Рев-Эрба альфа
| НР1Д1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | NR1D1 , EAR1, THRA1, THRAL, Ear-1, hRev, подсемейство ядерных рецепторов 1, член группы D 1, REVERBA, REVERBalpha | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 602408 ; МГИ : 2444210 ; Гомологен : 23324 ; Генные карты : NR1D1 ; ОМА : NR1D1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




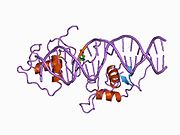
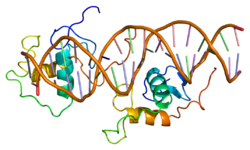
Rev-Erb альфа (Rev-Erbɑ) , также известный как член 1 группы D подсемейства 1 ядерных рецепторов (NR1D1), является одним из двух белков Rev-Erb в семействе ядерных рецепторов (NR) внутриклеточных факторов транскрипции . У людей REV-ERBɑ кодируется геном NR1D1 , который высоко консервативен у разных видов животных. [5]
Rev-Erbɑ играет важную роль в регуляции основных циркадных часов посредством репрессии элемента положительных часов Bmal1 . Он также регулирует несколько физиологических процессов под циркадным контролем, включая метаболические и иммунные пути. [6] [7] Rev-Erbɑ мРНК демонстрирует циркадные колебания в своей экспрессии, и она высоко экспрессируется у млекопитающих в мозге и метаболических тканях, таких как скелетные мышцы , жировая ткань и печень . [6] [8]
Открытие
[ редактировать ]Rev-Erbɑ был открыт в 1989 году Нобуюки Миядзимой и его коллегами, которые идентифицировали два erbA гомолога на хромосоме 17 человека , которые транскрибировались с противоположных цепей ДНК в одном и том же локусе . Один из генов кодировал белок, очень похожий на рецептор гормона щитовидной железы курицы , а другой, который они назвали Ear-1 , позже будет описан как Rev-Erbɑ. [9] Впервые белок был упомянут под названием Rev-Erbɑ в 1990 году Митчеллом А. Лазаром , Карен Э. Джонс и Уильямом В. Чином, которые выделили комплементарную ДНК скелетных мышц плода человека Rev-Erbɑ из библиотеки . Подобно гену у крыс, они обнаружили, что человеческий Rev-Erbɑ транскрибируется с цепи, противоположной альфа-рецептору гормона щитовидной железы человека (THRA, c-erbAα) . [10]
Rev-Erbɑ впервые был задействован в циркадном контроле в 1998 году, когда Аурелио Бальсалобре, Франческа Дамиола и Ули Шиблер продемонстрировали, что экспрессия Rev-Erbɑ в фибробластах крыс демонстрирует суточные ритмы. [11] Rev-Erbɑ был впервые идентифицирован как ключевой игрок в петле обратной связи трансляции транскрипции (TTFL) в 2002 году, когда эксперименты показали, что Rev-Erbɑ действует, подавляя транскрипцию гена Bmal1 , а экспрессия Rev-Erbɑ контролируется другими компонентами TTFL. Это сделало Rev-Erbɑ связующим звеном между положительным и отрицательным контурами TTFL. [12]
Генетика и эволюция
[ редактировать ]Ген NR1D1 (член 1 группы D подсемейства 1 ядерных рецепторов), расположенный на хромосоме 17 , кодирует белок REV-ERBɑ у человека. Он транскрибируется с противоположной цепи альфа-рецептора гормона щитовидной железы человека (THRA, c-erbAα), так что кДНК NR1D1 и THRA комплементарны по 269 основаниям. [10] Ген состоит из 7797 оснований с 8 экзонами , образующими только 1 сплайсинговый вариант . [5] Сам промотор NR1D1 содержит ответный элемент REV-ERB (RevRE), который позволяет регулировать экспрессию генов как посредством ауторегуляции , так и регуляции с помощью орфанного рецептора альфа, связанного с рецептором ретиноевой кислоты (RORɑ) , другого фактора транскрипции ядерного рецептора. [8] NR1D1 также содержит E-box на своем промоторе , который позволяет регулировать его с помощью BMAL1 . У людей NR1D1 (REV-ERBɑ) высоко экспрессируется в головном мозге и метаболических тканях, включая скелетные мышцы, жировую ткань и печень. [8] [6]
