Циклический ди-GMP
 | |
| Имена | |
|---|---|
| Систематическое название ИЮПАК (2R , 3R , 3aS , 7aR , 9R , 10R , 10aS , 14aR ) -2,9-бис(2-амино-6-оксо-1,6-дигидро-9H - пурин -9-ил)-3,5,10,12-тетрагидроксиоктагидро- 2H , 5H , 7H ,12H - 5λ 5 , 12 мин. 5 -дифуро[3,2- d :3',2'- j ][1,3,7,9,2,8]тетраоксадифосфациклододецин-5,12-дион | |
| Другие имена Циклический дигуанилат; 3',5'-циклическая дигуаниловая кислота; c-ди-GMP; 5ГП-5ГП | |
| Идентификаторы | |
3D model ( JSmol ) | |
| ХимическийПаук | |
ПабХим CID | |
Панель управления CompTox ( EPA ) | |
| Характеристики | |
| С 20 Ч 24 Н 10 О 14 П 2 | |
| Молярная масса | 690.09 g/mol |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
Циклический ди-ГМФ (также называемый циклическим дигуанилатом и с-ди- ГМФ ) представляет собой вторичный мессенджер , используемый в передаче сигналов у широкого спектра бактерий . [1] Известно, что циклический ди-ГМФ не используется архей и наблюдался только у эукариот в Dictyostelium . [2] Биологическая роль циклического ди-ГМФ была впервые обнаружена, когда он был идентифицирован как аллостерический активатор целлюлозосинтазы, обнаруженной в Gluconacetobacter xylinus, для производства микробной целлюлозы . [3]
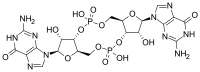
По структуре это цикл, содержащий всего два гуаниновых основания, связанных рибозой и фосфатом .
Функция
[ редактировать ]Контакт с поверхностями увеличивает количество c-di-GMP, который увеличивает транскрипцию , трансляцию и посттрансляцию экзополисахаридов (см . (EPS) и других компонентов матрицы внеклеточного полимерного вещества обзор Jenal et al 2017). [4] У бактерий определенные сигналы передаются путем синтеза или разложения циклического ди-ГМФ. Циклический ди-ГМФ синтезируется белками с дигуанилатциклазной активностью. Эти белки обычно имеют характерный мотив GGDEF , который относится к консервативной последовательности из пяти аминокислот . На деградацию циклического ди-ГМФ влияют белки с фосфодиэстеразной активностью. Эти белки имеют аминокислотный мотив EAL или HD-GYP. Процессы, которые, как известно, регулируются циклическим ди-ГМФ, по крайней мере, у некоторых организмов, включают образование биопленок (например, матрицы EPS, обнаруженные Steiner et al 2013), [4] подвижность (особенно переход от подвижного к сидячему состоянию, см. обзор Jenal et al, 2017) [4] и вирулентности производство фактора .
Регулирование
[ редактировать ]Циклические уровни ди-ГМФ регулируются с помощью различных механизмов. Многие белки с доменами GGDEF, EAL или HD-GYP встречаются с другими доменами, которые могут принимать сигналы, такими как домены PAS . Считается, что ферменты, которые расщепляют или синтезируют циклический ди-ГМФ, локализованы в определенных областях клетки, где они влияют на ресиверы в ограниченном пространстве. [1] У Gluconacetobacter xylinus c-ди-GMP стимулирует полимеризацию глюкозы в целлюлозу, являясь высокоаффинным аллостерическим активатором фермента целлюлозосинтазы. [3] Некоторые ферменты дигуанилатциклазы аллостерически ингибируются циклическим ди-ГМФ.
Циклические уровни ди-ГМФ регулируют другие процессы с помощью ряда механизмов. Целлюлосинтаза Gluconacetobacter xylinus аллостерически стимулируется циклическим ди-ГМФ, что представляет собой механизм, с помощью которого циклический ди-ГМФ может регулировать активность целлюлозо-синтазы. Было показано, что домен PilZ связывает циклический ди-ГМФ и, как полагают, участвует в зависимой от циклического ди-ГМФ регуляции, но механизм, с помощью которого он это делает, неизвестен. Недавние структурные исследования доменов PilZ двух видов бактерий показали, что домены PilZ резко меняют конформацию при связывании с циклическим ди-ГМФ. [5] [6] Это приводит к строгому выводу, что конформационные изменения в доменах PilZ позволяют регулировать активность целевых эффекторных белков (таких как целлюлозосинтаза) с помощью циклического ди-ГМФ. Рибопереключатели, называемые циклическим рибопереключателем ди-GMP-I и циклическим рибопереключателем ди-GMP-II, регулируют экспрессию генов в ответ на концентрации циклического ди-ГМФ у различных бактерий, но не у всех бактерий, которые, как известно, используют циклический ди-ГМФ.
