Протеин Sonic Hedgehog
| Тсс | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | SHH , HHG1, HLP3, HPE3, MCOPCB5, SMMCI, TPT, TPTPS, sonic hedgehog, Sonic hedgehog, ShhNC, сигнальная молекула sonic hedgehog | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 600725 ; МГИ : 98297 ; Гомологен : 30961 ; Генные карты : SHH ; ОМА : SHH – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Белок Sonic hedgehog ( SHH ) кодируется SHH геном . [5] Белок назван в честь персонажа видеоигры Sonic the Hedgehog .
Эта сигнальная молекула играет ключевую роль в регуляции эмбрионального морфогенеза у всех животных. SHH контролирует органогенез и организацию центральной нервной системы, конечностей, пальцев и многих других частей тела. Sonic hedgehog — это морфоген , который моделирует развивающийся эмбрион с использованием градиента концентрации, характеризуемого моделью французского флага . [6] Эта модель имеет неоднородное распределение молекул SHH, которое управляет различной судьбой клеток в зависимости от концентрации. Мутации в этом гене могут вызвать голопрозэнцефалию — нарушение расщепления в полушариях головного мозга. [7] как было продемонстрировано в эксперименте с использованием мышей, нокаутных по SHH, у которых средняя линия переднего мозга не развивалась, и вместо этого образовался только один сросшийся телэнцефалический пузырек. [8]
Sonic hedgehog по-прежнему играет роль в дифференцировке, пролиферации и поддержании взрослых тканей. Аномальная активация передачи сигналов SHH во взрослых тканях связана с различными типами рака, включая рак молочной железы , кожи , головного мозга , печени , желчного пузыря и многих других. [9]
Открытие и присвоение имени
[ редактировать ]Ген ежа ( hh ) был впервые идентифицирован у плодовой мухи Drosophila melanogaster в классических гейдельбергских скринингах Кристианы Нюсляйн-Фольхард и Эрика Вишауса , опубликованных в 1980 году. [10] Эти скрининги , благодаря которым исследователи получили Нобелевскую премию в 1995 году вместе с генетиком развития Эдвардом Б. Льюисом , идентифицировали гены, которые контролируют структуру сегментации эмбрионов дрозофилы . Мутантный фенотип hh с потерей функции приводит к тому, что эмбрионы покрываются зубчиками, т.е. небольшими заостренными выступами, напоминающими шипы ежа . Исследования, направленные на поиск эквивалента ежа у позвоночных, проведенные Филипом Ингэмом , Эндрю П. МакМахоном и Клиффордом Табином, выявили три гомологичных гена . [11] [12] [13] [14]
Два из этих генов, Desert Hedgehog и Indian Hedgehog , были названы в честь вида ежей, а sonic hedgehog был назван в честь персонажа видеоигры Sonic the Hedgehog . [15] [16] Ген был назван Робертом Риддлом, научным сотрудником лаборатории Табина , после того, как его жена Бетси Уайлдер пришла домой с журналом, содержащим рекламу первой игры серии, Sonic the Hedgehog (1991). [17] [18] [19] У рыбок данио два из трех генов hh дублированы позвоночных: SHH a [20] и ШХ б [21] (ранее описывавшийся как ежик Тиггивинкль , названный в честь миссис Тигги-Винкль , персонажа из Беатрикс Поттер детских книг ) и ihha и ihhb [22] (ранее описывавшийся как ёжик ехидны , названный в честь колючего муравьеда , а не в честь персонажа Ехидны Наклза из «Соник» франшизы ).
Функция
[ редактировать ]что из hh гомологов SHH Было обнаружено, играет наиболее важную роль в развитии, действуя как морфоген, участвующий в формировании паттерна многих систем, включая переднюю долю гипофиза , [23] мантия головного мозга, [24] спинной мозг , [25] легкие , [26] зубы [27] и таламус зоной - интраталамусной . [28] [29] У позвоночных развитие конечностей пальцев и , расположенной на задней стороне зависит от секреции звукового ежа зоной поляризующей активности эмбрионального зачатка конечности . [13] Мутации в гене звукового ежа человека SHH вызывают голопрозэнцефалию типа 3 HPE3 в результате потери вентральной срединной линии. Путь транскрипции sonic hedgehog также связан с образованием определенных видов раковых опухолей, включая эмбриональную мозжечка . опухоль [30] и медуллобластома , [31] а также прогрессирование опухолей простаты . [32] Для экспрессии SHH в развивающихся конечностях эмбриона морфоген, называемый факторами роста фибробластов должен секретироваться из апикального эктодермального гребня . [33]
Также было показано, что звуковой еж действует как сигнал для наведения аксонов . Было продемонстрировано, что SHH привлекает комиссуральные аксоны на вентральной средней линии развивающегося спинного мозга. [34] В частности, SHH привлекает аксоны ганглиозных клеток сетчатки (RGC) при низких концентрациях и отталкивает их при более высоких концентрациях. [35] Было показано, что отсутствие (неэкспрессия) SHH контролирует рост зарождающихся задних конечностей у китообразных. [36] ( киты и дельфины ).
Ген SHH является членом семейства генов hedgehog с пятью вариантами изменений последовательности ДНК или вариантами сплайсинга. [37] SHH расположен на седьмой хромосоме и инициирует выработку белка Sonic Hedgehog. [37] Этот белок посылает сигналы ближнего и дальнего действия эмбриональным тканям для регулирования развития. [38] Если ген SHH мутирован или отсутствует, белок Sonic Hedgehog не сможет должным образом выполнять свою работу. Звуковой ёж способствует росту клеток, их спецификации и формированию, структурированию и организации строения тела. [39] Этот белок функционирует как жизненно важная морфогенная сигнальная молекула и играет важную роль в формировании множества различных структур в развивающихся эмбрионах. [39] Ген SHH влияет на несколько основных систем органов, таких как нервная система, сердечно-сосудистая система, дыхательная система и костно-мышечная система. [37] [38] Мутации в гене SHH могут вызвать пороки развития компонентов этих систем, что может привести к серьезным проблемам в развивающемся эмбрионе. Например, мутации этого гена могут существенно повлиять на мозг и глаза и вызвать такие расстройства, как микрофтальмия и голопрозэнцефалия . [39] Микрофтальмия — это заболевание, поражающее глаза, которое приводит к появлению небольших, недоразвитых тканей в одном или обоих глазах. [39] Это может привести к различным проблемам: от колобомы до единственного маленького глаза или полного отсутствия глаз. [38] Голопрозэнцефалия — это состояние, которое чаще всего вызывается мутацией гена SHH , которая вызывает неправильное разделение или поворот левого и правого полушария мозга. [40] и дисморфия лица. [38] [39] Многие системы и структуры в значительной степени полагаются на правильную экспрессию гена SHH и последующего белка sonic hedgehog, благодаря чему он считается важным геном для развития.