Геномный анализ показывает, что ген NR1D1 присутствовал у самого недавнего общего предка всех животных, а ортологи присутствовали у 378 протестированных видов, включая шимпанзе, собак, мышей, крыс, кур, рыбок данио, лягушек и плодовых мух. [13] Сравнение с крысиным ортологом, Nr1d1 , указывает на высокую консервативность связывающих ДНК и карбокси-концевых доменов , а также сохранение транскрипции c-erbA альфа-2 и Rev-Erbɑ на противоположных цепях. [10] У человека NR1D1 имеет только один паралог , NR1D2 ( REV-ERBβ ), который расположен на хромосоме 3 и, вероятно, возник в результате дупликации . [14] Однако и NR1D1 , и NR1D2 являются членами семейства ядерных рецепторов, что указывает на то, что они имеют общее происхождение. Таким образом, NR1D1 функционально связан с другими генами ядерных рецепторов, такими как дельта-рецептор, активирующий пролифератор пероксисом ( PPARD ) и альфа-рецептор ретиноевой кислоты ( RARA ) . [13] Более того, исследования показали, что генетический локус NR1D1/THRA генетически связан с геном RARA . [6] [15]
Структура белка
[ редактировать ]человека Ген NR1D1 продуцирует белковый продукт (REV-ERBα) из 614 аминокислот . [5] REV-ERBα имеет 3 основных функциональных домена, включая ДНК-связывающий домен (DBD) и лиганд-связывающий домен (LBD) на С-конце, а также N-концевой домен, который позволяет модулировать активность. [16] [17] Эти три домена являются общей чертой белков ядерных рецепторов. [8]
Белки Rev-Erb уникальны среди других ядерных рецепторов тем, что они не имеют спирали на С-конце, которая необходима для рекрутирования и активации коактиватора ядерными рецепторами через их LBD. [18] Вместо этого Rev-Erbα взаимодействует через свой LBD с ко-репрессором ядерных рецепторов (NCoR) и другим близкородственным ко-репрессором, медиатором сайленсинга ретиноидных и тиреоидных рецепторов (SMRT) , хотя взаимодействие с NCoR более сильное из-за его структурной совместимости. [18] Гем , эндогенный лиганд Rev-Erbα, дополнительно стабилизирует взаимодействие с NCoR. [18] [8] Репрессия с помощью Rev-Erbα также требует взаимодействия с комплексом гистондеактилаза 3 класса I (HDAC3) - NCoR. Каталитическая активность HDAC3 активируется только тогда, когда он образует комплекс с NCoR или SMRT, поэтому Rev-Erbα должен взаимодействовать с этим комплексом, чтобы произошла репрессия генов посредством деацетилирования гистонов. [6] Пока неизвестно, играют ли другие HDAC роль в функции Rev-Erbα. [6] Rev-Erbα рекрутирует комплекс NCoR-HDAC3 посредством связывания специфической последовательности ДНК, обычно называемой RORE, из-за ее взаимодействия с активатором транскрипции - орфанным рецептором, связанным с рецептором ретиноевой кислоты (ROR) . Эта последовательность состоит из полусайта «AGGTCA», которому предшествует последовательность A/T. [18] Rev-Erbα связывается с большой бороздкой этой последовательности через свой домен DBD, который содержит два цинковых пальца типа C4 . [18] Rev-Erbα может подавлять активацию гена в качестве мономера посредством конкурентного связывания в этом сайте RORE, но для взаимодействия с NCoR и активной репрессии гена необходимы две молекулы Rev-Erbα. Это может происходить за счет связывания двух молекул Rev-Erbα с отдельными RORE или в результате более сильного взаимодействия посредством связывания отвечающего элемента , который представляет собой прямой повтор RORE (RevDR2). [18]
У мышей было показано, что N-концевой регуляторный домен содержит важный сайт фосфорилирования казеинкиназой 1 эпсилон (Csnk1e) , который способствует правильной локализации Rev-Erbα, и, кроме того, что этот домен необходим для активации ген белка щелевого соединения 1 ( GJA1 ) . [19] [20]
Функция
[ редактировать ]
Циркадный осциллятор
[ редактировать ]Было предложено, что Rev-Erbα координирует циркадные метаболические реакции. [21] Циркадные ритмы управляются взаимосвязанными регуляторными петлями транскрипции/трансляции (TTFL) , которые генерируют и поддерживают эти ежедневные ритмы, а Rev-Erbα участвует во вторичном TTFL у млекопитающих. Первичный TTFL содержит белки-активаторы транскрипции CLOCK и BMAL1 , которые способствуют ритмической экспрессии генов внутри этой петли, особенно per и крик . [22] Экспрессия этих генов затем действует посредством отрицательной обратной связи, ингибируя транскрипцию CLOCK:BMAL1. [12] Считается , что вторичный TTFL, в котором Rev-Erbα работает совместно с Rev-Erbβ и сиротским рецептором RORα , усиливает этот первичный TTFL за счет дальнейшей регуляции BMAL1. [23] RORα