ДЖЕЙД1
| ДЖЕЙД1 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | JADE1 , PHF17, семейство нефритов, палец PHD 1 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 610514 ; МГИ : 1925835 ; Гомологен : 18162 ; Генные карты : JADE1 ; ОМА : JADE1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||






JADE1 — это белок , который у человека кодируется JADE1 геном . [5] [6] [7] [8]
Семья
[ редактировать ]Небольшое семейство белков под названием «Ген апоптоза и дифференцировки» (JADE). [6] включает три члена, кодируемых отдельными генами: растительный гомеодомен-17 (PHF17, JADE1), PHF16 (JADE3) и PHF15 (JADE2). Все белки семейства JADE несут два заметных домена средней молекулы: канонический цинковый палец растительного гомеодомена (PHD) и расширенный PHD-подобный цинковый палец. Таким образом, JADE1 классифицируется как член семейства белков PHD. Известны два белковых продукта гена PHF17: полноразмерный JADE1 (JADE1L) и его сплайсинговый вариант, в котором отсутствует C-концевой фрагмент, также называемый короткой изоформой (JADE1S).
Открытие
[ редактировать ]Нагасе и др. клонировали и секвенировали 100 отдельных кДНК из библиотеки кДНК мозга плода, включая клон KIAA1807, который был обозначен PHF17. [9] Предсказанный белковый продукт этого клона, состоящий из 702 аминокислот, был подобен белку цинковых пальцев человека BR140 (BRPF1). [10] На основании анализа базы данных последовательностей исследование показало, что PHF17 может участвовать в пути управления нуклеиновыми кислотами. [9] Используя дрожжевой двухгибридный подход для поиска новых партнеров белкового продукта гена фон Хиппель-Линдау (pVHL), другое исследование идентифицировало кДНК, которая соответствует клону KIAA1807. [11] Белковый продукт этой кДНК получил название JADE1 (Jade-1, PHF17). [11] Выведенный белковый продукт кДНК JADE1 длиной 509 аминокислот был дополнительно подтвержден как физический партнер pVHL. [11] В ходе генетического скринингового исследования генов, участвующих в эмбриогенезе, был идентифицирован мышиный ортолог JADE1. [6] Это исследование впервые охарактеризовало ген JADE1 и определило новое семейство JADE. В ходе исследования были получены мыши с нокаутом гена JADE1.
Транскрипты Jade1 как у людей, так и у мышей подвергаются альтернативному сплайсингу и полиаденилированию с образованием двух основных транскриптов: полной мРНК длиной 6 т.п.н. и мРНК длиной 3,6 т.п.н. [6] Два полученных белковых продукта гена JADE1 были обозначены как JADE1S для короткой изоформы (что аналогично (3)) и JADE1L для длинной изоформы. Также обнаружено несколько второстепенных транскриптов. Анализ базы данных выявил два дополнительных паралога JADE1 и членов семейства JADE, JADE2 и JADE3. JADE3 идентичен белку E9, выявленному в более раннем независимом исследовании, которое предположило роль PHF16/JADE3/E9 в апоптозе в клетках рака молочной железы. [12]
JADE1 картирован на хромосоме 4 (4q26-q27). JADE1 консервативен, и его ортологи были обнаружены или предсказаны почти у всех многоклеточных животных. Структуру и последовательности генов, варианты, консервацию, ортологи и паралоги, филогенетическое дерево JADE1 и крупномасштабный скрининг тканевой экспрессии JADE1 можно найти в нескольких обширных базах данных ( https://www.genecards.org ; http://useast.ensembl. орг ).
Структура
[ редактировать ]Полноразмерный полипептид JADE1 несет один канонический и один расширенный домен цинковых пальцев PHD. [13] Другие домены включают N-концевой кандидатный домен PEST, энхансер полисотового домена и сигнал C-концевой ядерной локализации (NLS). [6] [11] (prosite.expasy.org). Белок JADE1 является мишенью посттрансляционных модификаций, включая фосфорилирование (рис. 1).
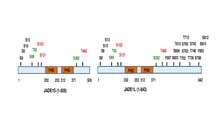
). Было идентифицировано, что шесть аминокислотных остатков фосфорилируются зависимым от клеточного цикла способом через киназы Aurora A. путь [14] [15] JADE1 является мишенью фосфорилирования казеинкиназой 2 (CK2). [16] Кроме того, с помощью высокопроизводительного скрининга и анализа in silico обнаруживаются множественные сайты фосфорилирования. Сводная схема сайтов фосфорилирования белков JADE1L и JADE1S со ссылками находится здесь. [15]
Белки, несущие тандемные канонические и расширенные пальцы PHD, образуют небольшое подсемейство внутри большого семейства белков PHD (www.gennames.org). Другие белки, несущие тандем пальцев PHD и родственные JADE1, включают белки, которые являются компонентами связывающих и модифицирующих хроматин комплексов BRPF1, BRPF3 и BRD1. [17] Кристаллическая структура доменов JADE1 PHD не решена. Канонический мотив пальца PHD имеет подпись C4HC3, представляет собой относительно небольшую, стабильную структуру и отличается от КОЛЬЦЕВОГО пальца типа C3HC4. Домены PHD способны распознавать и связывать специфический метилированный лизин гистона H3, что определяет эти домены как считыватели эпигенетического кода гистонов. [18] [19] [20] Доступны обзоры, подробно описывающие строение и свойства пальцев PHD. [21] [22] [23] [24]
Клеточная функция
[ редактировать ]Белки JADE1 многофункциональны и взаимодействуют с несколькими белками-партнерами.
