Псевдоузел
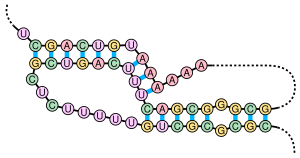

Псевдоузел содержащую представляет собой вторичную структуру нуклеиновой кислоты, по крайней мере две структуры «стебель-петля» , в которых половина одного стебля вставлена между двумя половинками другого стебля. Псевдоузел был впервые обнаружен у вируса желтой мозаики репы в 1982 году. [2] Псевдоузлы складываются в узловые трехмерные конформации, но не являются настоящими топологическими узлами . Эти структуры классифицируются как топология креста (X) в рамках топологии цепей , которая, в отличие от теории узлов, представляет собой подход, основанный на контактах.
Прогнозирование и идентификация
[ редактировать ]Структурная конфигурация псевдоузлов плохо поддается биовычислительному обнаружению из-за ее контекстно-зависимой или «перекрывающейся» природы. Спаривание оснований в псевдоузлах плохо вложено; то есть встречаются пары оснований, которые «перекрываются» друг с другом в положении последовательности. Это делает наличие псевдоузлов в последовательностях РНК более трудным для предсказания с помощью стандартного метода динамического программирования , который использует рекурсивную систему оценки для идентификации парных стеблей и, следовательно, большинство из них не могут обнаружить невложенные пары оснований. Новый метод стохастических контекстно-свободных грамматик страдает от той же проблемы. Таким образом, популярные методы прогнозирования вторичной структуры, такие как Mfold и Pfold, не будут прогнозировать структуры псевдоузлов, присутствующие в последовательности запроса; они идентифицируют только более стабильный из двух стеблей псевдоузлов.
Можно идентифицировать ограниченный класс псевдоузлов с помощью динамического программирования, но эти методы не являются исчерпывающими и хуже масштабируются в зависимости от длины последовательности, чем алгоритмы без псевдоузлов. [3] [4] Было показано, что общая проблема прогнозирования структур с наименьшей свободной энергией с псевдоузлами является NP-полной . [5] [6]
Биологическое значение
[ редактировать ]Некоторые важные биологические процессы основаны на молекулах РНК, образующих псевдоузлы, которые часто представляют собой РНК с обширной третичной структурой . Например, область псевдоузла РНКазы P является одним из наиболее консервативных элементов за всю эволюцию. Компонент теломеразной РНК содержит псевдоузел, критический для активности. [1] а некоторые вирусы используют структуру псевдоузла для формирования тРНК-подобного мотива для проникновения в клетку-хозяина. [7]
Представление псевдоузлов
[ редактировать ]Существует множество типов псевдоузлов, различающихся тем, как они пересекаются и сколько раз они пересекаются. Чтобы отразить эту разницу, псевдоузлы классифицируются на H-, K-, L-, M-типы, причем каждый последующий тип добавляет слой ступенчатой интеркаляции. Например, простой пример теломеразы P2b-P3 в статье представляет собой псевдоузел H-типа. [8]
Вторичная структура РНК обычно обозначается точечными скобками с парными круглыми скобками. () обозначающие пары оснований в основе и точки, обозначающие петли. Прерванные основы псевдоузлов означают, что такие обозначения необходимо расширить дополнительными скобками или даже буквами, чтобы можно было представить различные наборы основ. Одно из таких расширений использует в порядке вложения ([{<ABCDE для открытия и edcba>}]) для закрытия. [9] Структура двух (слегка различающихся) примеров теломеразы в этих обозначениях такова:
(((.(((((........))))).))). ....]]]]]]. drawing 1 CGCGCGCUGUUUUUCUCGCUGACUUUCAGCGGGCGA---AAAAAAUGUCAGCU 50 ALIGN |.||||||||||||||||||||||||| .|.| |||||| ||||||. 1ymo 1 ---GGGCUGUUUUUCUCGCUGACUUUCAGC--CCCAAACAAAAAA-GUCAGCA 47 ((((((........)))) )).........]]]]]].
Обратите внимание, что U-выпуклость на конце обычно присутствует в теломеразной РНК. Он был удален в модели решения 1ymo для повышения стабильности псевдоузла. [10]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Чен, Дж.Л.; Грейдер, CW (7 июня 2005 г.). «Функциональный анализ структуры псевдоузла теломеразной РНК человека» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (23): 8077–9. Бибкод : 2005PNAS..102.8080C . дои : 10.1073/pnas.0502259102 . ПМЦ 1149427 . ПМИД 15849264 .
- ^ Staple DW, Butcher SE (июнь 2005 г.). «Псевдоузлы: структуры РНК с разнообразными функциями» . ПЛОС Биол . 3 (6): е213. дои : 10.1371/journal.pbio.0030213 . ПМЦ 1149493 . ПМИД 15941360 .
- ^ Ривас Э, Эдди С. (1999). «Алгоритм динамического программирования для предсказания структуры РНК, включая псевдоузлы». J Мол Биол 285 (5): 2053–2068.
- ^ Диркс, Р.М. Пирс Н.А. (2004)Алгоритм расчета вероятностей спаривания оснований нуклеиновой кислоты, включая псевдоузлы. «J Вычислительная химия». 25:1295-1304, 2004.
- ^ Люнгсё РБ, Педерсен CN. (2000). «Предсказание псевдоузла РНК в энергетических моделях». J Comput Biol 7 (3–4): 409–427.
- ^ Люнгсё, РБ (2004). Сложность прогнозирования псевдоузлов в простых моделях. Доклад, представленный на ICALP.
- ^ Плейдж К.В., Ритвельд К., Бош Л. (1985). «Новый принцип сворачивания РНК, основанный на псевдоузлах» . Нуклеиновые кислоты Рез . 13 (5): 1717–31. дои : 10.1093/нар/13.5.1717 . ПМК 341107 . ПМИД 4000943 .
- ^ Кучарик, М; Хофакер, Иллинойс; Стадлер, ПФ; Цинь, Дж (15 января 2016 г.). «Псевдоузлы в ландшафтах сворачивания РНК» . Биоинформатика . 32 (2): 187–94. doi : 10.1093/биоинформатика/btv572 . ПМК 4708108 . ПМИД 26428288 .
- ^ Анчак, М; Попенда, М; Зок, Т; Зурковский, М; Адамяк, RW; Шашнюк, М (15 апреля 2018 г.). «Новые алгоритмы для представления сложных псевдоузловых структур РНК в точечных скобках» . Биоинформатика . 34 (8): 1304–1312. doi : 10.1093/биоинформатика/btx783 . ПМК 5905660 . ПМИД 29236971 .
- ^ Теймер, Калифорния; Блуа, Калифорния; Фейгон, Дж. (4 марта 2005 г.). «Структура псевдоузла теломеразной РНК человека обнаруживает консервативные третичные взаимодействия, необходимые для функционирования» . Молекулярная клетка . 17 (5): 671–82. doi : 10.1016/j.molcel.2005.01.017 . ПМИД 15749017 .
Внешние ссылки
[ редактировать ]