Рибопереключатель

В молекулярной биологии рибопереключатель — это регуляторный сегмент молекулы информационной РНК , который связывает небольшую молекулу , что приводит к изменению производства белков , кодируемых мРНК. [1] [2] [3] [4] Таким образом, мРНК, содержащая рибопереключатель, непосредственно участвует в регуляции собственной активности в ответ на концентрацию своей эффекторной молекулы. Открытие того, что современные организмы используют РНК для связывания небольших молекул и дискриминации близкородственных аналогов, расширило известные природные возможности РНК за пределы ее способности кодировать белки , катализировать реакции или связывать другие РНК или белковые макромолекулы .
В первоначальном определении термина «рибопереключатель» указывалось, что они напрямую определяют концентрации низкомолекулярных метаболитов . [5] Хотя это определение по-прежнему широко используется, некоторые биологи использовали более широкое определение, включающее другие цис-регуляторные РНК . Однако в этой статье будут обсуждаться только рибопереключатели, связывающие метаболиты.
Большинство известных рибопереключателей встречаются у бактерий , но функциональные рибопереключатели одного типа ( рибопереключатель TPP ) были обнаружены у архей, растений и некоторых грибов . Рибопереключатели TPP также были предсказаны у архей . [6] но не прошли экспериментальную проверку.
и открытия История
До открытия рибопереключателей механизм регуляции некоторых генов, участвующих в нескольких метаболических путях, оставался загадочным. Накапливающиеся данные все чаще указывают на беспрецедентную на тот момент идею о том, что участвующие мРНК могут напрямую связывать метаболиты, влияя на их собственную регуляцию. Эти данные включали консервативные вторичные структуры РНК, часто обнаруживаемые в нетранслируемых областях ( UTR ) соответствующих генов, а также успех процедур по созданию искусственных малых молекул-связывающих РНК, называемых аптамерами . [7] [8] [9] [10] [11] В 2002 году были опубликованы первые исчерпывающие доказательства существования нескольких классов рибопереключателей, включая анализы безбелкового связывания, а рибопереключатели, связывающиеся с метаболитами, были признаны новым механизмом регуляции генов. [5] [12] [13] [14]
Многие из самых ранних обнаруженных рибопереключателей соответствовали «мотивам» (паттернам) консервативных последовательностей в 5'-UTR , которые, по-видимому, соответствовали структурированной РНК. Например, сравнительный анализ вышестоящих областей нескольких генов, которые, как ожидается, будут совместно регулироваться, привел к описанию S-бокса. [15] (теперь рибопереключатель SAM-I), THI-box [9] (область внутри рибопереключателя TPP), элемент RFN [8] (теперь рибопереключатель FMN) и B 12 -бокс [16] (часть кобаламинового рибопереключателя), а в некоторых случаях экспериментальные демонстрации того, что они участвуют в регуляции генов посредством неизвестного механизма. Биоинформатика сыграла свою роль в недавних открытиях, благодаря растущей автоматизации базовой стратегии сравнительной геномики . Баррик и др. (2004) [17] использовали BLAST , чтобы найти НТО, гомологичные всем НТО в Bacillus subtilis . Некоторые из этих гомологичных наборов были проверены на предмет консервативной структуры, в результате чего было обнаружено 10 РНК-подобных мотивов. Три из них были позже экспериментально подтверждены как рибопереключатели glmS, глицин и PreQ1-I (см. ниже). Последующие усилия по сравнительной геномике с использованием дополнительных таксонов бактерий и усовершенствованных компьютерных алгоритмов выявили дополнительные рибопереключатели, которые экспериментально подтверждены, а также консервативные структуры РНК, которые, как предполагается, функционируют как рибопереключатели. [18] [19] [20]
Механизмы [ править ]
Рибопереключатели часто концептуально разделяют на две части: аптамер и платформу экспрессии. Аптамер напрямую связывает небольшую молекулу, и платформа экспрессии претерпевает структурные изменения в ответ на изменения аптамера. Платформа экспрессии — это то, что регулирует экспрессию генов.
Платформы экспрессии обычно отключают экспрессию генов в ответ на небольшую молекулу, но некоторые включают ее. Экспериментально были продемонстрированы следующие механизмы рибопереключения.
- Контролируемое рибопереключателем образование rho-независимых шпилек терминации транскрипции приводит к преждевременной терминации транскрипции.
