Транскрипционный фактор II H
| общий фактор транскрипции IIH, полипептид 1, 62 кДа | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | GTF2H1 | ||
| Альт. символы | БТФ2 | ||
| ген NCBI | 2965 | ||
| HGNC | 4655 | ||
| МОЙ БОГ | 189972 | ||
| RefSeq | НМ_005316 | ||
| ЮниПрот | P32780 | ||
| Другие данные | |||
| Локус | Хр. 11 п15.1-п14 | ||
| |||
| общий фактор транскрипции IIH, полипептид 2, 44 кДа | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | GTF2H2 | ||
| Альт. символы | БТФ2, ТФИИХ, БТФ2П44, Т-БТФ2П44 | ||
| ген NCBI | 2966 | ||
| HGNC | 4656 | ||
| МОЙ БОГ | 601748 | ||
| RefSeq | НМ_001515 | ||
| ЮниПрот | Q13888 | ||
| Другие данные | |||
| Локус | Хр. 5 q12.2-13.3 | ||
| |||
| общий фактор транскрипции IIH, полипептид 3, 34 кДа | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | GTF2H3 | ||
| Альт. символы | БТФ2, ТФИИХ | ||
| ген NCBI | 2967 | ||
| HGNC | 4657 | ||
| МОЙ БОГ | 601750 | ||
| RefSeq | НМ_001516 | ||
| ЮниПрот | Q13889 | ||
| Другие данные | |||
| Локус | Хр. 12 д24.31 | ||
| |||
Фактор транскрипции II H ( TF II H ) представляет собой важный белковый комплекс , играющий роль в транскрипции различных генов, кодирующих белки, и в путях эксцизионной репарации нуклеотидов ДНК (NER). TF II H впервые появился в 1989 году, когда общий фактор транскрипции-δ или основной фактор транскрипции 2 был охарактеризован как незаменимый фактор транскрипции in vitro. Этот фактор также был выделен из дрожжей и в 1992 году назван TF II H. [1] [2]
TF II H состоит из десяти субъединиц, 7 из которых ( ERCC2 /XPD, ERCC3 /XPB, GTF2H1 /p62, GTF2H4 /p52, GTF2H2 /p44, GTF2H3 /p34 и GTF2H5 /TTDA) образуют коровый комплекс. Субкомплекс киназы, активирующей циклин ( CDK7 , MAT1 и циклин H), связан с ядром через белок XPD. [3] Две субъединицы, ERCC2 /XPD и ERCC3/ XPB , обладают геликазной и АТФазной активностью и помогают создавать пузырь транскрипции . В пробирке эти субъединицы необходимы для транскрипции только в том случае, если матрица ДНК еще не денатурирована или не имеет сверхскрученной структуры .
Две другие субъединицы TF II H, CDK7 и циклин H , фосфорилируют сериновые аминокислоты домене РНК-полимеразы II в С-концевом и, возможно, в других белках, участвующих в клеточном цикле . Помимо жизненно важной функции инициации транскрипции, TF II H также участвует в эксцизионной репарации нуклеотидов .
История TF II H
[ редактировать ]До того, как TF II H определила его, у него было несколько названий. Он был выделен в 1989 году из печени крыс, известный по фактору транскрипции дельта. Когда он был идентифицирован в раковых клетках, он был известен в то время как основной фактор транскрипции 2. Кроме того, при выделении из дрожжей он был назван фактором транскрипции B. Наконец, в 1992 году он был известен как TF II H. [4]
Структура ТФ II H
[ редактировать ]TF II H представляет собой комплекс из десяти субъединиц; семь из этих субъединиц составляют «ядро», тогда как три составляют диссоциируемый модуль «CAK» (CDK-активирующая киназа). [5] Ядро состоит из субъединиц XPB, XPD, p62 , p52 , p44, p34 и p8 , тогда как CAK состоит из CDK7 , циклина H и MAT1 . [6]
Функции
[ редактировать ]Общая функция TF II H:
(NER)TF II H представляет собой общий фактор транскрипции, который привлекает РНК Pol II к промоторам генов. Он действует как геликаза, раскручивающая ДНК. Он также раскручивает ДНК после того, как повреждение ДНК было распознано либо путем глобального восстановления генома (GGR), либо путем транскрипционно-связанной репарации (TCR) NER. [8] [9] Очищенный TF II H играет роль в остановке дальнейшего синтеза РНК путем активации циклического пептида α-аманитина.