Ацетилирование гистонов
[ редактировать ]В 2004 г. сообщалось о функции JADE1 в ацетилировании гистонов и активации транскрипции, которая требовала второго вытянутого цинкового пальца PHD (16). JADE1 резко увеличивает уровни ацетилированного гистона H4 в хроматине, но не гистона H3, что является специфичностью, характерной для семейства MYST HAT TIP60 и HBO1. TIP60 физически связывается с JADE1 и усиливает функцию HAT JADE1 в живых клетках. TIP60 и JADE1 взаимно стабилизировали друг друга. Транскрипционная и HAT-активность JADE1 требует PHD2. Результаты свидетельствуют о роли JADE1 PHD2 в нацеливании на хроматин. [25] Кроме того, PHD2 JADE1 связывается с N-концевым хвостом гистона H3 в контексте хроматина независимо от статуса метилирования. [26]
Исследования, анализирующие нативные комплексы белков семейства пальцев PHD INhibitor of Growth (ING), показали, что белки ING4 и ING5 связаны с JADE1S и HAT HBO1, [27] в то время как ING3 связан с EPC1 (гомолог JADE1), TIP60 (гомолог HBO1) и несколькими другими партнерами. Оба комплекса также включали небольшой белок Eaf6. Биохимический анализ и анализ in silico комплексов, образованных HBO1 и TIP60, позволил предположить общую архитектуру и подтвердил роль JADE1 в массовом ацетилировании гистона H4. Характеристика функциональных взаимодействий JADE1 и HBO1 показывает структурное и функциональное сходство между комплексами (16, 19). Подобно TIP60, JADE1 и HBO1 взаимно стабилизируют друг друга. [28] JADE1 связывается с HBO1 и позволяет ему усиливать глобальное ацетилирование гистона H4, что требует интактного пальца PHD2. [28] Подобно HBO1, JADE1 отвечает за массовое ацетилирование гистона H4 в культивируемых клетках. H4K5, H4K12 и, скорее всего, H4K8 являются мишенями JADE1-зависимого ацетилирования в культивируемых клетках и in vivo. [14] [26] [29] В результате экспериментов с использованием подходов скрининга было предложено несколько потенциальных мишеней транскрипции JADE1. [26] [30] По данным скринингового геномного анализа, проведенного с помощью ChIP-чипа, комплекс JADE1L обнаруживается в основном вдоль кодирующих областей многих генов, а содержание JADE1L коррелирует в основном с H3K36me3 меткой гистона . Повышенная экспрессия JADE1L коррелирует с повышенным количеством H4acK8 в кодирующей области многих генов. [26] Два цинковых пальца PHD JADE1, по-видимому, связывают преимущественно неметилированный N-концевой пептид гистона. [26] [30] [31] Изоформы JADE1 образуют как минимум два разных комплекса: комплекс JADE1L-HBO1-ING4/5 и комплекс JADE1S-HBO1. [28] Из-за отсутствия С-концевого фрагмента JADE1S не способен связывать партнеров ING4/5. [28] Небольшой, менее охарактеризованный белок Eaf6 также является еще одним компонентом комплексов JADE1. [30]
Клеточный цикл
[ редактировать ]Известно, что ацетилирование N-концевых фрагментов объемного гистона H4 коррелирует с синтезом ДНК и делением клеток. [32] [33] [34] [35] [36] Несколько исследований подтверждают роль JADE1 в клеточном цикле, связанную с путем HBO1. [25] [29] И JADE1, и HBO1 независимо необходимы для ацетилирования гистона H4 в культивируемых клетках. [25] [27] [28] [29] Истощение белков JADE1 миРНК приводит к 1) снижению уровня массового ацетилирования гистона H4; 2) более медленные темпы синтеза ДНК в культивируемых клетках; [29] 3) снижение уровня общего и связанного с хроматином HBO1; [28] [29] 4) прекращение рекрутирования хроматина MCM7. [29] В соответствии с этими результатами, сверхэкспрессия JADE1L увеличивает связанный с хроматином белок MCM3. [37] Эффекты истощения JADE1 на события репликации ДНК аналогичны эффектам, описанным первоначально для HBO1. [38] и предполагает роль адаптера JADE1 в регуляции клеточного цикла, опосредованной HBO1.
Была высказана гипотеза о роли JADE1 в повреждении ДНК. Недавно обнаруженная некодирующая РНК lncRNA-JADE регулирует экспрессию JADE1 и обеспечивает функциональную связь между реакцией на повреждение ДНК (DDR) и объемным ацетилированием гистона H4. [39] Результаты подтверждают роль в синтезе ДНК, связанную с ацетилированием гистона H4. [39] В культивируемых клетках нокдаун lncRNA-JADE повышал чувствительность клеток к препаратам, повреждающим ДНК. На модели ксенотрансплантата опухоли у мышей нокдаун lncRNA-JADE ингибировал рост опухоли ксенотрансплантата молочной железы. В пилотном исследовании на людях более высокие уровни lncRNA-JADE, а также белка JADE1 были обнаружены в тканях рака молочной железы по сравнению с нормальными тканями. Наконец, более высокие уровни белка JADE1 обратно коррелировали с выживаемостью пациентов с раком молочной железы. Исследование предполагает, что днРНК-JADE может способствовать образованию опухолей молочной железы и что белок JADE1 опосредует, по крайней мере, часть этого эффекта. [39] JADE1 и цитокинез. JADE1S отрицательно регулирует цитокинез цикла эпителиальных клеток - функцию, специфичную для малой изоформы. [14] [15] Первый отчет, в котором предполагалась функция JADE1 при переходе G2/M/G1, показал, что во время поздней фазы G2 JADE1S подвергается фосфорилированию, связанному с его диссоциацией из хроматина в цитоплазму. Масс-спектральный анализ показал, что всего шесть отдельных аминокислотных остатков фосфорилируются митотической киназой. [14] Согласно фармакологическому анализу, фосфорилирование и компартментализация JADE1 регулируются путями Aurora A и Aurora B. [14] [15] Сообщалось о других киназах, которые могут играть определенную роль. [16] [40] По завершении митоза около телофазы основной пул белка JADE1S подвергается дефосфорилированию и повторно связывается с очевидно конденсирующимся хроматином внутри реформированных ядер. [14] Дискретный пул JADE1S связывается с бороздой расщепления и впоследствии появляется в средней части цитокинетического моста. [15] Только JADE1S, но не JADE1L или HBO1, был обнаружен в средней части тела клеток, подвергающихся цитокинезу. Пространственная регуляция JADE1S во время клеточного деления предполагает его роль в переходе от G2/M к G1, который включает цитокинез и окончательное отторжение. [15] [41] Цитокинез — заключительный этап клеточного цикла, который контролирует правильность деления клеточного содержимого, включая цитоплазму, мембрану и хроматин. Цитокинетический мост разрывается во время окончательного отторжения, которое происходит вблизи средней части тела и может занять до 2 часов. Цитокинез и окончательное отторжение жестко контролируются регуляторными белковыми комплексами и белками контрольных точек. За последнее десятилетие число сообщений о контроле цитокинеза росло. [42] [43] [44] [45] [46]
Роль JADE1 в цитокинезе была продемонстрирована с помощью нескольких функциональных анализов и моделей клеточных культур. [15] Профилирование ДНК с помощью FACS показало, что истощение JADE1S способствует скорости накопления G1-клеток в синхронно делящихся клетках HeLa. Истощение белка JADE1S в асинхронно делящихся клетках уменьшало долю цитокинетических клеток и увеличивало долю многоядерных клеток. Данные продемонстрировали, что JADE1 отрицательно контролирует цитокинез, предположительно способствуя задержке цитокинеза. Снижение регуляции JADE1 увеличивало количество многоядерных клеток, что свидетельствует о нарушении цитокинеза, тогда как умеренная сверхэкспрессия JADE1S увеличивала количество цитокинетических клеток, что свидетельствовало о цитокинетической задержке. Ингибирование киназы Aurora B специфическими низкомолекулярными препаратами приводило к высвобождению JADE1S-опосредованной цитокинетической задержки и способствовало прогрессированию отторжения. Поскольку Aurora B является ключевым регулятором NoCut, JADE1S, вероятно, регулирует цитокинез в контрольной точке отрыва. [15] [41] JADE1S, но не JADE1L или HBO1, был обнаружен в центросомах делящихся клеток на протяжении всего клеточного цикла, и ни один из этих белков не был обнаружен в ресничках. Напротив, в другом исследовании сообщалось о локализации JADE1 в ресничках и центросомах. [40] В исследовании не сообщалось о специфичности изоформы JADE1. [40] Центросомы являются центрами нуклеации цитоскелета. Передача сигналов центросом способствует определению формы клеток, подвижности, ориентации, полярности, плоскости деления и точности разделения сестринских хромосом во время митоза и цитокинеза. [47] [48]
