. Путь челюсть-квадрат- стремечко способен обнаруживать вибрации в ангстремном масштабе, несмотря на отсутствие наружного уха и отсутствие механизма согласования импеданса , обеспечиваемого косточками у других позвоночных. [69] [70] В черепе змеи мозг хорошо защищен. Поскольку ткани мозга могут быть повреждены через небо, эта защита особенно ценна. Твердый и цельный нейрокраниум змей закрыт спереди. [52] [ нужна страница ] [71]

Скелет большинства змей состоит исключительно из черепа, подъязычной кости , позвоночного столба и ребер, хотя у хенофидийских змей сохраняются остатки таза и задних конечностей. Подъязычная кость — небольшая кость, расположенная сзади и вентральнее черепа, в «шейной» области, которая служит местом прикрепления мышц языка змеи, как и у всех других четвероногих . В позвоночнике насчитывается от 200 до 400 позвонков, а иногда и больше. Каждый из позвонков тела имеет по два ребра, сочленяющихся с ними. Хвостовые позвонки сравнительно немногочисленны (часто менее 20% от общего числа) и лишены ребер. Позвонки имеют выступы, обеспечивающие прочное прикрепление мышц, что позволяет передвигаться без помощи конечностей.
Каудальная автотомия (самоампутация хвоста) — особенность, встречающаяся у некоторых ящериц, — у большинства змей отсутствует. [72] В тех редких случаях, когда у змей она действительно существует, каудальная автотомия является межпозвоночной (что означает разделение соседних позвонков), в отличие от таковой у ящериц, которая является внутрипозвоночной, т. е. разрыв происходит вдоль заранее определенной плоскости перелома, присутствующей на позвонке. [73] [74]
У некоторых змей, особенно удавов и питонов, имеются остатки задних конечностей в виде пары тазовых шпор . Эти небольшие когтеобразные выступы на каждой стороне клоаки являются внешней частью рудиментарного скелета задней конечности, который включает остатки подвздошной и бедренной костей.
Змеи — полифиодонты , зубы которых постоянно сменяются. [75]
Внутренние органы

- пищевод
- трахея
- трахея легкие
- рудиментарное левое легкое
- правое легкое
- сердце
- печень
- желудок
- воздушный мешок
- желчный пузырь
- поджелудочная железа
- селезенка
- кишечник
- яички
- почки
Змеи и другие рептилии, не относящиеся к архозаврам ( крокодилы , динозавры + птицы и их союзники), имеют трехкамерное сердце, которое управляет системой кровообращения через левое и правое предсердия и один желудочек. [76] Внутри желудочек разделен на три взаимосвязанные полости: артериальную полость, легочную полость и венозную полость. [77] Венозная полость получает дезоксигенированную кровь из правого предсердия, а артериальная полость получает насыщенную кислородом кровь из левого предсердия. Под венозной полой полостью находится легочная полость, которая перекачивает кровь в легочный ствол. [78]
Сердце змеи заключено в мешочек, называемый перикардом расположенный в развилке бронхов , . Сердце способно двигаться благодаря отсутствию диафрагмы; эта регулировка защищает сердце от потенциального повреждения, когда крупная проглоченная добыча проходит через пищевод . Селезенка и прикрепляется к желчному пузырю и поджелудочной железе фильтрует кровь. Тимус , расположенный в жировой ткани над сердцем, отвечает за выработку иммунных клеток в крови. Сердечно-сосудистая система змей уникальна наличием почечной портальной системы, в которой кровь из хвоста змеи проходит через почки, прежде чем вернуться в сердце. [79]
Кровеносная система змеи в основном аналогична системе кровообращения любого другого позвонка. Однако змеи не регулируют внутреннюю температуру своей крови. Названные хладнокровными, змеи на самом деле имеют кровь, которая реагирует на изменение температуры окружающей среды. Змеи могут регулировать температуру крови, двигаясь. Слишком долго под прямыми солнечными лучами кровь змей нагревается до предела. Оставшись во льду или снегу, змея может замерзнуть. В умеренных зонах с ярко выраженными сезонными изменениями змеи, сидевшие вместе, приспособились к натиску зимы. [52] [ нужна страница ]
Рудиментарное часто маленькое, а иногда даже отсутствует, поскольку трубчатые тела змей требуют , левое легкое чтобы все их органы были длинными и тонкими. [79] У большинства видов функционально только одно легкое. Это легкое содержит васкуляризированную переднюю часть и заднюю часть, которая не участвует в газообмене. [79] Это «мешотчатое легкое» используется в гидростатических целях для регулирования плавучести у некоторых водных змей, и его функция остается неизвестной у наземных видов. [79] Многие парные органы, такие как почки или репродуктивные органы , расположены внутри тела в шахматном порядке, один расположен впереди другого. [79]
Змея с ее особым расположением органов может достичь большей эффективности. [ по сравнению с? ] Например, легкие содержат в своей части, ближайшей к голове и горлу, орган, потребляющий кислород, а другая половина используется для резерва воздуха. Расположение пищевод-желудок-кишечник представляет собой прямую линию. Он заканчивается там, где открываются кишечный, мочевой и репродуктивный пути, в камере, называемой клоакой. [52] [ нужна страница ]
У змей нет лимфатических узлов . [79]
Яд

Кобры, гадюки и близкородственные виды используют яд, чтобы обездвижить, ранить или убить свою добычу. Яд представляет собой модифицированную слюну , доставляемую через клыки . [26] [80] : 243 Клыки «продвинутых» ядовитых змей, таких как гадюки и элапиды, полые, что позволяет более эффективно вводить яд, а клыки змей с задними клыками, таких как бумсланг, просто имеют бороздку на заднем крае для направления яда в рану. Змеиные яды часто специфичны для добычи, и их роль в самозащите второстепенна. [26] [80] : 243
Яд, как и все выделения слюны, является предварительным средством, которое инициирует расщепление пищи на растворимые соединения, способствуя правильному пищеварению. Даже неядовитые укусы змей (как и укусы любого животного) вызывают повреждение тканей. [26] [80] : 209
Некоторые птицы, млекопитающие и другие змеи (например, королевские змеи ), охотящиеся на ядовитых змей, выработали устойчивость и даже иммунитет к определенным ядам. [26] : 243 Ядовитые змеи включают три семейства змей и не составляют формальной таксономической классификационной группы.
Разговорный термин «ядовитая змея» , как правило, является неверным обозначением змей. Яд вдыхается или проглатывается, тогда как яд, вырабатываемый змеями, впрыскивается жертве через клыки. [81] Однако есть два исключения: Rhabdophis изолирует токсины от жаб, которых он поедает, а затем выделяет их из затылочных желез, чтобы отпугивать хищников; а небольшая необычная популяция подвязочных змей в американском штате Орегон сохраняет в печени достаточно токсинов от проглоченных тритонов , чтобы быть эффективно ядовитыми для мелких местных хищников (таких как вороны и лисы ). [82]
Змеиные яды представляют собой сложную смесь белков . [80] и хранятся в ядовитых железах на затылке. [82] У всех ядовитых змей эти железы открываются протоками в бороздчатые или полые зубы верхней челюсти. [26] : 243 [81] Белки потенциально могут представлять собой смесь нейротоксинов (поражающих нервную систему), гемотоксинов (поражающих систему кровообращения), цитотоксинов (поражающих непосредственно клетки), бунгаротоксинов (связанных с нейротоксинами, но также напрямую влияющих на мышечную ткань) и многие другие токсины, по-разному воздействующие на организм. [81] [80] Почти весь змеиный яд содержит гиалуронидазу — фермент, обеспечивающий быструю диффузию яда. [26] : 243
Ядовитые змеи, использующие гемотоксины, обычно имеют клыки в передней части рта, что облегчает им введение яда в жертву. [80] [81] Некоторые змеи, использующие нейротоксины (например, мангровая змея ), имеют клыки в задней части рта, загнутые назад. [83] Из-за этого змее становится сложнее использовать свой яд, а ученым - доить ее. [81] Однако элапиды, такие как кобры и крайты, являются протероглифами — у них есть полые клыки, которые нельзя поднять к передней части рта, и они не могут «колоть», как гадюка. Они должны фактически укусить жертву. [26] : 242
Было высказано предположение, что все змеи могут быть в определенной степени ядовитыми, причем безвредные змеи имеют слабый яд и не имеют клыков. [84] Согласно этой теории, большинство змей, помеченных как «неядовитые», считаются безвредными, поскольку у них либо отсутствует метод доставки яда, либо они не способны доставить достаточно яда, чтобы подвергнуть опасности человека. Теория постулирует, что змеи, возможно, произошли от общего предка ящерицы, который был ядовитым, а также что ядовитые ящерицы, такие как монстр Гила , бисерная ящерица , вараны и ныне вымершие мозазавры , возможно, произошли от этого же общего предка. Они разделяют эту « ядовитую кладу » с различными другими видами ящеров .
Ядовитые змеи делятся на два таксономических семейства:
- Элапиды – кобры , в том числе королевские кобры , крайты , мамбы , австралийские медноголовые , морские и коралловые змеи . [83]
- Гадюки – гадюки, гремучие змеи , медноголовые / хлопотные змеи и бушмастеры . [83]
Существует третье семейство, включающее опистоглифных (заднезубых) змей (как и большинство других видов змей):
- Колубриды – бумсланги , древесные ужа , виноградные змеи , кошачьи змеи , хотя не все колубриды ядовиты. [26] : 209 [83]
Воспроизведение
Хотя змеи используют широкий спектр репродуктивных способов, все они используют внутреннее оплодотворение . Это достигается с помощью парных раздвоенных гемипенов , которые в перевернутом виде хранятся в хвосте самца. [85] самки Гемипены часто имеют бороздки, крючки или шипы и предназначены для захвата стенок клоаки . [86] [85] Клитор самки змеи состоит из двух структур, расположенных между клоакой и ароматическими железами. [87]
Большинство видов змей откладывают яйца , которые бросают вскоре после кладки. Однако некоторые виды (например, королевская кобра) строят гнезда и после инкубации остаются рядом с вылупившимися птенцами. [85] Большинство питонов обвивают кладки яиц и остаются с ними до тех пор, пока не вылупятся. [88] Самка питона не будет покидать яйца, разве что изредка погреться на солнышке или попить воды. Она даже будет «дрожать», чтобы выработать тепло для инкубации яиц. [88]
Некоторые виды змей являются яйцеживородящими и сохраняют яйца в своем теле до тех пор, пока они не будут почти готовы вылупиться. [89] [90] Некоторые виды змей, такие как удав и зеленая анаконда, являются полностью живородящими и выкармливают детенышей через плаценту , а также через желточный мешок ; это весьма необычно для рептилий и обычно встречается у акул-реквиемов или плацентарных млекопитающих . [89] [90] Сохранение яиц и живорождение чаще всего связаны с более холодной средой. [85] [90]