- Сворачивание, опосредованное рибопереключателем, изолирует участок связывания рибосомы , тем самым ингибируя трансляцию .
- Рибопереключатель представляет собой рибозим , который расщепляет себя в присутствии достаточных концентраций своего метаболита.
- Альтернативные структуры рибопереключателя влияют на сплайсинг пре-мРНК.
- Рибопереключатель TPP в Neurospora crassa (гриб) контролирует альтернативный сплайсинг для условного образования восходящей открытой рамки считывания (uORF), тем самым влияя на экспрессию нижестоящих генов. [21]
- Рибопереключатель TPP в растениях модифицирует сплайсинг и альтернативный процессинг 3'-конца. [22] [23]
- Рибопереключатель Clostridium acetobutylicum регулирует соседний ген, который не является частью того же транскрипта мРНК. В этой регуляции рибопереключатель вмешивается в транскрипцию гена. Механизм неясен, но может быть вызван столкновениями между двумя единицами РНК-полимеразы, поскольку они одновременно транскрибируют одну и ту же ДНК. [24]
- Рибопереключатель Listeria monocytogenes регулирует экспрессию нижележащего гена. Однако транскрипты рибопереключателя впоследствии модулируют экспрессию гена, расположенного в другом месте генома. [25] Эта транс- регуляция происходит посредством спаривания оснований с мРНК дистального гена. По мере повышения температуры бактерии рибопереключатель плавится, обеспечивая транскрипцию. Неопубликованные исследования студентов создали аналогичный рибопереключатель или «термосенсор» посредством случайного мутагенеза последовательности Listeria monocytogenes. [26]
Типы [ править ]

Ниже приводится список экспериментально подтвержденных рибопереключателей, упорядоченных по лигандам.
- Рибопереключатель кобаламина (также B 12 элемент ), который связывает либо аденозилкобаламин (коферментная форма витамина B 12 ), либо аквокобаламин для регулирования биосинтеза кобаламина и транспорта кобаламина и подобных метаболитов, а также других генов.
- Рибопереключатели циклического AMP-GMP связывают сигнальную молекулу циклического AMP-GMP . Эти рибопереключатели структурно связаны с циклическими рибопереключателями ди-GMP-I (см. также «циклический ди-GMP» ниже).
- циклические рибопереключатели ди-АМФ (также называемые ydaO/yuaA ) связывают сигнальную молекулу циклического ди-АМФ .
- Рибопереключатели циклического ди-ГМФ связывают сигнальную молекулу циклического ди-ГМФ , чтобы регулировать различные гены, контролируемые этим вторым мессенджером. Известны два класса циклических рибопереключателей ди-GMP: циклические рибопереключатели ди-GMP-I и циклические рибопереключатели ди-GMP-II . Эти классы не кажутся структурно связанными.
- Фторидные рибопереключатели воспринимают ионы фтора и помогают выдерживать высокие уровни фторида .
- Рибопереключатель FMN (также RFN-элемент ) связывает флавинмононуклеотид (FMN) для регулирования и транспорта рибофлавина биосинтеза .
- glmS рибопереключатель , который представляет собой рибозим, который расщепляет себя при наличии достаточной концентрации глюкозамин-6-фосфата .
- Глутаминовые рибопереключатели связывают глютамин , чтобы регулировать гены, участвующие в метаболизме глютамина и азота , а также короткие пептиды с неизвестной функцией. Известны два класса глутаминовых рибопереключателей: мотив glnA РНК и мотив нисходящего пептида . Считается, что эти классы структурно связаны (см. обсуждения в этих статьях).
- Глициновый рибопереключатель связывает глицин, регулируя гены метаболизма глицина, включая использование глицина в качестве источника энергии. До 2012 года этот рибопереключатель считался единственным, который демонстрирует кооперативное связывание , поскольку он содержит смежные двойные аптамеры. Хотя кооперативность больше не доказана, причина возникновения двойных аптамеров все еще остается неоднозначной. [27]
- Лизиновый рибопереключатель (также L-box ) связывает лизин, регулируя биосинтез, катаболизм и транспорт лизина.
- Марганцевые рибопереключатели связывают марганца ионы .
- Нико-рибопереключатели связывают ионы металлов никеля и кобальта .