имеет те же элементы ответа, что и Rev-Erbα, но оказывает противоположный эффект на транскрипцию генов; Экспрессия BMAL1 подавляется Rev-Erbα и активируется RORα. [24] Экспрессия CLOCK:BMAL1 активирует транскрипцию NR1D1 , кодирующего белок Rev-Erbα. Повышенная экспрессия Rev-Erbα, в свою очередь, подавляет транскрипцию BMAL1, стабилизируя петлю. [25] Колеблющаяся экспрессия RORα и Rev-Erbα в супрахиазматическом ядре , главном циркадном хранителе времени у млекопитающих. [26] приводит к циркадному характеру экспрессии BMAL1. Занятость промотора BMAL1 этими двумя рецепторами является ключом к правильному синхронизации механизма основных часов у млекопитающих. [21]
Метаболизм
[ редактировать ]Rev-erbα играет роль в регуляции метаболизма всего организма посредством контроля липидного обмена , метаболизма желчных кислот и метаболизма глюкозы . [27] Rev-Erbα передает циркадные сигналы в метаболические и воспалительные регуляторные реакции и наоборот, хотя точные механизмы, лежащие в основе этой взаимосвязи, до конца не изучены. [21]
Rev-erbα регулирует экспрессию аполипопротеинов печени , белка, связывающего регуляторные элементы стерола жирных кислот , и элонгазы elovl3 посредством своей репрессивной активности. [28] [29] [30] Кроме того, подавление Rev-erbα связано со снижением уровня синтазы жирных кислот, ключевого регулятора липогенеза . [30] У мышей с дефицитом Rev-erbα наблюдается дислипидемия из-за повышенного уровня триглицеридов. [31] и полиморфизмы Rev-erbα у людей связаны с ожирением. [32] Rev-erbα также регулирует адипогенез белых и коричневых адипоцитов. [9] Транскрипция Rev-Erba индуцируется во время адипогенного процесса, а сверхэкспрессия Rev-erba усиливает адипогенез. Исследователи предположили, что роль Rev-erbα в функции адипоцитов может влиять на время таких процессов, как хранение липидов и липолиз, способствуя долгосрочным проблемам с контролем ИМТ. [28] Rev-erbα также регулирует метаболизм желчных кислот путем косвенного подавления Cyp7A1 , который кодирует первый фермент, контролирующий скорость основного пути биосинтеза желчных кислот. [21]
Rev-erbα играет как косвенную, так и прямую роль в метаболизме глюкозы. BMAL1 сильно влияет на выработку глюкозы и синтез гликогена, таким образом, посредством регуляции BMAL1 Rev-erbα косвенно регулирует синтез глюкозы. [33] Более непосредственно, экспрессия Rev-erbα в поджелудочной железе регулирует функцию α-клеток и β-клеток , которые производят глюкагон и инсулин соответственно. [34]
Мышцы и хрящи
[ редактировать ]Rev-erbα играет роль в миогенезе посредством взаимодействия с транскрипционным комплексом Nuclear Factor-T . [29] Он также подавляет экспрессию генов, участвующих в дифференцировке мышечных клеток, и экспрессируется циркадным образом в скелетных мышцах мышей. Потеря функции Rev-erbα снижает содержание и функцию митохондрий, что приводит к нарушению способности к физической нагрузке. Чрезмерная экспрессия приводит к улучшению. [34] [30]
Этот белок также участвует в целостности хряща. Из всех известных ядерных рецепторов Rev-erbα наиболее экспрессируется в хряще, пораженном остеоартритом. [35] Одно исследование показало, что у пациентов с остеоартритом уровни Rev-erbα снижены по сравнению с нормальным хрящом. [36] Исследования ревматоидного артрита (РА) выявили возможность лечения пациентов с РА агонистами Rev-erbα из-за их подавления разрушения костей и хрящей. [37]
Иммунная система
[ редактировать ]Rev-erbα способствует воспалительной реакции у млекопитающих. [34] В гладкомышечных клетках мышей этот белок усиливает экспрессию интерлейкина 6 (IL-6) и циклооксигеназы-2 . У людей он контролирует индуцированный липополисахаридами (ЛПС), эндотоксический ответ, посредством репрессии толл-подобного рецептора (TLR-4) , который запускает иммунный ответ на ЛПС. [28] [34] В головном мозге делеция Rev-erbα вызывает нарушение колебаний активации микроглии и увеличивает экспрессию провоспалительных транскриптов. [19]
