Рибопереключатель PreQ1
| Рибопереключатель PreQ1 | |
|---|---|
 Прогнозируемая вторичная структура и сохранение последовательности PreQ1 | |
| Идентификаторы | |
| Символ | Пред1 квартал |
| Рфам | RF00522 |
| Другие данные | |
| РНК Тип | Цис-рег ; рибопереключатель |
| Домен(ы) | Бактерии |
| ТАК | ТАК: 0000035 |
| PDB Структуры | ПДБе 2Л1В |
Рибопереключатель PreQ 1 -I представляет собой цис-действующий элемент, идентифицированный у бактерий , который регулирует экспрессию генов, участвующих в ( Q ) биосинтезе нуклеозида кевозина из GTP . [ 1 ] PreQ 1 (пре-кеуозин 1 ) является промежуточным звеном в пути кевозина, а рибопереключатель preQ 1 , как тип рибопереключателя , представляет собой элемент РНК, который связывает preQ 1 . Рибопереключатель preQ 1 отличается необычно маленьким аптамером по сравнению с другими рибопереключателями. Была определена его трехмерная структура с атомным разрешением с идентификатором PDB 2L1V . [ 2 ] [ 3 ]
PreQ 1 Классификация
[ редактировать ]Существуют три подкатегории рибопереключателя PreQ 1 : preQ 1 -I, preQ 1 -II и preQ 1 -III. PreQ 1 -I имеет явно небольшой аптамер длиной от 25 до 45 нуклеотидов. [ 4 ] по сравнению со структурами рибопереключателя PreQ 1 -II и preQ 1 рибопереключателя -III . Рибопереключатель PreQ 1 -II, обнаруженный только у Lactobacillales , имеет более крупную и сложную консенсусную последовательность и структуру, чем рибопереключатель preQ 1 -I, со средним числом 58 нуклеотидов, составляющих его аптамер, который образует целых пять подструктур спаренных оснований. [ 5 ] Рибопереключатель PreQ 1 -III имеет четкую структуру, а также имеет больший размер аптамера, чем рибопереключатель preQ 1 -I, в диапазоне размеров от 33 до 58 нуклеотидов. Рибопереключатель PreQ 1 -III имеет атипично организованный псевдоузел , который, по-видимому, не включает свою нижестоящую платформу экспрессии в сайте связывания рибосомы (RBS). [ 6 ]
История
[ редактировать ]PreQ 1 был впервые обнаружен как антикодоновая последовательность тРНК E.coli в 1972 году. [ 7 ] Рибопереключатель preQ1 не был впервые обнаружен до 2004 года. [ 8 ] и признан даже позже. [ 9 ] Первый зарегистрированный рибопереключатель preQ 1 был расположен в лидере оперона Bacillus subtilis ykvJKLM (queCDEF), который кодирует четыре гена, необходимые для производства кевозина. [ 8 ] Считается , что в этом организме связывание PreQ 1 рибопереключателя с аптамером вызывает преждевременное прекращение транскрипции внутри лидера, что снижает экспрессию этих генов. Позднее рибопереключатель preQ1 был идентифицирован как консервативная последовательность на 5'-UTR генов многих грамположительных бактерий и было доказано, что он связан с синтезом preQ 1 . [ 9 ]
В 2008 году второй класс рибопереключателей preQ 1 ( рибопереключатели PreQ1-II ) также был обнаружен как представитель мотива РНК COG4708 из Streptococcus pneumoniae R6. [ 10 ] Хотя PreQ 1 рибопереключатель -II также работает как промежуточный продукт биосинтеза кевозина, характеристики структурного и молекулярного распознавания отличаются от рибопереключателя preQ1-I, что указывает на то, что природные аптамеры, использующие разные структуры для связывания одного и того же метаболита, могут быть более распространены, чем известно в настоящее время. [ 10 ]
Структура и функции
[ редактировать ]

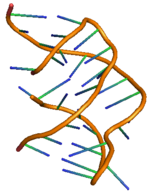
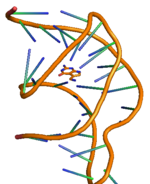
Рибопереключатель PreQ1 имеет два стебля и три петли, его подробная структура показана справа. [ 11 ] Рибопереключающее действие рибопереключателей preQ 1 у бактерий регулируется путем связывания метаболита preQ 1 с аптамерной областью, что приводит к структурным изменениям в информационной РНК (мРНК), которая управляет последующей генетической регуляцией. [ 12 ] Структура рибопереключателя preQ 1 имеет компактный псевдоузел H-типа , что существенно отличает его от других рибопереключателей на основе пурина . [ 12 ] preQ 1 Лиганд похоронен в ядре псевдоузла и стабилизирован за счет интеркаляции между спиральными стопками и взаимодействия водородных связей с гетероатомами. В отсутствие preQ1 область хвоста P2 находится вдали от области петли P2, и, следовательно, рибопереключатель находится в расстыкованном (частично пристыкованном) состоянии, тогда как при связывании preQ 1 с рибопереключателем две области P2 сближаются. вызывая полную стыковку рибопереключателя. Наблюдается , что этот механизм стыковки и расстыковки рибопереключателя с изменением концентрации лиганда preQ 1 контролирует передачу сигналов регуляции генов, широко известную как передача сигналов «ВКЛ» или «ВЫКЛ» для экспрессии генов. [ 11 ] [ 13 ] Наблюдается, что на механизм стыковки и расстыковки влияет не только лиганд, но и другие факторы, такие как соль Mg. [ 14 ] Как и любой другой рибопереключатель, два наиболее распространенных типа регуляции генов, опосредованных рибопереключателем preQ 1, связаны с ослаблением транскрипции или ингибированием инициации трансляции . Связывание лиганда с транскрипционным рибопереключателем у бактерий вызывает модификацию структуры рибопереключателя, что приводит к затруднению активности РНК-полимеразы, вызывая ослабление транскрипции . Аналогично, связывание лиганда с трансляционным рибопереключателем вызывает модификацию вторичной структуры единицы рибопереключателя, что приводит к затруднению связывания рибосомы и, следовательно, к ингибированию инициации трансляции .