пВХЛ
[ редактировать ]Первый белок-партнер JADE1S был идентифицирован в 2002 году в исследовании по поиску новых партнеров pVHL, который является супрессором опухоли. [11] Несколько последующих исследований охарактеризовали связывание и предоставили некоторую информацию о функциональных взаимодействиях JADE1-pVHL. [49] [50] [51] pVHL человека мутирует при наследственной болезни фон Хиппеля-Линдау и в большинстве спорадических светлоклеточных карцином почки. [52] [53] [54] [55] [56] Свойства и функции pVHL исследуются на протяжении многих десятилетий, и доступна обширная литература. Одной из наиболее известных функций pVHL является обеспечение убиквитинирования и протеосомной деградации белков. В качестве компонента комплекса убиквитинлигазы E3 pVHL связывается и нацеливается на несколько известных факторов, включая HIF1a и HIF2a для убиквитинирования. [55] О механизме активации HIF1a при гипоксии и роли pVHL в этом пути сообщалось более десяти лет назад. [57] Белок VHL был тщательно изучен и установлена связь возникающих в природе мутаций с раком. Были рассмотрены другие причинные HIF-1a-независимые пути pVHL. [58] Физическое взаимодействие pVHL-JADE1S было идентифицировано с помощью скринингового анализа дрожжевых гибридов и дополнительно подтверждено биохимически. Котрансфекция pVHL увеличила период полувыведения и количество белка JADE1S, что указывает на потенциальную положительную взаимосвязь. [11] Определенные усечения pVHL, вызванные раком, но не точечные мутации, снижают функцию стабилизации pVHL-JADE1, что указывает на связь с раком, связанным с pVHL. [51] Молекулярные пути и клеточное значение взаимодействий JADE1-pVHL недостаточно изучены. В 2008 году было опубликовано единственное исследование, описывающее внутреннюю убиквитин-лигазную активность JADE1S и убиквитинирование бета-катенина. [49] На основании этого исследования была предложена модель, согласно которой pVHL регулирует бета-катенин через JADE1, и для этой активности необходимы цинковые пальцы PHD.
Апоптоз
[ редактировать ]Была предложена функция JADE1S при апоптозе, но механизмы остаются неясными, а результаты трудно согласовать. [11] [30] [49] [50] [51] Согласно исследованиям, сверхэкспрессия JADE1 замедляет скорость клеточного роста и индуцирует белок остановки клеточного цикла p21. Несколько попыток создать надежные клеточные линии, стабильно экспрессирующие белок JADE1S, не увенчались успехом, предположительно из-за самоотбора негативных клеток. Напротив, другое исследование показывает, что подавление JADE1 снижает скорость синтеза ДНК в синхронно делящихся клетках. [28] [39] По данным непрямой иммунофлуоресценции и микроскопического анализа культивируемых клеток, перегрузка культивируемых клеток белком JADE1 вызывает клеточную токсичность и побочные эффекты. [15] Клетки претерпевают морфологические изменения, которые не напоминают апоптоз, но предполагают серьезное нарушение клеточного цикла, включая окрашивание клеток аномальной формы и крупные многодольковые ядра. [15] На основании JADE1S-опосредованной регуляции клеточного цикла рассматриваются и другие интерпретации: перегрузка JADE1 может вызывать длительное NoCut и остановку цитокинеза или серьезный дисбаланс клеточного цикла, а не прямую активацию транскрипции апоптоза. [15]
Биологическая роль
[ редактировать ]Биологическая роль JADE1 не выяснена. Ограниченное количество публикаций посвящено этому вопросу с использованием моделей на мышах. Наиболее полное исследование, опубликованное в 2003 году, выявило у мышей ортолог человеческого JADE1, Jade1, и исследовало экспрессию Jade1 во время эмбриогенеза мышей. [6] В поисках генов, регулируемых развитием, авторы использовали скрининговый анализ генных ловушек и идентифицировали мышиный Jade1 как ген, строго регулируемый во время эмбриогенеза. Вставка вектора в третий интрон гена Jade1 приводит к образованию укороченного белка, состоящего из 47 аминокислот. Инсерционная мутация генной ловушки привела к образованию продукта слияния репортера Jade1-бета-галактозидазы и нулевого аллеля Jade1. Хотя гомозиготы по интеграции генной ловушки не давали сильного фенотипа развития, продукт слияния выявил пространственно-временную экспрессию гена Jade1 в эмбриональных клетках мыши и тканях развивающегося эмбриона до 15,5 дпк. Кроме того, в исследовании сообщается об экспериментальном и сравнительном анализе in silico. транскриптов мРНК Jade1, структуры гена Jade1 и анализа ортологов белка Jade1 от мыши, человека и рыбки данио. [6] Экспрессия Jade1 была обнаружена во внеэмбриональной эктодерме и трофобласте, которые являются плацентарными компонентами, важными для васкулогенеза, а также в участках, обогащенных мультипотентными или тканеспецифичными предшественниками, включая нейральные предшественники (2). Динамика экспрессии репортера Jade1 в этих областях указывает на участие в детерминации и удлинении передне-задней оси, что является важным моментом исследования). [6] Потенциальная роль человеческого JADE1 в обновлении культур эмбриональных стволовых клеток и клеток эмбриональной карциномы была предложена в другом скрининговом исследовании, которое показало, что в культивируемых стволовых клетках активация пути транскрипционного фактора стволовых клеток OCT4 повышает экспрессию гена JADE1 наряду с факторами стволовых клеток NANOG. , PHC1, USP44 и SOX2. [59] Роль JADE1 в пролиферации эпителиальных клеток рассматривалась на мышиной модели острого повреждения и регенерации почек. [14] [29] Характер экспрессии и динамику HBO1-JADE1S/L исследовали в регенерирующих эпителиальных клетках канальцев. [29] Ишемия и реперфузионное повреждение приводили к первоначальному снижению уровней белков JADE1S, JADE1L и HBO1, которые возвращались к исходному уровню во время восстановления функции почек. Уровни экспрессии HBO1 и JADE1S восстанавливались, когда скорость пролиферации клеток достигала максимума, тогда как JADE1L восстанавливался после уменьшения объемной пролиферации. Временная экспрессия JADE1 коррелирует с ацетилированием гистона H4 (H4K5 и H4K12), но не с ацетилированием гистона H3 (H4K14), что позволяет предположить, что комплекс JADE1-HBO1 специфически маркирует H4 во время пролиферации эпителиальных клеток. Результаты исследования указывают на участие комплекса JADE1-HBO1 в остром повреждении почек и предполагают различную роль изоформ JADE1 во время восстановления эпителиальных клеток. [29]
Ассоциации заболеваний
[ редактировать ]Роль JADE1 в заболеваниях человека не выяснена. Недавнее исследование выявило новые субмикроскопические генетические изменения при миелофиброзе, раке костного мозга. [60] Исследование выявило семь новых делеций и транслокаций у небольшой группы пациентов с первичным миелофиброзом. JADE1 и соседний ген, называемый натриевым каналом и клатриновым линкером 1 (SCLT1), были значительно модифицированы. В результате мутации ген JADE1 имеет делеции интрона 5–6 и экзонов 6–11, в результате чего в JADE1 отсутствует большой фрагмент белка, начиная с цинкового пальца PHD. Связь с патогенезом находится в стадии изучения.В нескольких пилотных исследованиях экспрессию JADE1 изучали при раке толстой кишки и карциноме почек. Результаты этих исследований не всегда согласуются. Результаты некоторых исследований получены в основном на основе гистохимического анализа образцов опухолей с использованием антитела JADE1 с неустановленной специфичностью к JADE1 в целом и к JADE1S или JADE1L в частности. [61] [62] Результаты исследования с использованием алгоритма анализа микрочипов in silico показывают, что мРНК PHF17 может играть роль в развитии рака поджелудочной железы. [63] Эти многообещающие направления расследований требуют дальнейшего контроля и дополнительных оценок.