Многие иммунные и воспалительные белки демонстрируют циркадное колебательное поведение, и исследования показали, что мыши с дефицитом Rev-erba больше не проявляют этих колебаний, особенно в IL-6 , IL-12 , CCL5 , CXCL1 и CCL2 . [38] Rev-erbα также участвует в развитии врожденных лимфоидных клеток группы 3 (ILC3), которые играют роль в регуляции здоровья кишечника и отвечают за развитие лимфоидных клеток. REV-ERBα способствует экспрессии RORγt , а RORγt необходим для экспрессии ILC3. Rev-erbα высоко экспрессируется в подмножествах ILC3. [39]
Настроение и поведение
[ редактировать ]Rev-erbα участвует в регуляции памяти и настроения. У мышей с нокаутом Rev-erbα наблюдается дефицит краткосрочной, долгосрочной и контекстуальной памяти, что демонстрирует дефицит функции гиппокампа . [40] Кроме того, было высказано предположение, что Rev-erbα играет роль в регуляции выработки дофамина в среднем мозге и связанном с настроением поведении у мышей посредством репрессии транскрипции гена тирозингидроксилазы . [41] Дисфункция, связанная с дофамином, связана с расстройствами настроения, особенно с большим депрессивным расстройством , сезонным аффективным расстройством и биполярным расстройством . человека Генетические вариации локусов NR1D1 также связаны с началом биполярного расстройства. [41]
Rev-erbα был предложен в качестве мишени при лечении биполярного расстройства с помощью лития , который косвенно регулирует белок на посттрансляционном уровне. Литий ингибирует киназу гликогенсинтазы (GSK 3β) , фермент, который фосфорилирует и стабилизирует Rev-erbα. Связывание лития с GSK 3β затем дестабилизирует и изменяет функцию Rev-erbα. [41] Это исследование было вовлечено в разработку терапевтических средств для лечения аффективных расстройств, таких как литий при биполярном расстройстве. [30]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000126368 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000020889 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б с «Ген NR1D1 | Белок NR1D1 | Антитело NR1D1» . Генные карты . Проверено 06 мая 2021 г.
- ^ Перейти обратно: а б с д и ж Инь Л., Ву Н., Лазар М.А. (апрель 2010 г.). «Ядерный рецептор Rev-erbalpha: гемовый рецептор, который координирует циркадный ритм и метаболизм» . Передача сигналов ядерных рецепторов . 8 (1): e001. дои : 10.1621/nrs.08001 . ПМЦ 2858265 . ПМИД 20414452 .
- ^ Ван С., Ли Ф, Линь Ю, Ву Б (2020). «Нацеливание REV-ERBα на терапевтические цели: обещания и проблемы» . Тераностика . 10 (9): 4168–4182. дои : 10.7150/thno.43834 . ПМК 7086371 . ПМИД 32226546 .
- ^ Перейти обратно: а б с д и Беррис Т.П. (июль 2008 г.). «Ядерные гормональные рецепторы гема: REV-ERBalpha и REV-ERBbeta являются лиганд-регулируемыми компонентами часов млекопитающих» . Молекулярная эндокринология . 22 (7): 1509–20. дои : 10.1210/me.2007-0519 . ПМК 5419435 . ПМИД 18218725 .
- ^ Перейти обратно: а б Миядзима Н., Хориучи Р., Сибуя Й., Фукусигэ С., Мацубара К., Тоёшима К. и др. (апрель 1989 г.). «Два гомолога erbA, кодирующие белки с различной способностью связывания Т3, транскрибируются с противоположных цепей ДНК одного и того же генетического локуса». Клетка . 57 (1): 31–9. дои : 10.1016/0092-8674(89)90169-4 . ПМИД 2539258 . S2CID 19135678 .
- ^ Перейти обратно: а б с Лазарь М.А., Ходин Р.А., Дарлинг Д.С., Чин В.В. (март 1989 г.). «Новый член семейства рецепторов тироидных/стероидных гормонов кодируется противоположной цепью транскрипционной единицы крысиного c-erbA альфа» . Молекулярная и клеточная биология . 9 (3): 1128–36. дои : 10.1128/MCB.9.3.1128 . ПМК 362703 . ПМИД 2542765 .
- ^ Бальсалобре А., Дамиола Ф., Шиблер У. (июнь 1998 г.). «Сывороточный шок индуцирует экспрессию циркадных генов в клетках культуры тканей млекопитающих» . Клетка . 93 (6): 929–37. дои : 10.1016/s0092-8674(00)81199-x . ПМИД 9635423 . S2CID 12445337 .
- ^ Перейти обратно: а б Прейтнер Н., Дамиола Ф., Лопес-Молина Л., Закани Дж., Дюбул Д., Альбрехт У. и др. (июль 2002 г.). «Осиротный ядерный рецептор REV-ERBalpha контролирует циркадную транскрипцию в положительном звене циркадного осциллятора млекопитающих» . Клетка . 110 (2): 251–60. дои : 10.1016/s0092-8674(02)00825-5 . ПМИД 12150932 . S2CID 15224136 .
