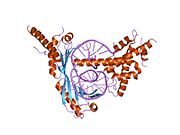
ТАТА-связывающий белок
| ТВП | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | TBP , GTF2D, GTF2D1, HDL4, SCA17, TFIID, TATA-бокс-связывающий белок, TATA-связывающий белок, TBP1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 600075 ; МГИ : 101838 ; Гомологен : 2404 ; GeneCards : TBP ; ОМА : TBP – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





| ТВП | |||
|---|---|---|---|
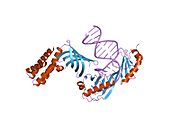
 кристаллическая структура тройного комплекса дрожжей brf1-tbp-ДНК | |||
| Идентификаторы | |||
| Символ | ТВП | ||
| Пфам | PF00352 | ||
| Пфам Клан | CL0407 | ||
| ИнтерПро | ИПР000814 | ||
| PROSITE | PDOC00303 | ||
| СКОП2 | 1 Тбп / СКОПе / СУПФАМ | ||
| |||
ТАТА -связывающий белок ( TBP ) представляет собой общий фактор транскрипции , который связывается с последовательностью ДНК, называемой ТАТА-боксом . Эта последовательность ДНК находится примерно на 30 пар оснований выше места начала транскрипции в некоторых эукариотических генов промоторах . [5]
Семейство генов ТБФ
[ редактировать ]ТБФ является членом небольшого семейства генов факторов, связанных с ТБФ. [6] Первый фактор, связанный с ТБФ (TRF/TRF1), был идентифицирован у плодовой мухи Drosophila , но, по-видимому, он специфичен для мух или насекомых. Впоследствии TBPL1 /TRF2 был обнаружен в геномах многих многоклеточных животных , тогда как геномы позвоночных кодируют третьего члена семейства позвоночных, TBPL2 /TRF3. В определенных типах клеток или на определенных промоторах ТБФ может быть заменен одним из этих связанных с ТБФ факторов, некоторые из которых взаимодействуют с ТАТА-боксом аналогично ТБФ.
Роль фактора транскрипции
[ редактировать ]TBP является субъединицей общего фактора транскрипции эукариот TFIID . TFIID является первым белком, который связывается с ДНК во время образования комплекса преинициации транскрипции РНК -полимеразы II (РНК Пол II). [7] Являясь одним из немногих белков в преинициационном комплексе, который связывает ДНК специфичным для последовательности образом, он помогает позиционировать РНК-полимеразу II над сайтом начала транскрипции гена. Однако, по оценкам, только 10–20% человеческих промоторов имеют TATA-боксы - большинство человеческих промоторов представляют собой TATA без генов домашнего хозяйства промоторы - поэтому TBP, вероятно, не единственный белок, участвующий в позиционировании РНК-полимеразы II. ТБФ к этим промоторам обеспечивается регуляторами генов «домашнего хозяйства». [8] [9] Интересно, что транскрипция инициируется в узкой области примерно на 30 п.н. ниже ТАТА-бокса на ТАТА-содержащих промоторах. [10] тогда как сайты начала транскрипции промоторов без ТАТА рассредоточены в пределах области длиной 200 п.н. [11] [9]
Связывание TFIID с ТАТА-полем в промоторной области гена инициирует рекрутирование других факторов, необходимых для начала транскрипции РНК Pol II. Некоторые из других рекрутированных факторов транскрипции включают TFIIA , TFIIB и TFIIF . Каждый из этих факторов транскрипции содержит несколько белковых субъединиц.
ТБФ также важен для транскрипции РНК-полимеразой I и РНК-полимеразой III и, следовательно, участвует в инициации транскрипции всеми тремя РНК-полимеразами. [12]
ТБФ участвует в плавлении ДНК (двухцепочечное разделение), изгибая ДНК на 80° (богатая АТ последовательность, с которой он связывается, облегчает плавление). TBP — необычный белок, поскольку он связывает малую бороздку с помощью β-листа.