Недавние данные свидетельствуют о том, что JADE1 может играть роль в нейродегенеративных таупатиях . [64] . В частности, локус JADE1 был идентифицирован в небольшом полногеномном исследовании ассоциаций на основе аутопсии у субъектов с первичной возрастной таупатией (PART). [65] Дальнейшие гистологические и биохимические исследования показали специфическое взаимодействие между JADE1 и изоформами ассоциированного с микротрубочками белка тау с четырьмя доменами связывания микротрубочек, но не с тремя. Модели дрозофилы in vivo показали, что нокдаун ортолога-носорога усугубляет фенотипы, связанные с тау-токсичностью, что предполагает защитную роль. Гистологические исследования показали, что JADE1 накапливается при большинстве таупатий, за исключением болезни Пика , которая примечательна тем, что отличается избирательным накоплением изоформ тау с тремя повторами домена, связывающего микротрубочки, к которым JADE1 имеет низкое сродство. Необходимы дальнейшие исследования, чтобы понять роль JADE1 в нейродегенерации.
Взаимодействия
[ редактировать ]С JADE1 взаимодействуют несколько белков, в том числе: MAPT, [66] пВХЛ, [11] СОВЕТ60, [25] ГБО1, ИНГ4, ИНГ5, [28] β-катенин, [49] НПХП4. [40]
Примечания
[ редактировать ]Версия этой статьи 2016 года была обновлена внешним экспертом в рамках модели двойной публикации. Соответствующая академическая рецензируемая статья была опубликована в журнале Gene и может цитироваться как: Мария В. Панченко (4 мая 2016 г.). «Структура, функции и регуляция пальца PHD 1 семейства нефритовых (JADE1)» . Джин . Серия обзоров Gene Wiki. 589 (1): 1–11. дои : 10.1016/J.GENE.2016.05.002 . ISSN 0378-1119 . ПМЦ 4903948 . ПМИД 27155521 . Викиданные Q38828242 . |
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000077684 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: Ensembl, выпуск 89: ENSMUSG00000025764 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Нагасе Т., Накаяма М., Накадзима Д., Кикуно Р., Охара О. (апрель 2001 г.). «Прогнозирование кодирующих последовательностей неопознанных генов человека. XX. Полные последовательности 100 новых клонов кДНК головного мозга, которые кодируют крупные белки in vitro» . Исследование ДНК . 8 (2): 85–95. дои : 10.1093/dnares/8.2.85 . ПМИД 11347906 .
- ^ Jump up to: а б с д и ж г час Цуанаку Э., Твиди С., Уилсон В. (декабрь 2003 г.). «Идентификация Jade1, гена, кодирующего белок цинковых пальцев PHD, в скрининге мутагенеза генной ловушки для генов, участвующих в развитии переднезадней оси» . Молекулярная и клеточная биология . 23 (23): 8553–2. дои : 10.1128/mcb.23.23.8553-8562.2003 . ПМК 262661 . ПМИД 14612400 .
- ^ Чжоу М.И., Ван Х., Росс Дж.Дж., Кузьмин И., Сюй С., Коэн Х.Т. (октябрь 2002 г.). «Супрессор опухоли фон Хиппель-Линдау стабилизирует новый растительный гомеодоменовый белок Jade-1» . Журнал биологической химии . 277 (42): 39887–98. дои : 10.1074/jbc.M205040200 . ПМИД 12169691 .
- ^ «Ген Энтрез: белок 17 пальца PHF17 PHD» .
- ^ Jump up to: а б Нагасе Т., Накаяма М., Накадзима Д., Кикуно Р., Охара О. (апрель 2001 г.). «Прогнозирование кодирующих последовательностей неопознанных генов человека. XX. Полные последовательности 100 новых клонов кДНК головного мозга, которые кодируют крупные белки in vitro» . Исследование ДНК . 8 (2): 85–95. дои : 10.1093/dnares/8.2.85 . ПМИД 11347906 .
- ^ Томпсон К.А., Ван Б., Аргрейвс В.С., Джанкотти Ф.Г., Шранк Д.П., Руослахти Е (февраль 1994 г.). «BR140, новый белок цинковых пальцев, гомологичный субъединице TAF250 TFIID» . Связь с биохимическими и биофизическими исследованиями . 198 (3): 1143–52. дои : 10.1006/bbrc.1994.1162 . ПМИД 7906940 .
- ^ Jump up to: а б с д и ж г час Чжоу М.И., Ван Х., Росс Дж.Дж., Кузьмин И., Сюй С., Коэн Х.Т. (октябрь 2002 г.). «Супрессор опухоли фон Хиппель-Линдау стабилизирует новый растительный гомеодоменовый белок Jade-1» . Журнал биологической химии . 277 (42): 39887–98. дои : 10.1074/jbc.M205040200 . ПМИД 12169691 .