- Рибопереключатели PreQ1 связывают пре-кеуозин 1 , чтобы регулировать гены, участвующие в синтезе или транспортировке этого предшественника к кевозину . Известны три совершенно различных класса рибопереключателей PreQ1: рибопереключатели PreQ1-I , рибопереключатели PreQ1-II и рибопереключатели PreQ1-III . Связывающий домен рибопереключателей PreQ1-I необычно мал среди природных рибопереключателей. Рибопереключатели PreQ1-II, которые встречаются только у некоторых видов родов Streptococcus и Lactococcus , имеют совершенно другую структуру и крупнее, как и рибопереключатели PreQ1-III.
- Пуриновые рибопереключатели связывают пурины , регулируя метаболизм и транспорт пуринов. Различные формы пуринового рибопереключателя связывают гуанин (форма, первоначально известная как G-бокс ) или аденин . Специфичность гуанина или аденина полностью зависит от взаимодействий Уотсона-Крика с одним пиримидином в рибопереключателе в положении Y74. В гуаниновом рибопереключателе этим остатком всегда является цитозин (т.е. С74), в адениновом остатке - всегда урацил (т.е. U74). Гомологичные типы пуриновых рибопереключателей связывают дезоксигуанозин , но имеют более существенные различия, чем мутация одного нуклеотида.
- Рибопереключатели SAH связывают S-аденозилгомоцистеин, регулируя гены, участвующие в переработке этого метаболита, который образуется при S-аденозилметионина в реакциях метилирования. использовании
- Рибопереключатели SAM связывают S-аденозилметионин (SAM), регулируя метионина биосинтез и транспорт и SAM. Известны три различных рибопереключателя SAM: SAM-I (первоначально называвшийся S-box ), SAM-II и рибопереключатель S MK box . SAM-I широко распространен у бактерий, но SAM-II обнаружен только у альфа- , бета- и некоторых гаммапротеобактерий . Рибопереключатель коробки S MK встречается только в отряде Lactobacillales . Эти три разновидности рибопереключателей не имеют явного сходства с точки зрения последовательности или структуры. Четвертая разновидность, рибопереключатели SAM-IV , по-видимому, имеет лиганд-связывающее ядро, подобное рибопереключателям SAM-I, но в контексте отдельного каркаса.
- Рибопереключатели SAM-SAH связывают как SAM, так и SAH с одинаковым сродством. Поскольку они всегда способны регулировать гены, кодирующие метионин-аденозилтрансферазу , было высказано предположение, что физиологически значимым является только их связывание с SAM.
- Тетрагидрофолатные рибопереключатели связывают тетрагидрофолат для регулирования синтеза и транспорта генов.
- Рибопереключатели TPP (также THI-box) связывают пирофосфат тиамина (TPP) для регулирования биосинтеза и транспорта тиамина , а также транспорта аналогичных метаболитов. Это единственный рибопереключатель, обнаруженный на сегодняшний день у эукариот. [28]
- Рибопереключатели ZMP/ZTP реагируют на ZMP и ZTP , которые являются побочными продуктами метаболизма пуринов de novo, когда уровни 10-формилтетрагидрофолата низкие.
Предполагаемые рибопереключатели:
- мотив Moco РНК Предполагается, что связывает кофактор молибдена , регулирует гены, участвующие в биосинтезе и транспорте этого кофермента, а также ферменты, которые используют его или его производные в качестве кофактора.
Галерея изображений вторичной структуры | |
|---|---|
|



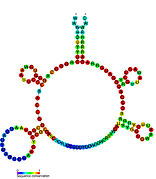

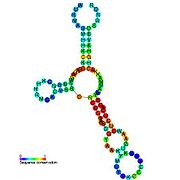
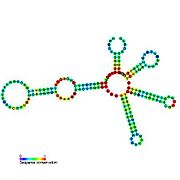
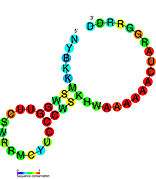


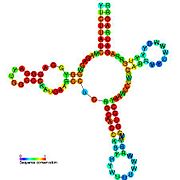
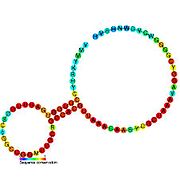
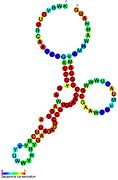

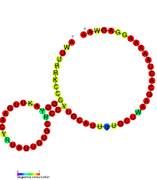


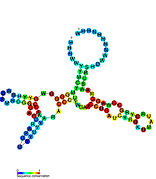



Рибопереключатели-кандидаты, связывающие метаболиты, были идентифицированы с помощью биоинформатики и имеют умеренно сложную вторичную структуру и несколько высококонсервативных положений нуклеотидов , поскольку эти особенности типичны для рибопереключателей, которые должны специфически связывать небольшую молекулу. Кандидаты на рибопереключатели также постоянно располагаются в 5'-UTR генов, кодирующих белки, и эти гены позволяют предположить связывание метаболитов, поскольку они также являются характеристиками большинства известных рибопереключателей. Гипотетические кандидаты на рибопереключатели, полностью соответствующие предыдущим критериям, следующие: мотив РНК crcB , мотив РНК manA , мотив РНК pfl , лидер ydaO/yuaA , мотив РНК yjdF , лидер ykkC-yxkD (и родственный мотив РНК ykkC-III) и yybP. -йкоЙ лидер . Функции этих гипотетических рибопереключателей остаются неизвестными.