Еще одной отличительной особенностью ТБФ является длинная цепочка глютаминов на N-конце белка. Эта область модулирует ДНК-связывающую активность С-конца, а модуляция ДНК-связывания влияет на скорость образования транскрипционного комплекса и инициацию транскрипции. Мутации, которые увеличивают количество CAG-повторов, кодирующих этот полиглутаминовый тракт , и, таким образом, увеличивают длину полиглутаминовой цепочки, связаны со спиноцеребеллярной атаксией 17, нейродегенеративным заболеванием, классифицируемым как полиглутаминовая болезнь . [13]
ДНК-белковые взаимодействия
[ редактировать ]Когда ТБФ связывается с ТАТА-боксом внутри ДНК , он искажает ДНК, вставляя боковые цепи аминокислот между парами оснований, частично раскручивая спираль и дважды перекручивая ее. Искажение достигается за счет большого количества поверхностных контактов между белком и ДНК. ТБФ связывается с отрицательно заряженными фосфатами в основной цепи ДНК через положительно заряженные лизина и аргинина аминокислотные остатки . Резкий изгиб ДНК возникает за счет проецирования четырех объемистых остатков фенилаланина в малую бороздку. По мере изгиба ДНК ее контакт с ТБФ увеличивается, тем самым усиливая взаимодействие ДНК-белок.
Напряжение, приложенное к ДНК в результате этого взаимодействия, инициирует плавление или разделение нитей. Поскольку эта область ДНК богата остатками аденина и тимина , которые соединяются в пары оснований только через две водородные связи , нити ДНК легче разделяются. Разделение двух цепей обнажает основания и позволяет РНК-полимеразе II начать транскрипцию гена .
С-конец ТБФ имеет спиральную форму, которая (неполностью) дополняет ТАТА-участок ДНК. Эта незавершенность позволяет ДНК пассивно склоняться к связыванию.
Информацию об использовании ТБФ в клетках см.: РНК-полимераза I , РНК-полимераза II и РНК-полимераза III .
Белково-белковые взаимодействия
[ редактировать ]Было показано, что ТАТА-связывающий белок взаимодействует с:
- БРФ1 , [14] [15]
- БТАФ1 , [16] [17]
- Си-Фос , [18]
- Си-джун , [19]
- ЭДФ1 , [20] [21] [22]
- ГТФ2Б (ТФИИБ), [23] [24]
- GTF2A1 ( субъединица 1 TFIIA ), [23] [25] [26] [27]
- GTF2F1 ( субъединица 1 TFIIF ) [16] [28] [29]
- GTF2H4 ( субъединица 4 TFIIH ), [16]
- Мдм2 , [30] [31]
- MSX1 , [32] [33] [34]
- НФЮБ , [35]
- Р53 , [36] [37]
- ПАКС6 , [37]
- ПОЛР2А , [16]
- ПОУ2Ф1 , [38]
- РЕЛА , [39] [40]
- НР2Б1 , [41]
- ТАФ1 , [35] [42] [43] [44]
- ТАФ4 , [45]
- ТАФ5 , [35] [43] [46]
- ТАФ6 , [35] [43] [45]
- ТАФ7 , [35] [45]
- ТАФ9 . [35] [47]
- ТАФ10 , [35] [43]
- ТАФ11 , [35] [48] [49]
- ТАФ13 , [48] и
- ТАФ15 . [50]
Сложная сборка
[ редактировать ]ТАТА-бокс , связывающий Белок (TBP), необходим для инициации транскрипции РНК- полимеразами I, II и III с промоторов с ТАТА-боксом или без него. [51] [52] В присутствии промотора без ТАТА ТБФ связывается с помощью ТБФ-ассоциированных факторов (TAF). [53] [54] TBP связывается с множеством факторов, включая общие факторы транскрипции TFIIA, -B, -D, -E и -H, с образованием огромных многосубъединичных пре-инициационных комплексов на коровом промоторе . Благодаря ассоциации с различными факторами транскрипции ТБФ может инициировать транскрипцию с разных РНК- полимераз . Существует несколько родственных TBP, включая TBP-подобные (TBPL) белки . [55]
Структура
[ редактировать ]С-концевое ядро ТБФ (~ 180 остатков) высококонсервативно и по 88 аминокислот содержит два повтора , которые образуют седловидную структуру , охватывающую ДНК; эта область связывается с ТАТА-боксом и взаимодействует с факторами транскрипции и регуляторными белками . [56] Напротив, N-концевая область различается как по длине, так и по последовательности .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000112592 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000014767 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Корнберг Р.Д. (2007). «Молекулярные основы транскрипции эукариот» . Учеб. Натл. акад. наук. США . 104 (32): 12955–61. Бибкод : 2007PNAS..10412955K . дои : 10.1073/pnas.0704138104 . ЧВК 1941834 . ПМИД 17670940 .