Паттерн центральной нервной системы
[ редактировать ]Сигнальная молекула sonic hedgehog (SHH) принимает на себя различные роли в формировании паттерна центральной нервной системы (ЦНС) во время позвоночных развития . Одной из наиболее характерных функций SHH является его роль в индукции пластинки дна и различных типов вентральных клеток внутри нервной трубки . [41] Нотохорда — структура , полученная из осевой мезодермы , — вырабатывает SHH, который перемещается внеклеточно в вентральную область нервной трубки и дает указание этим клеткам формировать пластинку дна. [42] Другой взгляд на индукцию пластинки дна предполагает, что некоторые клетки-предшественники, расположенные в хорде, встраиваются в нервную пластинку до ее формирования, позже давая начало пластинке дна. [43]
Нервная трубка сама по себе является исходной основой ЦНС позвоночных , а пластинка дна представляет собой специализированную структуру, расположенную в вентральной средней точке нервной трубки. Доказательства, подтверждающие, что хорда является сигнальным центром, получены в исследованиях, в которых вторая хорда имплантируется рядом с нервной трубкой in vivo, что приводит к образованию эктопической пластинки дна внутри нервной трубки. [44]
 Градиенты SHH и BMP в нервной трубке позвоночных
Градиенты SHH и BMP в нервной трубке позвоночных Эктопическое образование пластинки дна
Эктопическое образование пластинки дна Вентральные нервные домены нервной трубки
Вентральные нервные домены нервной трубки



Sonic hedgehog — это секретируемый белок , который опосредует сигнальную активность хорды и пластинки дна. [45] Исследования, включающие эктопическую экспрессию SHH in vitro [46] и в естественных условиях [47] индукции пластинки дна и дифференцировке мотонейронов приводят к и вентральных интернейронов . С другой стороны, у мышей, мутантов по SHH, отсутствуют характеристики вентрального отдела спинного мозга. [48] in vitro Блокирование передачи сигналов SHH с использованием антител против него демонстрирует сходные фенотипы. [47] SHH оказывает свое воздействие в зависимости от концентрации. [49] так что высокая концентрация SHH приводит к местному ингибированию клеточной пролиферации . [50] Это торможение приводит к тому, что пластинка дна становится тоньше по сравнению с боковыми областями нервной трубки . Более низкая концентрация SHH приводит к клеточной пролиферации и индукции различных типов вентральных нервных клеток. [47] Как только пластинка дна установится, клетки, находящиеся в этой области, впоследствии сами будут экспрессировать SHH. [50] создание градиента концентрации внутри нервной трубки.
SHH нет Хотя прямых доказательств существования градиента , косвенные доказательства имеются в виде визуализации экспрессии гена Patched ( Ptc ) , который кодирует лиганд- связывающий домен рецептора SHH. [51] по всей вентральной части нервной трубки. [52] Исследования in vitro показывают, что постепенные двух- и трехкратные изменения концентрации SHH приводят к появлению двигательных нейронов и различных подтипов межнейронов, обнаруженных в вентральном отделе спинного мозга. [53] Эти дополнительные изменения in vitro соответствуют расстоянию доменов от сигнальной ткани (хорды и пластинки дна), которая впоследствии дифференцируется в разные подтипы нейронов, как это происходит in vitro . [54] Предполагается, что градуированная передача сигналов SHH опосредуется через Gli семейство белков , которые являются гомологами позвоночных дрозофилы цинковые пальцы, , содержащего транскрипционного фактора Cubitus прерывания ( Ci ). Ci является важнейшим медиатором передачи сигналов hedgehog ( Hh ) у дрозофилы . [55] У позвоночных присутствуют три различных белка Gli, а именно. Gli1 , Gli2 и Gli3 , которые экспрессируются в нервной трубке. [56] У мышей, мутировавших по Gli1, наблюдается нормальное развитие спинного мозга, что указывает на то, что он не является посредником в обеспечении активности SHH. [57] Однако у мышей с мутацией Gli2 наблюдаются аномалии в вентральной части спинного мозга с тяжелыми дефектами в пластинке дна и самых вентральных интернейронах (V3). [58] Gli3 противодействует функции SHH дозозависимым образом, способствуя развитию подтипов дорсальных нейронов. Фенотипы мутантов SHH можно спасти с помощью двойного мутанта SHH/Gli3 . [59] Белки Gli имеют С-концевой домен активации и N-концевой репрессивный домен. [56] [60]
Предполагается, что SHH способствует активации функции Gli2 и ингибирует репрессивную активность Gli3. SHH также, по-видимому, способствует функции активации Gli3, но эта активность недостаточно сильна. [59] Ступенчатая концентрация SHH приводит к ступенчатой активности Gli 2 и Gli3, которые способствуют развитию вентральных и дорсальных подтипов нейронов в вентральном отделе спинного мозга. Данные, полученные от мутантов Gli3 и SHH/Gli3, показывают, что SHH в первую очередь регулирует пространственное ограничение доменов- предшественников , а не является индуктивным, поскольку мутанты SHH/Gli3 демонстрируют перемешивание типов клеток. [59] [61]
SHH также индуцирует другие белки, с которыми он взаимодействует, и эти взаимодействия могут влиять на чувствительность клетки к SHH. Белок, взаимодействующий с Hedgehog ( HHIP ), индуцируется SHH, что, в свою очередь, ослабляет его сигнальную активность. [62] Витронектин — еще один белок, индуцируемый SHH; он действует как обязательный кофактор передачи сигналов SHH в нервной трубке. [63]
В вентральной части нервной трубки имеется пять различных доменов-предшественников: V3 интернейроны , мотонейроны (MN), интернейроны V2 , V1 и V0 (в вентральном и дорсальном порядке). [53] Эти различные домены-предшественники устанавливаются посредством «связи» между различными классами гомеобокса транскрипционных факторов . (См. Тройничный нерв .) Эти факторы транскрипции реагируют на концентрацию градиента SHH. В зависимости от характера взаимодействия с SHH они подразделяются на две группы — класс I и класс II — и состоят из представителей семейств Pax , Nkx , Dbx и Irx . [50] Белки класса I репрессируются при разных порогах SHH, очерчивающих вентральные границы доменов-предшественников , тогда как белки класса II активируются при разных порогах SHH, очерчивающих дорсальную границу доменов. Избирательные перекрестно -репрессивные взаимодействия между белками класса I и класса II приводят к появлению пяти кардинальных вентральных подтипов нейронов. [64]
Важно отметить, что SHH — не единственная сигнальная молекула, оказывающая влияние на развивающуюся нервную трубку. Многие другие молекулы, пути и механизмы активны (например, RA , FGF , BMP ), и возможны сложные взаимодействия между SHH и другими молекулами. Предполагается, что BMP играют решающую роль в определении чувствительности нервных клеток к передаче сигналов SHH. Доказательства, подтверждающие это, получены в исследованиях с использованием ингибиторов BMP, которые изменяют судьбу клеток нервной пластинки при заданной концентрации SHH. [65] С другой стороны, мутация в антагонистах BMP (например, noggin ) вызывает серьезные дефекты самых вентральных характеристик спинного мозга, за которыми следует эктопическая экспрессия BMP в вентральной части нервной трубки. [66] Взаимодействия SHH с Fgf и RA еще не изучены детально на молекулярном уровне.