Вычислительные модели [ править ]
Рибопереключатели также исследовались с использованием методов in silico. [29] [30] [31] В частности, решения для предсказания рибопереключателей можно разделить на две большие категории:
- искатели генов рибопереключателей , то есть системы, нацеленные на обнаружение рибопереключателей посредством геномных проверок, в основном основанных на механизмах поиска мотивов. В эту группу входит Infernal, основной компонент базы данных Rfam . [32] и более конкретные инструменты, такие как RibEx. [33] или РибоСВ. [34]
- предикторы конформационного переключения , то есть методы, основанные на структурной классификации альтернативных структур, таких как paRNAss, [35] РНКформы [36] и РНАбор. [37] Кроме того, были предложены специфичные для семейства подходы к предсказанию структуры ВКЛ/ВЫКЛ. [38]
Инструмент СвиСпот [39] каким-то образом охватывает обе группы, поскольку использует конформационные предсказания для оценки присутствия рибопереключателей.
Гипотеза мира РНК
Рибопереключатели демонстрируют, что встречающаяся в природе РНК может специфически связывать небольшие молекулы — способность, которая, как многие ранее считали, была свойственна белкам или искусственно сконструированным РНК, называемым аптамерами . Таким образом, существование рибопереключателей во всех сферах жизни добавляет некоторую поддержку гипотезе мира РНК , согласно которой жизнь изначально существовала с использованием только РНК, а белки появились позже; эта гипотеза требует, чтобы все критические функции, выполняемые белками (включая связывание малых молекул), могли выполняться РНК. Было высказано предположение, что некоторые рибопереключатели могут представлять собой древние регуляторные системы или даже остатки рибозимов мира РНК , чьи связывающие домены консервативны. [13] [18] [40]
качестве мишени антибиотиков для В
Рибопереключатели могут стать мишенью для новых антибиотиков . Действительно, было показано, что некоторые антибиотики, механизм действия которых был неизвестен на протяжении десятилетий, действуют путем воздействия на рибопереключатели. [41] Например, когда антибиотик пиритиамин попадает в клетку, он метаболизируется в пирофосфат пиритиамина. Было показано, что пиритиаминпирофосфат связывает и активирует рибопереключатель TPP, заставляя клетку прекращать синтез и импорт TPP. Поскольку пиритиаминпирофосфат не заменяет TPP в качестве кофермента, клетка погибает.
Инженерные рибопереключатели [ править ]
Поскольку рибопереключатели являются эффективным методом контроля экспрессии генов в природных организмах, возник интерес к созданию искусственных рибопереключателей. [42] [43] [44] для промышленных и медицинских применений, таких как генная терапия . [45] [46]
См. также [ править ]
- РНК-термометр — еще один класс регуляторных сегментов мРНК, которые меняют конформацию в ответ на колебания температуры, тем самым обнажая или закрывая сайт связывания рибосомы.
Ссылки [ править ]
- ^ Нудлер Э., Миронов А.С. (январь 2004 г.). «Рибопереключатель контроля бактериального метаболизма». Тенденции биохимических наук . 29 (1): 11–17. дои : 10.1016/j.tibs.2003.11.004 . ПМИД 14729327 .
- ^ Такер Би Джей, Брейкер Р.Р. (июнь 2005 г.). «Рибопереключатели как универсальные элементы контроля генов». Современное мнение в области структурной биологии . 15 (3): 342–348. дои : 10.1016/j.sbi.2005.05.003 . ПМИД 15919195 .