- ^ Ахтар В., Винстра Г.Дж. (1 января 2011 г.). «Факторы, связанные с TBP: парадигма разнообразия в инициации транскрипции» . Клетка и биологические науки . 1 (1): 23. дои : 10.1186/2045-3701-1-23 . ПМЦ 3142196 . ПМИД 21711503 .
- ^ Ли Т.И., Молодой РА (2000). «Транскрипция генов, кодирующих эукариотические белки». Ежегодный обзор генетики . 34 : 77–137. дои : 10.1146/annurev.genet.34.1.77 . ПМИД 11092823 .
- ^ Лам К.К., Мюльпфордт Ф., Вакерисас Дж.М., Раджа С.Дж., Хольц Х., Ласкомб Н.М., Манке Т., Ахтар А. (2012). «Комплекс NSL регулирует гены домашнего хозяйства у дрозофилы» . ПЛОС Генетика . 8 (6): e1002736. дои : 10.1371/journal.pgen.1002736 . ПМЦ 3375229 . ПМИД 22723752 .
- ^ Перейти обратно: а б Лам К.К., Чунг Х.Р., Семпличио Г., Айер С.С., Гауб А., Бхардвадж В., Хольц Х., Георгиев П., Ахтар А. (февраль 2019 г.). «Нуклеосомный ландшафт, опосредованный комплексом NSL, необходим для поддержания точности транскрипции и подавления транскрипционного шума» . Гены и развитие . 33 (7–8): 452–465. дои : 10.1101/gad.321489.118 . ПМК 6446542 . ПМИД 30819819 .
- ^ Карнинчи П., Санделин А., Ленхард Б., Катаяма С., Симокава К., Поньявич Дж. и др. (июнь 2006 г.). «Полногеномный анализ архитектуры и эволюции промотора млекопитающих». Природная генетика . 38 (6): 626–35. дои : 10.1038/ng1789 . ПМИД 16645617 . S2CID 22205897 .
- ^ Ни Т., Коркоран Д.Л., Рэч Э.А., Сонг С., Спана Э.П., Гао Ю., Олер У., Чжу Дж. (июль 2010 г.). «Стратегия парного секвенирования для картирования сложного ландшафта инициации транскрипции» . Природные методы . 7 (7): 521–7. дои : 10.1038/nmeth.1464 . ПМК 3197272 . ПМИД 20495556 .
- ^ Ваннини А., Крамер П. (февраль 2012 г.). «Консервация между механизмами инициации транскрипции РНК-полимеразой I, II и III» . Молекулярная клетка . 45 (4): 439–46. doi : 10.1016/j.molcel.2012.01.023 . hdl : 11858/00-001M-0000-0015-483A-5 . ПМИД 22365827 .
- ^ «Ген Энтрез: белок, связывающий ТАТА-бокс TBP» .
- ^ Маккалок В., Хардин П., Пэн В., Рупперт Дж. М., Лобо-Рупперт С. М. (август 2000 г.). «Альтернативно сплайсированные варианты hBRF функционируют на разных промоторах РНК-полимеразы III» . ЭМБО Дж . 19 (15): 4134–43. дои : 10.1093/emboj/19.15.4134 . ПМК 306597 . ПМИД 10921893 .
- ^ Ван З, Рёдер Р.Г. (июль 1995 г.). «Структура и функция субъединицы человеческого транскрипционного фактора TFIIIB, которая является эволюционно консервативной и содержит как TFIIB-, так и домены, связанные с белком 2 группы с высокой подвижностью» . Учеб. Натл. акад. наук. США . 92 (15): 7026–30. Бибкод : 1995PNAS...92.7026W . дои : 10.1073/pnas.92.15.7026 . ПМК 41464 . ПМИД 7624363 .
- ^ Перейти обратно: а б с д Скалли Р., Андерсон С.Ф., Чао Д.М., Вэй В., Йе Л., Янг Р.А., Ливингстон Д.М., Парвин Дж.Д. (май 1997 г.). «BRCA1 является компонентом голофермента РНК-полимеразы II» . Учеб. Натл. акад. наук. США . 94 (11): 5605–10. Бибкод : 1997PNAS...94.5605S . дои : 10.1073/pnas.94.11.5605 . ПМК 20825 . ПМИД 9159119 .