Морфогенетическая активность
[ редактировать ]Зависимая от концентрации и времени, определяющая судьбу клеток активность SHH в вентральной части нервной трубки делает его ярким примером морфогена . У позвоночных передача сигналов SHH в вентральной части нервной трубки наиболее ответственна за индукцию клеток пластинки дна и мотонейронов . [67] SHH исходит из хорды и вентральной пластинки дна развивающейся нервной трубки, создавая градиент концентрации , который охватывает дорсо-вентральную ось и противодействует обратному градиенту Wnt , который определяет дорсальный отдел спинного мозга. [68] [69] Более высокие концентрации лиганда SHH обнаруживаются в наиболее вентральных частях нервной трубки и хорды, тогда как более низкие концентрации обнаруживаются в более дорсальных областях нервной трубки. [68] Градиент концентрации SHH был визуализирован в нервной трубке мышей, сконструированных для экспрессии слитого белка SHH::GFP, чтобы продемонстрировать это градуированное распределение SHH во время формирования вентрального паттерна нервной трубки. [70]
Считается, что градиент SHH работает, вызывая множество различных судеб клеток с помощью зависящего от концентрации и времени механизма, который индуцирует множество факторов транскрипции в вентральных клетках-предшественниках . [68] [70] Каждый из вентральных доменов-предшественников экспрессирует высоко индивидуализированную комбинацию транскрипционных факторов — Nkx2.2, Olig2, Nkx6.1, Nkx6.2, Dbx1, Dbx2, Irx3, Pax6 и Pax7 — которая регулируется градиентом SHH. Эти факторы транскрипции индуцируются последовательно по градиенту концентрации SHH в зависимости от количества и времени воздействия лиганда SHH. [68] Поскольку каждая популяция клеток-предшественников реагирует на разные уровни белка SHH, они начинают экспрессировать уникальную комбинацию транскрипционных факторов, которая приводит к дифференциации судеб нейрональных клеток. Эта SHH-индуцированная дифференциальная экспрессия генов создает четкие границы между дискретными доменами экспрессии транскрипционных факторов, которые в конечном итоге формируют структуру вентральной нервной трубки. [68]
Пространственный и временной аспект прогрессирующей индукции генов и судеб клеток в вентральной части нервной трубки иллюстрируется доменами экспрессии двух наиболее хорошо охарактеризованных транскрипционных факторов, Olig2 и Nkx2.2. [68] На ранних стадиях развития клетки вентральной срединной линии подвергаются воздействию низких концентраций SHH только в течение относительно короткого времени и экспрессируют транскрипционный фактор Olig2. [68] Экспрессия Olig2 быстро расширяется в дорсальном направлении одновременно с непрерывным дорсальным расширением градиента SHH с течением времени. [68] Однако по мере того, как морфогенетический фронт лиганда SHH перемещается и начинает расти более концентрированно, клетки, которые подвергаются воздействию более высоких уровней лиганда, реагируют выключением Olig2 и включением Nkx2.2, [68] создавая резкую границу между клетками, экспрессирующими фактор транскрипции Nkx2.2, вентрально по отношению к клеткам, экспрессирующим Olig2. Таким образом, считается, что каждый из доменов шести популяций клеток-предшественников последовательно формируется по всей нервной трубке градиентом концентрации SHH. [68] Взаимное ингибирование пар транскрипционных факторов, экспрессируемых в соседних доменах, способствует развитию резких границ; однако в некоторых случаях ингибирующая связь обнаруживалась даже между парами транскрипционных факторов из более удаленных доменов. В частности, NKX2-2, сообщается, что экспрессируемый в домене V3, ингибирует IRX3, экспрессируемый в V2 и других дорсальных доменах, хотя V3 и V2 разделены дополнительным доменом, называемым MN. [71]
Экспрессия SHH в лобно-носовой эктодермальной зоне (FEZ), которая является сигнальным центром, ответственным за паттерн развития верхней челюсти, регулирует краниофациальное развитие, опосредованно через семейство miR-199 в FEZ. В частности, SHH-зависимые сигналы головного мозга регулируют гены семейства miR-199, при этом подавление генов miR-199 увеличивает экспрессию SHH и приводит к более широким лицам, в то время как активация генов miR-199 снижает экспрессию SHH, что приводит к узким лицам. [72]
Развитие зубов
[ редактировать ]SHH играет важную роль в органогенезе и, что наиболее важно, черепно-лицевом развитии. Поскольку SHH является сигнальной молекулой, он в основном действует путем диффузии по градиенту концентрации, по-разному воздействуя на клетки. На ранних стадиях развития зубов SHH высвобождается из первичного эмалевого узла — сигнального центра — для предоставления позиционной информации как в латеральном, так и в плоскостном сигнальном паттерне при развитии зубов и регуляции роста зубных бугорков. [73] SHH, в частности, необходим для роста эпителиальных петель шейки матки, где внешний и внутренний эпителий соединяются и образуют резервуар для зубных стволовых клеток. После апоптоза первичных эмалевых узлов образуются вторичные эмалевые узлы. Вторичные узлы эмали секретируют SHH в сочетании с другими сигнальными молекулами, чтобы утолщать оральную эктодерму и начинать формировать сложные формы коронки зуба во время дифференцировки и минерализации. [74] В модели нокаутного гена отсутствие SHH указывает на голопрозэнцефалию . Однако SHH активирует нижестоящие молекулы Gli2 и Gli3. Мутантные эмбрионы Gli2 и Gli3 имеют аномальное развитие резцов, которые задерживаются на ранних стадиях развития зубов, а также маленьких коренных зубов. [75]
Развитие легких
[ редактировать ]Хотя SHH чаще всего связан с развитием мозга и пальцев конечностей, он также важен для развития легких. [76] [77] [78] [79] Исследования с использованием кПЦР и нокаутов продемонстрировали, что SHH способствует эмбриональному развитию легких. Ветвление легких млекопитающих происходит в эпителии развивающихся бронхов и легких. [80] [81] SHH экспрессируется по всей энтодерме передней кишки (самый внутренний из трех зародышевых листков) в дистальном эпителии, где развиваются эмбриональные легкие. [78] [81] Это позволяет предположить, что SHH частично ответственен за ветвление легких. Дополнительные доказательства роли SHH в разветвлении легких были получены с помощью кПЦР. Экспрессия SHH происходит в развивающихся легких примерно на 11-й день эмбриона и сильно выражена в зачатках легких плода, но низкая в развивающихся бронхах. [78] [81] У мышей с дефицитом SHH может развиться трахеопищеводная фистула (аномальное соединение пищевода и трахеи). [82] [78] Кроме того, модель мышей с двойным нокаутом (SHH-/- ) демонстрировала плохое развитие легких. В легких двойного нокаута SHH не наблюдалось дольчатости и ветвления (т.е. в аномальных легких развивалась только одна ветвь по сравнению с сильно разветвленным фенотипом дикого типа). [78]
Потенциальная регенеративная функция
[ редактировать ]Sonic hedgehog может играть роль в волосковых клеток регенерации млекопитающих . Модулируя активность белка ретинобластомы в улитке крысы, sonic hedgehog позволяет зрелым волосковым клеткам , которые обычно не могут вернуться в пролиферативное состояние, делиться и дифференцироваться. Белки ретинобластомы подавляют рост клеток, не давая клеткам вернуться в клеточный цикл , тем самым предотвращая пролиферацию. Подавление активности Rb, по-видимому, позволяет клеткам делиться. Таким образом, звуковой еж, известный как важный регулятор Rb, также может оказаться важным фактором в восстановлении волосковых клеток после повреждения. [83]
SHH важен для регуляции дермального адипогенеза клетками, усиливающими транзит волосяных фолликулов (HF-TAC). В частности, SHH индуцирует дермальный ангиогенез, воздействуя непосредственно на предшественников адипоцитов и способствуя их пролиферации посредством экспрессии гена рецептора γ, активируемого пролифератором пероксисом (Pparg). [84]
Обработка
[ редактировать ]SHH подвергается ряду стадий обработки, прежде чем он будет секретирован из клетки. Недавно синтезированный SHH весит 45 кДа и называется препробелком. Как секретируемый белок, он содержит короткую сигнальную последовательность на своем N-конце, которая распознается частицей распознавания сигнала во время транслокации в эндоплазматический ретикулум (ЭР), что является первым этапом секреции белка . После завершения транслокации сигнальная последовательность удаляется сигнальной пептидазой в ЭР. Там SHH подвергается автопроцессингу с образованием N-концевого сигнального домена массой 20 кДа (SHH-N) и C-концевого домена массой 25 кДа без известной сигнальной роли. [85] Расщепление катализируется протеазой С -концевого домена. В ходе реакции холестерина . к С-концу SHH-N присоединяется молекула [86] [87] Таким образом, С-концевой домен действует как интеин и холестерин-трансфераза. Другая гидрофобная группа , пальмитат , добавляется к альфа-амину N-концевого цистеина SHH-N. Эта модификация необходима для эффективной передачи сигнала, что приводит к 30-кратному увеличению эффективности по сравнению с непальмитилированной формой и осуществляется членом мембраносвязанного семейства О-ацилтрансфераз протеинцистеин-N-пальмитоилтрансферазой HHAT . [88]
Роботникинин
[ редактировать ]Был обнаружен потенциальный ингибитор сигнального пути Hedgehog, получивший название «Роботникинин» — в честь заклятого врага Sonic the Hedgehog и главного антагониста серии игр Sonic the Hedgehog, доктора Иво «Эггмана» Роботника . [89]
Бывшие споры вокруг имени
[ редактировать ]Этот ген связан с состоянием, известным как голопрозэнцефалия , которое может привести к серьезным дефектам головного мозга, черепа и лица, из-за чего некоторые врачи и ученые критикуют это название на том основании, что оно звучит слишком легкомысленно. Было отмечено, что упоминание о мутации гена звукового ежа может быть не очень хорошо воспринято при обсуждении серьезного заболевания с пациентом или его семьей. [17] [90] [91] Этот спор в значительной степени утих, и теперь это название обычно рассматривается как юмористический пережиток времени, когда еще не было быстрого и дешевого полного секвенирования генома и стандартизированной номенклатуры. [92] Проблему «неуместности» названий генов, таких как « Матери против декапентаплегии» , « Безумный край » и «Соник-ёж», в значительной степени можно избежать, используя стандартизированные сокращения при разговоре с пациентами и их семьями. [93]
Галерея
[ редактировать ] |  | 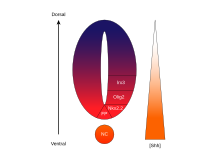 |
См. также
[ редактировать ]- Пикачурин , белок сетчатки, названный в честь Пикачу.