Транскрипционная регуляция
[ редактировать ]Ослабление транскрипции, опосредованное PreQ 1, контролируется динамическим переключением антитерминатора и терминаторной шпильки в рибопереключателе. [ 11 ] Предполагается, что для рибопереключателя preQ 1 из бактерий Bacillus subtilis (Bsu) антитерминатор будет менее стабильным, чем терминатор, поскольку добавление preQ 1 значительно смещает равновесие в сторону образования терминатора. [ 11 ] В присутствии preQ 1 3'-конец богатого аденином хвостового домена соединяется с центром шпильки P1, образуя псевдоузел H-типа . [ 11 ] В нативной структуре мРНК связывание preQ 1 с аптамерной областью рибопереключателя приводит к образованию терминаторной шпильки, которая заставляет РНК-полимеразу останавливать транскрипцию, процесс, который широко известен как OFF-регуляция генетической экспрессии или терминация транскрипции. [ 13 ]
Регламент перевода
[ редактировать ]Трансляция белка у прокариот инициируется связыванием 30S рибосомальной субъединицы с последовательностью Шайна-Дальгарно (SD) в мРНК . Опосредованное PreQ 1 ингибирование регуляции трансляции контролируется путем блокирования -Дальгарно последовательности мРНК Шайна для предотвращения связывания рибосомы с мРНК для трансляции . Связывание preQ1 с доменом аптамера способствует секвестрации части последовательности SD на 5'-конце стебля P2 домена аптамера, вызывая недоступность последовательности SD. [ 11 ] Трансляционный рибопереключатель бактерий Thermoanaerobacter tengcongensis ( Tte ) временно закрывается (предварительно стыковывается) в отсутствие preQ 1 , тогда как в присутствии preQ 1 принимается полностью стыкованное состояние. Это равновесие стыковки/расстыковки регулируется не только концентрацией лиганда, но и концентрацией соли Mg. [ 14 ] [ 15 ] Недоступность последовательности SD из-за образования псевдоузла в присутствии preQ 1 указывает на OFF-регуляцию генетической экспрессии в трансляционном рибопереключателе или ингибирование инициации трансляции.
Физиологическое значение в регуляции бактериальных генов
[ редактировать ]PreQ 1 Активность рибопереключателя у бактерий Tte можно измерить по уровням двух белков, находящихся в кодирующей области мРНК Tte , а именно TTE1564 и TTE1563. [ 16 ] Белки, расположенные ниже рибопереключателя preQ1 , биосинтезируют азотистое основание, называемое квеином , и нуклеозид кевозин ингибируется активацией рибопереключателя preQ1. Кеуин участвует в антикодоновой последовательности некоторых тРНК . [ 17 ] У бактерий гипермодифицированное нуклеиновое основание хвеин занимает первое положение антикодона или его колебательное положение в тРНК аспарагина , аспарагиновой кислоты , гистидина и тирозина . [ 18 ] У бактерий фермент тРНК-гуанинтрансгликозилаза (TGT) катализирует замену гуанина в положении 34 тРНК на квеин в первом положении аниткодона. [ 15 ] [ 16 ] Эукарии включают квеин в РНК, тогда как эубактерии включают preQ 1 , который затем подвергается модификации с образованием квеина. [ 17 ] Поскольку кевозин вырабатывается исключительно бактериями, эукариотические организмы должны получать запасы кевозина или его нуклеинового основания кевина из своего рациона, а бактерии - из кишечной микрофлоры . Последствием дефицита кевина или кевозина является неспособность вырабатывать модифицированную кевозином тРНК и, кроме того, неспособность клетки превращать фенилаланин в тирозин . [ 19 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Рот А., Винклер В.К., Регулски Э.Э., Ли Б.В., Лим Дж., Джона И., Баррик Дж.Е., Ритвик А., Ким Дж.Н., Велц Р., Ивата-Рейл Д., Брейкер Р.Р. (2007). «Рибосвитч, селективный в отношении предшественника кевозина preQ1, содержит необычно маленький аптамерный домен» . Nat Struct Мол Биол . 14 (4): 308–317. дои : 10.1038/nsmb1224 . ПМИД 17384645 . S2CID 32367656 .
