НЕ домен
| НЕ сбрасывать | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | НЕТ | ||
| Пфам | PF00989 | ||
| Пфам Клан | CL0183 | ||
| ИнтерПро | ИПР013767 | ||
| УМНЫЙ | НЕТ | ||
| PROSITE | PDOC50112 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 2phy / SCOPe / СУПФАМ | ||
| CDD | cd00130 | ||
| |||
Домен Per-Arnt-Sim ( PAS ) встречающийся — это белковый домен, во всех царствах жизни. [2] Как правило, домен PAS действует как молекулярный сенсор, посредством чего небольшие молекулы и другие белки связываются посредством связывания домена PAS. [3] [4] [5] Благодаря этой сенсорной способности домен PAS был показан как ключевой структурный мотив, участвующий в белок-белковых взаимодействиях циркадных часов , а также это общий мотив, обнаруживаемый в сигнальных белках, где он действует как сигнальный сенсор. [6] [7]
Открытие
[ редактировать ]Домены PAS обнаружены у большого количества организмов, от бактерий до млекопитающих. Домен PAS был назван в честь трех белков, в которых он был впервые обнаружен: [8]
- за период Циркадный белок
- Arnt - ядерный транслокатор арильного углеводородного рецептора
- Сим – целеустремленный белок
С момента первоначального открытия домена PAS у бактерий и эукариот было обнаружено большое количество сайтов связывания домена PAS. Подмножество белков PAS LOV реагирует на кислород, свет и напряжение. [9]
Структура
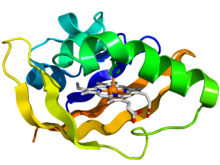
[ редактировать ]Хотя домен PAS демонстрирует некоторую степень вариабельности последовательности, трехмерная структура ядра домена PAS в целом консервативна. [10] Это ядро состоит из пятинитевого антипараллельного β-листа и нескольких α-спиралей. Структурные изменения в результате передачи сигналов преимущественно происходят внутри β-листа . Эти сигналы распространяются через α-спирали ядра к ковалентно связанному эффекторному домену. [11] В 1998 году архитектура ядра домена PAS была впервые охарактеризована в структуре фотоактивного желтого белка (PYP) из Halorhodospira halophila . [10] Во многих белках требуется димер доменов PAS, при этом один связывает лиганд, а другой опосредует взаимодействие с другими белками. [5]
Примеры PAS в организмах
[ редактировать ]Известные домены PAS имеют в среднем менее 20% идентичности парных последовательностей , что означает, что они удивительно различны. [10] Домены PAS часто обнаруживаются в белках с другими механизмами восприятия окружающей среды. Кроме того, многие домены PAS прикреплены к фоторецепторным клеткам. [12]
Бактерии
[ редактировать ]Часто в бактериальном царстве домены PAS располагаются на аминоконце сигнальных белков, таких как сенсорные гистидинкиназы , синтезаторы и гидролазы циклического ди-ГМФ , а также белки хемотаксиса, принимающие метил . [10]
Нейроспора
[ редактировать ]В присутствии света белые воротнички-1 (WC-1) и белые воротнички-2 (WC-2) димеризуются посредством доменов PAS, что активирует трансляцию FRQ . [13]
Дрозофила
[ редактировать ]В присутствии света CLK и CYC прикрепляются через домен PAS, активируя трансляцию PER, который затем связывается с Тимом через домен PER PAS. Следующие гены содержат PAS-связывающие домены: PER, Tim, CLK, CYC.
Арабидопсис
[ редактировать ]Домен PAS обнаружен в генах ZTL и NPH1. Эти домены очень похожи на домен PAS, обнаруженный в циркадно-ассоциированном белке WC-1 Neurospora . [14]
Млекопитающие
[ редактировать ]Циркадные часы, которые в настоящее время известны у млекопитающих, начинаются, когда свет активирует BMAL1 и CLK связывание через их домены PAS. Этот активаторный комплекс регулирует Per1, Per2 и Per3, все из которых имеют домены PAS, которые используются для связывания с криптохромами 1 и 2 ( семейство CRY 1,2 ). Следующие гены млекопитающих содержат PAS-связывающие домены: Per1, Per2, Per3, Cry1, Cry2, Bmal, Clk, Pasd1.
