Сигнальный путь ежа
| Белковый ежик | |||
|---|---|---|---|
| Идентификаторы | |||
| Организм | |||
| Символ | хх | ||
| ЮниПрот | Q02936 | ||
| |||
| Хеджирующий N-терминальный сигнал | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | HH_сигнал | ||
| Пфам | PF01085 | ||
| ИнтерПро | IPR000320 | ||
| КАТ | 3д1м | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 3д1м / ОБЛАСТЬ / СУПФАМ | ||
| |||
| См. также ИнтерПро : IPR001657 . | |||
— Сигнальный путь Hedgehog это сигнальный путь , который передает информацию эмбриональным клеткам, необходимую для правильной дифференцировки клеток . Различные части эмбриона имеют разные концентрации сигнальных белков ежа. Путь также играет роль у взрослых. Заболевания, связанные с нарушением работы этого пути, включают рак . [1] [2]
Сигнальный путь Hedgehog является одним из ключевых регуляторов развития животных и присутствует у всех билатерий . [3] Путь получил свое название от полипептидного лиганда , внутриклеточной сигнальной молекулы под названием Hedgehog ( Hh ), обнаруженной у плодовых мух рода Drosophila ; личинки плодовых мух, лишенные гена Hh Говорят, что , напоминают ежей . Hh дрозофилы — один из генных продуктов полярности сегментов , участвующий в создании основы строения тела мух . Молекула остается важной на более поздних стадиях эмбриогенеза и метаморфоза .
У млекопитающих есть три гомолога Hedgehog: Desert (DHH) , Indian (IHH) и Sonic (SHH) , из которых Sonic изучен лучше всего. Этот путь одинаково важен во время эмбрионального развития позвоночных и поэтому представляет интерес для эволюционной биологии развития . У нокаутных мышей, у которых отсутствуют компоненты этого пути, мозг , скелет , мускулатура , желудочно-кишечный тракт и легкие развиваются неправильно. Недавние исследования указывают на роль передачи сигналов Hedgehog в регуляции взрослых стволовых клеток, участвующих в поддержании и регенерации взрослых тканей . Этот путь также вовлечен в развитие некоторых видов рака . [1] Лекарства, специально нацеленные на передачу сигналов Hedgehog для борьбы с этим заболеванием, активно разрабатываются рядом фармацевтических компаний .
Открытие
[ редактировать ]
В 1970-х годах фундаментальной проблемой биологии развития было понимание того, как относительно простая яйцеклетка может дать начало сложному сегментированному строению тела. В конце 1970-х годов Кристиан Нюсляйн-Фольхард и Эрик Вишаус выделили мутации в генах, которые контролируют развитие сегментированной передне-задней оси тела мухи; [4] их метод «насыщающего мутагенеза» привел к открытию группы генов, участвующих в развитии сегментации тела , что помогло основать область эволюционной биологии развития . [5] В 1995 году они разделили Нобелевскую премию с Эдвардом Б. Льюисом за работу по изучению генетических мутаций в дрозофилы эмбриогенезе . [6]
Ген ежа дрозофилы ( hh ) был идентифицирован как один из нескольких генов, важных для создания различий между передней и задней частью отдельных сегментов тела. мухи Ген hh был независимо клонирован в 1992 году в лабораториях Джима Молера, Филипа Бичи , Томаса Б. Корнберга и Сайго Каору. Некоторые мутанты ежей приводят к образованию эмбрионов аномальной формы, которые необычно короткие и короткие по сравнению с эмбрионами дикого типа . Функция гена полярности сегмента ежа была изучена на предмет влияния на нормально поляризованное распределение кутикулярных зубцов личинок , а также на особенности придатков взрослых особей, таких как ноги и усики. [7] Вместо обычного рисунка зубчиков у личинок-мутантов ежей обычно наблюдаются «сплошные лужайки» из зубчиков (рис. 1). Внешний вид коротких и «волосатых» личинок послужил основанием для названия « еж ».
Плодовая мушка
[ редактировать ]

Механизм
[ редактировать ]Клетки насекомых экспрессируют полноразмерный «цинковые пальцы» транскрипционный фактор Cubitus прерывание (Ci), который образует комплекс с кинезин -подобным белком Costal-2 (Cos2) и локализуется в цитоплазме, связанной с клеточными микротрубочками (рис. 2). Комплекс SCF нацелен на полноразмерный белок Ci массой 155 кДа для протеосомно -зависимого расщепления, в результате которого образуется фрагмент массой 75 кДа (CiR). CiR накапливается в клетке и диффундирует в ядро , где действует как ко- репрессор для генов-мишеней Hedgehog ( Hh ). [8] Шаги, ведущие к протеолизу белка Ci, включают фосфорилирование белка Ci несколькими протеинкиназами ; PKA , GSK3β и CK1 (рис. 2). [9] Белок Slimb дрозофилы является частью комплекса SCF , который нацелен на белки для убиквитилирования . Slimm связывается с фосфорилированным белком Ci.
В отсутствие Hh (рис. 3) трансмембранный белок клеточной поверхности, называемый Patched (PTCH), предотвращает высокую экспрессию и активность 7-мембранного рецептора. [10] называется сглаженным (SMO). Patched имеет сходство последовательности с известными мембранными транспортными белками. Когда присутствует внеклеточный Hh (рис. 3), он связывается с Patched и ингибирует его, позволяя Smoothened накапливаться и ингибировать протеолитическое расщепление белка Ci. Этот процесс, скорее всего, включает прямое взаимодействие Smoothened и Costal-2 и может включать секвестрацию комплекса, содержащего белок Ci, в микродомен, где нарушаются этапы, ведущие к протеолизу белка Ci. [8] Механизм, с помощью которого связывание Hh с Patched приводит к повышению уровня Smoothened, не ясен (шаг 1 на рисунке 3). После привязки Hh к Patched уровни Smoothened значительно увеличиваются по сравнению с уровнем, поддерживаемым в ячейках, когда Patched не привязан к Hh. [11] Было высказано предположение, что фосфорилирование Smoothened играет роль в Hh-зависимой регуляции уровней Smoothened. [12]
В клетках с Hh-активированным Patched (рис. 3) интактный белок Ci накапливается в цитоплазме клетки, а уровни CiR снижаются, обеспечивая транскрипцию некоторых генов, таких как декапентаплегический (dpp, член семейства факторов роста BMP ). Для других генов, регулируемых Hh, экспрессия требует не только потери CiR, но и положительного действия нерасщепленного Ci, чтобы действовать как активатор транскрипции . [9] Costal-2 обычно важен для удержания белка Ci в цитоплазме, но взаимодействие Smoothened с Costal-2 позволяет некоторому количеству интактного белка Ci попасть в ядро. Белок Fused дрозофилы (Fu на рисунке 3) представляет собой протеинкиназу, которая связывается с Costal-2. Fused может ингибировать супрессор Fused (SUFU), который, в свою очередь, взаимодействует с Ci, регулируя транскрипцию генов в некоторых типах клеток. [13]
Роль
[ редактировать ]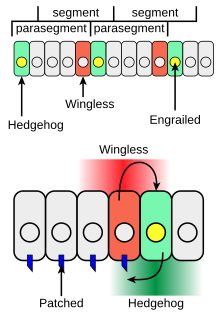
Еж играет роль в развитии сегментов тела личинок и формировании придатков взрослых особей. Во время формирования сегментов тела у развивающегося эмбриона дрозофилы полосы клеток, которые синтезируют транскрипционный фактор внедренный , также могут экспрессировать межклеточный сигнальный белок Hedgehog (зеленый на рисунке 4). Hedgehog не может уходить очень далеко от клеток, из которых он состоит, поэтому он активирует только тонкую полоску клеток, прилегающих к клеткам, экспрессирующим закреплённые клетки. Действуя таким образом, еж действует как паракринный фактор. Только клетки по одну сторону от клеток, экспрессирующих engrailed, способны реагировать на Hedgehog после взаимодействия Hh с рецепторным белком Patched (синий на рисунке 4).