- ^ Чикка Джей-Джей, Обл Д.Т., Пью Б.Ф. (март 1998 г.). «Клонирование и биохимическая характеристика TAF-172, человеческого гомолога дрожжевого Mot1» . Мол. Клетка. Биол . 18 (3): 1701–10. дои : 10.1128/MCB.18.3.1701 . ПМЦ 108885 . ПМИД 9488487 .
- ^ Мец Р., Баннистер А.Дж., Сазерленд Дж.А., Хагемайер С., О'Рурк ЕС, Кук А., Браво Р., Кузаридес Т. (сентябрь 1994 г.). «c-Fos-индуцированная активация промотора, содержащего ТАТА-бокс, включает прямой контакт с белком, связывающим ТАТА-бокс» . Мол. Клетка. Биол . 14 (9): 6021–9. дои : 10.1128/MCB.14.9.6021 . ПМК 359128 . ПМИД 8065335 .
- ^ Франклин CC, Маккалок А.В., Kraft AS (февраль 1995 г.). «In vitro ассоциация между семейством белков Jun и общими факторами транскрипции, TBP и TFIIB» . Биохим. Дж . 305 (3): 967–74. дои : 10.1042/bj3050967 . ПМК 1136352 . ПМИД 7848298 .
- ^ Брендель С., Гельман Л., Ауверкс Дж. (июнь 2002 г.). «Мультипротеиновый мостик-фактор-1 (MBF-1) является кофактором ядерных рецепторов, которые регулируют липидный обмен» . Мол. Эндокринол . 16 (6): 1367–77. дои : 10.1210/mend.16.6.0843 . ПМИД 12040021 .
- ^ Мариотти М., Де Бенедиктис Л., Эйвон Э., Майер Дж.А. (август 2000 г.). «Взаимодействие между фактором-1, связанным с дифференцировкой эндотелия, и кальмодулином in vitro и in vivo» . Ж. Биол. Хим . 275 (31): 24047–51. дои : 10.1074/jbc.M001928200 . ПМИД 10816571 .
- ^ Кабе Ю, Гото М, Сима Д, Имаи Т, Вада Т, Морохаши Ки, Сиракава М, Хиросе С, Ханда Х (ноябрь 1999 г.). «Роль человеческого MBF1 как коактиватора транскрипции» . Ж. Биол. Хим . 274 (48): 34196–202. дои : 10.1074/jbc.274.48.34196 . ПМИД 10567391 .
- ^ Перейти обратно: а б Тан Х, Сунь Х, Рейнберг Д., Эбрайт Р.Х. (февраль 1996 г.). «Белко-белковые взаимодействия при инициации транскрипции эукариот: структура преинициаторного комплекса» . Учеб. Натл. акад. наук. США . 93 (3): 1119–24. Бибкод : 1996PNAS...93.1119T . дои : 10.1073/pnas.93.3.1119 . ПМК 40041 . ПМИД 8577725 .
- ^ Бушнелл Д.А., Вестовер К.Д., Дэвис Р.Э., Корнберг Р.Д. (февраль 2004 г.). «Структурная основа транскрипции: сокристалл РНК-полимеразы II-TFIIB с длиной 4,5 ангстрем». Наука . 303 (5660): 983–8. Бибкод : 2004Sci...303..983B . дои : 10.1126/science.1090838 . ПМИД 14963322 . S2CID 36598301 .
- ^ ДеДжонг Дж., Бернштейн Р., Редер Р.Г. (апрель 1995 г.). «Общий фактор транскрипции человека TFIIA: характеристика кДНК, кодирующей малую субъединицу, и потребность в базальной и активированной транскрипции» . Учеб. Натл. акад. наук. США . 92 (8): 3313–7. Бибкод : 1995PNAS...92.3313D . дои : 10.1073/pnas.92.8.3313 . ПМК 42156 . ПМИД 7724559 .
- ^ Озер Дж., Мицурас К., Зерби Д., Кэри М., Либерман П.М. (июнь 1998 г.). «Фактор транскрипции IIA подавляет связанное с ТАТА-связывающим белком (TBP) факторное ингибирование связывания TBP с ДНК» . Ж. Биол. Хим . 273 (23): 14293–300. дои : 10.1074/jbc.273.23.14293 . ПМИД 9603936 .