Другие роли PAS млекопитающих
[ редактировать ]У млекопитающих оба домена PAS играют важную роль. PAS A отвечает за белок-белковые взаимодействия с другими белками домена PAS, тогда как PAS B играет более разностороннюю роль. Он опосредует взаимодействие с шаперонинами и другими небольшими молекулами, такими как диоксин , но домены PAS B в NPAS2 , гомологе гена clk дрозофилы, и фактор, индуцируемый гипоксией (HIF), также помогают опосредовать связывание лиганда . [12] Более того, было показано, что домены PAS, содержащие белок NPAS2, заменяют ген Clock у мутантных мышей, у которых полностью отсутствует ген Clock. [15]
Домен PAS также напрямую взаимодействует с BHLH . Обычно он расположен на С-конце белка BHLH. Домены PAS, содержащие белки BHLH, образуют белок BHLH-Pas, обычно обнаруживаемый и кодируемый в HIF, для которого требуются как домен PAS, так и домен BHLH, а также ген Clock. [16] [17] [18]
Ссылки
[ редактировать ]- ^ PDB : 1y28 ; Данхэм СМ, Диум Э.М., Такерман-младший, Гонсалес Дж., Скотт В.Г., Жиль-Гонсалес М.А. (июль 2003 г.). «Дистальный аргинин в чувствительных к кислороду доменах гема-PAS необходим для связывания лигандов, передачи сигнала и структуры» . Биохимия . 42 (25): 7701–8. дои : 10.1021/bi0343370 . ПМИД 12820879 . S2CID 14090693 .
- ^ Генри, Джонатан Т.; Кроссон, Шон (1 января 2011 г.). «Лигандсвязывающие домены PAS в геномном, клеточном и структурном контексте» . Ежегодный обзор микробиологии . 65 : 261–286. doi : 10.1146/annurev-micro-121809-151631 . ПМК 3298442 . ПМИД 21663441 .
- ^ Лю, Ю С.; Мачука, Майра А.; Бекхэм, Симона А.; Гинцбург, Менахем Дж.; Ружейникова, Анна (1 октября 2015 г.). «Структурная основа распознавания аминокислот и трансмембранной передачи сигналов сенсорными доменами тандемных хеморецепторов Per-Arnt-Sim (тандемный PAS)». Acta Crystallographica Раздел D. 71 (10): 2127–2136. Бибкод : 2015AcCrD..71.2127L . дои : 10.1107/S139900471501384X . ПМИД 26457436 .
- ^ Мёглич, Андреас; Айерс, Ребекка А.; Моффат, Кейт (14 октября 2009 г.). «Структура и механизм передачи сигналов доменов Per-ARNT-Sim» . Структура . 17 (10): 1282–1294. дои : 10.1016/j.str.2009.08.011 . ПМК 3092527 . ПМИД 19836329 .
- ^ Перейти обратно: а б Хенниг, Свен; Штраус, Хольгер М.; Ванселов, Катя; Йылдыз, Озкан; Шульце, Сабрина; Аренс, Джулия; Крамер, Ахим; Вольф, Ева (28 апреля 2009 г.). «Структурный и функциональный анализ взаимодействий доменов PAS часовых белков Drosophila PERIOD и Mouse PERIOD2» . ПЛОС Биология . 7 (4): e1000094. дои : 10.1371/journal.pbio.1000094 . ПМЦ 2671562 . ПМИД 19402751 .
- ^ Понтинг К.П., Аравинд Л. (ноябрь 1997 г.). «PAS: появляется многофункциональное семейство доменов» . Курс. Биол . 7 (11): Р674–7. дои : 10.1016/S0960-9822(06)00352-6 . ПМИД 9382818 . S2CID 14105830 .
- ^ Хефти М.Х., Франсуа К.Дж., де Врис С.К., Диксон Р., Вервурт Дж. (март 2004 г.). «Складка PAS. Переопределение домена PAS на основе структурного предсказания» . Евро. Дж. Биохим . 271 (6): 1198–208. дои : 10.1111/j.1432-1033.2004.04023.x . ПМИД 15009198 .