Клетки с Hh-активированным рецептором Patched синтезируют белок Wingless (красный на рисунке 4). Если эмбрион дрозофилы изменен так, что он продуцирует Hh во всех клетках, все компетентные клетки реагируют и образуют более широкую полосу экспрессирующих Wingless клеток в каждом сегменте. Ген «бескрылого» имеет расположенную выше область регуляции транскрипции, которая связывает транскрипционный фактор Ci Hh-зависимым образом, что приводит к увеличению транскрипции «бескрылого» (взаимодействие 2 на рисунке 3) в полосе клеток, прилегающей к полосе клеток, продуцирующих Hh. [14]
Бескрылый белок действует как внеклеточный сигнал и формирует структуру соседних рядов клеток, активируя его рецептор на клеточной поверхности Frizzled . Wingless действует на клетки, экспрессирующие закреплённую экспрессию, стабилизируя полосы закреплённой экспрессии. Wingless является членом Wnt семейства белков, передающих сигнал от клетки к клетке, . Взаимная передача сигналов Hedgehog и Wingless стабилизирует границу между парасегментами (Figure 4, вверху). Эффекты Wingless и Hedgehog на другие полосы клеток в каждом сегменте устанавливают позиционный код, который объясняет различные анатомические особенности вдоль передне-задней оси сегментов. [15]
Белок Wingless называют «бескрылым» из-за фенотипа некоторых мутантов бескрылых мух. Бескрылый и Еж функционируют вместе во время метаморфоза , чтобы координировать формирование крыльев. Hedgehog экспрессируется в задней части развивающихся конечностей дрозофилы . Еж также участвует в координации развития глаз, мозга, половых желез, кишечника и трахеи. Подавление активности ежа было связано с замедлением развития глаз у амфипод Gammarus minus . [16]
Аннелиды
[ редактировать ]Hedgehog также участвует в сегментации кольчатых червей; поскольку параллельная эволюция кажется маловероятной, это предполагает общее происхождение сегментации между двумя типами. [17] Хотя Hh не вызывает образование сегментов, он, по-видимому, стабилизирует сегментированные поля после их появления. [17]
Позвоночные животные
[ редактировать ]Механизм
[ редактировать ]
Sonic hedgehog (SHH) — наиболее изученный лиганд пути позвоночных. Большая часть того, что известно о передаче сигналов hedgehog, была установлена в результате изучения SHH. Он транслируется как предшественник ~45 кДа и подвергается автокаталитическому сигнального домена ~20 кДа процессингу (процесс «1» на рисунке 5) с образованием N-концевого (называемого SHH-N) и C-концевого домена ~25 кДа, о котором неизвестно. сигнальная роль. В ходе расщепления холестерина . присоединяется молекула к карбоксильному концу N-концевого домена [18] который участвует в транспортировке, секреции и взаимодействии с рецепторами лиганда. SHH может передавать сигналы аутокринным способом , влияя на клетки, в которых он вырабатывается. Секреция и последующая паракринная передача сигналов hedgehog требуют участия белка Dispatched (DISP) (процесс «2» на рисунке 5).
Когда SHH достигает клетки-мишени, он связывается с рецептором Patched-1 (PTCH1) (процесс «3» на рисунке 5, синяя молекула). В отсутствие лиганда PTCH1 ингибирует Smoothened (SMO), нижестоящий белок этого пути (Процесс «4»). Было высказано предположение, что SMO регулируется небольшой молекулой, клеточная локализация которой контролируется PTCH. [19] PTCH1 имеет гомологию с Нимана-Пика болезнью типа C1 ( NPC1 ), которая, как известно, транспортирует липофильные молекулы через мембрану. [20] PTCH1 имеет стерол- чувствительный домен (SSD), который, как было показано, необходим для подавления активности SMO. [21] Современная теория предполагает, что PTCH регулирует SMO, удаляя оксистерины из SMO. PTCH действует как стероловый насос и удаляет оксистерины, вырабатываемые 7-дегидрохолестеринредуктазой . [22] При связывании белка Hh или мутации SSD PTCH насос выключается, позволяя оксистеринам накапливаться вокруг SMO.

Такое накопление стеринов позволяет СМО становиться активными или оставаться на мембране в течение более длительного периода времени. Эта гипотеза подтверждается существованием ряда низкомолекулярных агонистов и антагонистов пути, действующего на SMO. Связывание SHH снимает ингибирование SMO, что приводит к активации факторов транскрипции GLI (Процесс «5»): активаторов Gli1 и Gli2 и репрессора Gli3 . Последовательность молекулярных событий, связывающих SMO с GLI, плохо изучена. Активированный GLI накапливается в ядре (Процесс «6») и контролирует транскрипцию генов-мишеней ежа (Процесс «7»). Недавно сообщалось, что PTCH1 подавляет транскрипцию генов-мишеней hedgehog посредством механизма, независимого от Smoothened . [23]
Помимо PTCH1, у млекопитающих имеется еще один рецептор ежа, PTCH2, идентичность последовательности которого с PTCH1 составляет 54%. [24] Все три млекопитающих-ежа связывают оба рецептора с одинаковым сродством , поэтому PTCH1 и PTCH2 не могут различать лиганды. Однако они различаются по моделям выражения. PTCH2 экспрессируется на гораздо более высоких уровнях в семенниках и опосредует там передачу сигналов пустынного ежа. [24] По-видимому, он выполняет отличную от PTCH1 роль передачи сигналов ниже по ходу транскрипции. В отсутствие связывания лиганда PTCH2 имеет пониженную способность ингибировать активность SMO. [25] Более того, сверхэкспрессия PTCH2 не заменяет мутировавший PTCH1 при базальноклеточной карциноме . [26]
У беспозвоночных, как и у дрозофилы , связывание Hedgehog с PTCH приводит к интернализации и секвестрации лиганда. [27] Следовательно, in vivo прохождение ежа через рецептивное поле, экспрессирующее рецептор, приводит к ослаблению сигнала, эффекту, называемому лиганд-зависимым антагонизмом (LDA). В отличие от дрозофилы , позвоночные обладают другим уровнем регуляции hedgehog посредством LDA, опосредованного Hh-взаимодействующим белком 1 (HHIP1). HHIP1 также секвестрирует лиганды hedgehog, но в отличие от PTCH не влияет на активность SMO. [28]
Роль
[ редактировать ]
Члены семейства ежей играют ключевую роль в самых разных процессах развития. [15] Одним из наиболее изученных примеров является действие ежа Соника во время развития конечностей позвоночных. Классические эксперименты [29] Сондерса и Гасселинга в 1968 году. [30] на развитие зачатка конечностей кур легли в основу концепции морфогена . Они показали, что идентичность пальцев на конечности цыпленка определялась диффузионным фактором, создаваемым зоной поляризационной активности (ZPA), небольшим участком ткани на заднем крае конечности. Развитие млекопитающих, по-видимому, шло по той же схеме. Позже было показано, что этим диффузионным фактором является еж Соника . Однако до недавнего времени оставалось неясным, как именно SHH определяет идентичность цифр. Текущая модель, предложенная Harfe et al. , [31] утверждает, что как концентрация, так и время воздействия SHH определяют, в какой палец разовьется ткань мыши эмбриона (рисунок 6).