Половой отбор у змей демонстрируется 3000 видами, каждый из которых использует разные тактики поиска партнеров. [91] Ритуальный бой между самцами за самок, с которыми они хотят спариваться , включает в себя топпинг - поведение, демонстрируемое большинством гадюк, при котором один самец обвивает вертикально поднятую переднюю часть тела своего противника и заставляет его опускаться вниз. Когда змеи переплетаются, часто происходит укус шеи. [92]
Факультативный партеногенез
Партеногенез — естественная форма размножения, при которой рост и развитие зародышей происходят без оплодотворения. Agkistrodon contortrix (медноголовый) и Agkistrodon piscivorus (ватноротый) могут размножаться путем факультативного партеногенеза , то есть они способны переключаться с полового способа размножения на бесполый . [93] Наиболее вероятным типом партеногенеза является аутомиксис с терминальным слиянием, процесс, при котором два терминальных продукта одного и того же мейоза сливаются с образованием диплоидной зиготы . по всему геному Этот процесс приводит к гомозиготности , экспрессии вредных рецессивных аллелей и часто к аномалиям развития. Как рожденные в неволе, так и дикие медноголовые и ватные ротики, по-видимому, способны к этой форме партеногенеза. [93]
Размножение у чешуйчатых рептилий почти исключительно половое. Мужчины обычно имеют пару ZZ хромосом, определяющих пол, а женщины - пару ZW. Однако колумбийский радужный удав ( Epicrates maurus ) также может размножаться путем факультативного партеногенеза, что приводит к образованию потомства самок WW. [94] Самки WW, вероятно, производятся терминальным автомиксисом.
Эмбриональное развитие

Эмбриональное развитие змеи первоначально следует тем же этапам, что и эмбрионы любого позвоночного . Эмбрион змеи начинается с зиготы , подвергается быстрому делению клеток, образует зародышевый диск , также называемый бластодиском, затем подвергается гаструляции , нейруляции и органогенезу . [96] Деление и пролиферация клеток продолжаются до тех пор, пока не разовьется ранний эмбрион змеи и не будет наблюдаться типичная форма тела змеи. [96] Эмбриологическое развитие змей отличается от других позвоночных множественными особенностями, двумя важными факторами являются удлинение тела и отсутствие развития конечностей.

Удлинение тела змей сопровождается значительным увеличением количества позвонков (у мышей их 60, а у змей — более 300). [95] Это увеличение позвонков происходит из-за увеличения сомитов во время эмбриогенеза, что приводит к увеличению числа развивающихся позвонков. [95] Сомиты образуются в пресомитной мезодерме благодаря набору колебательных генов, которые управляют часами сомитогенеза . Часы сомитогенеза змей работают с частотой, в 4 раза превышающей таковую у мыши (после поправки на время развития), создавая больше сомитов и, следовательно, больше позвонков. [95] Считается, что эта разница в тактовой частоте вызвана различиями в Lunatic экспрессии маргинального гена , гена, участвующего в часах сомитогенеза. [97]
Существует обширная литература, посвященная развитию/отсутствию развития конечностей у эмбрионов змей и экспрессии генов, связанной с различными стадиями. У базальных змей , таких как питон, эмбрионы на раннем этапе развития имеют зачаток задних конечностей , который развивается с некоторым количеством хряща и хрящевым тазовым элементом, однако перед вылуплением он дегенерирует. [98] Наличие рудиментарного развития предполагает, что некоторые змеи все еще подвергаются редукции задних конечностей, прежде чем они будут уничтожены. [99] У базальных змей нет зачатков передних конечностей, а также нет примеров зарождения зачатков передних конечностей змей у эмбриона, поэтому мало что известно об утрате этого признака. [99] Недавние исследования показывают, что уменьшение задних конечностей может быть связано с мутациями в энхансерах гена SSH . [99] однако другие исследования показали, что мутации в генах Hox или их энхансерах могут способствовать потере конечностей у змей. [95] Поскольку многочисленные исследования обнаружили доказательства того, что разные гены играют роль в потере конечностей у змей, вполне вероятно, что множественные мутации генов имели аддитивный эффект, приводящий к потере конечностей у змей. [100]
Поведение и история жизни

Зимний покой
В регионах, где зимы слишком холодные, чтобы змеи могли их терпеть, оставаясь активными, местные виды вступают в период брумации . В отличие от спячки , в которой спящие млекопитающие фактически спят, брумирующие рептилии бодрствуют, но неактивны. Отдельные змеи могут скапливаться в норах, под грудами камней или внутри упавших деревьев, либо большое количество змей может скапливаться в гибернакуле .
Кормление и диета



Все змеи строго плотоядны и охотятся на мелких животных, включая ящериц, лягушек, других змей, мелких млекопитающих, птиц, яйца, рыбу, улиток, червей и насекомых. [26] : 81 [27] [101] Змеи не могут кусать или разрывать пищу на куски, поэтому им приходится заглатывать добычу целиком. На пищевые привычки змеи во многом влияет размер тела; более мелкие змеи едят более мелкую добычу. Молодые питоны могут начать питаться ящерицами или мышами, а во взрослом возрасте превратиться в мелких оленей или антилоп.
змеи Челюсть представляет собой сложную структуру. Вопреки распространенному мнению, что змеи могут вывихивать челюсти, у них чрезвычайно гибкая нижняя челюсть , две половины которой не соединены жестко, и множество других суставов черепа, которые позволяют змее открывать пасть достаточно широко, чтобы глотать. добычу целиком, даже если она в диаметре больше самой змеи. [101] Например, африканская змея, питающаяся яйцами, имеет гибкие челюсти, приспособленные для поедания яиц, размер которых намного превышает диаметр ее головы. [26] : 81 У этой змеи нет зубов, но есть костные выступы на внутреннем крае позвоночника , которые она использует, чтобы разбить скорлупу при поедании яиц. [26] : 81
Большинство змей питаются разнообразными животными, но у определенных видов существует некоторая специализация. Королевские кобры и австралийский хоккей с мячом поедают других змей. У видов семейства Pareidae больше зубов на правой стороне рта, чем на левой, поскольку они в основном охотятся на улиток, а раковины обычно вращаются по часовой стрелке. [26] : 184 [102] [103]
У некоторых змей есть ядовитый укус, которым они убивают добычу перед тем, как ее съесть. [101] [104] Другие змеи убивают свою добычу сжиманием , [101] а некоторые проглатывают свою добычу, когда она еще жива. [26] : 81 [101]
процесс пищеварения ; После еды змеи впадают в спячку, позволяя начать [67] это интенсивная деятельность, особенно после поедания крупной добычи. У видов, которые питаются лишь спорадически, весь кишечник между приемами пищи переходит в сокращенное состояние для сохранения энергии. Затем пищеварительная система «настраивается» на полную мощность в течение 48 часов после потребления добычи. Будучи экзотермическими («хладнокровными»), температура окружающей среды играет важную роль в процессе пищеварения. Идеальная температура для переваривания пищи змеями — 30 °C (86 °F). В пищеварении у змей задействовано огромное количество метаболической энергии, например, температура поверхности тела южноамериканской гремучей змеи ( Crotalus durissus ) увеличивается на целых 1,2 ° C (2,2 ° F) во время процесса пищеварения. [105] Если змея потревожена после того, как недавно поела, она часто срыгивает добычу, чтобы избежать предполагаемой угрозы. Когда процесс пищеварения не нарушен, он очень эффективен; змеи пищеварительные ферменты растворяют и поглощают все, кроме волос (или перьев) и когтей жертвы, которые выводятся из организма вместе с отходами .
Капюшон и плевание
Капюшон (расширение области шеи) является визуальным сдерживающим фактором, чаще всего наблюдаемым у кобр (елапид), и в первую очередь контролируется реберными мышцами. [106] Надевание капюшона может сопровождаться плевком яда в сторону угрожающего объекта. [107] и создание специального звука; шипение. Исследования кобр в неволе показали, что 13–22% длины тела приподнимается во время надевания капюшона. [108]
Передвижение
Отсутствие конечностей не препятствует передвижению змей. Они разработали несколько различных способов передвижения для работы в определенных условиях. В отличие от походки конечностей животных, которые образуют континуум, каждый способ передвижения змей дискретен и отличается от других; переходы между режимами резкие. [109] [110]
Боковая волнистость

Боковая волнистость — единственный способ передвижения в воде и наиболее распространенный способ передвижения на суше. [110] В этом режиме тело змеи попеременно изгибается влево и вправо, что приводит к серии «волн», движущихся назад. [109] Хотя это движение кажется быстрым, редко регистрируется, что змеи двигаются со скоростью более двух длин тела в секунду, а часто и намного меньше. [111] Этот способ передвижения имеет те же чистые затраты на транспортировку (калории, сжигаемые на метр перемещения), что и бег ящериц той же массы. [112]
Наземные боковые волнистости - наиболее распространенный способ наземного передвижения большинства видов змей. [109] В этом режиме движущиеся назад волны сталкиваются с точками контакта с окружающей средой, такими как камни, ветки, неровности почвы и т. д. [109] Каждый из этих объектов окружающей среды, в свою очередь, генерирует силу реакции, направленную вперед и к средней линии змеи, что приводит к толчку вперед, в то время как боковые компоненты уравновешиваются. [113] Скорость этого движения зависит от плотности точек нажатия в окружающей среде, средняя плотность составляет около 8 [ нужны разъяснения ] по длине змеи идеально. [111] Скорость волны точно такая же, как и скорость змеи, и в результате каждая точка тела змеи повторяет путь точки перед ней, что позволяет змеям перемещаться через очень густую растительность и небольшие отверстия. [113]
Во время плавания волны становятся больше по мере движения вниз по телу змеи, и волна движется назад быстрее, чем змея движется вперед. [114] Тяга создается за счет прижатия тела к воде, что приводит к наблюдаемому скольжению. Несмотря на общее сходство, исследования показывают, что характер мышечной активации различен при водных и наземных боковых волнах, что дает основание называть их отдельными режимами. [115] Все змеи могут совершать боковые движения вперед (с волнами, движущимися назад), но наблюдалось только морские змеи, меняющие движение (движущиеся назад с волнами, движущимися вперед). [109]
боковой ветер

Чаще всего используется колубридными змеями ( колобридами , элапидами и гадюками ), когда змее приходится двигаться в среде, в которой нет неровностей, на которые можно было бы наталкиваться (что делает невозможными боковые волнистости), например, на скользкой илистой равнине или песчаной дюне. модифицированная форма боковой волнистости, при которой все сегменты тела, ориентированные в одном направлении, остаются в контакте с землей, в то время как другие сегменты поднимаются вверх, что приводит к своеобразному «перекатывающемуся» движению. [116] [117] Сайдвиндер движется вперед, создавая петлю, а затем подтягиваясь за нее. Опуская голову, змея получает рычаг, выпрямляясь и прижимаясь к земле, она продвигается вперед под углом, который готов к следующему прыжку. Голова и петля — это, по сути, две ноги, на которых ходит змея. Тело змеи, выглядящее примерно перпендикулярно ее направлению, может сбить с толку наблюдателя, поскольку предвзятое мнение может привести к тому, что можно связать движение змеи с головой, которая ведет, и телом, которое следует за ней. Кажется, что змея движется вбок, но куда именно движется змея, куда она хочет идти, голова ясно указывает. Змея оставляет за собой след, похожий на серию крючков, идущих один за другим. Змеи могут двигаться назад, чтобы отступить от врага, хотя обычно они этого не делают. [52] [ нужна страница ] Этот способ передвижения позволяет преодолеть скользкость песка или грязи, отталкиваясь только статическими частями тела, тем самым сводя к минимуму скольжение. [116] Статичность точек контакта можно показать по следам извивающейся змеи, на которых виден отпечаток каждой чешуи на брюхе, без всякого размытия. Этот способ передвижения имеет очень низкую калорийность, менее 1/3 . стоимости перемещения ящерицы на такое же расстояние [112] Вопреки распространенному мнению, нет никаких доказательств того, что боковой ветер связан с горячим песком. [116]
Концертина
Когда точки толчка отсутствуют, но недостаточно места для использования бокового движения из-за боковых ограничений, например, в туннелях, змеи полагаются на движение гармошкой. [109] [117] В этом режиме змея прижимает заднюю часть своего тела к стене туннеля, в то время как передняя часть змеи вытягивается и выпрямляется. [116] Затем передняя часть сгибается и образует точку опоры, а задняя выпрямляется и тянется вперед. Этот способ передвижения медленный и очень трудоемкий: затраты на боковые колебания на том же расстоянии в семь раз превышают затраты. [112] Такая высокая стоимость обусловлена повторяющимися остановками и стартами частей тела, а также необходимостью использования активных мышечных усилий для упирания в стены туннеля.
Древесный