- ^ Мёглич, Андреас; Айерс, Ребекка А.; Моффат, Кейт (14 октября 2009 г.). «Структура и механизм сигнализации доменов Per-ARNT-Sim» . Структура . 17 (10): 1282–1294. дои : 10.1016/j.str.2009.08.011 . ПМК 3092527 . ПМИД 19836329 .
- ^ Розато, Эцио; Таубер, Эран; Кириаку, Хараламбос П. (1 января 2006 г.). «Молекулярная генетика циркадных часов плодовых мушек» . Европейский журнал генетики человека . 14 (6): 729–738. дои : 10.1038/sj.ejhg.5201547 . ПМИД 16721409 .
- ^ Перейти обратно: а б с д Генри, Джонатан Т.; Кроссон, Шон (1 января 2011 г.). «Лиганд-связывающие PAS-домены в геномном, клеточном и структурном контексте» . Ежегодный обзор микробиологии . 65 : 261–286. doi : 10.1146/annurev-micro-121809-151631 . ПМК 3298442 . ПМИД 21663441 .
- ^ Мёглич, А; Айерс, РА; Моффат, К. (2009). «Структура и механизм сигнализации доменов Per-ARNT-Sim» . Структура . 17 (10): 1282–94. дои : 10.1016/j.str.2009.08.011 . ПМК 3092527 . ПМИД 19836329 .
- ^ Перейти обратно: а б Макинтош, Брайан; Хогенеш, Джон; Брэдфилд, Кристофер (2010). «Белки Per-Arnt-Sim млекопитающих в адаптации к окружающей среде». Ежегодный обзор физиологии . 72 : 625–645. doi : 10.1146/annurev-psyol-021909-135922 . ПМИД 20148691 .
- ^ Хармер, Стейси Л.; Панда, Сатчидананда; Кей, Стив А. (28 ноября 2003 г.). «Молекулярные основы циркадных ритмов». Ежегодный обзор клеточной биологии и биологии развития . 17 : 215–253. дои : 10.1146/annurev.cellbio.17.1.215 . ПМИД 11687489 .
- ^ Сомерс, Дэвид; Шульц, Томас; Кей, Стив; Милнамоу, Морин (2000). «ZEITLUPE кодирует новый ассоциированный с часами белок PAS из Arabidopsis» . Клетка . 101 (3): 319–329. дои : 10.1016/S0092-8674(00)80841-7 . ПМИД 10847686 . S2CID 3013788 .
- ^ Дебрюйн Дж.П., Нотон Э., Ламберт К.М., Мэйвуд Э.С., Уивер Д.Р., Репперт С.М. (май 2006 г.). «Потрясение часов: ЧАСЫ мыши не требуются для функции циркадного осциллятора» . Нейрон . 50 (3): 465–77. дои : 10.1016/j.neuron.2006.03.041 . ПМИД 16675400 . S2CID 19028601 .
- ^ Джонс, Сьюзен (1 января 2004 г.). «Обзор основных белков спираль-петля-спираль» . Геномная биология . 5 (6): 226. doi : 10.1186/gb-2004-5-6-226 . ПМК 463060 . ПМИД 15186484 .
- ^ Кэ, Циндун; Коста, Макс (1 ноября 2006 г.). «Фактор-1, индуцируемый гипоксией (HIF-1)». Молекулярная фармакология . 70 (5): 1469–1480. дои : 10.1124/моль.106.027029 . ПМИД 16887934 . S2CID 2522614 .
- ^ Ван, Г.Л.; Цзян, Б.Х.; Рю, EA; Семенца, Г.Л. (6 июня 1995 г.). «Фактор 1, индуцируемый гипоксией, представляет собой гетеродимер основная спираль-петля-спираль-PAS, регулируемый клеточным напряжением O2» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (12): 5510–5514. Бибкод : 1995PNAS...92.5510W . дои : 10.1073/pnas.92.12.5510 . ПМК 41725 . ПМИД 7539918 .