Цифры V, IV и часть III возникают непосредственно из клеток, экспрессирующих SHH во время эмбриогенеза . В этих клетках сигналы SHH аутокринны , и эти цифры развиваются правильно в отсутствие DISP, который необходим для внеклеточной диффузии лиганда. Эти цифры различаются в зависимости от продолжительности времени, в течение которого SHH продолжает выражаться. Самый задний палец V развивается из клеток, которые экспрессируют лиганд в течение наиболее длительного периода времени. Клетки пальца IV экспрессируют SHH в течение более короткого времени, а клетки пальца III еще короче. Цифра II развивается из клеток, подвергающихся воздействию умеренных концентраций внеклеточного SHH. Наконец, разработка Digit I не требует SHH. В каком-то смысле это программа по умолчанию для клеток зачатка конечностей.
Передача сигналов ежей остается важной и у взрослых. Было показано, что Sonic hedgehog способствует пролиферации взрослых стволовых клеток из различных тканей, включая примитивные гемопоэтические клетки, [32] молочная железа [33] и нейронный [34] стволовые клетки. Активация ежового пути необходима для перехода волосяного фолликула из фазы покоя в фазу роста. [35] Это не удалось из-за токсичности, обнаруженной на животных моделях. [36]
Болезнь человека
[ редактировать ]Нарушение передачи сигналов hedgehog во время эмбрионального развития либо из-за вредной мутации, либо из-за потребления тератогенов беременной матерью может привести к серьезным аномалиям развития. Голопрозэнцефалия , неспособность эмбрионального переднего мозга делиться с образованием полушарий головного мозга, встречается с частотой примерно 1 из 8000 живорождений и примерно 1 из 200 самопроизвольных абортов у людей и обычно связана с мутациями в генах, участвующих в пути ежа, в том числе ШХ и ПТЧ . [37] Циклопия , один из наиболее тяжелых дефектов голопрозэнцефалии ингибитора пути циклопамина беременными млекопитающими. , возникает в результате употребления [38]
Активация пути ежа вовлечена в развитие рака в различных органах, включая мозг , легкие , молочную железу , простату и кожу . Базальноклеточная карцинома , наиболее распространенная форма злокачественного новообразования , имеет наиболее тесную связь с передачей сигналов hedgehog. мутации потери функции в Patched и активирующие мутации в Smoothened . У пациентов с этим заболеванием были выявлены [39] Аномальная активация этого пути, вероятно, приводит к развитию заболевания за счет трансформации взрослых стволовых клеток в раковые стволовые клетки , которые дают начало опухоли. Исследователи рака надеются, что специфические ингибиторы передачи сигналов «ежик» обеспечат эффективную терапию широкого спектра злокачественных новообразований. [40] Связь между сигнальным путем hedgehog и развитием рака очень сложна. Тем не менее, ясно, что аберрантная активация передачи сигналов hedgehog приводит к росту, пролиферации и инвазии опухолевых клеток. [41] Помимо участия в развитии рака , путь ежа может также способствовать развитию серьезных респираторных заболеваний, таких как легочный фиброз . [42] и хроническая обструктивная болезнь легких . [43] [44] [45]
Нацеливание на путь ежа
[ редактировать ]Самый распространенный способ воздействовать на этот путь — модулировать SMO. Уже было показано, что антагонист и агонист SMO влияют на регуляцию нижележащего пути. Для лечения рака доступны несколько ингибиторов сигнального пути hedgehog, такие как висмодегиб и сонидегиб. Эти препараты считаются многообещающими методами лечения рака, особенно для пациентов с рефрактерным/распространенным раком. Ингибиторы SMO представляют собой потенциальное лечение некоторых типов рака. Однако из-за вредных и потенциально токсичных побочных эффектов ингибиторов SMO, неустановленной безопасности для детей и данных о том, что у некоторых пациентов развивается резистентность к ингибиторам SMO, необходимы новые классы лекарств. [41] Наиболее клинически продвинутые агенты, нацеленные на SMO, конкурентоспособны по отношению к циклопамину . Также было показано, что итраконазол ( Споранокс ) воздействует на СМО посредством механизма, отличного от циклопамина и висмодегиба . [46] Итраконазол (ITZ) ингибирует SMO при наличии мутаций, обеспечивающих устойчивость к висмодегибу и другим циклопамин -конкурентным антагонистам, таким как IPI-926 и LDE-225 Novartis. [47] ПТЧ [48] и Gli3 (5E1) [49] антитела также являются способом регулирования этого пути. Нижестоящий эффектор и сильный активатор транскрипции siRNA Gli1 использовался для ингибирования роста клеток и стимулирования апоптоза. [50] Также было показано, что триоксид мышьяка ( Трисенокс ) ингибирует передачу сигналов hedgehog, вмешиваясь в функцию Gli и транскрипцию. [51] [52]
Было идентифицировано несколько модификаторов окружающей среды передачи сигналов Hedgehog, которые представляют потенциальную опасность для здоровья или развития. Диетические алкалоиды, содержащиеся в томатах (томатодин), [53] картофель (соланидин), [53] пасленовые, такие как перец и баклажаны (соласодин). [53] и куркума (куркумин) [54] Было показано, что они противодействуют SMO и нарушают передачу сигналов Hedgehog. Кроме того, некоторые токсиканты окружающей среды могут блокировать передачу сигналов Hedgehog. Пиперонилбутоксид (ПБО) — это полусинтетическая пестицидная добавка, разработанная в 1940-х годах, которую можно найти в тысячах бытовых и сельскохозяйственных продуктов. [55] Несмотря на широкое использование, способность PBO ингибировать передачу сигналов hedgehog и действовать как мощный тератоген для развития не была признана до недавнего времени. [56] [57]
Метастазирование
[ редактировать ]Активация пути Hedgehog приводит к увеличению экспрессии белка улитки и уменьшению количества E-кадгерина и плотных контактов . [58]
Регуляция опухоли
[ редактировать ]Активация пути Hedgehog приводит к увеличению ангиогенных факторов (ангиопоэтина-1 и ангиопоэтина-2), [59] Циклины (циклин D1 и B1), [60] антиапоптотических генов и снижение апоптотических генов (Fas). [61] Дисфункция или аберрантная активация сигнального пути Hh связана с пороками развития и раком, включая синдром базальноклеточного невуса (BCNS), базальноклеточную карциному (BCC), медуллобластомы (MB), рабдомиосаркомы и менингиомы. Примерно треть злокачественных опухолей связана с аберрантной активацией пути Hh. Существует три предполагаемых механизма аберрантной активации передачи сигналов Hh при различных типах рака: тип I включает лиганд-независимую передачу сигналов из-за мутаций в Smo или отрицательных регуляторах, тип II включает лиганд-зависимую аутокринную/юктакринную передачу сигналов со сверхэкспрессией лиганда Hh и тип III. включает лиганд-зависимую паракринную передачу сигналов между опухолевыми клетками и стромальными клетками. Эти нарушения регуляции пути Hh могут привести к пролиферации, выживанию опухолевых клеток и присутствию раковых стволовых клеток, которые способствуют инициированию и прогрессированию опухоли. [62]
Клинические испытания
[ редактировать ]- Самый Модегиб [63] Одобрено FDA (январь 2012 г.) для лечения базальноклеточной карциномы .