Передвижение змей в древесных местообитаниях изучается лишь недавно. [118] Находясь на ветвях деревьев, змеи используют несколько способов передвижения в зависимости от вида и текстуры коры. [118] В общем, змеи будут использовать модифицированную форму передвижения гармошкой на гладких ветвях, но будут совершать боковые волнистые движения, если доступны точки контакта. [118] Змеи движутся быстрее по маленьким ветвям и при наличии точек контакта, в отличие от животных с конечностями, которые лучше себя чувствуют на больших ветвях с небольшим «беспорядком». [118]
Планирующие змеи ( Chrysopelea ) Юго-Восточной Азии взлетают с кончиков ветвей, расправляя ребра и совершая волнообразные движения в стороны, скользя между деревьями. [116] [119] [120] Эти змеи могут выполнять контролируемое планирование на сотни футов в зависимости от высоты запуска и даже поворачиваться в воздухе. [116] [119]
Прямолинейный
Самый медленный способ передвижения змеи - прямолинейное движение, которое также является единственным, при котором змее не нужно сгибать свое тело в стороны, хотя она может делать это при повороте. [121] В этом режиме чешуя живота поднимается и вытягивается вперед, а затем опускается вниз и тело натягивается на нее. Волны движения и стаза проходят назад, вызывая серию пульсаций на коже. [121] Ребра змеи не движутся при этом способе передвижения, и этот метод чаще всего используется крупными питонами , удавами и гадюками при преследовании добычи на открытой местности, поскольку движения змеи тонкие и их труднее обнаружить добыче таким образом. . [116]
Взаимодействие с людьми
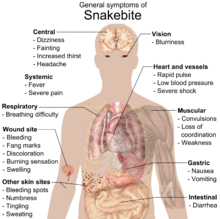
Кусать

Змеи обычно не охотятся на людей. Если они не испуганы и не ранены, большинство змей предпочитают избегать контакта и не нападают на людей. Неядовитые змеи, за исключением крупных удавов, не представляют угрозы для человека. Укус неядовитой змеи обычно безвреден; их зубы не приспособлены для разрывания или нанесения глубокой колотой раны, а скорее для хватания и удержания. Хотя при укусе неядовитой змеи существует возможность заражения и повреждения тканей, ядовитые змеи представляют гораздо большую опасность для человека. [26] : 209 Всемирная организация здравоохранения (ВОЗ) относит укус змеи к категории «других забытых состояний». [124]
Документально подтвержденные случаи смерти в результате укусов змей редки. Несмертельные укусы ядовитых змей могут привести к необходимости ампутации конечности или ее части. Из примерно 725 видов ядовитых змей во всем мире только 250 способны убить человека одним укусом. В Австралии в среднем случается только один смертельный укус змеи в год. В Индии за один год регистрируется 250 000 укусов змей, при этом первоначально зарегистрировано до 50 000 случаев смерти. [125] По оценкам ВОЗ, ежегодно в результате укусов змей умирают порядка 100 000 человек, а укусы змей ежегодно вызывают примерно в три раза больше ампутаций и других необратимых инвалидностей. [126]
Укусы змей серьезно угрожают здоровью людей, особенно в районах, где имеется большое разнообразие змей и ограничен доступ к медицинской помощи, таких как регион тропических лесов Амазонки в Южной Америке. [127] Змеиный укус классифицируется Всемирной организацией здравоохранения (ВОЗ) как «другие запущенные состояния». [128] Хотя зарегистрировано не так много смертей от укусов змей, укусы могут вызвать серьезные осложнения и необратимые нарушения. [128] Самым успешным средством лечения змеиных укусов по-прежнему является противоядие, изготовленное из змеиного яда. [128] Однако доступ к противояду сильно различается в зависимости от местоположения: сельские районы часто испытывают трудности как с ценой, так и с доступностью. [129] Клинические исследования, приготовление сыворотки и извлечение яда относятся к числу сложных процедур, связанных с производством противоядия. [129] Разработка альтернативных методов лечения, а также повышение доступности противоядия имеют важное значение для снижения глобального воздействия укусов змей на население. [130]
Заклинатели змей

В некоторых частях света, особенно в Индии, заклинание змей представляет собой придорожное представление, проводимое заклинателем. В таком представлении заклинатель змей несет корзину со змеей, которую он, по-видимому, очаровывает, играя мелодии на своем похожем на флейту музыкальном инструменте, на что змея реагирует. [131] На самом деле змея реагирует на движение флейты, а не на звук, который она издает, поскольку у змей нет внешних ушей (хотя внутренние уши у них есть). [131]
Закон о защите дикой природы 1972 года в Индии технически запрещает заклинание змей по причине уменьшения жестокого обращения с животными. Другие типы заклинателей змей используют шоу змей и мангустов , где два животных инсценируют драку; однако это случается не очень часто, поскольку животные могут быть серьезно ранены или убиты. Профессия заклинания змей вымирает в Индии из-за конкуренции со стороны современных форм развлечений и законов об окружающей среде, запрещающих эту практику. Многие индийцы никогда не видели очаровательных змей, и это становится сказкой прошлого. [131] [132] [133] [134]
Захват
Племя ирулас из Андхра-Прадеша и Тамил Наду в Индии занималось охотниками-собирателями в жарких, сухих равнинных лесах и на протяжении поколений практиковало искусство ловли змей. У них есть обширные знания о змеях в этой области. Обычно они ловят змей с помощью простой палки. Ранее Ирулас ловили тысячи змей для производства змеиной кожи. После полного запрета производства змеиной кожи в Индии и защиты всех змей в соответствии с Законом о защите дикой природы Индии 1972 года они сформировали Кооператив ловцов змей Ирула и переключились на ловлю змей для удаления яда, выпуская их в дикую природу после четыре извлечения. Собранный таким образом яд используется для производства спасительного противоядия, биомедицинских исследований и других лекарственных средств. [135] поедают Известно также , что ирулы некоторых пойманных ими змей и очень полезны при истреблении крыс в деревнях.
Несмотря на существование заклинателей змей, были также профессиональные ловцы змей или дрессировщики . В современном отлове змей герпетолог использует длинную палку с V-образным концом. Некоторые ведущие телешоу, такие как Билл Хааст , Остин Стивенс , Стив Ирвин и Джефф Корвин , предпочитают ловить их голыми руками.
Потребление

Потребление змеиного мяса и связанных с ним товаров является отражением многих культур по всему миру, особенно в азиатских странах, таких как Китай, Тайвань, Таиланд, Индонезия, Вьетнам и Камбоджа. Из-за предполагаемой пользы для здоровья и свойств афродизиака змеиное мясо часто считается деликатесом и употребляется в пищу. [136] Принято пить вино, смешанное с кровью змей, чтобы повысить мужественность и энергию. [136] Традиционная китайская медицина считает, что змеиное вино, традиционный напиток, настоянный на целых змеях, имеет лечебные свойства. [136] Истоки змеиного вина лежат в китайской культуре. Однако использование змеиных товаров порождает моральные вопросы относительно сохранения и благополучия животных. [137] Важно уделять внимание и регулировать устойчивый промысел змей в пищу человеку, особенно в районах, где популяция змей сокращается в результате деградации среды обитания и чрезмерной эксплуатации. [137]
Домашние животные
В западном мире некоторых змей держат в качестве домашних животных, особенно послушных видов, таких как шаровидный питон и кукурузная змея . Чтобы удовлетворить спрос, разведения в неволе была развита индустрия . Змеи, выращенные в неволе, считаются более предпочтительными, чем особи, пойманные в дикой природе, и, как правило, из них получаются лучшие домашние животные. [138] По сравнению с более традиционными видами домашних животных, змеи могут быть домашними животными, не требующими особого ухода; им требуется минимальное пространство, поскольку длина большинства распространенных видов не превышает 5 футов (1,5 м), и их можно кормить относительно нечасто - обычно один раз в пять-четырнадцать дней. Продолжительность жизни некоторых змей при правильном уходе превышает 40 лет.
Символизм


В древней Месопотамии , Нира бог-посланник Иштарана , изображался в виде змеи на кудуррусе , или пограничных камнях . [139] Изображения двух переплетенных змей распространены в шумерском и неошумерском искусстве. [139] и до сих пор время от времени появляются на цилиндрических печатях и амулетах вплоть до тринадцатого века до нашей эры. [139] Рогатая гадюка ( Cerastes cerastes ) появляется в касситском и неоассирийском кудуррусе . [139] и упоминается в ассирийских текстах как магическое защитное существо. [139] Драконоподобное существо с рогами, телом и шеей змеи, передними лапами льва и задними лапами птицы появляется в месопотамском искусстве от аккадского периода до эллинистического периода (323–31 до н.э.). [139] Это существо, известное на аккадском языке как mušḫuššu , что означает «яростный змей», использовалось как символ определенных божеств, а также как общая защитная эмблема. [139] Судя по всему, изначально он был слугой бога подземного мира Ниназу . [139] но позже стал помощником хурритского бога бури Тишпака , а также, позднее, сына Ниназу Нингишзида , вавилонского национального бога Мардука , бога-писца Набу и ассирийского национального бога Ашура. [139]
В египетской истории змея занимает первостепенную роль, наряду с нильской коброй, украшавшей в древности корону фараона. Ему поклонялись как одному из богов, а также использовали в зловещих целях: убийство противника и ритуальное самоубийство ( Клеопатра ). [140] Уроборос . был известным древнеегипетским символом змеи, проглотившей собственный хвост [141] Предшественником уробороса был «Многоликий», [141] змей с пятью головами, который, согласно «Амдуату» , старейшей из сохранившихся «Книг о загробной жизни» , как говорят, обвился вокруг трупа бога Солнца Ра, защищая его. [141] изображение «настоящего» уробороса происходит из позолоченных святынь в гробнице Тутанхамона Самое раннее сохранившееся . [141] в качестве символа. -гностиками В первые века нашей эры уроборос был принят христианами [141] а глава 136 «Пистис София» , раннего гностического текста, описывает «великого дракона, чей хвост находится во рту». [141] В средневековой алхимии уроборос стал типичным западным драконом с крыльями, ногами и хвостом. [141]
В Библии царь аммонитянский Нахаш , имя которого означает «Змей», изображен весьма негативно, как особо жестокий и презренный враг древних евреев.