- ^ Сунь X, Ма Д., Шелдон М., Юнг К., Рейнберг Д. (октябрь 1994 г.). «Восстановление активности TFIIA человека из рекомбинантных полипептидов: роль в транскрипции, опосредованной TFIID» . Генс Дев . 8 (19): 2336–48. дои : 10.1101/gad.8.19.2336 . ПМИД 7958900 .
- ^ Руперт С., Тиан Р. (ноябрь 1995 г.). «Человеческий TAFII250 взаимодействует с RAP74: значение для инициации РНК-полимеразы II» . Генс Дев . 9 (22): 2747–55. дои : 10.1101/gad.9.22.2747 . ПМИД 7590250 .
- ^ Малик С., Гермах М., Редер Р.Г. (март 1998 г.). «Динамическая модель функции коактиватора PC4 в транскрипции РНК-полимеразы II» . Учеб. Натл. акад. наук. США . 95 (5): 2192–7. Бибкод : 1998PNAS...95.2192M . дои : 10.1073/pnas.95.5.2192 . ЧВК 19292 . ПМИД 9482861 .
- ^ Тут С.Дж., Гудрич Дж.А., Тиан Р. (август 1997 г.). «Репрессия p53-опосредованной транскрипции с помощью MDM2: двойной механизм» . Генс Дев . 11 (15): 1974–86. дои : 10.1101/гад.15.11.1974 . ПМК 316412 . ПМИД 9271120 .
- ^ Левейяр Т., Василик Б. (декабрь 1997 г.). «С-концевая область MDM2 связывается с TAFII250 и необходима для регуляции MDM2 промотора циклина А» . Ж. Биол. Хим . 272 (49): 30651–61. дои : 10.1074/jbc.272.49.30651 . ПМИД 9388200 .
- ^ Шетти С., Такахаси Т., Мацуи Х., Айенгар Р., Рагоу Р. (май 1999 г.). «Транскрипционная ауторепрессия гена Msx1 опосредуется взаимодействием белка Msx1 с мультибелковым транскрипционным комплексом, содержащим ТАТА-связывающий белок, Sp1 и белок, связывающий элемент ответа цАМФ (CBP/p300)» . Биохим. Дж . 339 (3): 751–8. дои : 10.1042/0264-6021:3390751 . ПМК 1220213 . ПМИД 10215616 .
- ^ Чжан Х., Ху Г., Ван Х., Скиаволино П., Илер Н., Шен М.М., Абате-Шен С. (май 1997 г.). «Гетеродимеризация гомеопротеинов Msx и Dlx приводит к функциональному антагонизму» . Мол. Клетка. Биол . 17 (5): 2920–32. дои : 10.1128/mcb.17.5.2920 . ПМК 232144 . ПМИД 9111364 .
- ^ Чжан Х., Катрон К.М., Абате-Шен С. (март 1996 г.). «Роль гомеодомена Msx-1 в регуляции транскрипции: остатки в N-концевом плече опосредуют взаимодействие ТАТА-связывающего белка и репрессию транскрипции» . Учеб. Натл. акад. наук. США . 93 (5): 1764–9. Бибкод : 1996PNAS...93.1764Z . дои : 10.1073/pnas.93.5.1764 . ПМК 39855 . ПМИД 8700832 .
- ^ Перейти обратно: а б с д и ж г час Беллорини М., Ли Д.К., Дантонель Дж.К., Земзуми К., Редер Р.Г., Тора Л., Мантовани Р. (июнь 1997 г.). «Взаимодействия CCAAT, связывающие NF-Y-TBP: NF-YB и NF-YC требуют коротких доменов, прилегающих к их мотивам гистоновых складок, для ассоциации с основными остатками TBP» . Нуклеиновые кислоты Рез . 25 (11): 2174–81. дои : 10.1093/нар/25.11.2174 . ПМК 146709 . ПМИД 9153318 .
- ^ Сето Э., Ушева А., Замбетти Г.П., Моманд Дж., Хорикоши Н., Вайнманн Р., Левин А.Дж., Шенк Т. (декабрь 1992 г.). «Р53 дикого типа связывается с ТАТА-связывающим белком и подавляет транскрипцию» . Учеб. Натл. акад. наук. США . 89 (24): 12028–32. Бибкод : 1992PNAS...8912028S . дои : 10.1073/pnas.89.24.12028 . ПМК 50691 . ПМИД 1465435 .