- Zbtb7 , онкоген, который первоначально назывался « Покемон ».
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000164690 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000002633 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Мариго В., Робертс DJ, Ли С.М., Цукуров О, Леви Т, Гастье Дж.М., Эпштейн DJ, Гилберт DJ, Коупленд Н.Г., Зейдман CE (июль 1995 г.). «Клонирование, экспрессия и хромосомное расположение SHH и IHH: два человеческих гомолога гена полярности сегмента дрозофилы hedgehog». Геномика . 28 (1): 44–51. дои : 10.1006/geno.1995.1104 . ПМИД 7590746 .
- ^ Джагер Дж., Мартинес-Ариас А. (март 2009 г.). «Получение меры позиционной информации» . ПЛОС Биология . 7 (3): е81. дои : 10.1371/journal.pbio.1000081 . ПМК 2661971 . ПМИД 19338391 .
- ^ Нанни Л., Минг Дж.Э., Боциан М., Стейнхаус К., Бьянки Д.В., Ди-Смолдерс С., Джаннотти А., Имаидзуми К., Джонс К.Л., Кампо М.Д., Мартин Р.А., Майнеке П., Пирпонт М.Е., Робин Н.Х., Янг И.Д., Росслер Э., Муенке М. (декабрь 1999 г.). «Мутационный спектр гена sonic hedgehog при голопрозэнцефалии: мутации SHH вызывают значительную долю аутосомно-доминантной голопрозэнцефалии» . Молекулярная генетика человека . 8 (13): 2479–2488. дои : 10.1093/hmg/8.13.2479 . ПМИД 10556296 .
- ^ Блесс С., Сабо Н., Хаддад-Товоли Р., Чжоу Х., Альварес-Боладо Г. (06.01.2015). «Передача сигналов звукового ежа в развитии гипоталамуса мыши» . Границы нейроанатомии . 8 : 156. дои : 10.3389/fnana.2014.00156 . ПМК 4285088 . ПМИД 25610374 .
- ^ Дженг К.С., Чанг К.Ф., Линь СС (январь 2020 г.). «Передача сигналов Sonic Hedgehog в органогенезе, опухолях и опухолевом микроокружении» . Международный журнал молекулярных наук . 21 (3): 758. doi : 10.3390/ijms21030758 . ПМК 7037908 . ПМИД 31979397 .
- ^ Нюсляйн-Фольхард С., Вишаус Е (октябрь 1980 г.). «Мутации, влияющие на количество и полярность сегментов у дрозофилы». Природа . 287 (5785): 795–801. Бибкод : 1980Natur.287..795N . дои : 10.1038/287795a0 . ПМИД 6776413 . S2CID 4337658 .
- ^ Краусс С., Конкордет Дж.П., Ингхэм П.В. (декабрь 1993 г.). «Функционально консервативный гомолог гена полярности сегмента дрозофилы hh экспрессируется в тканях с поляризующей активностью эмбрионов рыбок данио». Клетка . 75 (7): 1431–1444. дои : 10.1016/0092-8674(93)90628-4 . ПМИД 8269519 . S2CID 46266627 .
- ^ Эшелард Ю., Эпштейн Д.Д., Сен-Жак Б., Шен Л., Молер Дж., МакМахон Дж.А., МакМахон А.П. (декабрь 1993 г.). «Sonic hedgehog, член семейства предполагаемых сигнальных молекул, участвует в регуляции полярности ЦНС». Клетка . 75 (7): 1417–1430. дои : 10.1016/0092-8674(93)90627-3 . ПМИД 7916661 . S2CID 6732599 .
- ^ Перейти обратно: а б Риддл Р.Д., Джонсон Р.Л., Лауфер Э., Табин С. (декабрь 1993 г.). «Звуковой ёж опосредует поляризующую активность ZPA». Клетка . 75 (7): 1401–1416. дои : 10.1016/0092-8674(93)90626-2 . ПМИД 8269518 . S2CID 4973500 .
- ^ Анжер Н. (11 января 1994 г.). «Биологи находят ключевые гены, которые формируют структуру эмбрионов» . Нью-Йорк Таймс . Архивировано из оригинала 17 августа 2017 г. Проверено 18 февраля 2017 г.
- ^ Анвуд Р. (6 сентября 2007 г.). Эму не может идти назад . Эбери Пресс. стр. 113–114. ISBN 978-0-09-192151-4 . Архивировано из оригинала 7 мая 2017 г. Проверено 6 октября 2016 г.
- ^ Симонит Т (15 декабря 2005 г.). «Покемон блокирует имя гена» . Природа . 438 (897): 897. Бибкод : 2005Natur.438..897S . дои : 10.1038/438897a . ПМИД 16355177 . S2CID 37632823 .
- ^ Перейти обратно: а б «Ген по имени Соник» . Нью-Йорк Таймс . 11 января 1994 г. Архивировано из оригинала 28 февраля 2018 г. Проверено 18 февраля 2017 г.
- ^ Кин А., Табин С. (12 апреля 2004 г.). «Клифф Тейбин: Интервью о Супер Сонике» . Еженедельный шум. Архивировано из оригинала 10 ноября 2005 года . Проверено 24 апреля 2014 г.
- ^ Риддл Р. Гениально: Ген Циклопа . BBC.co.uk. Беседовала Кэт Арни. Радио Би-би-си.