Древние греки использовали Горгонейон , изображение отвратительного лица со змеями вместо волос, как апотропный символ для отражения зла. [142] В греческом мифе, описанном Псевдо-Аполлодором в его «Библиотеке» , Медуза была Горгоной со змеями вместо волос, чей взгляд обращал в камень всех, кто смотрел на нее, и была убита героем Персеем . [143] [144] [145] поэта Овидия римского В «Метаморфозах» говорится , что Медуза когда-то была прекрасной жрицей Афины , которую Афина превратила в змееволосого монстра после того, как она была изнасилована богом Посейдоном в храме Афины. [146] В другом мифе, на который ссылается беотийский поэт Гесиод и подробно описывает Псевдо-Аполлодор, говорится, что герой Геракл убил Лернейскую гидру . [147] [148] многоголовый змей, обитавший в болотах Лерны . [147] [148]
В легендарном рассказе об основании Фив упоминается чудовищная змея, охраняющая источник, из которого новое поселение должно было черпать воду. В битве со змеей и ее убийстве все товарищи основателя Кадма погибли, что привело к появлению термина « Кадмейская победа » (то есть победа, влекущая за собой собственную гибель). [149]

Три медицинских символа со змеями, которые используются до сих пор, — это Чаша Гигиеи , символизирующая аптеку, а также Кадуцей и Жезл Асклепия , которые являются символами, обозначающими медицину в целом. [66]
Одна из этимологий, предложенных для общего женского имени Линда, заключается в том, что оно может происходить от древнегерманского Lindi или Linda , что означает змея.
Индию часто называют страной змей, и она богата традициями в отношении змей. [150] Змеям и сегодня поклоняются как богам, и многие женщины выливают молоко в змеиные ямы (несмотря на отвращение змей к молоку). [150] Кобру можно увидеть на шее Шивы , а Вишну часто изображают спящим на семиголовой змее или в кольцах змеи. [151] В Индии также есть несколько храмов исключительно для кобр, которых иногда называют Наградж (Король змей), и считается, что змеи являются символами плодородия. Каждый год проводится индуистский фестиваль Наг Панчами , в этот день почитают змей и молятся им. См. также Нага . [152]
Змея – одно из 12 небесных животных китайского зодиака в китайском календаре . [153]
Многие древние перуанские культуры поклонялись природе. [154] В своем искусстве они подчеркивали животных и часто изображали змей. [155]
Религия