- Сонидегиб одобрен FDA (июль 2015 г.) для лечения базальноклеточного рака .
- Итраконазол [64]
Эволюция
[ редактировать ]
Ланцетники , являющиеся примитивными хордовыми , обладают только одним гомологом дрозофилы Hh (рисунок 7). С другой стороны, у позвоночных есть несколько лигандов Hedgehog, которые попадают в три подгруппы — Desert , Indian и Sonic , каждая из которых представлена одним геном млекопитающих. Это следствие двух раундов дупликации всего генома, произошедших на ранних этапах истории эволюции позвоночных. [65] Два таких события дали бы четыре гомологичных гена, один из которых, должно быть, был утерян. Пустынные ежи являются наиболее близкими родственниками дрозофилы Hh . У некоторых видов произошли дополнительные дупликации генов. [15] такие как рыбка данио Danio rerio , у которой есть дополнительный ген tiggywinkle hedgehog в звуковой группе. Различные линии позвоночных адаптировали ежей к уникальным процессам развития. Например, гомолог X.laevis полосатого ежа участвует в регенерации конечности саламандры . [66]
shh претерпел ускоренную эволюцию в линии приматов, ведущей к человеку. [67] Дорус и др. выдвинули гипотезу, что это позволило осуществить более сложную регуляцию белка и, возможно, сыграло роль в увеличении объема и сложности человеческого мозга.
Семейство рецепторов WNT Frizzled имеет некоторое сходство последовательности с Smoothened . [68] Smoothened, по-видимому, является функционально отличающимся членом суперсемейства рецепторов, связанных с G-белком (GPCR). Были рассмотрены другие сходства между сигнальными путями WNT и Hh. [69] Нусс заметил, что «сигнальная система, основанная на модифицированных липидами белках и специфических мембранных транслокаторах, является древней и, возможно, была основателем сигнальных систем Wnt и Hh».
Было высказано предположение, что передача сигналов у беспозвоночных и позвоночных ниже по течению от Smoothened значительно различается. [70] Роль Suppressor of Fused (SUFU) повышена у позвоночных по сравнению с дрозофилой , где его роль относительно незначительна. Costal-2 особенно важен для дрозофилы . Протеинкиназа Fused является регулятором SUFU у дрозофилы , но может не играть роли в пути Hh у позвоночных. [71] У позвоночных передача сигналов Hh сильно вовлечена в развитие ресничек . [72]
Поразительная эволюция уровня домена присутствует в семействе белков Hedgehog, N-концевом домене (Hedge) и C-концевом домене (Hog), которые позже были сращены вместе в единую транскрипционную единицу. [73] [74] Домен Hog содержит последовательность под названием Hint (Hedgehog INTein), которая по последовательности и функциям аналогична бактериальным и грибковым интеинам . [75] Домен Hog присутствует во многих ветвях эукариот, т.е. в красных водорослях, мхах, динофлагеллятах, якобидах и других одноклеточных эукариотах. [76] Хоанофлагелляты содержат ген hoglet, который также кодирует C-концевой домен ежа Hog. Однако хоанофлагелляты и низшие эукариоты не содержат каких-либо областей, подобных хедж-домену, что позволяет предположить, что свиньи эволюционировали первыми. [75] [76] Poriferans имеют как хедж-подобные белки (называемые хеджинговыми), так и свиноподобные белки, но они существуют как две совершенно отдельные транскрипционные единицы. [73] [74] Книдарии содержат гены хеджинга и свиньи, но также имеют полный ген ежа, что указывает на то, что хедж и свинья были сращены в ежа после последнего общего предка пориферан и книдарий. [74]
Билатерии не содержат генов изгороди, что позволяет предположить, что они были потеряны в результате делеции до того, как эта ветвь отделилась от других многоклеточных животных. [3] Однако гены, содержащие домен Hog без домена Hedge, присутствуют в нескольких билатеральных линиях. Они обнаружены у Lophotrochozoa и Nematoda . [77] Гены, подобные Hedgehog, 2 гомолога Patched и гены, родственные Patched, существуют у червя C. elegans . [78] [79] Было показано, что эти гены кодируют белки, которые играют роль в развитии C. elegans . [78] [79] В то время как нематоды Enoplea сохранили настоящего Hedgehog, Chromadoreans потеряли архетипического Hedgehog и вместо этого развили расширенный репертуар из 61 дивергентного полуортологичного гена с новыми N-концевыми доменами, связанными с Hog. [76] [77] Эти N-концевые домены, связанные с Hog у C. elegans, были впоследствии классифицированы: сначала Warthog (WRT) и Groundhog (GRD), а затем Ground-like (GRL) и Quahog (QUA). [78] [79] C. elegans , как и другие виды нематод, утратили GPCR Smoothened. [78] [79]
Предполагается, что основным источником сигнального пути Hedgehog является бактериальный путь регуляции гопаноидов , которые являются общими липидными компонентами бактерий и являются структурными аналогами стероидов . [80]
См. также
[ редактировать ]- Sonic hedgehog , наиболее изученный лиганд пути позвоночных.
- Сглаженный , консервативный GPCR . компонент пути
- Netpath - тщательно подобранный ресурс о путях передачи сигналов у человека.
- Ингибиторы передачи сигналов Hh
- Циклопамин — небольшая молекула природного происхождения.
- Сонидегиб
- Висмодегиб одобрен для лечения базальноклеточного рака .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Джеймисон С., Мартинелли Дж., Папаяннидис С., Кортес Дж.Э. (сентябрь 2020 г.). «Ингибиторы пути ежа: новый терапевтический класс для лечения острого миелолейкоза» . Открытие рака крови . 1 (2): 134–145. дои : 10.1158/2643-3230.BCD-20-0007 . ПМЦ 8447269 . ПМИД 34661144 .
- ^ Кимбалл Дж.В. (18 февраля 2008 г.). «Сигнальный путь ежа» . Страницы биологии Кимбалла . Архивировано из оригинала 26 июня 2008 года.
- ^ Перейти обратно: а б Ингхэм П.В., Накано Ю., Сегер С. (июнь 2011 г.). «Механизмы и функции передачи сигналов ежей через многоклеточные животные». Обзоры природы. Генетика . 12 (6): 393–406. дои : 10.1038/nrg2984 . ПМИД 21502959 . S2CID 33769324 .