Трихотиодистрофия
[ редактировать ]Мутация в генах ERCC3 ( XPB ), ERCC2 ( XPD ) или GTF2H5 ( TTDA ) вызывает трихотиодистрофию — состояние, характеризующееся светочувствительностью , ихтиозом , ломкостью волос и ногтей, умственными нарушениями, снижением фертильности и/или низким ростом. [10]
Болезнь
[ редактировать ]Известно, что генетический полиморфизм генов, кодирующих субъединицы TF II H, связан с повышенной предрасположенностью к раку во многих тканях, например; ткань кожи, ткань молочной железы и ткань легких. Мутации в субъединицах (таких как XPD и XPB) могут привести к множеству заболеваний, включая пигментную ксеродермию (XP) или XP в сочетании с синдромом Кокейна . [11] Помимо генетических вариаций, кодируемые вирусом белки также нацелены на TF II H. [12]
восстановление ДНК
[ редактировать ]TF II H участвует в эксцизионной репарации нуклеотидов (NER), открывая двойную спираль ДНК после первоначального распознавания повреждения. NER — это многоэтапный путь, который устраняет широкий спектр различных повреждений, нарушающих нормальное спаривание оснований, включая объемные химические повреждения и повреждения, вызванные УФ-излучением. Лица с мутационными дефектами в генах, определяющих белковые компоненты, катализирующие путь NER, включая компоненты TF II H, часто демонстрируют признаки преждевременного старения. [10] [13] (см. теорию старения, связанную с повреждением ДНК ).
Ингибиторы
[ редактировать ]мощных биоактивных натуральных продуктах, таких как триптолид, которые ингибируют транскрипцию млекопитающих посредством ингибирования субъединицы XPB общего фактора транскрипции TF II H в качестве конъюгата глюкозы для воздействия на гипоксические раковые клетки с повышенной экспрессией переносчика глюкозы. Недавно сообщалось о [14]
Механизм TF II H восстановления поврежденной последовательности ДНК
[ редактировать ]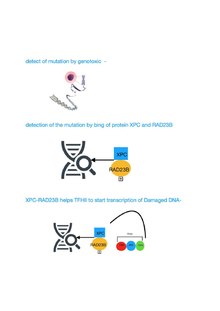
Ссылки
[ редактировать ]- ^ Флорес О, Лу Х, Рейнберг Д (февраль 1992 г.). «Факторы, участвующие в специфической транскрипции РНК-полимеразой II млекопитающих. Идентификация и характеристика фактора IIH» . Журнал биологической химии . 267 (4): 2786–93. дои : 10.1016/S0021-9258(18)45947-9 . ПМИД 1733973 .
- ^ Ким Т.К., Эбрайт Р.Х. , Рейнберг Д. (май 2000 г.). «Механизм плавления АТФ-зависимого промотора транскрипционным фактором IIH». Наука . 288 (5470): 1418–22. Бибкод : 2000Sci...288.1418K . дои : 10.1126/science.288.5470.1418 . ПМИД 10827951 .
- ^ Ли Т.И., Молодой РА (2000). «Транскрипция генов, кодирующих эукариотические белки». Ежегодный обзор генетики . 34 : 77–137. дои : 10.1146/annurev.genet.34.1.77 . ПМИД 11092823 .
- ^ Римель Дж. К., Таатьес DJ (июнь 2018 г.). «Незаменимый и многофункциональный комплекс TFIIH» . Белковая наука . 27 (6): 1018–1037. дои : 10.1002/pro.3424 . ПМЦ 5980561 . ПМИД 29664212 .
- ^ Драпкин Р., Рирдон Дж.Т., Ансари А., Хуанг Дж.К., Завел Л., Ан К., Санкар А., Рейнберг Д. (апрель 1994 г.). «Двойная роль TFIIH в эксцизионной репарации ДНК и в транскрипции РНК-полимеразой II». Природа . 368 (6473): 769–72. Бибкод : 1994Natur.368..769D . дои : 10.1038/368769a0 . ПМИД 8152490 . S2CID 4363484 .