Хост-ассоциация
[ редактировать ]Циклический ди-ГМФ связан с ассоциацией хозяина у многих видов Pseudomonas . В эксперименте, в котором Pseudomonas lurida выращивали вместе с нематодой-хозяином Caenorhabditis elegans, в определенных генах наблюдались генетические мутации, которые повышали уровень c-di-GMP, вызывая появление специализированного морфотипа хозяина. Мутации, влияющие на регуляцию c-di-GMP, были обнаружены при полногеномном секвенировании. Гены wspE и wspF оперона wsp обнаруживали мутации, которые повышали экспрессию c-di-GMP. Кроме того, было установлено, что мутации в гене rph также влияют на экспрессию c-di-GMP, что является новым открытием, поскольку ранее rph не был связан с регуляцией c-di-GMP. [7]
Обзор роли c-di-GMP в Caulobacter crescentus , Pseudomonas aeruginosa , Komagataeibacter xylinus / Gluconacetobacter xylinus , Myxococcus xanthus , Bdellovibrio бактериоворус и Pseudomonas fluorescens см. в Jenal et al 2017. [4]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Тамайо Р., Пратт Дж.Т., Камилли А. (2007). «Роль циклического дигуанилата в регуляции бактериального патогенеза» . Ежегодный обзор микробиологии . 61 : 131–148. дои : 10.1146/annurev.micro.61.080706.093426 . ПМЦ 2776827 . ПМИД 17480182 .
- ^ Чен Ж., Шаап П. (август 2012 г.). «Прокариотный мессенджер c-di-GMP запускает дифференцировку стебельчатых клеток у Dictyostelium» . Природа . 488 (7413): 680–683. Бибкод : 2012Natur.488..680C . дои : 10.1038/nature11313 . ПМЦ 3939355 . ПМИД 22864416 .
- ^ Jump up to: а б Росс П., Вейнхаус Х., Алони Ю., Михаэли Д., Вайнбергер-Оана П., Майер Р. и др. (1987). «Регуляция синтеза целлюлозы у Acetobacter xylinum с помощью циклической дигуаниловой кислоты». Природа . 325 (6101): 279–281. Бибкод : 1987Natur.325..279R . дои : 10.1038/325279a0 . ПМИД 18990795 . S2CID 4305512 .
- ^ Jump up to: а б с д Лавенти Би Джей, Дженал Ю (сентябрь 2020 г.). «Поверхностное зондирование и адаптация бактерий». Ежегодный обзор микробиологии . 74 (1). Годовые обзоры : 735–760. doi : 10.1146/annurev-micro-012120-063427 . ПМИД 32905753 . S2CID 221622525 .
- ^ Бенах Дж., Сваминатан С.С., Тамайо Р., Хандельман С.К., Фолта-Стогнев Э., Рамос Дж.Э. и др. (декабрь 2007 г.). «Структурная основа передачи сигнала циклического дигуанилата доменами PilZ» . Журнал ЭМБО . 26 (24). Европейская организация молекулярной биологии: 5153–5166. дои : 10.1038/sj.emboj.7601918 . ПМК 2140105 . ПМИД 18034161 .
- ^ Ко Дж., Рю К.С., Ким Х., Шин Дж.С., Ли Дж.О., Чеонг С. и др. (апрель 2010 г.). «Структура PP4397 раскрывает молекулярную основу различных способов связывания c-di-GMP белками домена Pilz». Журнал молекулярной биологии . 398 (1): 97–110. дои : 10.1016/j.jmb.2010.03.007 . ПМИД 20226196 .
- ^ Обенг Н., Червински А., Шютц Д., Михельс Дж., Лейперт Дж., Бансепт Ф. и др. (октябрь 2023 г.). «Бактериальный c-di-GMP играет ключевую роль в установлении симбиоза хозяина и микроба» . Природная микробиология . 8 (10): 1809–1819. дои : 10.1038/s41564-023-01468-x . ПМЦ 10522488 . ПМИД 37653009 .