- ^ Селей Дж., Сото А.М., Гек П., Десронвил М., Прехтль Н.В., Вейл Б.С., Зонненшайн С. (март 2000 г.). «Идентификация человеческих эстроген-индуцируемых транскриптов, которые потенциально опосредуют апоптотический ответ при раке молочной железы». Журнал биохимии стероидов и молекулярной биологии . 72 (3–4): 89–102. дои : 10.1016/s0960-0760(00)00025-x . ПМИД 10775800 . S2CID 25912630 .
- ^ «Белок Джейд-1 (Q6IE81)» . ИнтерПро .
- ^ Jump up to: а б с д и ж г Сиривардана Н.С., Мейер Р., Хаваси А., Домингес И., Панченко М.В. (2014). «Зависимое от клеточного цикла перемещение хроматина комплекса гистонацетилтрансферазы (HAT) HBO1-JADE1» . Клеточный цикл . 13 (12): 1885–901. дои : 10.4161/cc.28759 . ПМЦ 4111752 . ПМИД 24739512 .
- ^ Jump up to: а б с д и ж г час я дж к Сиривардана Н.С., Мейер Р.Д., Панченко М.В. (2015). «Новая функция JADE1S в цитокинезе эпителиальных клеток» . Клеточный цикл . 14 (17): 2821–34. дои : 10.1080/15384101.2015.1068476 . ПМЦ 4612376 . ПМИД 26151225 .
- ^ Jump up to: а б Боргал Л., Риншен М.М., Дафингер С., Хофф С., Райнерт М.Дж., Ламкемейер Т., Лиенкамп С.С., Бенцинг Т., Шермер Б. (сентябрь 2014 г.). «Казеинкиназа 1 α фосфорилирует регулятор Wnt Jade-1 и модулирует его активность» . Журнал биологической химии . 289 (38): 26344–56. дои : 10.1074/jbc.M114.562165 . ПМК 4176241 . ПМИД 25100726 .
- ^ «JADE1 – Поиск – Homo sapiens – Браузер генома Ensembl 84» .
- ^ Высоцка Дж., Свигут Т., Сяо Х., Милн Т.А., Квон С.Ю., Лэндри Дж., Кауэр М., Тэкетт А.Дж., Чайт Б.Т., Баденхорст П., Ву С., Эллис К.Д. (июль 2006 г.). «Палец PHD NURF сочетает триметилирование лизина 4 гистона H3 с ремоделированием хроматина». Природа . 442 (7098): 86–90. Бибкод : 2006Natur.442...86W . дои : 10.1038/nature04815 . ПМИД 16728976 . S2CID 4389087 .
- ^ Ши X, Хонг Т, Уолтер К.Л., Эвальт М., Мичишита Э., Хунг Т., Карни Д., Пенья П., Лан Ф., Каадиге М.Р., Лакост Н., Кайру С., Давразу Ф., Саха А., Кэрнс Б.Р., Айер Д.Е., Кутателадзе Т.Г. , Ши Ю, Коте Дж, Чуа К.Ф., Гозани О (июль 2006 г.). «Домен ING2 PHD связывает метилирование лизина 4 гистона H3 с активной репрессией генов» . Природа . 442 (7098): 96–9. Бибкод : 2006Natur.442...96S . дои : 10.1038/nature04835 . ПМК 3089773 . ПМИД 16728974 .
- ^ Таверна С.Д., Илин С., Роджерс Р.С., Тэнни Дж.С., Лаванда Х., Ли Х., Бейкер Л., Бойл Дж., Блэр Л.П., Чейт Б.Т., Патель DJ, Эйчисон Дж.Д., Тэкетт А.Дж., Эллис CD (декабрь 2006 г.). «Связывание пальца Yng1 PHD с триметилированным по K4 H3 способствует активности NuA3 HAT по K14 H3 и транскрипции в подмножестве целевых ORF» . Молекулярная клетка . 24 (5): 785–96. doi : 10.1016/j.molcel.2006.10.026 . ПМК 4690528 . ПМИД 17157260 .
- ^ Санчес Р., Чжоу М.М. (июль 2011 г.). «Палец PHD: универсальный читатель эпигенома» . Тенденции биохимических наук . 36 (7): 364–72. дои : 10.1016/j.tibs.2011.03.005 . ПМК 3130114 . ПМИД 21514168 .
- ^ Кван А.Х., Гелл Д.А., Вергер А., Кроссли М., Мэтьюз Дж.М., Маккей Дж.П. (июль 2003 г.). «Создание белкового каркаса из пальца доктора философии» . Структура . 11 (7): 803–13. дои : 10.1016/s0969-2126(03)00122-9 . ПМИД 12842043 .
- ^ Мэнсфилд Р.Э., Массельман К.А., Кван А.Х., Оливер С.С., Гарске А.Л., Давразу Ф., Дену Дж.М., Кутателадзе Т.Г., Маккей Дж.П. (апрель 2011 г.). «Растительные гомеодомены (PHD) пальцы CHD4 представляют собой модули, связывающие гистон H3, с предпочтением немодифицированного H3K4 и метилированного H3K9» . Журнал биологической химии . 286 (13): 11779–91. дои : 10.1074/jbc.M110.208207 . ПМК 3064229 . ПМИД 21278251 .
- ^ Мэтьюз Дж.М., Бхати М., Лехтомаки Э., Мэнсфилд Р.Э., Кубедду Л., Маккей Дж.П. (2009). «Для танго нужны двое: структура и функции доменов LIM, RING, PHD и MYND». Текущий фармацевтический дизайн . 15 (31): 3681–96. дои : 10.2174/138161209789271861 . ПМИД 19925420 .
- ^ Jump up to: а б с д Панченко М.В., Чжоу М.И., Коэн Х.Т. (декабрь 2004 г.). «Партнер фон Хиппель-Линдау Jade-1 представляет собой коактиватор транскрипции, связанный с активностью гистон-ацетилтрансферазы» . Журнал биологической химии . 279 (53): 56032–41. дои : 10.1074/jbc.M410487200 . ПМИД 15502158 .
- ^ Jump up to: а б с д и Саксук Н., Аввакумов Н., Шампань К.С., Хунг Т., Дойон Ю., Кайру С., Пакет Э., Улла М., Ландри А.Дж., Коте В., Ян XJ, Гозани О., Кутателадзе Т.Г., Коте Дж. (январь 2009 г.). «Комплексы HBO1 HAT нацелены на хроматин во всех областях кодирования гена посредством множественных взаимодействий пальца PHD с хвостом гистона H3» . Молекулярная клетка . 33 (2): 257–65. doi : 10.1016/j.molcel.2009.01.007 . ПМЦ 2677731 . ПМИД 19187766 .
- ^ Jump up to: а б Дойон Ю, Кайру С, Улла М, Лэндри А.Дж., Коте В., Селлек В., Лейн В.С., Тан С., Ян XJ, Коте Дж. (январь 2006 г.). «Белки-супрессоры опухолей ING являются важными регуляторами ацетилирования хроматина, необходимыми для экспрессии и сохранения генома» . Молекулярная клетка . 21 (1): 51–64. doi : 10.1016/j.molcel.2005.12.007 . ПМИД 16387653 .