- ^ «Данио ЩХа» . Университет Орегона. Архивировано из оригинала 25 июня 2009 г.
- ^ «Данио ШХб» . Университет Орегона. Архивировано из оригинала 26 июня 2009 г.
- ^ Карри П.Д., Ингхэм П.В. (август 1996 г.). «Индукция определенного типа мышечных клеток белком, похожим на ежа, у рыбок данио». Природа . 382 (6590): 452–455. Бибкод : 1996Natur.382..452C . дои : 10.1038/382452a0 . ПМИД 8684485 . S2CID 4271898 .
- ^ Херцог В., Цзэн Х., Леле З., Зоннтаг С., Тинг Дж.В., Чанг С.И., Хаммершмидт М. (февраль 2003 г.). «Формирование аденогипофиза у рыбок данио и его зависимость от звукового ежа» . Биология развития . 254 (1): 36–49. дои : 10.1016/S0012-1606(02)00124-0 . ПМИД 12606280 .
- ^ Раш Б.Г., Гроув Э.А. (октябрь 2007 г.). «Образование спинного конечного мозга: роль звукового ежа?» . Журнал неврологии . 27 (43): 11595–11603. doi : 10.1523/JNEUROSCI.3204-07.2007 . ПМК 6673221 . ПМИД 17959802 .
- ^ Льюис К.Е., Эйзен Дж.С. (сентябрь 2001 г.). «Передача сигналов Hedgehog необходима для первичной индукции мотонейронов у рыбок данио». Разработка . 128 (18): 3485–3495. дои : 10.1242/dev.128.18.3485 . PMID 11566854 .
- ^ Вулперт Л. (2015). Принципы развития (5-е изд.). Издательство Оксфордского университета. п. 500.
- ^ Дассул Х.Р., Льюис П., Бей М., Маас Р., МакМахон А.П. (ноябрь 2000 г.). «Звуковой еж регулирует рост и морфогенез зуба». Разработка . 127 (22): 4775–4785. дои : 10.1242/dev.127.22.4775 . ПМИД 11044393 .
- ^ Шолпп С., Вольф О., Бранд М., Ламсден А. (март 2006 г.). «Передача сигналов ежей из zona limitans Intrathalamica управляет формированием паттерна промежуточного мозга рыбок данио» . Разработка . 133 (5): 855–864. дои : 10.1242/dev.02248 . ПМИД 16452095 . S2CID 27550686 .
- ^ Раш Б.Г., Гроув Э.А. (ноябрь 2011 г.). «Shh и Gli3 регулируют образование телэнцефально-диэнцефального соединения и подавляют перешеек-подобный источник сигналов в переднем мозге» . Биология развития . 359 (2): 242–250. дои : 10.1016/j.ydbio.2011.08.026 . ПМЦ 3213684 . ПМИД 21925158 .
- ^ Тейлор М.Д., Норткотт П.А., Коршунов А., Ремке М., Чо Ю.Дж., Клиффорд С.С., Эберхарт К.Г., Парсонс Д.В., Рутковски С., Гаджар А., Эллисон Д.В., Лихтер П., Гилбертсон Р.Дж., Померой С.Л., Кул М., Пфистер С.М. (апрель 2012 г.) ). «Молекулярные подгруппы медуллобластомы: современный консенсус» . Акта Нейропатологика . 123 (4): 465–472. дои : 10.1007/s00401-011-0922-z . ПМЦ 3306779 . ПМИД 22134537 .
- ^ ДеСуза Р.М., Джонс Б.Р., Лоуис С.П., Куриан К.М. (22 июля 2014 г.). «Детская медуллобластома - обновленная информация о молекулярной классификации, способствующая таргетной терапии» . Границы онкологии . 4 : 176. doi : 10.3389/fonc.2014.00176 . ПМК 4105823 . ПМИД 25101241 .
- ^ Любик А.А., Нури М., Труонг С., Гаффари М., Адомат Х.Х., Кори Э., Кокс М.Э., Ли Н., Ганс Э.С., Йенки П., Фам С., Буттян Р. (январь 2017 г.). «Паракринная передача сигналов звукового ежа вносит значительный вклад в приобретенный стероидогенез в микроокружении опухоли простаты» . Международный журнал рака . 140 (2): 358–369. дои : 10.1002/ijc.30450 . ПМИД 27672740 . S2CID 2354209 .
- ^ Риддл Р.Д., Табин С. (февраль 1999 г.). «Как развиваются конечности». Научный американец . 280 (2): 74–79. Бибкод : 1999SciAm.280b..74R . doi : 10.1038/scientificamerican0299-74 . ПМИД 9924814 .
- ^ Чаррон Ф., Стайн Э., Чон Дж., МакМахон А.П., Тессье-Лавин М. (апрель 2003 г.). «Морфоген sonic hedgehog представляет собой аксональный хемоаттрактант, который сотрудничает с нетрином-1 в управлении аксонами по средней линии» . Клетка . 113 (1): 11–23. дои : 10.1016/S0092-8674(03)00199-5 . ПМИД 12679031 . S2CID 13909497 .
- ^ Колпак А., Чжан Дж., Бао З.З. (март 2005 г.). «Sonic hedgehog оказывает двойное влияние на рост аксонов ганглиев сетчатки в зависимости от его концентрации» . Журнал неврологии . 25 (13): 3432–3441. doi : 10.1523/JNEUROSCI.4938-04.2005 . ПМК 1564194 . ПМИД 15800198 .
- ^ Тьювиссен Дж.Г., Кон М.Дж., Стивенс Л.С., Баджпай С., Хейнинг Дж., Хортон В.Е. (май 2006 г.). «Основы развития потери задних конечностей у дельфинов и происхождение строения тела китообразных» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (22): 8414–8418. Бибкод : 2006PNAS..103.8414T . дои : 10.1073/pnas.0602920103 . ПМК 1482506 . ПМИД 16717186 .
- ^ Перейти обратно: а б с «ENSG00000164690» . ГРЧ38 . Выпуск Ensembl 99. Архивировано из оригинала 1 ноября 2020 года . Проверено 9 апреля 2020 г.
- ^ Перейти обратно: а б с д "UniprotKB - Q15465 (SHH_HUMAN)" . ЮниПрот . Консорциум ЮниПрот. Архивировано из оригинала 31 мая 2020 года . Проверено 9 апреля 2020 г.
- ^ Перейти обратно: а б с д и «Ген SHH» . Домашний справочник по генетике . Национальная медицинская библиотека США. Архивировано из оригинала 2 апреля 2020 года . Проверено 9 апреля 2020 г.
- ^ де Люссане М.Х., Оссе Дж.В. (2012). «Наследственное осевое скручивание объясняет контралатеральную переднюю часть и перекрест зрительных нервов у позвоночных». Биология животных . 62 (2): 193–216. arXiv : 1003.1872 . дои : 10.1163/157075611X617102 . S2CID 7399128 .
- ^ Литингтунг И., Чан С. (октябрь 2000 г.). «Контроль активности Shh и передачи сигналов в нервной трубке». Динамика развития . 219 (2): 143–154. doi : 10.1002/1097-0177(2000)9999:9999<::AID-DVDY1050>3.0.CO;2-Q . ПМИД 11002335 . S2CID 221646338 .
- ^ Плачек М. (август 1995 г.). «Роль хорды и пластинки дна в индуктивных взаимодействиях». Текущее мнение в области генетики и развития . 5 (4): 499–506. дои : 10.1016/0959-437X(95)90055-L . ПМИД 7580143 .