- ^ Драпкин Р., Рирдон Дж.Т., Ансари А., Хуанг Дж.К., Завел Л., Ан К., Санкар А., Рейнберг Д. (апрель 1994 г.). «Двойная роль TFIIH в эксцизионной репарации ДНК и в транскрипции РНК-полимеразой II». Природа . 368 (6473): 769–72. Бибкод : 1994Natur.368..769D . дои : 10.1038/368769a0 . ПМИД 8152490 . S2CID 4363484 .
- ^ Jump up to: а б Compe E, Egly JM (май 2012 г.). «TFIIH: когда транскрипция встретила восстановление ДНК» . Обзоры природы. Молекулярно-клеточная биология . 13 (6): 343–54. дои : 10.1038/nrm3350 . ПМИД 22572993 . S2CID 29077515 .
- ^ Хугстратен Д., Нигг А.Л., Хит Х., Маллендерс Л.Х., ван Дрил Р., Хоймейкерс Дж.Х., Вермюлен В., Хаутсмюллер А.Б. (ноябрь 2002 г.). «Быстрое переключение TFIIH между транскрипцией РНК-полимеразой I и II и репарацией ДНК in vivo» . Молекулярная клетка . 10 (5): 1163–74. дои : 10.1016/s1097-2765(02)00709-8 . ПМИД 12453423 .
- ^ Ассфальг Р., Лебедев А., Гонсалес О.Г., Шеллинг А., Кох С., Ибен С. (январь 2012 г.). «TFIIH является фактором элонгации РНК-полимеразы I» . Исследования нуклеиновых кислот . 40 (2): 650–9. дои : 10.1093/nar/gkr746 . ПМЦ 3258137 . ПМИД 21965540 .
- ^ Jump up to: а б Тейл А.Ф., Хоймейкерс Дж.Х., Вермюлен В. (ноябрь 2014 г.). «TTDA: большое влияние маленького белка». Экспериментальные исследования клеток . 329 (1): 61–8. doi : 10.1016/j.yexcr.2014.07.008 . ПМИД 25016283 .
- ^ О К.С., Хан С.Г., Ясперс Н.Г., Раамс А., Уэда Т., Леманн А., Фридманн П.С., Эммерт С., Грачев А., Лахлан К., Лукассан А., Бейкер CC, Кремер К.Х. (ноябрь 2006 г.). «Фенотипическая гетерогенность гена ДНК-хеликазы XPB (ERCC3): пигментная ксеродермия без синдрома Коккейна и с ним» . Человеческая мутация . 27 (11): 1092–103. дои : 10.1002/humu.20392 . ПМИД 16947863 . S2CID 22852219 .
- ^ Ле Мэй Н., Дубаэле С., Пройетти Де Сантис Л., Биллекок А., Булой М., Эгли Дж. М. (февраль 2004 г.). «Фактор транскрипции TFIIH, мишень для вируса геморрагической лихорадки долины Рифт» . Клетка . 116 (4): 541–50. дои : 10.1016/s0092-8674(04)00132-1 . ПМИД 14980221 . S2CID 14312462 .
- ^ Эдифици Д., Шумахер Б. (август 2015 г.). «Нестабильность генома в развитии и старении: результаты эксцизионной репарации нуклеотидов у людей, мышей и червей» . Биомолекулы . 5 (3): 1855–69. дои : 10.3390/biom5031855 . ПМЦ 4598778 . ПМИД 26287260 .
- ^ Датан Э, Минн И, Пэн X, Хэ QL, Ан Х, Ю Б, Помпер МГ, Лю Джо (2020). «Конъюгат глюкозы-триптолида избирательно воздействует на раковые клетки в условиях гипоксии» . iScience . 23 (9): 101536. Бибкод : 2020iSci...23j1536D . дои : 10.1016/j.isci.2020.101536 . ПМК 7509213 . ПМИД 33083765 .
Внешние ссылки
[ редактировать ]- Транскрипция + Фактор + TFIIH в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
Эта по биохимии статья незавершена . Вы можете помочь Википедии, расширив ее . |