- ^ Перейти обратно: а б Йейтс А.Д., Ачутан П., Аканни В., Аллен Дж., Аллен Дж., Альварес-Харрета Дж. и др. (январь 2020 г.). «Ансамбль 2020» . Исследования нуклеиновых кислот . 48 (Д1): Д682–Д688. дои : 10.1093/nar/gkz966 . ПМК 7145704 . ПМИД 31691826 .
- ^ Ко Ю.С., Мур Д.Д. (апрель 1999 г.). «Связь генов рецепторов ядерных гормонов NR1D2, THRB и RARB: свидетельства древней крупномасштабной дупликации». Геномика . 57 (2): 289–92. дои : 10.1006/geno.1998.5683 . ПМИД 10198169 .
- ^ Кайну Т., Энмарк Э., Густафссон Дж.А., член парламента Пелто-Хуикко (декабрь 1996 г.). «Локализация сиротских рецепторов Rev-ErbA в мозге». Исследования мозга . 743 (1–2): 315–319. дои : 10.1016/S0006-8993(96)00507-0 . ПМИД 9017260 . S2CID 43554925 .
- ^ Бейтман А., Мартин М., Орчард С., Магран М., Агиветова Р., Ахмад С. и др. (январь 2021 г.). «UniProt: универсальная база знаний о белках в 2021 году» . Исследования нуклеиновых кислот . 49 (Д1): Д480–Д489. дои : 10.1093/nar/gkaa1100 . ПМЦ 7778908 . ПМИД 33237286 .
{{cite journal}}: CS1 maint: переопределенная настройка ( ссылка ) - ^ «ГомолоГен – NCBI» . www.ncbi.nlm.nih.gov . Проверено 06 мая 2021 г.
- ^ Перейти обратно: а б с д и ж Эверетт Л.Дж., Лазар М.А. (ноябрь 2014 г.). «Ядерный рецептор Rev-erbα: вверх, вниз и вокруг» . Тенденции в эндокринологии и обмене веществ . 25 (11): 586–92. дои : 10.1016/j.tem.2014.06.011 . ПМЦ 4252361 . ПМИД 25066191 .
- ^ Перейти обратно: а б Гриффин П., Дмитрий Дж.М., Шихан П.В., Лананна Б.В., Го С., Робинетт М.Л. и др. (март 2019 г.). «Белок циркадных часов Rev-erbα регулирует нейровоспаление» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (11): 5102–5107. Бибкод : 2019PNAS..116.5102G . дои : 10.1073/pnas.1812405116 . ПМК 6421453 . ПМИД 30792350 .
- ^ Негоро Х., Окинами Т., Канемацу А., Имамура М., Табата Ю., Огава О. (январь 2013 г.). «Роль доменов Rev-erbα в трансактивации промотора коннексина43 с помощью Sp1». Письма ФЭБС . 587 (1): 98–103. Бибкод : 2013FEBSL.587...98N . дои : 10.1016/j.febslet.2012.11.021 . hdl : 2433/168617 . ПМИД 23201262 . S2CID 30249508 .
- ^ Перейти обратно: а б с д Дуэз Х., Стальс Б. (декабрь 2009 г.). «Рев-эрб-альфа: интегратор циркадных ритмов и метаболизма» . Журнал прикладной физиологии . 107 (6): 1972–80. doi : 10.1152/japplphysicalol.00570.2009 . ПМЦ 2966474 . ПМИД 19696364 .
- ^ Андреани Т.С., Ито Т.К., Йилдирим Э., Хванбо Д.С., Аллада Р. (декабрь 2015 г.). «Генетика циркадных ритмов» . Клиники медицины сна . 10 (4): 413–21. дои : 10.1016/j.jsmc.2015.08.007 . ПМЦ 4758938 . ПМИД 26568119 .
- ^ Гийомон Ф., Дарденте Х., Жигер В., Чермакян Н. (октябрь 2005 г.). «Дифференциальный контроль циркадной транскрипции Bmal1 ядерными рецепторами REV-ERB и ROR» . Журнал биологических ритмов . 20 (5): 391–403. дои : 10.1177/0748730405277232 . ПМИД 16267379 . S2CID 33279857 .
- ^ Солт Л.А., Кожетин DJ, Беррис Т.П. (апрель 2011 г.). «REV-ERB и ROR: молекулярные связи между циркадными ритмами и липидным гомеостазом» . Будущая медицинская химия . 3 (5): 623–38. дои : 10.4155/fmc.11.9 . ПМК 3134326 . ПМИД 21526899 .