- ^ Витрещак А.Г., Родионов Д.А., Миронов А.А., Гельфанд М.С. (январь 2004 г.). «Рибопереключатели: старейший механизм регуляции экспрессии генов?». Тенденции в генетике . 20 (1): 44–50. CiteSeerX 10.1.1.312.9100 . дои : 10.1016/j.tig.2003.11.008 . ПМИД 14698618 .
- ^ Бати RT (июнь 2006 г.). «Структуры регуляторных элементов мРНК». Современное мнение в области структурной биологии . 16 (3): 299–306. дои : 10.1016/j.sbi.2006.05.001 . ПМИД 16707260 .
- ↑ Перейти обратно: Перейти обратно: а б Нахви А., Сударсан Н., Эберт М.С., Зоу Х, Браун К.Л., Брейкер Р.Р. (сентябрь 2002 г.). «Генетический контроль с помощью мРНК, связывающей метаболиты» . Химия и биология . 9 (9): 1043–1049. дои : 10.1016/S1074-5521(02)00224-7 . ПМИД 12323379 .
- ^ Сударсан Н., Баррик Дж.Э., Брейкер Р.Р. (июнь 2003 г.). «Метаболит-связывающие домены РНК присутствуют в генах эукариот» . РНК . 9 (6): 644–647. дои : 10.1261/rna.5090103 . ПМК 1370431 . ПМИД 12756322 .
- ^ Ноу X, Каднер Р.Дж. (июнь 2000 г.). «Аденозилкобаламин ингибирует связывание рибосом с РНК btuB» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (13): 7190–7195. Бибкод : 2000PNAS...97.7190N . дои : 10.1073/pnas.130013897 . ПМК 16521 . ПМИД 10852957 .
- ↑ Перейти обратно: Перейти обратно: а б Гельфанд М.С., Миронов А.А., Йомантас Ю., Козлов Ю.И., Перумов Д.А. (ноябрь 1999 г.). «Консервативный структурный элемент РНК, участвующий в регуляции генов синтеза бактериального рибофлавина». Тенденции в генетике . 15 (11): 439–442. дои : 10.1016/S0168-9525(99) 01856-9 ПМИД 10529804 .
- ↑ Перейти обратно: Перейти обратно: а б Миранда-Риос Х, Наварро М, Соберон М (август 2001 г.). «Консервативная структура РНК (ти-бокс) участвует в регуляции экспрессии генов биосинтеза тиамина у бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (17): 9736–9741. дои : 10.1073/pnas.161168098 . ПМК 55522 . ПМИД 11470904 .
- ^ Стормо Г.Д., Джи Й (август 2001 г.). «Действуют ли мРНК в качестве прямых сенсоров малых молекул, контролируя их экспрессию?» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (17): 9465–9467. Бибкод : 2001PNAS...98.9465S . дои : 10.1073/pnas.181334498 . ПМЦ 55472 . ПМИД 11504932 .
- ^ Голд Л, Браун Д, Хэ Ю, Штатланд Т, Сингер БС, Ву Ю (январь 1997 г.). «От форм олигонуклеотидов к геномному SELEX: новые биологические регуляторные петли» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (1): 59–64. Бибкод : 1997ПНАС...94...59Г . дои : 10.1073/pnas.94.1.59 . ЧВК 19236 . ПМИД 8990161 .
- ^ Миронов А.С., Гусаров И., Рафиков Р., Лопес Л.Е., Шаталин К., Кренева Р.А., Перумов Д.А., Нудлер Е (ноябрь 2002 г.). «Ощущение малых молекул с помощью зарождающейся РНК: механизм контроля транскрипции у бактерий» . Ячейка 111 (5): 747–756. дои : 10.1016/S0092-8674(02) 01134-0 ПМИД 12464185 .
- ↑ Перейти обратно: Перейти обратно: а б Винклер В., Нахви А., Брейкер Р.Р. (октябрь 2002 г.). «Производные тиамина напрямую связывают информационные РНК, регулируя экспрессию бактериальных генов». Природа . 419 (6910): 952–956. Бибкод : 2002Natur.419..952W . дои : 10.1038/nature01145 . ПМИД 12410317 . S2CID 4408592 .