Змеи используются в индуизме как часть ритуального поклонения. [157] На ежегодном фестивале Наг Панчами участники поклоняются либо живым кобрам, либо изображениям Нагов . Господь Шива на большинстве изображений изображен со змеей, обвивающей его шею. [158] Пураническая литература включает в себя различные истории, связанные со змеями, например, говорят, что Шеша держит на своих капюшонах все планеты Вселенной и постоянно воспевает славу Вишну изо всех своих уст. Другими известными змеями в индуизме являются Васуки , Такшака , Каркотака и Пингала . Термин Нага используется для обозначения существ, принимающих форму больших змей в индуизме и буддизме . [159]
Змеи широко почитались во многих культурах, например, в Древней Греции , где змей считали целителем. [160] Асклепий носил вокруг своей палочки змеиную рану - символ, который сегодня можно увидеть на многих машинах скорой помощи. [161] В иудаизме змея медная также является символом исцеления, спасения жизни от неминуемой смерти. [162]
С религиозной точки зрения змея и ягуар были, возможно, самыми важными животными в древней Мезоамерике . [163] «В состояниях экстаза лорды танцуют змеиный танец; огромные нисходящие змеи украшают и поддерживают здания от Чичен-Ицы до Теночтитлана , а на языке науатль, слово коатль означающее змею или близнеца, является частью первичных божеств, таких как Мишкоатль , Кетцалькоатль и Коатликуэ ». [164] В календарях майя и ацтеков пятый день недели назывался Днем Змеи.
В некоторых частях христианства искупительная работа Иисуса Христа сравнивается со спасением жизни через созерцание Нехуштана (медного змея). [165] Укротители змей используют змей как неотъемлемую часть церковного богослужения, чтобы продемонстрировать свою веру в божественную защиту. Однако чаще всего в христианстве змей изображался как представитель зла и коварных заговоров, как это видно из описания в книге Бытия змея, искушающего Еву в Эдемском саду . [166] Предполагается, что Святой Патрик изгнал всех змей из Ирландии во время обращения страны в христианство в V веке, что объясняет отсутствие там змей. [167]
В христианстве и иудаизме змея появляется в первой книге Библии, когда змей появляется перед Адамом и Евой и искушает их запретным плодом с Древа познания . [166] Змея возвращается в Книге Исход, когда Моисей превращает свой посох в змею в знак Божьей силы, а позже, когда он делает Нехуштан, бронзовую змею на шесте, которая, глядя на нее, исцеляла людей от укусов змей, которые преследовал их в пустыне. Змей в последний раз появляется, символизируя сатану, в Книге Откровения : «И схватил дракона, старого змея, который есть дьявол и сатана, и связал его на тысячу лет». [168]
В неоязычестве и Викке змея рассматривается как символ мудрости и знаний. [169] Кроме того, змеи иногда ассоциируются с Гекатой , греческой богиней колдовства . [170]
Лекарство
Несколько соединений змеиных ядов исследуются в качестве потенциальных средств лечения или профилактики боли, рака, артрита, инсульта, болезней сердца, гемофилии и гипертонии, а также для контроля кровотечения (например, во время операции). [171] [172] [173]
См. также
- Безногие позвоночные
- Список семей Змеиных
- Список змей
- офиология
- Змеебот
- Теория обнаружения змей
- Спинальная остеоартропатия (болезнь рептилий)
- Новая энциклопедия змей
- Змеи Европы и Змеи Европы Wikibooks:Змеи Европы , викикнига
Ссылки
- ↑ Перейти обратно: Перейти обратно: а б Сян А.Ю., полевой ди-джей, Вебстер Т.Х., Бельке А.Д., Дэвис М.Б., Расикот Р.А., Готье Дж.А. (май 2015 г.). «Происхождение змей: раскрытие экологии, поведения и истории эволюции ранних змей с использованием геномики, феноменомики и летописи окаменелостей» . Эволюционная биология BMC . 15 (1): 87. Бибкод : 2015BMCEE..15...87H . дои : 10.1186/s12862-015-0358-5 . ПМЦ 4438441 . ПМИД 25989795 .
- ^ Ридер Т.В., Таунсенд Т.М., Малкахи Д.Г., Нунан Б.П., Вуд PL, Sites JW, Wiens JJ (2015). «Комплексный анализ разрешает конфликты по поводу филогении чешуйчатых рептилий и выявляет неожиданные места размещения ископаемых таксонов» . ПЛОС ОДИН . 10 (3): e0118199. Бибкод : 2015PLoSO..1018199R . дои : 10.1371/journal.pone.0118199 . ПМЦ 4372529 . ПМИД 25803280 .
- ^ Винс Джей-Джей, Брэндли MC, Ридер Т.В. (январь 2006 г.). «Почему черта развивается несколько раз внутри клады? Повторяющаяся эволюция змееподобной формы тела у чешуйчатых рептилий» (PDF) . Эволюция; Международный журнал органической эволюции . 60 (1): 123–41. дои : 10.1554/05-328.1 . ПМИД 16568638 . S2CID 17688691 . Архивировано (PDF) из оригинала 2 февраля 2023 года . Проверено 21 января 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Баушо, Роланд, изд. (1994). Змеи: естественная история . Нью-Йорк: Sterling Publishing Co., Inc., с. 220 . ISBN 978-1-4027-3181-5 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к «Результаты поиска по высшему таксону: змея» . Reptile-database.org . Архивировано из оригинала 11 апреля 2021 года . Проверено 7 марта 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б Хеджес С.Б. (4 августа 2008 г.). «На нижнем пределе размера змей: два новых вида нитчатых змей (Squamata: Leptotyphlopidae: Leptotyphlops) с Малых Антильских островов» (PDF) . Зоотакса . 1841 : 1–30. дои : 10.11646/zootaxa.1841.1.1 . Архивировано (PDF) из оригинала 13 августа 2008 г. Проверено 4 августа 2008 г.
- ↑ Перейти обратно: Перейти обратно: а б Фредрикссон, генеральный директор (2005). «Хищничество сетчатого питона на солнечных медведей в Восточном Калимантане, индонезийское Борнео» . Зоологический бюллетень Raffles . 53 (1): 165–168. Архивировано из оригинала 9 июля 2014 года.
- ↑ Перейти обратно: Перейти обратно: а б Руководитель Дж.Дж., Блох Дж.И., Гастингс А.К., Бурк Дж.Р., Кадена Э.А., Эррера Ф.А. и др. (февраль 2009 г.). «Гигантская змея-боид из неотропов палеоцена обнаруживает более высокие температуры за экваториальными температурами». Природа . 457 (7230): 715–7. Бибкод : 2009Natur.457..715H . дои : 10.1038/nature07671 . ПМИД 19194448 . S2CID 4381423 .
- ^ Перкинс С. (27 января 2015 г.). «Обнаружены окаменелости древнейших известных змей» . news.sciencemag.org . Архивировано из оригинала 30 января 2015 года . Проверено 29 января 2015 г.
- ^ Колдуэлл М.В., Нидам Р.Л., Палчи А., Апестегия С. (январь 2015 г.). «Самые старые известные змеи из средней юры-нижнего мела дают представление об эволюции змей» . Природные коммуникации . 6 (5996): 5996. Бибкод : 2015NatCo...6.5996C . дои : 10.1038/ncomms6996 . hdl : 11336/37995 . ПМИД 25625704 .
- ^ Прато-IE: * (s)nēg-o- , Значение: змея, древнеиндийский: nagá- m. 'змея', германское: * snēk-a- м., * snak-an- м., * snak-ō f.; * snak-a- vb., русс. значение: жаба ( змея ), Ссылки: WP ( Vergleichendes Wörterbuch der indogermanischen Sprachen ) II 697 f.
- ^ «змея (сущ.)» . etymonline.com . Архивировано из оригинала 19 июля 2010 года . Проверено 22 сентября 2009 г.
- ^ «Определение змеи» . Интернет-словарь Мерриам-Вебстера . Архивировано из оригинала 17 октября 2007 года . Проверено 12 октября 2006 г.
- ^ Гамкрелидзе Томас В.; Иванов, Вячеслав В. (2010). Индоевропейцы и индоевропейцы: реконструкция и исторический анализ протоязыка и протокультуры . Вальтер де Грюйтер. п. 193. ИСБН 9783110815030 .
- ↑ Перейти обратно: Перейти обратно: а б «Змеи» . Интегрированная таксономическая информационная система . Проверено 4 апреля 2017 г.
- ^ Паф Ф.Х. (2002) [1992]. Герпетология: Третье издание . Пирсон Прентис Холл . ISBN 978-0-13-100849-6 .
- ^ Датта, Дебаджит; Баджпай, Сунил (18 апреля 2024 г.). «Самая крупная известная змея-мадцойид из теплого эоценового периода Индии предполагает межконтинентальное распространение в Гондване» . Научные отчеты . 14 (1): 8054. Бибкод : 2024NatSR..14.8054D . дои : 10.1038/s41598-024-58377-0 . ISSN 2045-2322 . ПМИД 38637509 .
- ^ Ли М.С., Хьюгалл А.Ф., Лоусон Р., Скэнлон Дж.Д. (2007). «Филогения змей (Serpentes): объединение морфологических и молекулярных данных по вероятности, байесовский анализ и анализ экономии». Систематика и биоразнообразие . 5 (4): 371–389. Бибкод : 2007SyBio...5..371L . дои : 10.1017/S1477200007002290 . hdl : 2440/44258 . S2CID 85912034 .
- ↑ Перейти обратно: Перейти обратно: а б МакДиармид Р.В., Кэмпбелл Дж.А., Туре Т. 1999. Мировые виды змей: таксономический и географический справочник, том. 1. Лига герпетологов. 511 стр. ISBN 1-893777-00-6 (серия). ISBN 1-893777-01-4 (объем).
- ^ Спалс, С.; Хауэлл, К.; Древес, Р.; Эш, Дж. (2004). Полевой справочник по рептилиям Восточной Африки . Лондон: ISBN A&C Black Publishers Ltd. 0-7136-6817-2 .
- ^ Elapidae в базе данных рептилий Reptarium.cz . По состоянию на 3 декабря 2008 г.
- ^ Буш, Брайан; Мэриан, Брэд; Браун-Купер, Роберт; Робинсон, Дэвид (2007). Рептилии и лягушки в кустах: Юго-Западная Австралия . Издательство Университета Западной Австралии . стр. 243, 244. ISBN. 978-1-920694-74-6 .
- ↑ Перейти обратно: Перейти обратно: а б с Название: Паскаль О.; Сингхал, Сонал; Грандлер, Майкл С.; Коста, Габриэль К.; Пайрон, Р. Александр; Колстон, Тимоти Дж.; Грандлер, Мэгги Р.; Пратес, Иван; Степанова, Наташа; Джонс, Марк Э.Х.; Кавальканти, Лукас БК; Колли, Гуарино Р.; Ди-Пои, Николя; Доннеллан, Стивен С.; Мориц, Крейг; Мескита, Дэниел О.; Пианка, Эрик Р.; Смит, Стивен А.; Витт, Лори Дж.; Рабоски, Дэниел Л. (23 февраля 2024 г.). «Макроэволюционная особенность змей» . Наука . 383 (6685): 918–923. Бибкод : 2024Sci...383..918T . дои : 10.1126/science.adh2449 . ПМИД 38386744 . Проверено 22 марта 2024 г.
- ^ Дюран, Дж. Ф. (2004). Происхождение змей . Геонауки Африки. Том. Абстрактный. Йоханнесбург, Южная Африка: Университет Витватерсранда . п. 187.
- ^ Видаль, Н.; Ярость, Ж.-К.; Кулу, А.; Хеджес, С.Б. (2009). «Змеи (Serpentes)». В Хеджесе, Южная Каролина; Кумар, С. (ред.). Древо времени жизни . Издательство Оксфордского университета . стр. 390–397.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п Мертенс, Дж. М. (1987). Живые змеи мира в цвете . Нью-Йорк: Стерлинг Паблишерс. ISBN 0-8069-6460-Х .
- ↑ Перейти обратно: Перейти обратно: а б с д и Санчес А. «Диапсиды III: Змеи» . Веб-сайт отца Санчеса, посвященный естественной истории Вест-Индии . Архивировано из оригинала 27 ноября 2007 года . Проверено 26 ноября 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б «Новая ископаемая змея с ногами» . База данных ЮНЕП ВЦМК . Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки . Архивировано из оригинала 25 декабря 2007 года . Проверено 29 ноября 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б Холман Дж. А. (2000). Ископаемые змеи Северной Америки (Первое изд.). Блумингтон, Индиана: Издательство Университета Индианы . стр. 284–323. ISBN 978-0253337214 .