- ^ Оиси Ю., Хаяси С., Исагава Т., Осима М., Ивама А., Симба С. и др. (август 2017 г.). «Bmal1 регулирует воспалительные реакции в макрофагах путем модуляции транскрипции энхансерной РНК» . Научные отчеты . 7 (1): 7086. Бибкод : 2017НатСР...7.7086О . дои : 10.1038/s41598-017-07100-3 . ПМК 5539165 . ПМИД 28765524 .
- ^ Гастингс М.Х., Мэйвуд Э.С., Бранкаччо М. (август 2018 г.). «Генерация циркадных ритмов в супрахиазматическом ядре». Обзоры природы. Нейронаука . 19 (8): 453–469. дои : 10.1038/s41583-018-0026-z . ПМИД 29934559 . S2CID 49357675 .
- ^ Дуэз Х., Стальс Б. (январь 2008 г.). «Rev-erb Alpha дает временной сигнал метаболизму». Письма ФЭБС . 582 (1): 19–25. Бибкод : 2008FEBSL.582...19D . дои : 10.1016/j.febslet.2007.08.032 . ПМИД 17765229 . S2CID 84204023 .
- ^ Перейти обратно: а б с Дуэз Х., Стальс Б. (август 2010 г.). «Ядерные рецепторы, связывающие циркадные ритмы и кардиометаболический контроль» . Атеросклероз, тромбоз и сосудистая биология . 30 (8): 1529–34. дои : 10.1161/ATVBAHA.110.209098 . ПМК 3056213 . ПМИД 20631353 .
- ^ Перейти обратно: а б Уэлч Р.Д., Биллон С., Камерик А., Беррис Т.П., Флавени Калифорния (14 мая 2020 г.). «Гетерозиготность Rev-erbα дает дозозависимое фенотипическое преимущество у мышей» . ПЛОС ОДИН . 15 (5): e0227720. Бибкод : 2020PLoSO..1527720W . дои : 10.1371/journal.pone.0227720 . ПМЦ 7224546 . ПМИД 32407314 .
- ^ Перейти обратно: а б с д Марчиано Д.П., Чанг М.Р., Корсо К.А., Госвами Д., Лам В.К., Паскаль Б.Д. и др. (февраль 2014 г.). «Терапевтический потенциал модуляторов ядерных рецепторов для лечения метаболических нарушений: PPARγ, ROR и Rev-erbs» . Клеточный метаболизм . 19 (2): 193–208. дои : 10.1016/j.cmet.2013.12.009 . ПМИД 24440037 .
- ^ Распе Э., Дуэз Х., Мансен А., Фонтен С., Фьеве С., Фрюшар Ж.К. и др. (декабрь 2002 г.). «Идентификация Rev-erbalpha как физиологического репрессора транскрипции гена apoC-III» . Журнал исследований липидов . 43 (12): 2172–9. doi : 10.1194/jlr.M200386-JLR200 . ПМИД 12454280 .
- ^ Руано Э.Г., Канивелл С., Виейра Э (4 августа 2014 г.). «Полиморфизм REV-ERB ALPHA связан с ожирением среди мужского населения Испании, страдающего ожирением» . ПЛОС ОДИН . 9 (8): е104065. Бибкод : 2014PLoSO...9j4065R . дои : 10.1371/journal.pone.0104065 . ПМК 4121274 . ПМИД 25089907 .
- ^ Калсбек А., ла Флер С., Флайерс Е (июль 2014 г.). «Циркадный контроль метаболизма глюкозы» . Молекулярный метаболизм . 3 (4): 372–83. doi : 10.1016/j.molmet.2014.03.002 . ПМК 4060304 . ПМИД 24944897 .
- ^ Перейти обратно: а б с д Лазарь М.А. (2016). «Rev-erbs: круглосуточная интеграция метаболизма». В Сассон-Корси П., Кристен Ю. (ред.). Время обмена веществ и гормонов . Исследования и перспективы эндокринных взаимодействий. Чам: Международное издательство Springer. стр. 63–70. дои : 10.1007/978-3-319-27069-2_7 . ISBN 978-3-319-27068-5 . ПМИД 28892343 .
- ^ Маркс Р. (2018). «Циркадные часы: потенциальная роль в целостности и разрушении хряща» . Международный журнал ортопедии . 5 (4): 936–942. doi : 10.17554/j.issn.2311-5106.2018.05.280 . ISSN 2311-5106 .
- ^ Инь Л., Лазарь М.А. (июнь 2005 г.). «Осиротный ядерный рецептор Rev-erbalpha рекрутирует корепрессор N-CoR/гистондеацетилазы 3 для регуляции циркадного гена Bmal1» . Молекулярная эндокринология . 19 (6): 1452–9. дои : 10.1210/me.2005-0057 . ПМИД 15761026 .