- ^ Кляйн DJ, Эдвардс Т.Э., Ферре-Д'Амаре А.Р. (март 2009 г.). «Кокристаллическая структура рибопереключателя preQ1 класса I обнаруживает псевдоузел, распознающий существенное гипермодифицированное нуклеиновое основание» . Нат. Структура. Мол. Биол . 16 (3): 343–344. дои : 10.1038/nsmb.1563 . ПМЦ 2657927 . ПМИД 19234468 .
- ^ Канг М., Петерсон Р., Фейгон Дж. (март 2009 г.). «Структурный взгляд на рибопереключатель биосинтеза кевозина, модифицированного нуклеотида, обнаруженного в антикодоне тРНК» . Мол. Клетка . 33 (6): 784–790. doi : 10.1016/j.molcel.2009.02.019 . ПМИД 19285444 .
- ^ «РИБОВЫКЛЮЧАТЕЛИ: КЛАССИФИКАЦИЯ, ФУНКЦИИ И ПОДХОД INSILICO». Международный журнал фармацевтических наук и исследований (IJPSR) . 1 (9): 414. 2010.
- ^ МакКаун, ПиДжей; Лян, Джей-Джей; Вайнберг, З; Брейкер, Р.Р. (17 июля 2014 г.). «Структурное, функциональное и таксономическое разнообразие трех классов рибопереключателей PreQ1» . Химия и биология . 21 (7): 880–889. doi : 10.1016/j.chembiol.2014.05.015 . ПМЦ 4145258 . ПМИД 25036777 .
- ^ Либерман, Джозеф А.; Суддала, Кришна К.; Айтенфису, Асаминев; Чан, Дален; Белашов Иван А.; Салим, Мохаммед; Мэтьюз, Дэвид Х.; Спитале, Роберт С.; Вальтер, Нильс Г. (07 июля 2015 г.). «Структурный анализ рибопереключателя preQ1 класса III выявил аптамер, удаленный от сайта связывания рибосомы, регулируемого быстрой динамикой» . Труды Национальной академии наук . 112 (27): E3485–E3494. дои : 10.1073/pnas.1503955112 . ISSN 0027-8424 . ПМК 4500280 . ПМИД 26106162 .
- ^ Харада, Фумио; Нисимура, Сусуму (январь 1972 г.). «Возможные антикодонные последовательности тРНКГис, тРНКАсн и тРНКАсп из Escherichia coli. Универсальное присутствие нуклеозида О в первом положении антикодонов этих переносчиков рибонуклеиновой кислоты». Биохимия . 11 (2): 301–308. дои : 10.1021/bi00752a024 . ПМИД 4550561 .
- ^ Jump up to: а б Ридер Дж.С., Мецгар Д., Шиммель П., де Креси-Лагард В. (2004). «Идентификация четырех генов, необходимых для биосинтеза модифицированного нуклеозида кевозина» . Ж. Биол. Хим . 279 (8): 6280–6285. дои : 10.1074/jbc.M310858200 . ПМИД 14660578 .
- ^ Jump up to: а б Рот, Адам; Винклер, Уэйд К.; Регульски, Элизабет Э.; Ли, Бобби В.К.; Лим, Джинсу; Йона, Инбал; Баррик, Джеффри Э.; Ритвик, Анкита; Ким, Джейн Н. (1 апреля 2007 г.). «Рибопереключатель, селективный в отношении предшественника кевозина preQ1, содержит необычно маленький аптамерный домен». Структурная и молекулярная биология природы . 14 (4): 308–317. дои : 10.1038/nsmb1224 . ISSN 1545-9993 . ПМИД 17384645 . S2CID 32367656 .
- ^ Jump up to: а б Мейер, Мишель М.; Рот, Адам; Червин, Стефани М.; Гарсия, Джордж А.; Брейкер, Рональд Р. (1 апреля 2008 г.). «Подтверждение второго природного класса аптамеров preQ1 у бактерий Streptococcaceae» . РНК . 14 (4): 685–695. дои : 10.1261/rna.937308 . ISSN 1355-8382 . ПМК 2271366 . ПМИД 18305186 .