- ^ Нюсляйн-Фольхард С., Вишаус Е (октябрь 1980 г.). «Мутации, влияющие на количество и полярность сегментов у дрозофилы». Природа . 287 (5785): 795–801. Бибкод : 1980Natur.287..795N . дои : 10.1038/287795a0 . ПМИД 6776413 . S2CID 4337658 .
- ^ Артур В. (февраль 2002 г.). «Новые концептуальные рамки эволюционной биологии развития». Природа . 415 (6873): 757–764. Бибкод : 2002Natur.415..757A . дои : 10.1038/415757a . ПМИД 11845200 . S2CID 4432164 .
- ^ Нобелевская премия 1995 г. за открытие генетического контроля раннего эмбрионального развития.
- ^ Молер Дж. (декабрь 1988 г.). «Требования к гену сегментной полярности hedgehog для формирования рисунка кутикулы личинки и взрослой особи дрозофилы» . Генетика . 120 (4): 1061–1072. дои : 10.1093/генетика/120.4.1061 . ПМЦ 1203569 . ПМИД 3147217 .
- ^ Перейти обратно: а б Коллинз RT, Коэн С.М. (май 2005 г.). «Генетический скрининг дрозофилы для выявления новых компонентов сигнального пути ежа» . Генетика . 170 (1): 173–184. дои : 10.1534/genetics.104.039420 . ПМЦ 1449730 . ПМИД 15744048 .
- ^ Перейти обратно: а б Лам Л., Бичи, Пенсильвания (июнь 2004 г.). «Сеть реагирования Hedgehog: датчики, коммутаторы и маршрутизаторы». Наука . 304 (5678): 1755–1759. Бибкод : 2004Sci...304.1755L . CiteSeerX 10.1.1.476.3902 . дои : 10.1126/science.1098020 . ПМИД 15205520 . S2CID 13949436 .
- ^ Чен В., Рен XR, Нельсон К.Д., Барак Л.С., Чен Дж.К., Бичи П.А. и др. (декабрь 2004 г.). «Зависимая от активности интернализация сглаживания, опосредованная бета-аррестином 2 и GRK2». Наука . 306 (5705): 2257–2260. Бибкод : 2004Sci...306.2257C . дои : 10.1126/science.1104135 . ПМИД 15618519 . S2CID 12823611 .
- ^ Альседо Дж., Зоу Ю., Нолл М. (август 2000 г.). «Посттранскрипционная регуляция сглаживания является частью механизма самокоррекции в сигнальной системе Hedgehog» . Молекулярная клетка . 6 (2): 457–465. дои : 10.1016/S1097-2765(00)00044-7 . ПМИД 10983991 .
- ^ Апионишев С., Катанаева Н.М., Маркс С.А., Кальдерон Д., Томлинсон А. (январь 2005 г.). «Сглаженные сайты фосфорилирования дрозофилы, необходимые для передачи сигнала Hedgehog». Природная клеточная биология . 7 (1): 86–92. дои : 10.1038/ncb1210 . ПМИД 15592457 . S2CID 5958856 .
- ^ Хо К.С., Суяма К., Фиш М., Скотт М.П. (март 2005 г.). «Дифференциальная регуляция транскрипции гена-мишени Hedgehog с помощью Costal2 и супрессора Fused» . Разработка . 132 (6): 1401–1412. дои : 10.1242/dev.01689 . ПМИД 15750186 .
- ^ Фон Олен Т., Лессинг Д., Нусс Р., Хупер Дж.Э. (март 1997 г.). «Передача сигналов Hedgehog регулирует транскрипцию посредством Cubitus Interruptus, белка, специфичного для последовательности ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (6): 2404–2409. Бибкод : 1997PNAS...94.2404V . дои : 10.1073/pnas.94.6.2404 . ПМК 20100 . ПМИД 9122207 .
- ^ Перейти обратно: а б с Ингхэм П.В., МакМахон А.П. (декабрь 2001 г.). «Передача сигналов ежа в развитии животных: парадигмы и принципы» . Гены и развитие . 15 (23): 3059–3087. дои : 10.1101/gad.938601 . ПМИД 11731473 .
- ^ Аспирас AC, Прасад Р., Фонг Д.В., Карлини Д.Б., Анджелини Д.Р. (май 2012 г.). «Параллельное снижение экспрессии гена развития глаз hedgehog в отдельно выведенных пещерных популяциях амфипод Gammarus minus». Журнал эволюционной биологии . 25 (5): 995–1001. дои : 10.1111/j.1420-9101.2012.02481.x . ПМИД 22462461 . S2CID 37541721 .
- ^ Перейти обратно: а б Дрей Н., Тессмар-Райбл К., Ле Гуар М., Виберт Л., Христодулу Ф., Шипани К. и др. (июль 2010 г.). «Передача сигналов Hedgehog регулирует формирование сегментов у кольчатых червей Platynereis» . Наука . 329 (5989): 339–342. Бибкод : 2010Sci...329..339D . дои : 10.1126/science.1188913 . ПМК 3182550 . ПМИД 20647470 .
- ^ Банавали Н.К. (март 2020 г.). «Механизм модификации холестерина лиганда Hedgehog» . Журнал вычислительной химии . 41 (6): 520–527. дои : 10.1002/jcc.26097 . ПМИД 31823413 .
- ^ Тайпале Дж., Купер М.К., Маити Т., Бичи П.А. (август 2002 г.). «Patched действует каталитически, подавляя активность Smoothened». Природа . 418 (6900): 892–897. Бибкод : 2002Natur.418..892T . дои : 10.1038/nature00989 . ПМИД 12192414 . S2CID 4362029 .
- ^ Дэвис Дж.П., Чен Ф.В., Иоанну Ю.А. (декабрь 2000 г.). «Активность трансмембранного молекулярного насоса белка C1 Нимана-Пика». Наука . 290 (5500): 2295–2298. Бибкод : 2000Sci...290.2295D . дои : 10.1126/science.290.5500.2295 . ПМИД 11125140 .
- ^ Стратт Х., Томас С., Накано Ю., Старк Д., Нив Б., Тейлор А.М., Ингхэм П.В. (апрель 2001 г.). «Мутации в домене Patched, чувствительном к стеролу, предполагают роль везикулярного транспорта в регуляции Smoothened» . Современная биология . 11 (8): 608–613. дои : 10.1016/S0960-9822(01)00179-8 . ПМИД 11369206 . S2CID 235311937 .
- ^ Коркоран Р.Б., член парламента Скотта (май 2006 г.). «Оксистерины стимулируют передачу сигнала Sonic hedgehog и пролиферацию клеток медуллобластомы» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (22): 8408–8413. Бибкод : 2006PNAS..103.8408C . дои : 10.1073/pnas.0602852103 . ПМК 1462959 . ПМИД 16707575 .
- ^ Рахнама Ф., Симокава Т., Лаут М., Финта С., Когерман П., Теглунд С. и др. (февраль 2006 г.). «Ингибирование активации гена GLI1 с помощью Patched1» . Биохимический журнал . 394 (Часть 1): 19–26. дои : 10.1042/BJ20050941 . ПМК 1385998 . ПМИД 16229683 .