- ^ Винклер В.К., Коэн-Чаламиш С., Брейкер Р.Р. (декабрь 2002 г.). «Структура мРНК, которая контролирует экспрессию генов путем связывания FMN» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (25): 15908–15913. Бибкод : 2002PNAS...9915908W . дои : 10.1073/pnas.212628899 . ПМК 138538 . ПМИД 12456892 .
- ^ Гранди Ф.Дж., Хенкин Т.М. (ноябрь 1998 г.). «S-бокс-регулон: новая глобальная система контроля терминации транскрипции генов биосинтеза метионина и цистеина в грамположительных бактериях» . Молекулярная микробиология . 30 (4): 737–749. дои : 10.1046/j.1365-2958.1998.01105.x . ПМИД 10094622 .
- ^ Франклунд К.В., Каднер Р.Дж. (июнь 1997 г.). «Множественные транскрибируемые элементы контролируют экспрессию гена btuB Escherichia coli» . Журнал бактериологии . 179 (12): 4039–4042. дои : 10.1128/jb.179.12.4039-4042.1997 . ПМК 179215 . ПМИД 9190822 .
- ^ Баррик Дж.Э., Корбино К.А., Винклер В.К., Нави А., Мандал М., Коллинз Дж., Ли М., Рот А., Сударсан Н., Йона И., Викайзер Дж.К., Брейкер Р.Р. (апрель 2004 г.). «Новые мотивы РНК предполагают расширение возможностей рибопереключателей в генетическом контроле бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (17): 6421–6426. Бибкод : 2004PNAS..101.6421B . дои : 10.1073/pnas.0308014101 . ПМК 404060 . ПМИД 15096624 .
- ↑ Перейти обратно: Перейти обратно: а б Корбино К.А., Баррик Дж.Э., Лим Дж., Велц Р., Такер Б.Дж., Пускарц И., Мандал М., Рудник Н.Д., Брейкер Р.Р. (2005). «Доказательства существования второго класса S-аденозилметиониновых рибопереключателей и других регуляторных мотивов РНК у альфа-протеобактерий» . Геномная биология . 6 (8): 70 рандов. дои : 10.1186/gb-2005-6-8-r70 . ПМЦ 1273637 . ПМИД 16086852 .
- ^ Вайнберг З., Баррик Дж.Э., Яо З., Рот А., Ким Дж.Н., Гор Дж., Ван Дж.К., Ли Э.Р., Блок К.Ф., Сударсан Н., Неф С., Томпа М., Руццо В.Л., Брейкер Р.Р. (2007). «Идентификация 22 структурированных РНК-кандидатов в бактериях с использованием конвейера сравнительной геномики CMfinder» . Исследования нуклеиновых кислот . 35 (14): 4809–4819. дои : 10.1093/нар/gkm487 . ПМК 1950547 . ПМИД 17621584 .
- ^ Вайнберг З., Ван Дж. И., Бог Дж., Ян Дж., Корбино К., Мой Р. Х., Брейкер Р. Р. (март 2010 г.). «Сравнительная геномика выявила 104 кандидата в структурированные РНК из бактерий, архей и их метагеномов» . Геномная биология . 11 (3): С31. дои : 10.1186/gb-2010-11-3-r31 . ПМЦ 2864571 . ПМИД 20230605 .
- ^ Чеа М.Т., Вахтер А., Сударсан Н., Брейкер Р.Р. (май 2007 г.). «Контроль альтернативного сплайсинга РНК и экспрессии генов с помощью эукариотических рибопереключателей». Природа . 447 (7143): 497–500. Бибкод : 2007Natur.447..497C . дои : 10.1038/nature05769 . ПМИД 17468745 . S2CID 4393918 .
- ^ Вахтер А., Тунч-Оздемир М., Гроув BC, Грин П.Дж., Шинтани Д.К., Брейкер Р.Р. (ноябрь 2007 г.). «Рибосвич-контроль экспрессии генов в растениях путем сплайсинга и альтернативного 3'-концевого процессинга мРНК» . Растительная клетка . 19 (11): 3437–3450. дои : 10.1105/tpc.107.053645 . ПМК 2174889 . ПМИД 17993623 .
- ^ Бокобза С., Адато А., Мандель Т., Шапира М., Нудлер Э., Ахарони А. (ноябрь 2007 г.). «Рибосвич-зависимая регуляция генов и ее эволюция в царстве растений» . Гены и развитие . 21 (22): 2874–2879. дои : 10.1101/gad.443907 . ПМК 2049190 . ПМИД 18006684 .