- ^ Jump up to: а б с д и ж г час Фой Р.Л., Сонг И.Ю., Читалия В.К., Коэн Х.Т., Саксук Н., Кайру С., Вазири С., Коте Ж., Панченко М.В. (октябрь 2008 г.). «Роль Jade-1 в комплексе гистонацетилтрансферазы (HAT) HBO1» . Журнал биологической химии . 283 (43): 28817–26. дои : 10.1074/jbc.M801407200 . ПМК 2570895 . ПМИД 18684714 .
- ^ Jump up to: а б с д и ж г час я Хаваси А., Хегеле Дж.А., Галл Дж.М., Блэкмон С., Ичимура Т., Бонеджио Р.Г., Панченко М.В. (январь 2013 г.). «Гистонацетилтрансфераза (HAT) HBO1 и JADE1 в регенерации эпителиальных клеток» . Американский журнал патологии . 182 (1): 152–62. дои : 10.1016/j.ajpath.2012.09.017 . ПМЦ 3532714 . ПМИД 23159946 .
- ^ Jump up to: а б с д Аввакумов Н., Лалонд М.Э., Саксук Н., Пакет Э., Гласс К.С., Ландри А.Дж., Дойон Ю., Кайру С., Робитайл Г.А., Ришар Д.Е., Ян XJ, Кутателадзе Т.Г., Коте Дж. (февраль 2012 г.). «Консервативные молекулярные взаимодействия внутри комплексов ацетилтрансферазы HBO1 регулируют пролиферацию клеток» . Молекулярная и клеточная биология . 32 (3): 689–703. дои : 10.1128/MCB.06455-11 . ПМК 3266594 . ПМИД 22144582 .
- ^ Лалонд М.Э., Аввакумов Н., Гласс К.С., Жонкас Ф.Х., Саксук Н., Холлидей М., Пакет Э, Ян К., Тонг К., Кляйн Б.Дж., Тан С., Ян XJ, Кутателадзе Т.Г., Коте Дж. (сентябрь 2013 г.). «Обмен ассоциированными факторами приводит к изменению специфичности хвоста гистонов ацетилтрансферазы HBO1» . Гены и развитие . 27 (18): 2009–24. дои : 10.1101/gad.223396.113 . ПМЦ 3792477 . ПМИД 24065767 .
- ^ Меги ПК, Морган Б.А., Смит М.М. (июль 1995 г.). «Гистон H4 и поддержание целостности генома» . Гены и развитие . 9 (14): 1716–27. дои : 10.1101/gad.14.9.1716 . ПМИД 7622036 .
- ^ Маки Н., Цонис П.А., Агата К. (16 сентября 2010 г.). «Изменения глобальных модификаций гистонов во время дедифференцировки при регенерации хрусталика тритона» . Молекулярное видение . 16 : 1893–7. ПМЦ 2956703 . ПМИД 21031136 .
- ^ Ясенчакова З., Мейстер А., Уолтер Дж., Тернер Б.М., Шуберт И. (ноябрь 2000 г.). «Ацетилирование гистонов H4 эухроматина и гетерохроматина зависит от клеточного цикла и коррелирует с репликацией, а не с транскрипцией» . Растительная клетка . 12 (11): 2087–100. дои : 10.1105/tpc.12.11.2087 . ПМК 150160 . ПМИД 11090211 .
- ^ Кларк А.С., Лоуэлл Дж.Э., Джейкобсон С.Дж., Пиллус Л. (апрель 1999 г.). «Esa1p — это незаменимая гистон-ацетилтрансфераза, необходимая для развития клеточного цикла» . Молекулярная и клеточная биология . 19 (4): 2515–26. дои : 10.1128/mcb.19.4.2515 . ПМК 84044 . ПМИД 10082517 .
- ^ Чой Дж.С., Тобе Б.Т., Ха Дж.Х., Крон С.Дж. (ноябрь 2001 г.). «Yng2p-зависимая активность ацетилирования гистона H4 NuA4 необходима для митотической и мейотической прогрессии» . Журнал биологической химии . 276 (47): 43653–62. дои : 10.1074/jbc.M102531200 . ПМИД 11544250 .
- ^ Миотто Б., Струл К. (январь 2010 г.). «Активность ацетилазы гистонов HBO1 необходима для лицензирования репликации ДНК и ингибируется геминином» . Молекулярная клетка . 37 (1): 57–66. doi : 10.1016/j.molcel.2009.12.012 . ПМЦ 2818871 . ПМИД 20129055 .
- ^ Иидзука М., Мацуи Т., Такисава Х., Смит М.М. (февраль 2006 г.). «Регулирование лицензирования репликации ацетилтрансферазой Hbo1» . Молекулярная и клеточная биология . 26 (3): 1098–108. дои : 10.1128/MCB.26.3.1098-1108.2006 . ПМК 1347032 . ПМИД 16428461 .
- ^ Jump up to: а б с д Ван Г, Ху Х, Лю Ю, Хань С., Суд А.К., Калин Г.А., Чжан Х, Лу Х (октябрь 2013 г.). «Новая некодирующая РНК lncRNA-JADE связывает передачу сигналов о повреждении ДНК с ацетилированием гистона H4» . Журнал ЭМБО . 32 (21): 2833–47. дои : 10.1038/emboj.2013.221 . ПМЦ 3817469 . ПМИД 24097061 .
- ^ Jump up to: а б с д Боргал Л., Хаббиг С., Хацольд Дж., Либау М.К., Дафингер С., Сакареа И., Хаммершмидт М., Бенцинг Т., Шермер Б. (июль 2012 г.). «Цилиарный белок нефроцистин-4 перемещает канонический регулятор Wnt Jade-1 в ядро, чтобы отрицательно регулировать передачу сигналов β-катенина» . Журнал биологической химии . 287 (30): 25370–80. дои : 10.1074/jbc.M112.385658 . ПМК 3408186 . ПМИД 22654112 .
- ^ Jump up to: а б Прекерис Р. (2015). «Cut или NoCut: роль JADE1S в регулировании контрольной точки отрыва» . Клеточный цикл . 14 (20): 3219. дои : 10.1080/15384101.2015.1089074 . ПМЦ 4825624 . ПМИД 26327571 .
- ^ Агромайор М., Мартин-Серрано Дж. (сентябрь 2013 г.). «Знать, когда действовать: механизмы, контролирующие цитокинетическое отторжение». Тенденции в клеточной биологии . 23 (9): 433–41. дои : 10.1016/j.tcb.2013.04.006 . ПМИД 23706391 .