- ^ Перейти обратно: а б Карпентер Д., Стоун Д.М., Браш Дж., Райан А., Арманини М., Франц Г. и др. (ноябрь 1998 г.). «Характеристика двух исправленных рецепторов семейства белков ежей позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (23): 13630–13634. Бибкод : 1998PNAS...9513630C . дои : 10.1073/pnas.95.23.13630 . ПМК 24870 . ПМИД 9811851 .
- ^ Рахнама Ф., Тофтгард Р., Зафиропулос П.Г. (март 2004 г.). «Отличные роли вариантов сплайсинга PTCH2 в передаче сигналов Hedgehog» . Биохимический журнал . 378 (Часть 2): 325–334. дои : 10.1042/BJ20031200 . ПМЦ 1223965 . ПМИД 14613484 .
- ^ Зафиропулос П.Г., Унден А.Б., Рахнама Ф., Холлингсворт Р.Э., Тофтгард Р. (февраль 1999 г.). «PTCH2, новый исправленный ген человека, подвергающийся альтернативному сплайсингу и повышающий регуляцию при базальноклеточной карциноме». Исследования рака . 59 (4): 787–792. ПМИД 10029063 .
- ^ Инкардона Дж. П., Ли Дж. Х., Робертсон С. П., Энга К., Капур Р. П., Ролинк Х. (октябрь 2000 г.). «Рецепторно-опосредованный эндоцитоз растворимого и мембраносвязанного ежа Sonic с помощью Patched-1» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (22): 12044–12049. Бибкод : 2000PNAS...9712044I . дои : 10.1073/pnas.220251997 . ПМК 17291 . ПМИД 11027307 .
- ^ Чон Дж., МакМахон AP (январь 2005 г.). «Рост и структура нервной трубки млекопитающих регулируются частично перекрывающимися действиями обратной связи антагонистов hedgehog patched 1 и Hhip1» . Разработка . 132 (1): 143–154. дои : 10.1242/dev.01566 . ПМИД 15576403 .
- ^ Сондерс Дж.В. (август 1948 г.). «Проксимально-дистальная последовательность происхождения частей куриного крыла и роль эктодермы». Журнал экспериментальной зоологии . 108 (3): 363–403. Бибкод : 1948JEZ...108..363S . дои : 10.1002/jez.1401080304 . ПМИД 18882505 .
- ^ Сондерс Дж.В., Гасселинг М.Т. (1968). «Эктодермально-мезенхимальные взаимодействия в возникновении симметрии конечностей». В Флейшмайере Р., Биллингеме Р.Э. (ред.). Эпителиально-мезенхимальные взаимодействия . Балтимор: Уильямс и Уилкинс. стр. 78–97.
- ^ Харф Б.Д., Шерц П.Дж., Ниссим С., Тиан Х., МакМахон А.П., Табин С.Дж. (август 2004 г.). «Доказательства временного градиента Shh, основанного на расширении, при определении идентичности пальцев позвоночных» . Клетка . 118 (4): 517–528. дои : 10.1016/j.cell.2004.07.024 . ПМИД 15315763 . S2CID 16280983 .
- ^ Бхардвадж Г., Мердок Б., Ву Д., Бейкер Д.П., Уильямс К.П., Чедвик К. и др. (февраль 2001 г.). «Sonic hedgehog индуцирует пролиферацию примитивных гемопоэтических клеток человека посредством регуляции BMP». Природная иммунология . 2 (2): 172–180. дои : 10.1038/84282 . ПМИД 11175816 . S2CID 27907370 .
- ^ Лю С., Донту Г., Мантл И.Д., Патель С., Ан Н.С., Джексон К.В. и др. (июнь 2006 г.). «Передача сигналов Hedgehog и Bmi-1 регулируют самообновление нормальных и злокачественных стволовых клеток молочной железы человека» . Исследования рака . 66 (12): 6063–6071. дои : 10.1158/0008-5472.CAN-06-0054 . ПМЦ 4386278 . ПМИД 16778178 .
- ^ Ан С., Джойнер А.Л. (октябрь 2005 г.). «Анализ in vivo покоящихся взрослых нервных стволовых клеток, реагирующих на Sonic hedgehog». Природа . 437 (7060): 894–897. Бибкод : 2005Natur.437..894A . дои : 10.1038/nature03994 . ПМИД 16208373 . S2CID 4431022 .
- ^ Паладини Р.Д., Салех Дж., Цянь С., Сюй Г.С., Рубин Л.Л. (октябрь 2005 г.). «Модуляция роста волос с помощью низкомолекулярных агонистов сигнального пути ежа» . Журнал исследовательской дерматологии . 125 (4): 638–646. дои : 10.1111/j.0022-202X.2005.23867.x . ПМИД 16185261 .
- ^ http://phx.corporate-ir.net/phoenix.zhtml?c=123198&p=irol-newsArticle&ID=997941&highlight= [ нужна полная цитата ]
- ^ Интернет-менделевское наследование у человека (OMIM): голопрозэнцефалия - 236100
- ^ Килер РФ (октябрь 1978 г.). «Циклопамин и родственные стероидные алкалоиды тератогены: их возникновение, структурные взаимоотношения и биологические эффекты». Липиды . 13 (10): 708–715. дои : 10.1007/BF02533750 . ПМИД 723484 . S2CID 1119899 .
- ^ Се Дж., Муроне М., Луох С.М., Райан А., Гу Q, Чжан С. и др. (январь 1998 г.). «Активация сглаженных мутаций при спорадической базальноклеточной карциноме». Природа . 391 (6662): 90–92. Бибкод : 1998Natur.391...90X . дои : 10.1038/34201 . ПМИД 9422511 . S2CID 205003240 .
- ^ Чен Дж. К., Тайпале Дж., Янг К. Э., Маити Т., Бичи П. А. (октябрь 2002 г.). «Маломолекулярная модуляция активности Smoothened» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (22): 14071–14076. Бибкод : 2002PNAS...9914071C . дои : 10.1073/pnas.182542899 . ПМЦ 137838 . ПМИД 12391318 .
- ^ Перейти обратно: а б Шкода А.М., Симович Д., Карин В., Кардум В., Вранич С., Серман Л. (февраль 2018 г.). «Роль сигнального пути Hedgehog при раке: всесторонний обзор» . Боснийский журнал фундаментальных медицинских наук . 18 (1): 8–20. дои : 10.17305/bjbms.2018.2756 . ПМК 5826678 . ПМИД 29274272 .
- ^ Сигна Н., Фаррохи Мошаи Э., Брайер С., Маршаль-Сомма Дж., Вемо-Стервину Л., Фабр А. и др. (декабрь 2012 г.). «Механизм системы ежа контролирует трансформирующий фактор роста-β-зависимую дифференцировку миофибробластов у людей: участие в идиопатическом легочном фиброзе» . Американский журнал патологии . 181 (6): 2126–2137. дои : 10.1016/j.ajpath.2012.08.019 . ПМИД 23031257 .
- ^ Ван С., де Мохель Н.С., Кристенсон С.А., Кассандра М., Мун Р., Брамвелл А.Н. и др. (октябрь 2018 г.). «Распространение ежа нарушает мезенхимальную идентичность и вызывает фенотип эмфиземы» . Журнал клинических исследований . 128 (10): 4343–4358. дои : 10.1172/JCI99435 . ПМК 6159975 . ПМИД 29999500 .