- ^ Тейе М.А., Лапуант Ф., Ле Дуарен Н.М. (сентябрь 1998 г.). «Пересмотр взаимосвязей между хордой и пластинкой дна в развитии позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (20): 11733–11738. Бибкод : 1998PNAS...9511733T . дои : 10.1073/pnas.95.20.11733 . ПМК 21709 . ПМИД 9751734 .
- ^ ван Страатен Х.В., Хеккинг Дж.В., Торс Ф., Вирц-Хессельс Э.Л., Друккер Дж. (октябрь 1985 г.). «Индукция дополнительной пластинки дна в нервной трубке». Acta Morphologica Neerlando-Scandinavica . 23 (2): 91–97. ПМИД 3834777 .
- ^ Паттен И., Плачек М. (ноябрь 2000 г.). «Роль ежа Соника в формировании рисунка нервной трубки» . Клеточные и молекулярные науки о жизни . 57 (12): 1695–1708. дои : 10.1007/PL00000652 . ПМЦ 11146859 . ПМИД 11130176 . S2CID 20950575 .
- ^ Марти Э., Бамкрот Д.А., Такада Р., МакМахон А.П. (май 1995 г.). «Потребность в форме 19K Sonic hedgehog для индукции различных типов вентральных клеток в эксплантатах ЦНС». Природа . 375 (6529): 322–325. Бибкод : 1995Natur.375..322M . дои : 10.1038/375322a0 . ПМИД 7753196 . S2CID 4362006 .
- ^ Перейти обратно: а б с Эриксон Дж., Мортон С., Каваками А., Ролинк Х., Джесселл Т.М. (ноябрь 1996 г.). «Два критических периода передачи сигналов Sonic Hedgehog, необходимые для спецификации идентичности мотонейронов» . Клетка . 87 (4): 661–673. дои : 10.1016/S0092-8674(00)81386-0 . ПМИД 8929535 . S2CID 11578260 .
- ^ Чан С., Литингтунг Й., Ли Э., Янг К.Е., Корден Дж.Л., Вестфаль Х., Бичи П.А. (октябрь 1996 г.). «Циклопия и дефектное осевое паттернирование у мышей, у которых отсутствует функция гена Sonic hedgehog». Природа . 383 (6599): 407–413. Бибкод : 1996Natur.383..407C . дои : 10.1038/383407a0 . ПМИД 8837770 . S2CID 4339131 .
- ^ Плачек М., Тессье-Лавин М., Ямада Т., Джесселл Т., Додд Дж. (ноябрь 1990 г.). «Мезодермальный контроль идентичности нервных клеток: индукция пластинки дна хордой». Наука . 250 (4983): 985–988. Бибкод : 1990Sci...250..985P . дои : 10.1126/science.2237443 . ПМИД 2237443 .
- ^ Перейти обратно: а б с Уилсон Л., Мэден М. (июнь 2005 г.). «Механизмы формирования дорсовентрального паттерна в нервной трубке позвоночных» . Биология развития . 282 (1): 1–13. дои : 10.1016/j.ydbio.2005.02.027 . ПМИД 15936325 .
- ^ Стоун Д.М., Хайнс М., Арманини М., Суонсон Т.А., Гу К., Джонсон Р.Л., Скотт М.П., Пенника Д., Годдард А., Филлипс Х., Нолл М., Хупер Дж.Е., де Соваж Ф., Розенталь А. (ноябрь 1996 г.). «Запатченный ген-супрессор опухоли кодирует кандидатный рецептор для ежа Соника». Природа . 384 (6605): 129–134. Бибкод : 1996Natur.384..129S . дои : 10.1038/384129a0 . ПМИД 8906787 . S2CID 4342540 .
- ^ Мариго В., Табин CJ (сентябрь 1996 г.). «Регуляция регуляции звукового ежа в развивающейся нервной трубке» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (18): 9346–9351. Бибкод : 1996PNAS...93.9346M . дои : 10.1073/pnas.93.18.9346 . ПМК 38430 . ПМИД 8790332 .
- ^ Перейти обратно: а б Эриксон Дж., Бриско Дж., Рашбасс П., ван Хейнинген В., Джесселл Т.М. (1997). «Градуированная передача сигналов звукового ежа и спецификация судьбы клеток в вентральной части нервной трубки». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 62 : 451–466. дои : 10.1101/SQB.1997.062.01.053 . ПМИД 9598380 .
- ^ Эриксон Дж., Рашбасс П., Шедл А., Бреннер-Мортон С., Каваками А., ван Хейнинген В., Джесселл Т.М., Бриско Дж. (июль 1997 г.). «Pax6 контролирует идентичность клеток-предшественников и судьбу нейронов в ответ на градуированную передачу сигналов Shh» . Клетка . 90 (1): 169–180. дои : 10.1016/S0092-8674(00)80323-2 . ПМИД 9230312 . S2CID 17054686 .
- ^ Лам Л., Бичи, Пенсильвания (июнь 2004 г.). «Сеть реагирования Hedgehog: датчики, коммутаторы и маршрутизаторы». Наука . 304 (5678): 1755–1759. Бибкод : 2004Sci...304.1755L . CiteSeerX 10.1.1.476.3902 . дои : 10.1126/science.1098020 . ПМИД 15205520 . S2CID 13949436 .
- ^ Перейти обратно: а б Руис и Альтаба А (июнь 1998 г.). «Комбинаторная функция гена Gli в пластинке дна и нейрональная индукция Sonic hedgehog». Разработка . 125 (12): 2203–2212. дои : 10.1242/dev.125.12.2203 . ПМИД 9584120 .
- ^ Пак Х.Л., Бай С., Платт К.А., Матис М.П., Бигли А., Хуэй К.С., Накашима М., Джойнер А.Л. (апрель 2000 г.). «Мутанты Gli1 мыши жизнеспособны, но имеют дефекты передачи сигналов SHH в сочетании с мутацией Gli2». Разработка . 127 (8): 1593–1605. дои : 10.1242/dev.127.8.1593 . ПМИД 10725236 .
- ^ Матис М.П., Эпштейн DJ, Парк Х.Л., Платт К.А., Джойнер А.Л. (август 1998 г.). «Gli2 необходим для индукции пластинки дна и прилегающих клеток, но не для большинства вентральных нейронов в центральной нервной системе мыши». Разработка . 125 (15): 2759–2770. дои : 10.1242/dev.125.15.2759 . ПМИД 9655799 .
- ^ Перейти обратно: а б с Литингтунг И., Чан С. (октябрь 2000 г.). «Спецификация типов вентральных нейронов опосредована антагонистическим взаимодействием между Shh и Gli3». Природная неврология . 3 (10): 979–985. дои : 10.1038/79916 . ПМИД 11017169 . S2CID 10102370 .
- ^ Сасаки Х., Нишизаки Ю., Хуэй С., Накафуку М., Кондо Х. (сентябрь 1999 г.). «Регуляция активности Gli2 и Gli3 с помощью аминоконцевого домена репрессии: роль Gli2 и Gli3 в качестве первичных медиаторов передачи сигналов Shh». Разработка . 126 (17): 3915–3924. дои : 10.1242/dev.126.17.3915 . ПМИД 10433919 .
- ^ Перссон М., Стаматаки Д., Те Вельшер П., Андерссон Э., Бёзе Дж., Рютер У., Эриксон Дж., Бриско Дж. (ноябрь 2002 г.). «Дорсально-вентральный паттерн спинного мозга требует активности репрессора транскрипции Gli3» . Гены и развитие . 16 (22): 2865–2878. дои : 10.1101/gad.243402 . ЧВК 187477 . ПМИД 12435629 .