- ^ Элия Н., Суграт Р., Сперлин Т.А., Херли Дж.Х., Липпинкотт-Шварц Дж. (март 2011 г.). «Динамика эндосомального сортировочного комплекса, необходимого для транспортного механизма (ESCRT) во время цитокинеза, и его роль в отторжении» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (12): 4846–51. Бибкод : 2011PNAS..108.4846E . дои : 10.1073/pnas.1102714108 . ПМК 3064317 . ПМИД 21383202 .
- ^ Фаббро М., Чжоу Б.Б., Такахаши М., Сарцевич Б., Лал П., Грэм М.Э., Габриэлли Б.Г., Робинсон П.Дж., Нигг Э.А., Оно Ю., Ханна К.К. (октябрь 2005 г.). «Cdk1/Erk2- и Plk1-зависимое фосфорилирование центросомного белка Cep55 необходимо для его рекрутирования в среднее тело и цитокинеза» . Развивающая клетка . 9 (4): 477–88. дои : 10.1016/j.devcel.2005.09.003 . ПМИД 16198290 .
- ^ Грин Р.А., Палух Э., Огема К. (2012). «Цитокинез в животных клетках». Ежегодный обзор клеточной биологии и биологии развития . 28 : 29–58. doi : 10.1146/annurev-cellbio-101011-155718 . ПМИД 22804577 .
- ^ Ху С.К., Кофлин М., Митчисон Т.Дж. (март 2012 г.). «Сборка среднего тела и ее регуляция во время цитокинеза» . Молекулярная биология клетки . 23 (6): 1024–34. doi : 10.1091/mbc.E11-08-0721 . ПМК 3302730 . ПМИД 22278743 .
- ^ Нигг Э.А., Стернс Т. (октябрь 2011 г.). «Центросомный цикл: биогенез центриолей, дупликация и присущие асимметрии» . Природная клеточная биология . 13 (10): 1154–60. дои : 10.1038/ncb2345 . ПМЦ 3947860 . ПМИД 21968988 .
- ^ Слудер Г., Ходжаков А. (декабрь 2010 г.). «Дупликация центриолей: аналоговый контроль в эпоху цифровых технологий» . Международная клеточная биология . 34 (12): 1239–45. дои : 10.1042/CBI20100612 . ПМК 3051170 . ПМИД 21067522 .
- ^ Jump up to: а б с д Читалия В.К., Фой Р.Л., Бахшмид М.М., Цзэн Л., Панченко М.В., Чжоу М.И., Бхарти А., Селдин Д.С., Лекер Ш.Х., Домингес И., Коэн Х.Т. (октябрь 2008 г.). «Jade-1 ингибирует передачу сигналов Wnt путем убиквитилирования бета-катенина и опосредует ингибирование пути Wnt с помощью pVHL» . Природная клеточная биология . 10 (10): 1208–16. дои : 10.1038/ncb1781 . ПМЦ 2830866 . ПМИД 18806787 .
- ^ Jump up to: а б Чжоу М.И., Фой Р.Л., Читалия В.К., Чжао Дж., Панченко М.В., Ван Х., Коэн Х.Т. (август 2005 г.). «Джад-1, кандидат в супрессоры опухолей почек, способствующий апоптозу» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (31): 11035–40. Бибкод : 2005PNAS..10211035Z . дои : 10.1073/pnas.0500757102 . ПМК 1182408 . ПМИД 16046545 .
- ^ Jump up to: а б с Чжоу М.И., Ван Х., Фой Р.Л., Росс Дж.Дж., Коэн Х.Т. (февраль 2004 г.). «Стабилизация белка Jade-1 опухолевым супрессором фон Хиппеля-Линдау (VHL) происходит посредством гомеодоменов растений и зависит от мутации VHL» . Исследования рака . 64 (4): 1278–86. дои : 10.1158/0008-5472.can-03-0884 . ПМИД 14973063 .
- ^ Кросси П.А., Ричардс Ф.М., Фостер К., Грин Дж.С., Проуз А., Латиф Ф., Лерман М.И., Збар Б., Аффара Н.А., Фергюсон-Смит М.А. (август 1994 г.). «Идентификация внутригенных мутаций в гене-супрессоре опухоли при болезни фон Хиппель-Линдау и корреляция с фенотипом заболевания». Молекулярная генетика человека . 3 (8): 1303–8. дои : 10.1093/hmg/3.8.1303 . ПМИД 7987306 .
- ^ Фостер К., Проуз А., ван ден Берг А., Флеминг С., Халсбек М.М., Кросси П.А., Ричардс Ф.М., Кэрнс П., Аффара Н.А., Фергюсон-Смит М.А. (декабрь 1994 г.). «Соматические мутации гена-супрессора опухоли при болезни фон Гиппеля-Линдау при несемейном светлоклеточном раке почки». Молекулярная генетика человека . 3 (12): 2169–73. дои : 10.1093/hmg/3.12.2169 . ПМИД 7881415 .
- ^ Дуан Д.Р., Хамфри Дж.С., Чен Д.И., Венг Ю., Сукегава Дж., Ли С., Гнарра Дж.Р., Линехан В.М., Клаузнер Р.Д. (июль 1995 г.). «Характеристика продукта гена-супрессора опухоли VHL: локализация, образование комплекса и влияние естественных инактивирующих мутаций» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (14): 6459–63. Бибкод : 1995PNAS...92.6459D . дои : 10.1073/pnas.92.14.6459 . ПМК 41537 . ПМИД 7604013 .
- ^ Jump up to: а б Максвелл П.Х., Визенер М.С., Чанг Г.В., Клиффорд СК, Во Э.К., Кокман М.Е., Вайкофф СС, Пью К.В., Махер Э.Р., Рэтклифф П.Дж. (май 1999 г.). «Белок-супрессор опухоли VHL нацелен на факторы, индуцируемые гипоксией, для кислородзависимого протеолиза». Природа . 399 (6733): 271–5. Бибкод : 1999Natur.399..271M . дои : 10.1038/20459 . ПМИД 10353251 . S2CID 4427694 .
- ^ Латиф Ф., Тори К., Гнарра Дж., Яо М., Дух Ф.М., Оркатт М.Л., Стэкхаус Т., Кузьмин И., Моди В., Гейл Л. (май 1993 г.). «Идентификация гена-супрессора опухоли при болезни фон Хиппель-Линдау». Наука . 260 (5112): 1317–20. Бибкод : 1993Sci...260.1317L . дои : 10.1126/science.8493574 . ПМИД 8493574 .
- ^ Яаккола П., Моул Д.Р., Тиан Ю.М., Уилсон М.И., Гилберт Дж., Гаскелл С.Дж., фон Кригсхайм А., Хебестрайт Х.Ф., Мукерджи М., Шофилд К.Дж., Максвелл П.Х., Пью К.В., Рэтклифф П.Дж. (апрель 2001 г.). «Нацеливание HIF-альфа на комплекс убиквитилирования фон Хиппель-Линдау посредством O2-регулируемого пролилгидроксилирования» . Наука . 292 (5516): 468–72. Бибкод : 2001Sci...292..468J . дои : 10.1126/science.1059796 . ПМИД 11292861 . S2CID 20914281 .