- ^ Ансель Дж., Бельгасеми Р., Перотин Дж.М., Диабасана З., Дьюри С., Девольф М. и др. (август 2020 г.). «Передача звукового ежа как потенциальный эндобронхиальный биомаркер при ХОБЛ» . Респираторные исследования . 21 (1): 207. дои : 10.1186/s12931-020-01478-x . ПМЦ 7412648 . ПМИД 32767976 .
- ^ Бельгачеми Р., Лучка Э., Ансель Дж., Диабасана З., Перотин Дж.М., Жермен А. и др. (январь 2020 г.). «Дифференцировка эпителиальных клеток дыхательных путей зависит от недостаточной передачи сигналов Hedgehog при ХОБЛ» . Электронная биомедицина . 51 : 102572. doi : 10.1016/j.ebiom.2019.11.033 . ПМК 6931110 . ПМИД 31877414 .
- ^ Ким Дж., Тан Дж.И., Гонг Р., Ким Дж., Ли Дж.Дж., Клемонс К.В. и др. (апрель 2010 г.). «Итраконазол, широко используемый противогрибковый препарат, который подавляет активность пути Hedgehog и рост рака» . Раковая клетка . 17 (4): 388–399. дои : 10.1016/j.ccr.2010.02.027 . ПМК 4039177 . ПМИД 20385363 .
- ^ Ким Дж., Афтаб Б.Т., Тан Дж.И., Ким Д., Ли А.Х., Резаи М. и др. (январь 2013 г.). «Итраконазол и триоксид мышьяка ингибируют активацию пути Hedgehog и рост опухоли, связанный с приобретенной устойчивостью к сглаженным антагонистам» . Раковая клетка . 23 (1): 23–34. дои : 10.1016/j.ccr.2012.11.017 . ПМЦ 3548977 . ПМИД 23291299 .
- ^ Накамура М., Кубо М., Янаи К., Миками Ю., Икебе М., Нагай С. и др. (2007). «Антитела против патч-1 подавляют сигнальный путь ежа и пролиферацию рака поджелудочной железы». Противораковые исследования . 27 (6А): 3743–3747. ПМИД 17970037 .
- ^ Хант Р., Брагина О., Дрюс М., Касак Л., Тиммуск С., Валкна А. и др. (август 2007 г.). «Получение и характеристика мышиных моноклональных антител 5E1 против фактора транскрипции человека GLI3». Гибридома . 26 (4): 231–240. дои : 10.1089/hyb.2007.0507 . ПМИД 17725385 .
- ^ Стекка Б., Мас С., Руис и Альтаба А. (май 2005 г.). «Вмешательство в передачу сигналов HH-GLI ингибирует рак простаты» . Тенденции молекулярной медицины . 11 (5): 199–203. doi : 10.1016/j.molmed.2005.03.004 . ПМИД 15882606 .
- ^ Бошан Э.М., Рингер Л., Булут Г., Саджван К.П., Холл М.Д., Ли Ю.К. и др. (январь 2011 г.). «Триоксид мышьяка подавляет рост раковых клеток человека и развитие опухолей у мышей, блокируя путь Hedgehog/GLI» . Журнал клинических исследований . 121 (1): 148–160. дои : 10.1172/JCI42874 . ПМК 3007144 . ПМИД 21183792 .
- ^ Ким Дж., Ли Джей-Джей, Ким Дж., Гарднер Д., Бичи П.А. (июль 2010 г.). «Мышьяк противодействует пути Hedgehog, предотвращая накопление ресничек и снижая стабильность транскрипционного эффектора Gli2» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (30): 13432–13437. Бибкод : 2010PNAS..10713432K . дои : 10.1073/pnas.1006822107 . ПМЦ 2922148 . ПМИД 20624968 .
- ^ Перейти обратно: а б с Липински Р.Дж., Бушман В. (август 2010 г.). «Идентификация ингибиторов передачи сигналов Hedgehog, имеющих значимое воздействие на человека, путем скрининга малых молекул» . Токсикология in vitro . 24 (5). Токсикол in vitro: 1404–1409. дои : 10.1016/j.tiv.2010.04.011 . ПМК 2891024 . ПМИД 20434536 .
- ^ Эламин М.Х., Шинвари З., Хендрайани С.Ф., Аль-Хинди Х., Аль-Шаил Э., Хафага Ю. и др. (март 2010 г.). «Куркумин ингибирует сигнальный путь Sonic Hedgehog и запускает апоптоз в клетках медуллобластомы». Молекулярный канцерогенез . 49 (3): 302–314. дои : 10.1002/mc.20604 . ПМИД 20025076 . S2CID 31459213 .
- ^ «Решение о перерегистрации пиперонилбутоксида» (PDF) . УСЕПА .
- ^ Ван Дж., Лу Дж., Мук Р.А., Чжан М., Чжао С., Барак Л.С. и др. (август 2012 г.). «Синергист инсектицидов пиперонилбутоксид ингибирует передачу сигналов ежей: оценка химических рисков» . Токсикологические науки . 128 (2): 517–523. doi : 10.1093/toxsci/kfs165 . ПМЦ 3493191 . ПМИД 22552772 .
- ^ Эверсон Дж.Л., Сан М.Р., Финк Д.М., Хейн Г.В., Мельберг К.Г., Нельсон К.Ф. и др. (октябрь 2019 г.). «Оценка токсичности воздействия пиперонилбутоксида на развитие, направленная на передачу сигналов Sonic Hedgehog и морфогенез переднего мозга и лица у мышей: исследование in vitro и in vivo » . Перспективы гигиены окружающей среды . 127 (10): 107006. doi : 10.1289/EHP5260 . ПМК 6867268 . ПМИД 31642701 .
- ^ Ли X, Дэн В., Nail CD, Бейли С.К., Краус М.Х., Рупперт Дж.М., Лобо-Рупперт С.М. (январь 2006 г.). «Индукция улитки — это ранний ответ на Gli1, который определяет эффективность эпителиальной трансформации» . Онкоген . 25 (4): 609–621. дои : 10.1038/sj.onc.1209077 . ПМЦ 1361531 . ПМИД 16158046 .
- ^ Ли С.В., Московиц М.А., Симс-младший (март 2007 г.). «Sonic hedgehog обратно регулирует экспрессию ангиопоэтина-1 и ангиопоэтина-2 в фибробластах» . Международный журнал молекулярной медицины . 19 (3): 445–451. дои : 10.3892/ijmm.19.3.445 . ПМИД 17273793 .
- ^ Адольф С., Хетерингтон Р., Эллис Т., Уэйнрайт Б. (февраль 2006 г.). «Patched1 действует как привратник, способствуя развитию клеточного цикла» . Исследования рака . 66 (4): 2081–2088. дои : 10.1158/0008-5472.CAN-05-2146 . ПМИД 16489008 .
- ^ Атар М., Ли С., Тан Х., Чи С., Чжан Х., Ким А.Л. и др. (октябрь 2004 г.). «Ингибирование сглаженной передачи сигналов предотвращает базальноклеточную карциному, индуцированную ультрафиолетом B, посредством регуляции экспрессии Fas и апоптоза» . Исследования рака . 64 (20): 7545–7552. дои : 10.1158/0008-5472.CAN-04-1393 . ПМИД 15492281 .