- ^ Чуанг П.Т., МакМахон А.П. (февраль 1999 г.). «Передача сигналов Hedgehog у позвоночных, модулируемая индукцией белка, связывающего Hedgehog». Природа . 397 (6720): 617–621. Бибкод : 1999Natur.397..617C . дои : 10.1038/17611 . ПМИД 10050855 . S2CID 204991314 .
- ^ Понс С., Марти Э (январь 2000 г.). «Sonic hedgehog взаимодействует с белком внеклеточного матрикса витронектином, вызывая дифференцировку спинальных мотонейронов». Разработка . 127 (2): 333–342. дои : 10.1242/dev.127.2.333 . ПМИД 10603350 .
- ^ Бриско Дж., Пьерани А., Джесселл Т.М., Эриксон Дж. (май 2000 г.). «Код гомеодоменного белка определяет идентичность клеток-предшественников и судьбу нейронов в вентральной части нервной трубки» . Клетка . 101 (4): 435–445. дои : 10.1016/S0092-8674(00)80853-3 . ПМИД 10830170 . S2CID 17295764 .
- ^ Лием К.Ф., Джесселл Т.М., Бриско Дж. (ноябрь 2000 г.). «Регуляция активности формирования нейронного паттерна звукового ежа с помощью секретируемых ингибиторов BMP, экспрессируемых хордой и сомитами». Разработка . 127 (22): 4855–4866. дои : 10.1242/dev.127.22.4855 . ПМИД 11044400 .
- ^ МакМахон Дж.А., Такада С., Циммерман Л.Б., Фан СМ, Харланд Р.М., МакМахон А.П. (май 1998 г.). «Опосредованный Noggin антагонизм передачи сигналов BMP необходим для роста и формирования паттерна нервной трубки и сомитов» . Гены и развитие . 12 (10): 1438–1452. дои : 10.1101/gad.12.10.1438 . ПМК 316831 . ПМИД 9585504 .
- ^ Ролинк Х., Портер Дж.А., Чан С., Танабэ Ю., Чанг Д.Т., Бичи П.А., Джесселл Т.М. (май 1995 г.). «Индукция пластинки пола и двигательных нейронов различными концентрациями аминоконцевого продукта расщепления аутопротеолиза звукового ежа» . Клетка . 81 (3): 445–455. дои : 10.1016/0092-8674(95)90397-6 . ПМИД 7736596 . S2CID 18937753 .
- ^ Перейти обратно: а б с д и ж г час я дж Рибес В., Бриско Дж. (август 2009 г.). «Установление и интерпретация ступенчатой передачи сигналов Sonic Hedgehog во время формирования паттерна нервной трубки позвоночных: роль отрицательной обратной связи» . Перспективы Колд-Спринг-Харбор в биологии . 1 (2): а002014. doi : 10.1101/cshperspect.a002014 . ПМК 2742090 . ПМИД 20066087 .
- ^ Мурояма И., Фудзихара М., Икея М., Кондо Х., Такада С. (март 2002 г.). «Передача сигналов Wnt играет важную роль в спецификации нейронов дорсального отдела спинного мозга» . Гены и развитие . 16 (5): 548–553. дои : 10.1101/gad.937102 . ПМК 155351 . ПМИД 11877374 .
- ^ Перейти обратно: а б Чемберлен С.Э., Чон Дж., Го С., Аллен Б.Л., МакМахон А.П. (март 2008 г.). «Происходящий из нотохорды Shh концентрируется в тесной связи с апикально расположенным базальным тельцем в нервных клетках-мишенях и образует динамический градиент во время формирования нейронного паттерна» . Разработка . 135 (6): 1097–1106. дои : 10.1242/dev.013086 . ПМИД 18272593 . S2CID 17431502 .
- ^ Ловрич А., Гао И., Юхас Б., Бок И., Бирн Х.М., Диньес А., Ковач К.А. (ноябрь 2014 г.). «Булево моделирование выявляет новые регуляторные связи между факторами транскрипции, управляющими развитием вентральной части спинного мозга» . ПЛОС ОДИН . 9 (11): е111430. Бибкод : 2014PLoSO...9k1430L . дои : 10.1371/journal.pone.0111430 . ПМЦ 4232242 . ПМИД 25398016 .
- ^ Ричбург Х.А., Ху Д.П., Сюй Ю, Барчак А.Дж., Маркучио Р.С. (сентябрь 2020 г.). «Семейство миР-199 способствует регуляции экспрессии звукового ежа во время черепно-лицевого развития» . Динамика развития . 249 (9): 1062–1076. дои : 10.1002/dvdy.191 . ПМЦ 7484444 . ПМИД 32391617 .
- ^ Нанси А. (2012). Гистология полости рта Тен Кейт: развитие, структура и функции (8-е изд.). Сент-Луис, Миссури: Эльзевир. ISBN 978-0-323-07846-7 .
- ^ Теслефф I (май 2003 г.). «Эпителиально-мезенхимальная передача сигналов, регулирующая морфогенез зубов» . Журнал клеточной науки . 116 (Часть 9): 1647–1648. дои : 10.1242/jcs.00410 . ПМИД 12665545 . S2CID 45648812 .
- ^ Хардкасл З., Мо Р., Хуэй CC, Шарп П.Т. (август 1998 г.). «Сигнальный путь Shh в развитии зубов: дефекты у мутантов Gli2 и Gli3». Разработка . 125 (15): 2803–2811. дои : 10.1242/dev.125.15.2803 . ПМИД 9655803 .
- ^ Вулперт Л. (2015). Принципы развития (5-е изд.). Издательство Оксфордского университета. п. 500. ИСБН 978-0-19-967814-3 .
- ^ Беллуски С., Фурута Ю., Раш М.Г., Хендерсон Р., Винньер Дж., Хоган Б.Л. (январь 1997 г.). «Участие Sonic hedgehog (Shh) в росте и морфогенезе эмбриональных легких мышей». Разработка . 124 (1): 53–63. дои : 10.1242/dev.124.1.53 . ПМИД 9006067 .
- ^ Перейти обратно: а б с д и Пепичелли К.В., Льюис П.М., МакМахон А.П. (сентябрь 1998 г.). «Sonic hedgehog регулирует морфогенез ветвления в легких млекопитающих» . Современная биология . 8 (19): 1083–1086. Бибкод : 1998CBio....8.1083P . дои : 10.1016/S0960-9822(98)70446-4 . ПМИД 9768363 . S2CID 12711144 .
- ^ Уайт AC, Сюй Дж, Инь Ю, Смит С, Шмид Г, Орниц ДМ (апрель 2006 г.). «Передача сигналов FGF9 и SHH координирует рост и развитие легких посредством регуляции отдельных мезенхимальных доменов» . Разработка . 133 (8): 1507–1517. дои : 10.1242/dev.02313 . ПМИД 16540513 . S2CID 23839558 .
- ^ Миура Т (2008). «Моделирование морфогенеза ветвления легких». Актуальные темы биологии развития . 81 : 291–310. дои : 10.1016/S0070-2153(07)81010-6 . ISBN 9780123742537 . ПМИД 18023732 .
- ^ Перейти обратно: а б с Куглер М.К., Джойнер А.Л., Лумис К.А., Мангер Дж.С. (январь 2015 г.). «Сигнализация звукового ежа в легких. От развития к болезни» . Американский журнал респираторной клеточной и молекулярной биологии . 52 (1): 1–13. дои : 10.1165/rcmb.2014-0132TR . ПМК 4370254 . ПМИД 25068457 .