- ^ Фонтен С., Дюбуа Г., Дюге И., Хелледи Т., Ву-Дак Н., Жервуа П. и др. (сентябрь 2003 г.). «Орфанный ядерный рецептор Rev-Erbalpha представляет собой ген-мишень гамма-рецептора, активирующего пролифератор пероксисом (PPAR), и способствует индуцированной PPARgamma дифференцировке адипоцитов» . Журнал биологической химии . 278 (39): 37672–80. дои : 10.1074/jbc.M304664200 . ПМИД 12821652 . S2CID 82056456 .
- ^ Шайерманн С., Кунисаки Ю., Френетт П.С. (март 2013 г.). «Циркадный контроль иммунной системы» . Обзоры природы. Иммунология . 13 (3): 190–8. дои : 10.1038/nri3386 . ПМК 4090048 . ПМИД 23391992 .
- ^ Ван К., Робинетт М.Л., Биллон С., Коллинз П.Л., Бандо Дж.К., Фачи Дж.Л. и др. (октябрь 2019 г.). «Зависимое от циркадного ритма и независимое от циркадного ритма влияние молекулярных часов на врожденные лимфоидные клетки 3 типа» . Наука Иммунология . 4 (40): eaay7501. doi : 10.1126/sciimmunol.aay7501 . ПМК 6911370 . ПМИД 31586012 .
- ^ Джагер Дж., О'Брайен В.Т., Манлав Дж., Кризман Э.Н., Фанг Б., Герхарт-Хайнс З. и др. (апрель 2014 г.). «Поведенческие изменения и дофаминергическая дисрегуляция у мышей, у которых отсутствует ядерный рецептор Rev-erbα» . Молекулярная эндокринология . 28 (4): 490–8. дои : 10.1210/me.2013-1351 . ПМЦ 3968406 . ПМИД 24552589 .
- ^ Перейти обратно: а б с Чунг С., Ли Э.Дж., Юн С., Чхве Х.К., Пак С.Б., Сон Х.Дж. и др. (май 2014 г.). «Влияние циркадного ядерного рецептора REV-ERBα на выработку дофамина в среднем мозге и регуляцию настроения» . Клетка . 157 (4): 858–68. дои : 10.1016/j.cell.2014.03.039 . ПМИД 24813609 . S2CID 3334962 .
Дальнейшее чтение
[ редактировать ]- Лоде В., Бег А., Анри-Дютуа С., Жубель А., Мартин П., Стехелин Д. и др. (март 1991 г.). «Геномная организация гена альфа-рецептора гормона щитовидной железы человека (c-erbA-1)» . Исследования нуклеиновых кислот . 19 (5): 1105–12. дои : 10.1093/нар/19.5.1105 . ПМЦ 333788 . ПМИД 1850510 .
- Миядзима Н., Хориучи Р., Сибуя Й., Фукусигэ С., Мацубара К., Тоёшима К. и др. (апрель 1989 г.). «Два гомолога erbA, кодирующие белки с различной способностью связывания Т3, транскрибируются с противоположных цепей ДНК одного и того же генетического локуса». Клетка . 57 (1): 31–9. дои : 10.1016/0092-8674(89)90169-4 . ПМИД 2539258 . S2CID 19135678 .
- Адельман Г., Бег А., Стелен Д., Лоде В. (апрель 1996 г.). «Функциональный чувствительный элемент Rev-erb альфа, расположенный в промоторе Rev-erb альфа человека, опосредует репрессирующую активность» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (8): 3553–8. Бибкод : 1996PNAS...93.3553A . дои : 10.1073/pnas.93.8.3553 . ПМК 39648 . ПМИД 8622974 .
- Даунс М., Берк Л.Дж., Бэйли П.Дж., Маскат Дж.Е. (ноябрь 1996 г.). «Два домена взаимодействия рецепторов в корепрессоре, N-CoR/RIP13, необходимы для эффективного взаимодействия с Rev-erbA альфа и RVR: физическая ассоциация зависит от области E орфанных рецепторов» . Исследования нуклеиновых кислот . 24 (22): 4379–86. дои : 10.1093/нар/24.22.4379 . ПМК 146280 . ПМИД 8948627 .
- Берк Л.Дж., Даунс М., Лаудет В., Маскат Дж.Э. (февраль 1998 г.). «Идентификация и характеристика новой области взаимодействия корепрессоров в RVR и Rev-erbA альфа» . Молекулярная эндокринология . 12 (2): 248–62. дои : 10.1210/mend.12.2.0061 . ПМИД 9482666 .
- Чжао К., Хорасанизаде С., Миёши Ю., Лазар М.А., Растинеджад Ф. (май 1998 г.). «Структурные элементы комплекса орфанный ядерный рецептор-ДНК» . Молекулярная клетка . 1 (6): 849–61. дои : 10.1016/S1097-2765(00)80084-2 . ПМИД 9660968 .