- ^ Йи, Хунъюй; Норелл, Марк А. (2015). «Роющее происхождение современных змей» . Достижения науки . 1 (10): e1500743. Бибкод : 2015SciA....1E0743Y . дои : 10.1126/sciadv.1500743 . ПМЦ 4681343 . ПМИД 26702436 . S2CID 8912706 .
- ↑ Перейти обратно: Перейти обратно: а б Мак Дауэлл С. (1972). «Эволюция языка змей и ее влияние на происхождение змей». Эволюционная биология . Том. 6. стр. 191–273. дои : 10.1007/978-1-4684-9063-3_8 . ISBN 978-1-4684-9065-7 .
- ^ Апестегия С., Захер Х. (апрель 2006 г.). «Меловая наземная змея с крепкими задними конечностями и крестцом» . Природа . 440 (7087): 1037–40. Бибкод : 2006Natur.440.1037A . дои : 10.1038/nature04413 . ПМИД 16625194 . S2CID 4417196 . Архивировано из оригинала 18 декабря 2007 года.
- ^ «Необычайная окаменелость черепа раскрывает тайны эволюции змей» . 20 ноября 2019 года. Архивировано из оригинала 24 февраля 2024 года . Проверено 24 февраля 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б с Мертенс Р. (1961). «Лантанотус: важная ящерица в эволюции». Журнал музея Саравака . 10 : 320–322.
- ^ Джонатан В. (24 июля 2014 г.). «Предок четвероногих змей 'рыл норы' » . BBC Наука и окружающая среда . Архивировано из оригинала 26 июля 2015 года . Проверено 24 июля 2015 г.
- ^ Ён Э (23 июля 2015 г.). «Ископаемая змея с четырьмя ногами» . Нэшнл Географик . Архивировано из оригинала 23 июля 2015 года . Проверено 24 июля 2015 г.
- ^ Мартилл Д.М., Тишлингер Х., Лонгрич Н.Р. (июль 2015 г.). «ЭВОЛЮЦИЯ. Четвероногие змеи из раннего мела Гондваны» . Наука . 349 (6246): 416–9. Бибкод : 2015Sci...349..416M . дои : 10.1126/science.aaa9208 . ПМИД 26206932 . S2CID 25822461 . Архивировано из оригинала 2 августа 2021 года . Проверено 20 марта 2020 г.
- ^ «Знаменитое открытие ископаемого четвероногого змея, оказывается, имеет неожиданный поворот в истории» . www.msn.com . Архивировано из оригинала 18 ноября 2021 года . Проверено 18 ноября 2021 г.
- ^ Видал Н., Хеджес С.Б. (май 2004 г.). «Молекулярные доказательства наземного происхождения змей» . Слушания. Биологические науки . 271 (Приложение 4): С226-9. дои : 10.1098/rsbl.2003.0151 . ПМК 1810015 . ПМИД 15252991 .
- ^ Колдуэлл М.В., Нидам Р.Л., Палчи А., Апестегия С. (январь 2015 г.). «Самые старые известные змеи из средней юры-нижнего мела дают представление об эволюции змей» . Природные коммуникации . 6 (1): 5996. Бибкод : 2015NatCo...6.5996C . дои : 10.1038/ncomms6996 . hdl : 11336/37995 . ПМИД 25625704 .
- ^ Бергманн, Филип Дж.; Моринага, Ген (март 2019 г.). «Конвергентная эволюция змееподобных форм различными путями эволюции у чешуйчатых рептилий». Эволюция . 73 (3): 481–496. дои : 10.1111/evo.13651 . ПМИД 30460998 .
- ^ «Что безногая мышь говорит нам об эволюции змей» . Атлантика . Архивировано из оригинала 24 октября 2016 года . Проверено 25 октября 2016 г.
- ^ «Раньше у змей были ноги и руки… пока не произошли эти мутации» . Живая наука . Архивировано из оригинала 22 октября 2016 года . Проверено 22 октября 2016 г. .
- ^ Лил Ф., Кон М.Дж. (ноябрь 2016 г.). «Потеря и повторное появление ног у змей в результате модульной эволюции Sonic hedgehog и усилителей HOXD» . Современная биология . 26 (21): 2966–2973. Бибкод : 2016CBio...26.2966L . дои : 10.1016/j.cub.2016.09.020 . ПМИД 27773569 .
- ^ Квон Э.З., Камнева О.К., Мело У.С., Бароцци И., Остервальдер М., Маннион Б.Дж. и др. (октябрь 2016 г.). «Прогрессирующая потеря функции усилителя конечностей во время эволюции змеи» . Клетка . 167 (3): 633–642.e11. дои : 10.1016/j.cell.2016.09.028 . ПМЦ 5484524 . PMID 27768887 .
- ^ «База данных рептилий» . www.reptile-database.org . Архивировано из оригинала 2 ноября 2015 года . Проверено 6 марта 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б Конант Р. , Коллинз Дж. (1991). Полевое руководство по рептилиям и земноводным Восточной и Центральной Северной Америки . Бостон: Компания Houghton Mifflin . ISBN 978-0-395-58389-0 .
- ^ Информационный центр естествознания; Оклендский военный мемориальный музей. «Вопросы естествознания» . Оклендский военный мемориальный музей | Тамаки Паенга Хира . Окленд , Новая Зеландия: Оклендский военный мемориальный музей . Вопрос: Есть ли змеи в Новой Зеландии? Архивировано из оригинала 12 июля 2012 года . Проверено 26 апреля 2012 г.
- ^ Ривас Дж.А. (2000). История жизни зеленой анаконды ( Eunectes murinus ) с акцентом на ее репродуктивную биологию (PDF) ( докторская диссертация). Университет Теннесси . Архивировано из оригинала (PDF) 3 марта 2016 года . Проверено 12 декабря 2014 г.
- ^ Бобак С.М., Гайер С. (февраль 2003 г.). «Эмпирические доказательства оптимального размера тела змей». Эволюция; Международный журнал органической эволюции . 57 (2): 345–51. doi : 10.1554/0014-3820(2003)057[0345:EEFAOB]2.0.CO;2 . ПМИД 12683530 . S2CID 198156987 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Коггер и Даут 1992 , с. 180.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час Кэмпбелл и Шоу, 1974 .
- ↑ Перейти обратно: Перейти обратно: а б Кэмпбелл и Шоу 1974 , с. 19.
- ↑ Перейти обратно: Перейти обратно: а б Зденек, Кристина Н.; Стейплс, Тимоти; Привет, Крис; Бурк, Лахлан Н.; Кандуссо, Дамиан (2023). «Звуковой сад: как змеи реагируют на воздушные и наземные звуки» . ПЛОС ОДИН . 18 (2): e0281285. Бибкод : 2023PLoSO..1881285Z . дои : 10.1371/journal.pone.0281285 . ПМЦ 9928108 . ПМИД 36787306 .
- ^ «Чувства рептилий: понимание их мира» . Petplace.com . 18 мая 2015 года. Архивировано из оригинала 19 февраля 2015 года . Проверено 9 января 2016 г.
- ^ «Морские змеи вернули цветовое зрение посредством редкой генетической эволюции» . 12 июля 2023 года. Архивировано из оригинала 28 сентября 2023 года . Проверено 28 сентября 2023 г.
- ^ «Некоторые морские змеи вновь обрели способность видеть больше цветов» . Архивировано из оригинала 28 сентября 2023 года . Проверено 28 сентября 2023 г.
- ^ «Змеиные глаза: новый взгляд на визуальную адаптацию» . ScienceDaily . 16 августа 2016 г. Архивировано из оригинала 18 декабря 2019 г. . Проверено 18 декабря 2019 г.
- ^ Симойнс, Бруно Ф.; и др. (октябрь 2016 г.). «Визуальные пигменты, глазные фильтры и эволюция змеиного зрения» . Молекулярная биология и эволюция . 33 (10). Издательство Оксфордского университета : 2483–2495. дои : 10.1093/molbev/msw148 . hdl : 1983/2008257f-b043-4206-a44b-39ebbdd1bea8 . ПМИД 27535583 . Архивировано из оригинала 23 марта 2021 года . Проверено 1 марта 2021 г.
- ^ Смит, Малкольм А. Фауна Британской Индии, включая Цейлон и Бирму . Том I, Лориката и Тестудины. п. 30.
- ↑ Перейти обратно: Перейти обратно: а б с д «Змеи склизкие?» . szgdocent.org . Архивировано из оригинала 5 августа 2006 года.
- ^ Аллен В.Л., Бэддели Р., Скотт-Сэмюэл Н.Э., Катхилл И.С. (2013). «Эволюция и функция разнообразия узоров у змей» . Поведенческая экология . 24 (5): 1237–1250. дои : 10.1093/beheco/art058 . ISSN 1465-7279 .
- ↑ Перейти обратно: Перейти обратно: а б «Общая информация о змеях» . sdgfp.info . Архивировано из оригинала 25 ноября 2007 года.
- ^ «ZooPax: Вопрос масштаба: Часть III» . Whozoo.org . Архивировано из оригинала 16 января 2016 года . Проверено 9 января 2016 г.
- ^ Баувенс, Дирк; Ван Дамм, Рауль; Верхейен, Рудольф Ф. (1989). «Синхронизация весенней линьки с началом брачного поведения у самцов ящериц Lacerta vivipara» . Журнал герпетологии . 23 (1): 89–91. дои : 10.2307/1564326 . ISSN 0022-1511 . JSTOR 1564326 . Проверено 29 апреля 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б Уилкокс Р.А., Уизем Э.М. (апрель 2003 г.). «Символ современной медицины: почему одна змея больше двух». Анналы внутренней медицины . 138 (8): 673–7. CiteSeerX 10.1.1.731.8485 . дои : 10.7326/0003-4819-138-8-200304150-00016 . ПМИД 12693891 . S2CID 19125435 .
- ↑ Перейти обратно: Перейти обратно: а б с Розенфельд (1989), с. 11.
- ^ Кэмпбелл и Шоу 1974 , с. 11.
- ^ Фридель П., Янг Б.А., ван Хеммен Дж.Л. (февраль 2008 г.). «Слуховая локализация наземных колебаний у змей». Письма о физических отзывах . 100 (4): 048701. Бибкод : 2008PhRvL.100d8701F . дои : 10.1103/physrevlett.100.048701 . ПМИД 18352341 .
- ^ Зыга Л (13 февраля 2008 г.). «Пустынная змея слышит челюстью шаги мыши» . Физика.орг . Архивировано из оригинала 10 октября 2011 года.
- ^ Хартлайн PH (апрель 1971 г.). «Физиологические основы обнаружения звука и вибрации у змей» (PDF) . Журнал экспериментальной биологии . 54 (2): 349–71. дои : 10.1242/jeb.54.2.349 . ПМИД 5553415 . Архивировано (PDF) из оригинала 17 декабря 2008 г.
- ^ Коггер, Х. 1993. Фауна Австралии. Том. 2А Амфибии и Рептилии. Австралийские исследования биологических ресурсов, Канберра.
- ^ Арнольд Э.Н. (1984). «Эволюционные аспекты сбрасывания хвоста у ящериц и их родственников». Журнал естественной истории . 18 (1): 127–169. Бибкод : 1984JNatH..18..127A . дои : 10.1080/00222938400770131 .
- ^ Ананьева Н.Б., Орлов Н.Л. (1994). «Хвостовая аутотомия у колубридной змеи Xenochrophis piscator из Вьетнама». Российский герпетологический журнал . 1 (2).
- ^ Гаэте М., Такер А.С. (2013). «Организованное появление нескольких поколений зубов у змей нарушается за счет активации передачи сигналов Wnt/бета-катенин» . ПЛОС ОДИН . 8 (9): e74484. Бибкод : 2013PLoSO...874484G . дои : 10.1371/journal.pone.0074484 . ПМК 3760860 . ПМИД 24019968 .
- ^ Дженсен Б., Мурман А.Ф., Ван Т (май 2014 г.). «Строение и функции сердца ящериц и змей». Биологические обзоры Кембриджского философского общества . 89 (2): 302–36. дои : 10.1111/brv.12056 . ПМИД 23998743 . S2CID 20035062 .
- ^ Бурггрен WW (1 февраля 1987 г.). «Форма и функция в циркуляции рептилий» . Интегративная и сравнительная биология . 27 (1): 5–19. дои : 10.1093/icb/27.1.5 . ISSN 1540-7063 .
- ^ Матур П. (1944). «Анатомия сердца рептилий. Часть I. Варанский варан (Линн.)» . Учеб. Индийский акад. Наука . Секта. Б 20: 1–29. Архивировано из оригинала 10 мая 2019 года . Проверено 10 мая 2019 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Мадер Д. (июнь 1995 г.). «Анатомия рептилий». Рептилии . 3 (2): 84–93.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Оливейра, Ана Л.; Вьегас, Матильда Ф.; да Силва, Сауло Л.; Соарес, Андреймар М.; Рамос, Мария Дж.; Фернандес, Педро А. (июль 2022 г.). «Химия змеиного яда и его лечебный потенциал» . Обзоры природы Химия . 6 (7): 451–469. дои : 10.1038/s41570-022-00393-7 . ISSN 2397-3358 . ПМЦ 9185726 . ПМИД 35702592 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Фрайберг и Уоллс 1984 , с. 125.