- ^ Кардозо В.В., Лю Дж. (май 2006 г.). «Регуляция раннего морфогенеза легких: вопросы, факты и противоречия» . Разработка . 133 (9): 1611–1624. дои : 10.1242/dev.02310 . ПМИД 16613830 . S2CID 18195361 .
- ^ Лу Н, Чен Ю, Ван Цз, Чен Г, Линь Ц, Чэнь Цзы, Ли Х (январь 2013 г.). «Sonic hedgehog инициирует регенерацию волосковых клеток улитки посредством подавления белка ретинобластомы» . Связь с биохимическими и биофизическими исследованиями . 430 (2): 700–705. дои : 10.1016/j.bbrc.2012.11.088 . ПМЦ 3579567 . ПМИД 23211596 .
- ^ Чжан Б., Цай П.С., Гонсалес-Селейро М., Чунг О., Бумар Б., Пердигото К.Н., Ежкова Е., Сюй Ю.К. (октябрь 2016 г.). «Транзитно-амплифицирующие клетки волосяных фолликулов управляют одновременным производством дермальных адипоцитов с помощью Sonic Hedgehog» . Гены и развитие . 30 (20): 2325–2338. дои : 10.1101/gad.285429.116 . ПМК 5110998 . ПМИД 27807033 .
- ^ Бамкрот Д.А., Такада Р., МакМахон А.П. (апрель 1995 г.). «Протеолитическая обработка дает две секретируемые формы звукового ежа» . Молекулярная и клеточная биология . 15 (4): 2294–2303. дои : 10.1128/MCB.15.4.2294 . ПМК 230457 . ПМИД 7891723 .
- ^ Ингхэм П.В., Накано Ю., Сегер С. (июнь 2011 г.). «Механизмы и функции передачи сигналов ежей через многоклеточные животные». Обзоры природы. Генетика . 12 (6): 393–406. дои : 10.1038/nrg2984 . ПМИД 21502959 . S2CID 33769324 .
- ^ Портер Дж. А., Янг К. Э., Бичи П. А. (октябрь 1996 г.). «Холестериновая модификация сигнальных белков ежей в развитии животных». Наука . 274 (5285): 255–259. Бибкод : 1996Sci...274..255P . дои : 10.1126/science.274.5285.255 . ПМИД 8824192 . S2CID 11125394 .
- ^ Пепинский Р.Б., Цзэн С., Вен Д., Рэйхорн П., Бейкер Д.П., Уильямс К.П., Бикслер С.А., Эмброуз С.М., Гарбер Э.А., Миатковски К., Тейлор Ф.Р., Ван Э.А., Галдес А. (май 1998 г.). «Идентификация модифицированной пальмитиновой кислотой формы человеческого ежа Соника» . Журнал биологической химии . 273 (22): 14037–14045. дои : 10.1074/jbc.273.22.14037 . ПМИД 9593755 . S2CID 22783982 .
- ^ Стэнтон Б.З., Пэн Л.Ф., Малуф Н., Накаи К., Ван X, Даффнер Дж.Л., Таверас К.М., Хайман Дж.М., Ли С.В., Келер А.Н., Чен Дж.К., Фокс Дж.Л., Мандинова А., Шрайбер С.Л. (март 2009 г.). «Небольшая молекула, которая связывает Hedgehog и блокирует его передачу сигналов в клетках человека» . Химическая биология природы . 5 (3): 154–156. дои : 10.1038/nchembio.142 . ПМК 2770933 . ПМИД 19151731 .
- ^ Маклин К. (январь 2006 г.). «Юмор названий генов, потерянных при переводе пациентам» . Природа . 439 (7074): 266. Бибкод : 2006Natur.439..266M . дои : 10.1038/439266d . ПМИД 16421543 . S2CID 19861153 .
- ^ Коэн М.М. (июль 2006 г.). «Проблемы именования генов». Американский журнал медицинской генетики. Часть А. 140 (13): 1483–1484. дои : 10.1002/ajmg.a.31264 . ПМИД 16718675 . S2CID 221388561 .
- ^ Белый М (26 сентября 2014 г.). «Sonic Hedgehog, DICER и проблема с именованием генов» . psmag.com . Тихоокеанский стандарт. Архивировано из оригинала 12 ноября 2020 года . Проверено 24 декабря 2020 г.
- ^ Хопкин М. (6 ноября 2006 г.). «Проблемные имена генов будут удалены» . Природа : news061106–2. дои : 10.1038/news061106-2 . S2CID 86514270 . Архивировано из оригинала 31 декабря 2020 года . Проверено 24 декабря 2020 г.
Дальнейшее чтение
[ редактировать ]- Дорус С., Андерсон Дж.Р., Валлендер Э.Дж., Гилберт С.Л., Чжан Л., Чемник Л.Г., Райдер О.А., Ли В., Лан Б.Т. (июль 2006 г.). «Sonic Hedgehog, ключевой ген развития, претерпел интенсивную молекулярную эволюцию у приматов» . Молекулярная генетика человека . 15 (13): 2031–2037. дои : 10.1093/hmg/ddl123 . ПМИД 16687440 .
- Гилберт С.Ф. (2000). Биология развития (6-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-243-6 .
- Ким Дж, Ким П, Хуэй CC (май 2001 г.). «Ассоциация VACTERL: уроки пути ежа Соника». Клиническая генетика . 59 (5): 306–315. дои : 10.1034/j.1399-0004.2001.590503.x . ПМИД 11359461 . S2CID 34304310 .
- Мортон Дж. П., Льюис BC (июль 2007 г.). «Тсс-сигнал и рак поджелудочной железы: значение для терапии?» . Клеточный цикл . 6 (13): 1553–1557. дои : 10.4161/cc.6.13.4467 . ПМИД 17611415 . S2CID 4670615 .
- Муллор Дж.Л., Санчес П., Руис и Альтаба А. (декабрь 2002 г.). «Пути и последствия: передача сигналов ежа при болезнях человека». Тенденции в клеточной биологии . 12 (12): 562–569. дои : 10.1016/S0962-8924(02)02405-4 . ПМИД 12495844 .
- Нанни Л., Мин Дж.Э., Ду Ю., Холл Р.К., Олдред М., Банкир А., Мюнке М. (июль 2001 г.). «Мутация SHH связана с одиночным срединным центральным резцом верхней челюсти: исследование 13 пациентов и обзор литературы». Американский журнал медицинской генетики . 102 (1): 1–10. doi : 10.1002/1096-8628(20010722)102:1<1::AID-AJMG1336>3.0.CO;2-U . ПМИД 11471164 .
- Уильямс Дж. А. (декабрь 2005 г.). «Еж и травма спинного мозга». Мнение экспертов о терапевтических целях . 9 (6): 1137–1145. дои : 10.1517/14728222.9.6.1137 . ПМИД 16300466 . S2CID 5548531 .
Внешние ссылки
[ редактировать ]- Вводная статья о SHH в колледже Дэвидсон.
- Открытие заново биологии: Блок 7 Генетика развития.. Стенограмма экспертного интервью, интервью с доктором философии Джоном Инкардоной .. объяснение открытия и названия гена звукового ежа
- «Sonic Hedgehog» поначалу звучало смешно .. New York Times, 12 ноября 2006 г..
- Запись GeneReviews/NCBI/NIH/UW об обзоре анофтальмии/микрофтальмии
- SHH – звуковой еж Национальная медицинская библиотека США
- Обзор всей структурной информации, доступной в PDB для UniProt : Q15465 (белок Hedgehog Human Sonic) на PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q62226 (белок Mouse Sonic hedgehog) в PDBe-KB .
галерея PDB |
|---|