- Сирк М.Л., Чжао К., Растинежад Ф. (октябрь 2001 г.). «Деформируемость ДНК как признак распознавания в элементе отклика реверберации». Биохимия . 40 (43): 12833–43. дои : 10.1021/bi011086r . ПМИД 11669620 .
- Косте Х., Родригес Х.К. (июль 2002 г.). «Рецептор орфанного ядерного гормона Rev-erbalpha регулирует промотор аполипопротеина CIII человека» . Журнал биологической химии . 277 (30): 27120–9. дои : 10.1074/jbc.M203421200 . hdl : 2445/105711 . ПМИД 12021280 .
- Делерив П., Чин В.В., Суен К.С. (сентябрь 2002 г.). «Идентификация Reverb(alpha) как нового целевого гена ROR(alpha)» . Журнал биологической химии . 277 (38): 35013–8. дои : 10.1074/jbc.M202979200 . ПМИД 12114512 .
- Распе Э., Маутино Г., Дюваль С., Фонтен С., Дуэз Х., Барбье О. и др. (декабрь 2002 г.). «Транкрипционная регуляция экспрессии гена Rev-erbalpha человека с помощью сиротского ядерного рецептора ретиноевой кислоты, связанного с орфанным рецептором альфа» . Журнал биологической химии . 277 (51): 49275–81. дои : 10.1074/jbc.M206215200 . ПМИД 12377782 .
- Распе Э., Дуэз Х., Мансен А., Фонтен С., Фьеве С., Фрюшар Ж.К. и др. (декабрь 2002 г.). «Идентификация Rev-erbalpha как физиологического репрессора транскрипции гена apoC-III» . Журнал исследований липидов . 43 (12): 2172–9. doi : 10.1194/jlr.M200386-JLR200 . ПМИД 12454280 .
- Шопен-Деланной С., Тено С., Делоне Ф., Бюзин Е., Бег А., Дутерк-Кокийо М. и др. (апрель 2003 г.). «Специфический и необычный сигнал ядерной локализации в ДНК-связывающем домене орфанных рецепторов Rev-erb» . Журнал молекулярной эндокринологии . 30 (2): 197–211. дои : 10.1677/jme.0.0300197 . ПМИД 12683943 .
- Фонтен С., Дюбуа Г., Дюге И., Хелледи Т., Ву-Дак Н., Жервуа П. и др. (сентябрь 2003 г.). «Орфанный ядерный рецептор Rev-Erbalpha представляет собой ген-мишень гамма-рецептора, активирующего пролифератор пероксисом (PPAR), и способствует индуцированной PPARgamma дифференцировке адипоцитов» . Журнал биологической химии . 278 (39): 37672–80. дои : 10.1074/jbc.M304664200 . ПМИД 12821652 .
- Джонсон Дж.М., Касл Дж., Гаррет-Энгеле П., Кан З., Лёрч П.М., Armor CD и др. (декабрь 2003 г.). «Полногеномное исследование альтернативного сплайсинга пре-мРНК человека с помощью микрочипов экзонных соединений». Наука . 302 (5653): 2141–4. Бибкод : 2003Sci...302.2141J . CiteSeerX 10.1.1.1017.9438 . дои : 10.1126/science.1090100 . ПМИД 14684825 . S2CID 10007258 .
- Мигита Х., Морсер Дж., Каваи К. (март 2004 г.). «Rev-erbalpha активирует гены, реагирующие на NF-kappaB, в гладкомышечных клетках сосудов» . Письма ФЭБС . 561 (1–3): 69–74. Бибкод : 2004FEBSL.561...69M . дои : 10.1016/S0014-5793(04)00118-8 . ПМИД 15013753 . S2CID 84456190 .
- Ченг Х., Ханна Х., О ЕС, Хикс Д., Миттон К.П., Сваруп А. (август 2004 г.). «Фоторецептор-специфичный ядерный рецептор NR2E3 действует как активатор транскрипции в стержневых фоторецепторах». Молекулярная генетика человека . 13 (15): 1563–75. дои : 10.1093/hmg/ddh173 . ПМИД 15190009 .
- Босолей С.А., Едриховски М., Шварц Д., Элиас Дж.Е., Виллен Дж., Ли Дж. и др. (август 2004 г.). «Крупномасштабная характеристика ядерных фосфопротеинов клеток HeLa» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (33): 12130–5. Бибкод : 2004PNAS..10112130B . дои : 10.1073/pnas.0404720101 . ПМК 514446 . ПМИД 15302935 .
Внешние ссылки
[ редактировать ]- NR1D1+белок+человек Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
галерея PDB |
|---|