- ^ Перейти обратно: а б Цвекл А., Кашанчи Ф., Брейди Дж. Н., Пятигорский Дж. (июнь 1999 г.). «Взаимодействие Pax-6 с белком, связывающим ТАТА-бокс, и белком ретинобластомы». Инвестируйте. Офтальмол. Вис. Наука . 40 (7): 1343–50. ПМИД 10359315 .
- ^ Цвиллинг С., Аннвайлер А., Вирт Т. (май 1994 г.). «Домены POU факторов транскрипции Oct1 и Oct2 опосредуют специфическое взаимодействие с TBP» . Нуклеиновые кислоты Рез . 22 (9): 1655–62. дои : 10.1093/нар/22.9.1655 . ПМК 308045 . ПМИД 8202368 .
- ^ Гермах М., Малик С., Редер Р.Г. (июнь 1998 г.). «Участие компонентов TFIID и США в активации транскрипции промотора вируса иммунодефицита человека с помощью NF-kappaB и Sp1» . Мол. Клетка. Биол . 18 (6): 3234–44. дои : 10.1128/mcb.18.6.3234 . ПМЦ 108905 . ПМИД 9584164 .
- ^ Шмитц М.Л., Стельцер Г., Альтманн Х., Майстернст М., Бауерле П.А. (март 1995 г.). «Взаимодействие COOH-концевого домена трансактивации p65 NF-каппа B с ТАТА-связывающим белком, транскрипционным фактором IIB и коактиваторами» . Ж. Биол. Хим . 270 (13): 7219–26. дои : 10.1074/jbc.270.13.7219 . ПМИД 7706261 .
- ^ Шульман И.Г., Чакраварти Д., Жугилон Х., Ромо А., Эванс Р.М. (август 1995 г.). «Взаимодействия между ретиноидным рецептором X и консервативной областью ТАТА-связывающего белка опосредуют гормонозависимую трансактивацию» . Учеб. Натл. акад. наук. США . 92 (18): 8288–92. Бибкод : 1995PNAS...92.8288S . дои : 10.1073/pnas.92.18.8288 . ПМК 41142 . ПМИД 7667283 .
- ^ Зигерт Дж.Л., Роббинс П.Д. (январь 1999 г.). «Rb ингибирует внутреннюю киназную активность фактора TAFII250, связанного с ТАТА-связывающим белком» . Мол. Клетка. Биол . 19 (1): 846–54. дои : 10.1128/MCB.19.1.846 . ПМК 83941 . ПМИД 9858607 .
- ^ Перейти обратно: а б с д Руперт С., Ван Э.Х., Цзян Р. (март 1993 г.). «Клонирование и экспрессия человеческого TAFII250: фактор, связанный с ТБФ, участвующий в регуляции клеточного цикла» . Природа . 362 (6416): 175–9. Бибкод : 1993Natur.362..175R . дои : 10.1038/362175a0 . ПМИД 7680771 . S2CID 4364676 .
- ^ О'Брайен Т., Тиан Р. (май 1998 г.). «Функциональный анализ N-концевого киназного домена TAFII250 человека» . Мол. Клетка . 1 (6): 905–11. дои : 10.1016/S1097-2765(00)80089-1 . ПМИД 9660973 .
- ^ Перейти обратно: а б с Пуантуд Х.К., Менгус Дж., Бранкорсини С., Монако Л., Парвинен М., Сассоне-Корси П., Дэвидсон I (май 2003 г.). «Внутриклеточная локализация TAF7L, паралога субъединицы TFIID фактора транскрипции TAF7, регулируется в процессе развития во время дифференцировки мужских зародышевых клеток». Дж. Клеточная наука . 116 (Часть 9): 1847–58. дои : 10.1242/jcs.00391 . ПМИД 12665565 . S2CID 24519687 .
- ^ Тао Й., Гермах М., Мартинес Э., Эльгешлегер Т., Хасегава С., Такада Р., Ямамото Т., Хорикоши М., Рёдер Р.Г. (март 1997 г.). «Специфические взаимодействия и потенциальные функции человеческого TAFII100» . Ж. Биол. Хим . 272 (10): 6714–21. дои : 10.1074/jbc.272.10.6714 . ПМИД 9045704 .