- ^ Шкода А.М., Симович Д., Карин В., Кардум В., Вранич С., Серман Л. (февраль 2018 г.). «Роль сигнального пути Hedgehog при раке: всесторонний обзор» . Боснийский журнал фундаментальных медицинских наук . 18 (1): 8–20. дои : 10.17305/bjbms.2018.2756 . ПМК 5826678 . ПМИД 29274272 .
- ^ Номер клинического исследования NCT00636610 «Исследование GDC-0449 (ингибитор пути Hedgehog) с одновременной химиотерапией и бевацизумабом в качестве терапии первой линии при метастатическом колоректальном раке» на ClinicalTrials.gov
- ^ Номер клинического исследования NCT01108094 «Пилотное исследование биомаркеров для оценки эффективности итраконазола у пациентов с базальноклеточной карциномой» на сайте ClinicalTrials.gov.
- ^ Вада Х., Макабе К. (2006). «Дупликация генома ранних позвоночных как возможная хроника эволюционной истории нервного гребня» . Международный журнал биологических наук . 2 (3): 133–141. дои : 10.7150/ijbs.2.133 . ПМЦ 1474148 . ПМИД 16763673 .
- ^ Старк Д.Р., Гейтс П.Б., Брокс Дж.П., Ферретти П. (июль 1998 г.). «Член семейства ежей проявляется во всех регенерирующих и развивающихся конечностях» . Динамика развития . 212 (3): 352–363. doi : 10.1002/(SICI)1097-0177(199807)212:3<352::AID-AJA3>3.0.CO;2-G . ПМИД 9671939 .
- ^ Дорус С., Андерсон Дж.Р., Валлендер Э.Дж., Гилберт С.Л., Чжан Л., Чемник Л.Г. и др. (июль 2006 г.). «Sonic Hedgehog, ключевой ген развития, претерпел интенсивную молекулярную эволюцию у приматов» . Молекулярная генетика человека . 15 (13): 2031–2037. дои : 10.1093/hmg/ddl123 . ПМИД 16687440 .
- ^ Граул Р.К., Сади В. (2001). «Эволюционные взаимоотношения между рецепторами, связанными с G-белком, с использованием подхода с кластерной базой данных» . AAPS PharmSci . 3 (2): 25–42. дои : 10.1208/ps030212 . ПМЦ 2779559 . ПМИД 11741263 .
- ^ Нусс Р. (ноябрь 2003 г.). «Wnts и Hedgehogs: липид-модифицированные белки и сходство механизмов передачи сигналов на поверхности клетки» . Разработка . 130 (22): 5297–5305. дои : 10.1242/dev.00821 . ПМИД 14530294 .
- ^ Варджосало М., Ли С.П., Тайпале Дж. (февраль 2006 г.). «Расхождение механизма передачи сигнала ежей между дрозофилой и млекопитающими» . Развивающая клетка . 10 (2): 177–186. дои : 10.1016/j.devcel.2005.12.014 . ПМИД 16459297 .
- ^ Чен М.Х., Гао Н., Каваками Т., Чуанг П.Т. (август 2005 г.). «Мыши с дефицитом слитого гомолога не проявляют фенотипов, указывающих на нарушение передачи сигналов hedgehog во время эмбрионального развития» . Молекулярная и клеточная биология . 25 (16): 7042–7053. дои : 10.1128/MCB.25.16.7042-7053.2005 . ПМК 1190231 . ПМИД 16055716 .
- ^ Хуанфу Д., Андерсон К.В. (январь 2006 г.). «Передача сигналов от Smo к Ci/Gli: сохранение и расхождение путей Hedgehog от дрозофилы к позвоночным» . Разработка . 133 (1): 3–14. дои : 10.1242/dev.02169 . ПМИД 16339192 .
- ^ Перейти обратно: а б Матус Д.К., Мэги Ч.Р., Панг К., Мартиндейл М.К., Томсен Г.Х. (январь 2008 г.). «Семейство генов Hedgehog книдарий Nematostella vectensis и значение для понимания эволюции пути многоклеточных Hedgehog» . Биология развития . 313 (2): 501–518. дои : 10.1016/j.ydbio.2007.09.032 . ПМК 2288667 . ПМИД 18068698 .
- ^ Перейти обратно: а б с Адамска М., Матус Д.К., Адамски М., Грин К., Рохсар Д.С., Мартиндейл М.К., Дегнан Б.М. (октябрь 2007 г.). «Эволюционное происхождение белков ежа» . Современная биология . 17 (19): Р836–Р837. дои : 10.1016/j.cub.2007.08.010 . ПМИД 17925209 . S2CID 15158554 .
- ^ Перейти обратно: а б Снелл Э.А., Брук Н.М., Тейлор В.Р., Казан Д., Филипп Х., Холланд П.В. (февраль 2006 г.). «Необычный белок хоанофлагеллят, выделяемый в результате автокаталитической обработки Hedgehog» . Слушания. Биологические науки . 273 (1585): 401–407. дои : 10.1098/rspb.2005.3263 . ПМК 1560198 . ПМИД 16615205 .
- ^ Перейти обратно: а б с Бурглин Т.Р. (март 2008 г.). «Эволюция генов ежа и родственных ему генов, их происхождение от белков Hog у предковых эукариот и открытие нового мотива намека» . БМК Геномика . 9 (127): 127. дои : 10.1186/1471-2164-9-127 . ПМК 2362128 . ПМИД 18334026 .
- ^ Перейти обратно: а б Бурглин Т.Р. (2008). «Семейство белков Hedgehog» . Геномная биология . 9 (11): 241. doi : 10.1186/gb-2008-9-11-241 . ПМК 2614485 . ПМИД 19040769 .
- ^ Перейти обратно: а б с д Зугасти О, Раджан Дж, Кувабара П.Е. (октябрь 2005 г.). «Функция и расширение гомологов, связанных с Patched и Hedgehog, у C. elegans» . Геномные исследования . 15 (10): 1402–1410. дои : 10.1101/гр.3935405 . ПМК 1240083 . ПМИД 16204193 .
- ^ Перейти обратно: а б с д Бурглин Т.Р., Кувабара Ч.Е. (январь 2006 г.). «Гомологи сигнальной сети Hh у C. elegans» . Червячная книга : 1–14. дои : 10.1895/wormbook.1.76.1 . ПМЦ 4781598 . ПМИД 18050469 .
- ^ Хаусманн Г., фон Меринг С., Баслер К. (июнь 2009 г.). «Сигнальный путь ежа: откуда он взялся?» . ПЛОС Биология . 7 (6): e1000146. дои : 10.1371/journal.pbio.1000146 . ПМЦ 2698682 . ПМИД 19564910 .
Внешние ссылки
[ редактировать ]- Рамирес-Вебер ФА. «База данных путей ежа» . Сан-Франциско, Калифорния: Государственный университет Сан-Франциско. Архивировано из оригинала 16 июля 2006 года.
- «Схема сигнальных путей ежа» . Новус Биологикс .
| Базы данных органов управления : Национальные |
|---|