- ^ Андре Г., Эвен С., Путцер Х., Бургьер П., Кру С., Данчин А., Мартин-Верстраете И., Сутурина О. (октябрь 2008 г.). «Рибопереключатели S-box и T-box и антисмысловая РНК контролируют оперон метаболизма серы Clostridium acetobutylicum» . Исследования нуклеиновых кислот . 36 (18): 5955–5969. дои : 10.1093/нар/gkn601 . ПМК 2566862 . ПМИД 18812398 .
- ^ Ло Э., Дюссурже О., Грипенланд Дж., Вайткявичюс К., Тьенсуу Т., Мандин П., Репойла Ф., Бухрисер К., Коссарт П., Йоханссон Дж. (ноябрь 2009 г.). «Транс-действующий рибопереключатель контролирует экспрессию регулятора вирулентности PrfA в Listeria monocytogenes» . Клетка . 139 (4): 770–779. дои : 10.1016/j.cell.2009.08.046 . ПМИД 19914169 .
- ^ «Команда:BYU Provo/Results — 2011.igem.org» . 2011.igem.org .
- ^ Шерман Э.М., Эскиаки Дж., Эльсайед Дж., Йе Дж.Д. (март 2012 г.). «Энергетически выгодное взаимодействие лидер-линкер отменяет кооперативность связывания лигандов в глициновых рибопереключателях» . РНК . 18 (3): 496–507. дои : 10.1261/rna.031286.111 . ПМЦ 3285937 . ПМИД 22279151 .
- ^ Бокобза С.Е., Ахарони А. (октябрь 2008 г.). «Включение света на растительных рибопереключателях». Тенденции растениеводства . 13 (10): 526–33. Бибкод : 2008TPS....13..526B . doi : 10.1016/j.tplants.2008.07.004 . ПМИД 18778966 .
- ^ Баррик Дж. Э. (2009). «Прогнозирование регуляции рибопереключателя в геномном масштабе». Рибопереключатели . Методы молекулярной биологии. Том. 540. стр. 1–13. дои : 10.1007/978-1-59745-558-9_1 . ISBN 978-1-934115-88-6 . ПМИД 19381548 .
- ^ Бараш Д., Габданк I (январь 2010 г.). «Минимизация энергии применительно к рибопереключателям: перспективы и проблемы» . Биология РНК . 7 (1): 90–97. дои : 10.4161/rna.7.1.10657 . ПМИД 20061789 .
- ^ Чен, Ши-Цзе; Берк, Дональд Х; Адамяк, RW (2015). Вычислительные методы понимания рибопереключателей / Методы энзимологии, том 553 . Академическая пресса. ISBN 978-0-12-801618-3 .
- ^ Навроцкий Е.П., Кольбе Д.Л., Эдди С.Р. (май 2009 г.). «Инфернал 1.0: вывод о выравнивании РНК» . Биоинформатика . 25 (10): 1335–1337. doi : 10.1093/биоинформатика/btp157 . ПМЦ 2732312 . ПМИД 19307242 .
- ^ Абреу-Гуджер С., Мерино Е (июль 2005 г.). «RibEx: веб-сервер для поиска рибопереключателей и других консервативных бактериальных регуляторных элементов» . Исследования нуклеиновых кислот . 33 (проблема с веб-сервером): W690-2. дои : 10.1093/nar/gki445 . ПМК 1160206 . ПМИД 15980564 .
- ^ Чанг Т.Х., Хуан Х.Д., Ву Л.К., Йе КТ, Лю Б.Дж., Хорнг Дж.Т. (июль 2009 г.). «Вычислительная идентификация рибопереключателей на основе консервативных функциональных последовательностей и конформаций РНК» . РНК . 15 (7): 1426–1430. дои : 10.1261/rna.1623809 . ПМК 2704089 . ПМИД 19460868 .
- ^ Восс Б., Мейер С., Гигерих Р. (июль 2004 г.). «Оценка предсказуемости конформационного переключения РНК» . Биоинформатика . 20 (10): 1573–1582. doi : 10.1093/биоинформатика/bth129 . ПМИД 14962925 .
- ^ Янссен С., Гигерих Р. (февраль 2015 г.). «Студия РНК форм» . Биоинформатика . 31 (3): 423–425. doi : 10.1093/биоинформатика/btu649 . ПМК 4308662 . ПМИД 25273103 .