- ↑ Перейти обратно: Перейти обратно: а б Фрайберг и Уоллс 1984 , с. 123.
- ↑ Перейти обратно: Перейти обратно: а б с д Фрайберг и Уоллс 1984 , с. 126.
- ^ Фрай Б.Г., Видал Н., Норман Дж.А., Вонк Ф.Дж., Шейб Х., Рамджан С.Ф. и др. (февраль 2006 г.). «Ранняя эволюция ядовитой системы ящериц и змей». Природа . 439 (7076): 584–8. Бибкод : 2006Natur.439..584F . дои : 10.1038/nature04328 . ПМИД 16292255 . S2CID 4386245 .
- ↑ Перейти обратно: Перейти обратно: а б с д Капула (1989), с. 117.
- ^ Олдридж Р.Д., Север DM (19 апреля 2016 г.). Репродуктивная биология и филогения змей . ЦРК Пресс . ISBN 978-1-4398-5833-2 – через Google Книги .
- ^ Фауэлл, Меган Дж.; Сандерс, Кейт Л.; Бреннан, Патрисия Л.Р.; Кроу-Ридделл, Дженна М. (21 декабря 2022 г.). «Первые свидетельства наличия гемиклиторов у змей» . Труды Королевского общества Б. 289 (1989). дои : 10.1098/rspb.2022.1702 . ПМЦ 9748774 . ПМИД 36515117 .
- ↑ Перейти обратно: Перейти обратно: а б Коггер и Даут 1992 , с. 186.
- ↑ Перейти обратно: Перейти обратно: а б Капула (1989), с. 118.
- ↑ Перейти обратно: Перейти обратно: а б с Коггер и Даут 1992 , с. 182.
- ^ Шайн Р., Лангкилде Т., Мейсон Р.Т. (2004). «Тактика ухаживания подвязочных змей: как морфология и поведение самца влияют на успех его спаривания?». Поведение животных . 67 (3): 477–83. дои : 10.1016/j.anbehav.2003.05.007 . S2CID 4830666 .
- ^ Блуэн-Демерс Дж., Гиббс Х.Л., Уэзерхед П.Дж. (2005). «Генетические доказательства полового отбора у черных крысиных змей Elaphe obsoleta ». Поведение животных . 69 (1): 225–34. дои : 10.1016/j.anbehav.2004.03.012 . S2CID 3907523 .
- ↑ Перейти обратно: Перейти обратно: а б Бут В., Смит К.Ф., Эскридж П.Х., Хосс С.К., Мендельсон-младший, Шуэтт Г.В. (декабрь 2012 г.). «Факультативный партеногенез обнаружен у диких позвоночных» . Письма по биологии . 8 (6): 983–5. дои : 10.1098/rsbl.2012.0666 . ПМЦ 3497136 . ПМИД 22977071 .
- ^ Бут В., Миллион Л., Рейнольдс Р.Г., Бургхардт Г.М., Варго Э.Л., Шал С. и др. (2011). «Последовательные непорочные зачатия в новом мире боид-змея, колумбийский радужный удав, Epicrates maurus» . Журнал наследственности . 102 (6): 759–63. doi : 10.1093/jhered/esr080 . ПМИД 21868391 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Уолтеринг, Йост М. (2012). «От ящерицы к змее: за эволюцией экстремального строения тела» . Современная геномика . 13 (4): 289–299. дои : 10.2174/138920212800793302 . ПМК 3394116 . ПМИД 23204918 .
- ↑ Перейти обратно: Перейти обратно: а б Зер, Дэвид Р. (20 июля 1962 г.). «Этапы нормального развития обыкновенной подвязочной змеи Thamnophis sirtalis sirtalis». Копейя . 1962 (2): 322–329. дои : 10.2307/1440898 . JSTOR 1440898 .
- ^ Гомес, Селин; Озбудак, Эртугрул М.; Вундерлих, Джошуа; Бауманн, Диана; Льюис, Джулиан; Пурке, Оливье (17 июля 2008 г.). «Контроль количества сегментов у эмбрионов позвоночных» . Природа . 454 (7202): 335–339. Бибкод : 2008Natur.454..335G . дои : 10.1038/nature07020 . ISSN 0028-0836 . ПМИД 18563087 . S2CID 4373389 . Архивировано из оригинала 26 марта 2023 года . Проверено 30 апреля 2022 г.
- ^ Буфнер, Джулия К.; Бухтова, Марсела; Фу, Кэтрин; Диверт, Вирджиния; Халлгримссон, Бенедикт; Ричман, Джой М. (июнь 2007 г.). «Эмбриональное развитие Python sebae – I: критерии стадирования и макроскопический морфогенез скелета головы и конечностей». Зоология . 110 (3): 212–230. Бибкод : 2007Zool..110..212B . дои : 10.1016/j.zool.2007.01.005 . ПМИД 17499493 .
- ↑ Перейти обратно: Перейти обратно: а б с Леал, Франциска; Кон, Мартин Дж. (январь 2018 г.). «Эволюционное, генетическое и геномное понимание эволюционной потери конечностей у змей». Бытие . 56 (1): e23077. дои : 10.1002/dvg.23077 . ПМИД 29095557 . S2CID 4510082 .
- ^ Квон Евгений З.; Камнева Ольга К.; Мело, Уира С.; Бароцци, Ирос; Остервальдер, Марко; Мэннион, Брэндон Дж.; Тиссьер, Вирджиния; Пикл, Кэтрин С.; Плайзер-Фрик, Ингрид; Ли, Элизабет А.; Като, Момоэ (октябрь 2016 г.). «Прогрессирующая потеря функции усилителя конечностей во время эволюции змеи» . Клетка . 167 (3): 633–642.e11. дои : 10.1016/j.cell.2016.09.028 . ПМЦ 5484524 . PMID 27768887 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Белер и Кинг 1979 , с. 581.
- ^ Хосо М., Асами Т., Хори М. (апрель 2007 г.). «Змеи-правши: конвергентная эволюция асимметрии для функциональной специализации» . Письма по биологии . 3 (2): 169–72. дои : 10.1098/rsbl.2006.0600 . ПМЦ 2375934 . ПМИД 17307721 .
- ^ Пайрон Р.А., Бербринк Ф.Т., Винс Дж.Дж. (апрель 2013 г.). «Филогения и пересмотренная классификация Squamata, включая 4161 вид ящериц и змей» . Эволюционная биология BMC . 13 (1): 93. Бибкод : 2013BMCEE..13...93P . дои : 10.1186/1471-2148-13-93 . ПМК 3682911 . ПМИД 23627680 .
- ^ Freiberg & Walls 1984 , стр. 125–127.
- ^ Таттерсолл Г.Дж., Милсом В.К., Эйб А.С., Брито С.П., Андраде Д.В. (февраль 2004 г.). «Термогенез пищеварения у гремучих змей» . Журнал экспериментальной биологии . 207 (Часть 4): 579–85. дои : 10.1242/jeb.00790 . ПМИД 14718501 .
- ^ Янг, Брюс А.; Кардонг, Кеннет В. (май 2010 г.). «Функциональная морфология капюшона кобры». Журнал экспериментальной биологии . 213 (9): 1521–1528. дои : 10.1242/jeb.034447 . ПМИД 20400637 .
- ^ Янг, Брюс А.; Данлэп, Карен; Кениг, Кристен; Певица Мередит (15 сентября 2004 г.). «Щёчная пряжка: функциональная морфология плевания ядом кобры». Журнал экспериментальной биологии . 207 (20): 3483–3494. дои : 10.1242/jeb.01170 . ПМИД 15339944 .
- ^ Насури, Алиреза; Шахбаззаде, Делавар; Цубота, Тосио; Янг, Брюс А. (зима 2016 г.). «Защитное поведение Naja oxiana с комментариями к визуальным проявлениям кобр» . Герпетологический вестник (138). Архивировано из оригинала 3 мая 2021 года . Проверено 3 мая 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Коггер и Даут 1992 , с. 175.
- ↑ Перейти обратно: Перейти обратно: а б Грей Дж (декабрь 1946 г.). «Механизм передвижения змей». Журнал экспериментальной биологии . 23 (2): 101–20. дои : 10.1242/jeb.23.2.101 . ПМИД 20281580 .
- ↑ Перейти обратно: Перейти обратно: а б Хекротте С (1967). «Взаимосвязь температуры тела, размера и скорости ползания обыкновенной подвязочной змеи Thamnophis s. sirtalis». Копейя . 23 (4): 759–763. дои : 10.2307/1441886 . JSTOR 1441886 .
- ↑ Перейти обратно: Перейти обратно: а б с Уолтон М., Джейн Б.К., Беннет А.Ф. (август 1990 г.). «Энергетическая стоимость передвижения без конечностей». Наука . 249 (4968): 524–7. Бибкод : 1990Sci...249..524W . дои : 10.1126/science.249.4968.524 . ПМИД 17735283 . S2CID 17065200 .
- ↑ Перейти обратно: Перейти обратно: а б Грей, Дж.; Лиссманн, HW (февраль 1950 г.). «Кинетика передвижения ужа». Журнал экспериментальной биологии . 26 (4): 354–367. дои : 10.1242/jeb.26.4.354 .
- ^ Грей Дж (1953). «Волнообразное движение». Ежеквартальный журнал микроскопической науки . 94 : 551–578.
- ^ Джейн, Британская Колумбия (август 1988 г.). «Мышечные механизмы передвижения змей: электромиографическое исследование боковых волнистости флоридской полосатой водяной змеи (Nerodia fasciata) и желтой крысиной змеи (Elaphe obsoleta)». Журнал морфологии . 197 (2): 159–81. дои : 10.1002/jmor.1051970204 . ПМИД 3184194 . S2CID 25729192 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Коггер и Даут 1992 , с. 177.
- ↑ Перейти обратно: Перейти обратно: а б Джейн, Британская Колумбия (1986). «Кинематика передвижения наземных змей». Копейя . 1986 (4): 915–927. дои : 10.2307/1445288 . JSTOR 1445288 .
- ↑ Перейти обратно: Перейти обратно: а б с д Эстли ХК, Джейн БК (ноябрь 2007 г.). «Влияние диаметра и наклона насеста на кинематику, работоспособность и способы древесного передвижения кукурузных змей (Elaphe guttata)». Журнал экспериментальной биологии . 210 (Часть 21): 3862–72. дои : 10.1242/jeb.009050 . ПМИД 17951427 . S2CID 18129284 .
- ↑ Перейти обратно: Перейти обратно: а б Фрайберг и Уоллс 1984 , с. 135.
- ^ Соча Джей-Джей (август 2002 г.). «Скользящий полет на райском дереве змеи». Природа . 418 (6898): 603–4. Бибкод : 2002Natur.418..603S . дои : 10.1038/418603а . ПМИД 12167849 . S2CID 4424131 .
- ↑ Перейти обратно: Перейти обратно: а б Коггер и Даут 1992 , с. 176.
- ↑ Перейти обратно: Перейти обратно: а б «Змеиные укусы» . MedlinePlus.gov . Архивировано из оригинала 4 декабря 2010 года . Проверено 9 марта 2010 г. от Тинтиналли Дж.Э., Келен Г.Д., Стапчински Дж.С., ред. Неотложная медицина: Комплексное учебное пособие . 6-е изд. Нью-Йорк, штат Нью-Йорк: МакГроу Хилл; 2004 г. Дата обновления: 27 февраля 2008 г. Обновлено: Стивен С. Акоста, доктор медицинских наук, отделение неотложной медицины, Медицинский центр Портленда, штат Вирджиния, Портленд, Орегон. Обзор предоставлен VeriMed Healthcare Network. Также проверено Дэвидом Зивом, доктором медицинских наук, MHA, медицинским директором ADAM, Inc.
- ^ «Первая помощь при укусе змеи – укус змеи» . Health-care-clinic.org . Архивировано из оригинала 16 января 2016 года . Проверено 9 января 2016 г.
- ^ ВОЗ. «17 забытых тропических болезней» . ВОЗ . Всемирная организация здравоохранения . Архивировано из оригинала 22 февраля 2014 года . Проверено 24 октября 2014 г.
- ^ Синха К. (25 июля 2006 г.). «Земли заклинателей змей больше нет…» The Times of India . Архивировано из оригинала 11 августа 2011 года.
- ^ «Отравление змеиным укусом» . Всемирная организация здравоохранения . Архивировано из оригинала 18 апреля 2017 года . Проверено 27 октября 2017 г.
- ^ «Тропический лес Амазонки» . Education.nationalgeographic.org . Проверено 23 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б с «Змеиный укус» . www.who.int . Проверено 23 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б Уоррелл, Дэвид А. (январь 2010 г.). «Укус змеи». Ланцет . 375 (9708): 77–88. дои : 10.1016/S0140-6736(09)61754-2 . ПМИД 20109866 .
- ^ Уильямс, Дэвид Дж.; Фаиз, Мохд Абул; Абела-Риддер, Бернадетт; Эйнсворт, Стюарт; Бульфоне, Томмазо К.; Никерсон, Андреа Д.; Хабиб, Абдулразак Г.; Юнгханс, Томас; Фань, Хуэй Вэнь; Тернер, Майкл; Харрисон, Роберт А.; Уоррелл, Дэвид А. (21 февраля 2019 г.). «Стратегия глобально скоординированного ответа на приоритетную забытую тропическую болезнь: отравление змеиными укусами» . PLOS Забытые тропические болезни . 13 (2): e0007059. дои : 10.1371/journal.pntd.0007059 . ISSN 1935-2735 . ПМК 6383867 . ПМИД 30789906 .
- ↑ Перейти обратно: Перейти обратно: а б с Багла П. (23 апреля 2002 г.). «Индийские заклинатели змей исчезают, обвиняя экологические законы, телевидение» . Национальные географические новости . Архивировано из оригинала 18 декабря 2007 года . Проверено 26 ноября 2007 г.