- ^ Мартинес Э., Палхан В.Б., Тьернберг А., Лимар Э.С., Гампер А.М., Кунду Т.К., Хаит Б.Т., Рёдер Р.Г. (октябрь 2001 г.). «Комплекс STAGA человека представляет собой коактиватор транскрипции, ацетилирующий хроматин, который взаимодействует с факторами сплайсинга пре-мРНК и факторами, связывающими повреждение ДНК, in vivo» . Мол. Клетка. Биол . 21 (20): 6782–95. дои : 10.1128/MCB.21.20.6782-6795.2001 . ПМК 99856 . ПМИД 11564863 .
- ^ Перейти обратно: а б Менгус Дж., Мэй М., Жак Икс, Стауб А., Тора Л., Шамбон П., Дэвидсон I (апрель 1995 г.). «Клонирование и характеристика hTAFII18, hTAFII20 и hTAFII28: три субъединицы человеческого транскрипционного фактора TFIID» . ЭМБО Дж . 14 (7): 1520–31. дои : 10.1002/j.1460-2075.1995.tb07138.x . ПМЦ 398239 . ПМИД 7729427 .
- ^ Мэй М., Менгус Г., Лавин А.С., Шамбон П., Дэвидсон И. (июнь 1996 г.). «Человеческий TAF(II28) способствует стимуляции транскрипции посредством активации функции 2 ретиноидных X-рецепторов» . ЭМБО Дж . 15 (12): 3093–104. дои : 10.1002/j.1460-2075.1996.tb00672.x . ПМК 450252 . ПМИД 8670810 .
- ^ Хоффманн А., Редер Р.Г. (июль 1996 г.). «Клонирование и характеристика человеческого TAF20/15. Множественные взаимодействия предполагают центральную роль в формировании комплекса TFIID» . Ж. Биол. Хим . 271 (30): 18194–202. дои : 10.1074/jbc.271.30.18194 . ПМИД 8663456 .
- ^ Хоххаймер А., Тиан Р. (июнь 2003 г.). «Диверсифицированные комплексы инициации транскрипции расширяют селективность промотора и тканеспецифическую экспрессию генов» . Гены и развитие . 17 (11): 1309–20. дои : 10.1101/gad.1099903 . ПМИД 12782648 .
- ^ Пью Б.Ф. (сентябрь 2000 г.). «Контроль экспрессии генов посредством регуляции ТАТА-связывающего белка». Джин . 255 (1): 1–14. дои : 10.1016/s0378-1119(00)00288-2 . ПМИД 10974559 .
- ^ Уивер, Роберт Франклин (1 января 2012 г.). Молекулярная биология . МакГроу-Хилл. ISBN 9780073525327 . OCLC 789601172 .
- ^ Громче Р.К., Хе Ю., Лопес-Бланко-младший, Фанг Дж., Чакон П., Ногалес Э. (март 2016 г.). «Структура TFIID, связанного с промотором, и модель сборки прединициаторного комплекса человека» . Природа . 531 (7596): 604–9. Бибкод : 2016Natur.531..604L . дои : 10.1038/nature17394 . ПМЦ 4856295 . ПМИД 27007846 .
- ^ Дэвидсон I (июль 2003 г.). «Генетика ТБФ и факторов, связанных с ТБФ». Тенденции биохимических наук . 28 (7): 391–8. дои : 10.1016/S0968-0004(03)00117-8 . ПМИД 12878007 .
- ^ Николов Д.Б., Ху Ш., Лин Дж., Гаш А., Хоффманн А., Хорикоши М., Чуа Н.Х., Редер Р.Г., Берли С.К. (ноябрь 1992 г.). «Кристаллическая структура белка, связывающего ТАТА-бокс TFIID». Природа . 360 (6399): 40–6. Бибкод : 1992Natur.360...40N . дои : 10.1038/360040a0 . ПМИД 1436073 . S2CID 4307043 .
Внешние ссылки
[ редактировать ]- Запись GeneReviews/NCBI/NIH/UW о спиноцеребеллярной атаксии типа 17
- Интерактивная структура TBP на сайте Massey.ac.nz
- PDB Молекула месяца ТАТА-связывающий белок
- TATA-Box+Binding+Protein в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ФакторБук ТВП
- Обзор всей структурной информации, доступной в PDB для UniProt : P20226 (человеческий TATA-бокс-связывающий белок) на PDBe-KB .
галерея PDB |
|---|