- ^ Фрейхульт Э., Моултон В., Клот П. (август 2007 г.). «Больцмановская вероятность структурных соседей РНК и обнаружение рибопереключателя» . Биоинформатика . 23 (16): 2054–2062. doi : 10.1093/биоинформатика/btm314 . ПМИД 17573364 .
- ^ Клот П., Лу Ф., Лоренц В.А. (апрель 2012 г.). «Максимально ожидаемая точность структурных соседей вторичной структуры РНК» . БМК Биоинформатика . 13 (Приложение 5): S6. дои : 10.1186/1471-2105-13-S5-S6 . ПМК 3358666 . ПМИД 22537010 .
- ^ Барсакки М., Новоа Э.М., Келлис М., Бекини А. (ноябрь 2016 г.). «SwiSpot: моделирование рибопереключателей путем выявления последовательностей переключения» . Биоинформатика . 32 (21): 3252–3259. doi : 10.1093/биоинформатика/btw401 . hdl : 11568/817190 . ПМИД 27378291 .
- ^ Кокрейн Дж.К., Strobel SA (июнь 2008 г.). «Эффекторы рибопереключателя как кофакторы белковых ферментов» . РНК . 14 (6): 993–1002. дои : 10.1261/rna.908408 . ПМК 2390802 . ПМИД 18430893 .
- ^ Блаунт К.Ф., Брейкер Р.Р. (декабрь 2006 г.). «Рибопереключатели как мишени антибактериальных препаратов». Природная биотехнология . 24 (12): 1558–1564. дои : 10.1038/nbt1268 . ПМИД 17160062 . S2CID 34398395 .
- ^ Бауэр Г., Зюсс Б. (июнь 2006 г.). «Спроектированные рибопереключатели как новые инструменты молекулярной биологии». Журнал биотехнологии . 124 (1): 4–11. doi : 10.1016/j.jbiotec.2005.12.006 . ПМИД 16442180 .
- ^ Диксон Н., Дункан Дж. Н., Гирлингс Т., Данстан М. С., Маккарти Дж. Э., Лейс Д., Миклфилд Дж. (февраль 2010 г.). «Реинжиниринг ортогонально-селективных рибопереключателей» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (7): 2830–2835. Бибкод : 2010PNAS..107.2830D . дои : 10.1073/pnas.0911209107 . ПМЦ 2840279 . ПМИД 20133756 .
- ^ Верхуниг А., Керхер Д., Бок Р. (апрель 2010 г.). «Индуцируемая экспрессия генов из пластидного генома с помощью синтетического рибопереключателя» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (14): 6204–6209. Бибкод : 2010PNAS..107.6204V . дои : 10.1073/pnas.0914423107 . ПМК 2852001 . ПМИД 20308585 .
- ^ Кетцер П., Кауфманн Дж.К., Энгельхардт С., Боссов С., фон Калле С., Хартиг Дж.С., Унгерехтс Г., Неттельбек Д.М. (февраль 2014 г.). «Искусственные рибопереключатели для экспрессии генов и контроля репликации ДНК и РНК-вирусов» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (5): E554–562. Бибкод : 2014PNAS..111E.554K . дои : 10.1073/pnas.1318563111 . ПМЦ 3918795 . ПМИД 24449891 .
- ^ Штробель Б., Клаузер Б., Хартиг Дж.С., Ламла Т., Гантнер Ф., Кройц С. (октябрь 2015 г.). «Опосредованное рибопереключателем ослабление трансгенной цитотоксичности увеличивает выход аденоассоциированного вирусного вектора в клетках HEK-293» . Молекулярная терапия . 23 (10): 1582–1591. дои : 10.1038/mt.2015.123 . ПМЦ 4817922 . ПМИД 26137851 .
Дальнейшее чтение [ править ]
- Ферре-Д'Амаре, Адриан Р.; Винклер, Уэйд К. (2011). «Глава 5. Роль ионов металлов в регуляции рибопереключателями». В Астрид Сигель, Хельмут Сигель и Роланд К.О. Сигель (ред.). Структурная и каталитическая роль ионов металлов в РНК . Ионы металлов в науках о жизни. Том. 9. Кембридж, Великобритания: Издательство RSC. стр. 141–173. дои : 10.1039/9781849732512-00141 . ISBN 978-1-84973-094-5 . ПМЦ 3454353 . ПМИД 22010271 .
| Базы данных органов управления : Национальные |
|---|