- ^ Хардинг Л. (2 апреля 2002 г.). «Змеиные проделки теряют свое очарование» . Хранитель . Архивировано из оригинала 16 мая 2020 года . Проверено 16 апреля 2020 г.
- ^ Чандра С. (12 ноября 2013 г.). «Индийские заклинатели змей находятся на грани вымирания» . Индия сегодня . Архивировано из оригинала 3 мая 2020 года . Проверено 16 апреля 2020 г.
- ^ Бертон, Морис; Бертон, Роберт (2002). «Блеф заклинателя змей». Международная энциклопедия дикой природы . Том. 4 (3-е изд.). Маршалл Кавендиш. п. 482. ИСБН 9780761472704 . Архивировано из оригинала 18 августа 2016 года — через Google Книги .
- ^ Уитакер, Ромул; Капитан Ашок (2004). Змеи Индии: Полевой справочник . стр. 11–13.
- ↑ Перейти обратно: Перейти обратно: а б с «Вино с остротой» . www.bbc.com . Проверено 8 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б «Статьи с тегом Змеи | Смитсоновский журнал» . www.smithsonianmag.com . Проверено 8 марта 2024 г.
- ^ Эрнест С., Цуг Г.Р., Гриффин, доктор медицинских наук (1996). Змеи под вопросом: Книга ответов Смитсоновского института . Вашингтон, округ Колумбия: Смитсоновские книги. п. 203 . ISBN 978-1-56098-648-5 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я Блэк Джей, Грин А (1992). Боги, демоны и символы Древней Месопотамии: Иллюстрированный словарь . Остин, Техас: Издательство Техасского университета . стр. 166–168. ISBN 978-0714117058 .
- ^ Сара Прюитт (10 марта 2020 г.). «Действительно ли Клеопатра умерла от укуса змеи?» . History.com .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Хорнунг, Эрик (2001). Тайные знания Египта: их влияние на Запад . Итака, Нью-Йорк и Лондон, Англия: Издательство Корнельского университета . стр. 13, 44. ISBN. 978-0-8014-3847-9 . Архивировано из оригинала 19 февраля 2024 года . Проверено 20 октября 2020 г. - через Google Книги .
- ^ Финни, Эдвард младший (1971). «Битва Персея с горгонами». Труды и труды Американской филологической ассоциации . 102 : 445–463. дои : 10.2307/2935950 . JSTOR 2935950 .
- ^ Кинсли, Дэвид (1989). Зеркало богинь: видения божества с востока и запада . Олбани, Нью-Йорк: Издательство Нью-Йоркского государственного университета . п. 151. ИСБН 978-0-88706-836-2 . Архивировано из оригинала 25 апреля 2023 года . Проверено 20 октября 2020 г. - через Google Книги .
- ^ Диси, Сьюзен (2008). Афина . Нью-Йорк, Нью-Йорк и Лондон, Англия: Routledge . ISBN 978-0-415-30066-7 . Архивировано из оригинала 19 февраля 2024 года . Проверено 20 октября 2020 г. - через Google Книги .
- ^ Псевдо-Аполлодор, Библиотека 2.37, 38, 39.
- ^ Силиг, Бет Дж. (август 2002 г.). «Похищение Медузы в храме Афины: аспекты триангуляции у девушки». Международный журнал психоанализа . 83 (4): 895–911. doi : 10.1516/3NLL-UG13-TP2J-927M . ПМИД 12204171 . S2CID 28961886 .
- ↑ Перейти обратно: Перейти обратно: а б Уэст, Мартин Личфилд (2007). Индоевропейская поэзия и миф . Оксфорд, Англия: Издательство Оксфордского университета . п. 258 . ISBN 978-0-19-928075-9 .
- ↑ Перейти обратно: Перейти обратно: а б Огден, Дэниел (2013). Дракон : миф о драконе и культ змеи в древнегреческом и римском мирах . Оксфорд, Англия: Издательство Оксфордского университета . стр. 28–29. ISBN 978-0-19-955732-5 . Архивировано из оригинала 19 февраля 2024 года . Проверено 20 октября 2020 г. - через Google Книги .
- ^ Микаэла Джанан (2009). Отражения в змеином глазу Фивы в «Метаморфозах» Овидия . ОУП Оксфорд. ISBN 9780191572258 .
- ↑ Перейти обратно: Перейти обратно: а б Дин 1833 , с. 61.
- ^ Дин 1833 , стр. 62–64.
- ^ Джеймс Чемберс, изд. (2015). Праздничные символы и обычаи, 5-е изд . Издательство информационной базы. п. 782. ИСБН 9780780813656 .
- ^ «Китайский календарь» . timeanddate.com . Архивировано из оригинала 15 августа 2017 года . Проверено 1 июня 2017 г.
- ^ Бенсон, Элизабет (1972). Мочика: культура Перу . Лондон: Темза и Гудзон . ISBN 978-0-500-72001-1 .
- ^ Беррин К., Музей Ларко (1997). Дух древнего Перу: сокровища из археологического музея Рафаэля Ларко Эрреры Нью-Йорк: Темза и Гудзон . ISBN 978-0-500-01802-6 .
- ^ Шарер RJ , Traxler LP (2006). Древние майя (6-е (полностью переработанное) изд.). Стэнфорд, Калифорния: Издательство Стэнфордского университета . п. 619 . ISBN 978-0-8047-4817-9 . OCLC 57577446 .
- ^ Керкар, Раджендра П. (4 августа 2011 г.). «Сегодня индусы объединяются, чтобы поклоняться змеиному богу» . Таймс оф Индия . Архивировано из оригинала 7 мая 2021 года . Проверено 3 марта 2021 г.
- ^ Айер, Гаятри (22 августа 2019 г.). «Каково значение змеи на шее Господа Шивы?» . TimesNowNews.com . Архивировано из оригинала 15 апреля 2021 года . Проверено 2 марта 2021 г.
- ^ «нага | Индуистская мифология» . Британника . Архивировано из оригинала 4 сентября 2022 года . Проверено 1 июля 2022 г.
- ^ Цукалас, Грегори; Андрутсос, Джордж (2019). «Асклепий и змея как токсикологические символы в Древней Греции и Риме». Токсикология в древности . стр. 257–265. дои : 10.1016/B978-0-12-815339-0.00017-2 . ISBN 978-0-12-815339-0 .
- ^ «Происхождение символа скорой медицинской помощи» . Mediband.com . 26 февраля 2019 года. Архивировано из оригинала 26 марта 2023 года . Проверено 26 февраля 2021 г.
- ^ Числа 21:6–21:9.
- ^ Викери, Кирби (1 августа 2019 г.). «Мезоамериканский змей» . Мансанильо Сан . Архивировано из оригинала 20 января 2021 года . Проверено 3 марта 2021 г.
- ^ Миллер, Мэри (1993). Боги и символы Древней Мексики и майя . Лондон: Темза и Гудзон . ISBN 978-0-500-27928-1 .
- ^ Иоанна 3:14
- ↑ Перейти обратно: Перейти обратно: а б Бытие 3:1
- ^ Дрю, апрель (24 февраля 2019 г.). «Действительно ли Святой Патрик изгнал всех змей из Ирландии?» . IrishCentral.com . Архивировано из оригинала 10 марта 2021 года . Проверено 2 марта 2021 г.
- ^ Откровение 20:2.
- ^ «ДИСКУССИЯ О КОЛДОВСТВЕ, НЕОЯЗЫЧЕСТВЕ ВИККИ И АФРИКАНСКИХ ТРАДИЦИЯХ» . People.ucalgary.ca . Архивировано из оригинала 9 июня 2022 года . Проверено 1 июля 2022 г.
- ^ Зейфферт, Оскар (1901). Словарь классических древностей: мифология, религия, литература и искусство (6-е изд.). Лебедь Зонненшайн и Ко . п. 271 . Проверено 2 января 2022 г.
- ^ Вьяс В.К., Брамбхатт К., Бхатт Х., Пармар У. (февраль 2013 г.). «Терапевтический потенциал змеиного яда в терапии рака: современные перспективы» . Азиатско-Тихоокеанский журнал тропической биомедицины . 3 (2): 156–62. дои : 10.1016/S2221-1691(13)60042-8 . ПМЦ 3627178 . ПМИД 23593597 .
- ^ Голландия JS (февраль 2013 г.). «Укус, который лечит» . Нэшнл Географик . Архивировано из оригинала 25 мая 2018 года.
- ^ Уилкокс С. (2016). Ядовитый . Научный американец . ISBN 978-0374283377 . Архивировано из оригинала 24 мая 2018 года . Проверено 24 мая 2018 г.
Библиография
- Белер Дж.Л., Кинг ФВ (1979). Полевое руководство Общества Одюбона по рептилиям и амфибиям Северной Америки . Нью-Йорк: Альфред А. Кнопф . п. 581 . ISBN 978-0-394-50824-5 .
- Кэмпбелл, Шелдон; Шоу, Чарльз Э. (1974). Змеи американского Запада . Нью-Йорк: Альфред А. Кнопф . ISBN 978-0-394-48882-0 .
- Коггер Х. , Цвайфель Р. (1992). Рептилии и амфибии . Сидней: Уэлдон Оуэн. ISBN 978-0-8317-2786-4 .
- Дин, Джон (1833). Поклонение Змею . Уайтфиш, Монтана: Издательство Кессинджер. п. 412. ИСБН 978-1-56459-898-1 . Архивировано из оригинала 9 октября 2019 года . Проверено 2 декабря 2007 г.
- Фрайберг М., Уоллс Дж. (1984). Мир ядовитых животных . Нью-Джерси: Публикации TFH. ISBN 978-0-87666-567-1 .
Дальнейшее чтение
- Снегирь Т (2000). Полная мифология снегиря . Лондон: Канцлер Пресс. п. 679. ИСБН 978-0-7537-0381-6 . Архивировано из оригинала 9 февраля 2009 года.
- Капула М., Белер Дж.Л. (1989). Путеводитель Саймона и Шустера по рептилиям и амфибиям мира . Нью-Йорк: Саймон и Шустер . ISBN 978-0-671-69098-4 .
- Коборн Дж (1991). Атлас змей мира . Нью-Джерси: Публикации TFH. ISBN 978-0-86622-749-0 .
- Конант Р. , Коллинз Дж. (1991). Полевое руководство по рептилиям и земноводным Восточной и Центральной Северной Америки . Бостон: Компания Houghton Mifflin . ISBN 978-0-395-58389-0 .
- Дитмарс, Раймонд Л. (1906). Ядовитые змеи США: как их отличить . Нью-Йорк: ER Sanborn. п. 11.
- Дитмарс, Раймонд Л. (1931). Змеи мира . Нью-Йорк: Макмиллан. п. 11 . ISBN 978-0-02-531730-7 .
- Дитмарс Р.Л. (1933). Рептилии мира: крокодилы, ящерицы, змеи, черепахи и черепахи Восточного и Западного полушарий . Нью-Йорк: Макмиллан. п. 321.
- Дитмарс Р.Л. , Бриджес В. (1935). Праздник охотников на змей . Нью-Йорк: Д. Эпплтон и компания. п. 309.
- Дитмарс Р.Л. (1939). Полевая книга североамериканских змей . Гарден-Сити, Нью-Йорк: Doubleday, Doran & Co. 305.
- Гиббонс Дж.В., Гиббонс В. (1983). У них кровь стынет в жилах: приключения с рептилиями и амфибиями . Алабама: Издательство Университета Алабамы . п. 164 . ISBN 978-0-8173-0135-4 .
- Мэттисон С (2007). Новая энциклопедия змей . Нью-Джерси: Издательство Принстонского университета . п. 272. ИСБН 978-0-691-13295-2 .
- МакДиармид Р.В., Кэмпбелл Дж.А., Туре Т. (1999). Мировые виды змей: таксономический и географический справочник . Том. 1. Лига герпетологов. п. 511. ИСБН 978-1-893777-00-2 .
- Мертенс Дж (1987). Живые змеи мира в цвете . Нью-Йорк: Стерлинг. ISBN 978-0-8069-6461-4 .
- Нобрега Алвес Р.Р., Сильва Виейра В.Л., Сантана Г.Г. (2008). «Рептилии, используемые в традиционной народной медицине: последствия сохранения». Биоразнообразие и сохранение . 17 (8): 2037–2049. Бибкод : 2008BiCon..17.2037D . дои : 10.1007/s10531-007-9305-0 . S2CID 42500066 .
- О'Ши, Марк (2023). Змеи мира: Путеводитель для каждой семьи . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-24066-4 .
- Уитакер Р. (1996). Змеи вокруг нас, тамильский . Национальный книжный фонд. ISBN 978-81-237-1905-4 .
- Розенфельд А (1989). Экзотические домашние животные . Нью-Йорк: Саймон и Шустер . п. 293 . ISBN 978-0-671-47654-0 .
- Спалс С., Отделение B (1995). Опасные змеи Африки . Остров Санибел, Флорида: Издательство Ральфа Кертиса. п. 192. ИСБН 978-0-88359-029-4 .
Внешние ссылки
- «Библиография к «Змеям» » . Библиотека наследия биоразнообразия.
- «Змеи» . Интегрированная таксономическая информационная система .
- «Американские змеи» . еПрирода. Архивировано из оригинала 15 марта 2008 года.
- «Змеи Индийского субконтинента» . Фотогалерея Naturemagics Кералы. Архивировано из оригинала 21 марта 2009 года . Проверено 8 января 2007 г.
- «Герпетологическая база данных» . Шведский музей естественной истории.
- BBC Nature: новости о змеях и видеоклипы из программ BBC прошлого и настоящего.
- Основы систематики змей в журнале «Жизнь коротка, но змеи длинны»