- ^ Jump up to: а б с д и ж Эйххорн, Кэтрин Д.; Кан, Миджон; Фейгон, Джули (01 октября 2014 г.). «Структура и функции рибопереключателей preQ1» . Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1839 (10): 939–950. дои : 10.1016/j.bbagrm.2014.04.019 . ПМК 4177978 . ПМИД 24798077 .
- ^ Jump up to: а б Серганов, Александр; Патель, Диншоу Дж. (11 мая 2012 г.). «Принципы распознавания метаболитов и молекулярные механизмы, лежащие в основе функции рибопереключателя» . Ежегодный обзор биофизики . 41 (1): 343–370. doi : 10.1146/annurev-biophys-101211-113224 . ISSN 1936-122Х . ПМЦ 4696762 . ПМИД 22577823 .
- ^ Jump up to: а б Ридер, Ульрике; Крейц, Кристоф; Микура, Рональд (15 июня 2010 г.). «Складывание транскрипционно действующего рибопереключателя PreQ1» . Труды Национальной академии наук . 107 (24): 10804–10809. дои : 10.1073/pnas.0914925107 . ISSN 0027-8424 . ПМК 2890745 . ПМИД 20534493 .
- ^ Jump up to: а б Суддала, Кришна К.; Ринальди, Арли Дж.; Фэн, Цзюнь; Мусто, Энтони М.; Эйххорн, Кэтрин Д.; Либерман, Джозеф А.; Ведекинд, Джозеф Э.; Аль-Хашими, Хашим М.; Брукс, Чарльз Л. (1 декабря 2013 г.). «Отдельные транскрипционные и трансляционные рибопереключатели preQ1 принимают сходные предварительно свернутые ансамбли, которые следуют различным путям сворачивания в одну и ту же структуру, связанную с лигандом» . Исследования нуклеиновых кислот . 41 (22): 10462–10475. дои : 10.1093/nar/gkt798 . ISSN 0305-1048 . ПМЦ 3905878 . ПМИД 24003028 .
- ^ Jump up to: а б Суддала, Кришна К.; Ван, Цзяруй; Хоу, Цянь; Вальтер, Нильс Г. (11 ноября 2015 г.). «Mg2 + сдвигает лиганд-опосредованное сворачивание рибопереключателя от индуцированного соответствия к конформационному отбору» . Журнал Американского химического общества . 137 (44): 14075–14083. дои : 10.1021/jacs.5b09740 . ISSN 0002-7863 . ПМК 5098500 . ПМИД 26471732 .
- ^ Jump up to: а б Ринальди, Арли Дж.; Лунд, Пол Э.; Бланко, Марио Р.; Вальтер, Нильс Г. (19 января 2016 г.). «Последовательность Шайна-Дальгарно одиночных мРНК, регулируемых рибопереключателями, демонстрирует лиганд-зависимые всплески доступности» . Природные коммуникации . 7 : 8976. Бибкод : 2016NatCo...7.8976R . дои : 10.1038/ncomms9976 . ISSN 2041-1723 . ПМЦ 4735710 . ПМИД 26781350 .
- ^ Jump up to: а б Киттендорф, Джеффри Д.; Сграджа, Таня; Рейтер, Клаус; Клебе, Герхард; Гарсия, Джордж А. (24 октября 2003 г.). «Основная роль аспартата 264 в катализе тРНК-гуанинтрансгликозилазой из Escherichia coli» . Журнал биологической химии . 278 (43): 42369–42376. дои : 10.1074/jbc.M304323200 . ISSN 0021-9258 . ПМИД 12909636 .
- ^ Дженкинс, Джермейн Л.; Кручинска, Иоланта; Маккарти, Рид М.; Бандарян, Ваге; Ведекинд, Джозеф Э. (15 июля 2011 г.). «Сравнение аптамера рибопереключателя PreQ1 в связанном с метаболитами и свободном состояниях с последствиями для регуляции генов» . Журнал биологической химии . 286 (28): 24626–24637. дои : 10.1074/jbc.M111.230375 . ISSN 0021-9258 . ПМК 3137038 . ПМИД 21592962 .
- ^ Ракович, Татьяна; Боланд, Койлин; Бернштейн, Илана; Чиквана, Вимбай М.; Ивата-Рейл, Дирк; Келли, Винсент П. (3 июня 2011 г.). «Дефицит кевозина у эукариот ставит под угрозу выработку тирозина за счет повышенного окисления тетрагидробиоптерина» . Журнал биологической химии . 286 (22): 19354–19363. дои : 10.1074/jbc.M111.219576 . ISSN 0021-9258 . ПМК 3103313 . ПМИД 21487017 .