- ^ Госсэйдж Л., Эйзен Т., Махер Э.Р. (январь 2015 г.). «VHL, история гена-супрессора опухолей». Обзоры природы. Рак . 15 (1): 55–64. дои : 10.1038/nrc3844 . ПМИД 25533676 . S2CID 19312746 .
- ^ Юнг М., Петерсон Х., Чавес Л., Калем П., Лерах Х., Вило Дж., Аджайе Дж. (21 мая 2010 г.). «Подход к интеграции данных для картирования регуляторных сетей гена OCT4, действующих в эмбриональных стволовых клетках и клетках эмбриональной карциномы» . ПЛОС ОДИН . 5 (5): е10709. Бибкод : 2010PLoSO...510709J . дои : 10.1371/journal.pone.0010709 . ПМЦ 2873957 . ПМИД 20505756 .
- ^ Лашо Т., Джонсон С.Х., Смит Д.И., Криспино Дж.Д., Парданани А., Васматзис Г., Теффери А. (сентябрь 2013 г.). «Идентификация субмикроскопических генетических изменений и точное картирование точек разрыва при миелофиброзе с использованием секвенирования парных пар с высоким разрешением» . Американский журнал гематологии . 88 (9): 741–6. дои : 10.1002/ajh.23495 . ПМИД 23733509 . S2CID 5232311 .
- ^ Лянь X, Дуань X, Ву X, Ли C, Чен С, Ван С, Цай Ю, Венг Z (август 2012 г.). «Экспрессия и клиническое значение нижестоящих генов фон Хиппель-Линдау: Jade-1 и β-катенин, связанных с почечно-клеточной карциномой». Урология . 80 (2): 485.e7–13. дои : 10.1016/j.urology.2012.02.024 . ПМИД 22516360 .
- ^ Лим С.Р., Гуи Б.Х., Сингх М., Гам Л.Х. (ноябрь 2011 г.). «Анализ дифференциально экспрессируемых белков при колоректальном раке с использованием колонки с гидроксиапатитом и SDS-PAGE». Прикладная биохимия и биотехнология . 165 (5–6): 1211–24. дои : 10.1007/s12010-011-9339-3 . ПМИД 21863284 . S2CID 13272576 .
- ^ Лю П.Ф., Цзян В.Х., Хан Ю.Т., Хэ Л.Ф., Чжан Х.Л., Рен Х. (28 августа 2015 г.). «Комплексный анализ микроРНК-мРНК аденокарциномы протоков поджелудочной железы» . Генетика и молекулярные исследования . 14 (3): 10288–97. doi : 10.4238/2015.28.14 августа . ПМИД 26345967 .
- ^ Фаррелл, Курт; Ким, Сунгхо; и др. (январь 2022 г.). «Полногеномное исследование ассоциаций и функциональная проверка указывают на причастность JADE1 к таупатии» . Акта Нейропатологика . 143 (1): 33–53. дои : 10.1007/s00401-021-02379-z . ПМЦ 8786260 . ПМИД 34719765 .
- ^ Крери, Джон Ф.; Трояновский, Джон К.; и др. (декабрь 2014 г.). «Первичная возрастная таупатия (ЧАСТЬ): распространенная патология, связанная со старением человека» . Акта Нейропатологика . 128 (6): 755–766. дои : 10.1007/s00401-014-1349-0 . ПМЦ 4257842 . ПМИД 25348064 .
- ^ Крери, Джон Ф.; Трояновский, Джон К.; и др. (декабрь 2014 г.). «Первичная возрастная таупатия (ЧАСТЬ): распространенная патология, связанная со старением человека» . Акта Нейропатологика . 128 (6): 755–766. дои : 10.1007/s00401-014-1349-0 . ПМЦ 4257842 . ПМИД 25348064 .
Дальнейшее чтение
[ редактировать ]- Цуанаку Э., Твиди С., Уилсон В. (декабрь 2003 г.). «Идентификация Jade1, гена, кодирующего белок цинковых пальцев PHD, в скрининге мутагенеза генной ловушки для генов, участвующих в развитии переднезадней оси» . Молекулярная и клеточная биология . 23 (23): 8553–2. дои : 10.1128/MCB.23.23.8553-8562.2003 . ПМК 262661 . ПМИД 14612400 .
- Панченко М.В., Чжоу М.И., Коэн Х.Т. (декабрь 2004 г.). «Партнер фон Хиппель-Линдау Jade-1 представляет собой коактиватор транскрипции, связанный с активностью гистон-ацетилтрансферазы» . Журнал биологической химии . 279 (53): 56032–41. дои : 10.1074/jbc.M410487200 . ПМИД 15502158 .
- Чжоу М.И., Фой Р.Л., Читалия В.К., Чжао Дж., Панченко М.В., Ван Х., Коэн Х.Т. (август 2005 г.). «Джад-1, кандидат в супрессоры опухолей почек, способствующий апоптозу» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (31): 11035–40. Бибкод : 2005PNAS..10211035Z . дои : 10.1073/pnas.0500757102 . ПМК 1182408 . ПМИД 16046545 .
- Дойон Ю, Кайру С, Улла М, Лэндри А.Дж., Коте В., Селлек В., Лейн В.С., Тан С., Ян XJ, Коте Дж. (январь 2006 г.). «Белки-супрессоры опухолей ING являются важными регуляторами ацетилирования хроматина, необходимыми для экспрессии и сохранения генома» . Молекулярная клетка . 21 (1): 51–64. doi : 10.1016/j.molcel.2005.12.007 . ПМИД 16387653 .
- Лим Дж., Хао Т., Шоу С., Патель А.Дж., Сабо Г., Руал Дж.Ф., Фиск С.Дж., Ли Н., Смоляр А., Хилл Д.Э., Барабаси А.Л., Видал М., Зогби Х.Ю. (май 2006 г.). «Сеть белок-белкового взаимодействия при наследственных атаксиях человека и нарушениях дегенерации клеток Пуркинье» . Клетка . 125 (4): 801–14. дои : 10.1016/j.cell.2006.03.032 . ПМИД 16713569 . S2CID 13709685 .
- Олсен Дж.В., Благоев Б., Гнад Ф., Мачек Б., Кумар С., Мортенсен П., Манн М. (ноябрь 2006 г.). «Глобальная, in vivo и сайт-специфическая динамика фосфорилирования в сигнальных сетях» . Клетка . 127 (3): 635–48. дои : 10.1016/j.cell.2006.09.026 . ПМИД 17081983 . S2CID 7827573 .
Внешние ссылки
[ редактировать ]- PHF17+белок+человек Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .