RAR-связанный сиротский рецептор альфа
| ПЛЕВАТЬ | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | RORA , NR1F1, ROR1, ROR2, ROR3, RZR-АЛЬФА, RZRA, RAR-связанный орфанный рецептор A, IDDECA | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 600825 ; МГИ : 104661 ; Гомологен : 56594 ; GeneCards : RORA ; ОМА : РОРА - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Связанный с RAR орфанный рецептор альфа ( RORα ), также известный как NR1F1 (подсемейство ядерных рецепторов 1, группа F, член 1), представляет собой ядерный рецептор , который у людей кодируется RORA геном . [5] RORα участвует в регуляции транскрипции некоторых генов, участвующих в циркадных ритмах . [6] У мышей RORα необходим для развития мозжечка. [7] [8] путем прямой регуляции генов, экспрессируемых в клетках Пуркинье. [9] Он также играет важную роль в развитии врожденных лимфоидных клеток 2 типа (ILC2), а у мутантных животных наблюдается дефицит ILC2. [10] [11] Кроме того, клетки ILC3 и Th17 от мышей с дефицитом RORα, хотя и присутствуют в нормальных количествах, дефектны в производстве цитокинов . [12]
Открытие
[ редактировать ]Первые три человеческие изоформы RORα были первоначально клонированы и охарактеризованы как ядерные рецепторы в 1994 году Жигером и его коллегами, когда их структура и функция были впервые изучены. [13]
В начале 2000-х годов различные исследования показали, что RORα демонстрирует ритмические паттерны экспрессии в циркадном цикле в печени , почках , сетчатке и легких . [14] млекопитающих является циркадным Интересно, что примерно в это же время было обнаружено, что содержание RORα в супрахиазматическом ядре . [15] RORα необходим для нормальных циркадных ритмов у мышей . [16] демонстрируя его значение в хронобиологии .
Структура
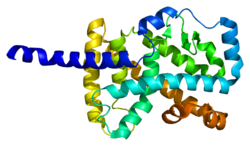
[ редактировать ]Белок, кодируемый этим геном, является членом подсемейства ядерных рецепторов гормонов NR1. [16] У людей были идентифицированы 4 изоформы RORα, которые генерируются посредством альтернативного сплайсинга и использования промотора и демонстрируют дифференциальную тканеспецифическую экспрессию. Белковая содержащего структура RORα состоит из четырех канонических функциональных групп: N-концевого (A/B) домена, ДНК -связывающего домена, два цинковых пальца , шарнирного домена и C-концевого лиганд -связывающего домена. В семействе ROR ДНК-связывающий домен высококонсервативен, а лиганд-связывающий домен консервативен лишь умеренно. [14] Различные изоформы RORα обладают разной специфичностью связывания и силой транскрипционной активности. [5]
Регуляция циркадного ритма
[ редактировать ]Основные млекопитающих циркадные часы представляют собой петлю отрицательной обратной связи , состоящую из Per1/Per2 , Cry1/Cry2 , Bmal1 и Clock . [15] Эта петля обратной связи стабилизируется посредством другой петли, включающей регуляцию транскрипции Bmal1 . [17] Трансактивация Bmal1 REV регулируется через вышестоящий элемент ответа ROR/REV-ERB (RRE) в промоторе Bmal1 , с которым связываются RORα и -ERBα . [17] Эта стабилизирующая регуляторная петля сама по себе индуцируется гетеродимером Bmal1/Clock , который индуцирует транскрипцию RORα и REV-ERBα . [15] RORα, который активирует транскрипцию Bmal1 , и REV-ERBα, который подавляет транскрипцию Bmal1 , конкурируют за связывание с RRE. [17] эта петля обратной связи, регулирующая экспрессию Bmal1 Считается, что , стабилизирует механизм основных часов, помогая защитить его от изменений в окружающей среде . [17]
Механизм
[ редактировать ]Специфическая ассоциация с элементами ROR (RORE) в регуляторных областях необходима для функции RORα как активатора транскрипции. [18] RORα достигает этого путем специфического связывания с консенсусным основным мотивом в RORE, RGGTCA. Это взаимодействие возможно благодаря ассоциации первого цинкового пальца RORα с мотивом ядра в большой бороздке, P-box, и ассоциации его C-концевого продолжения с AT-богатой областью в 5'-области RORE. [16]
Гомология
[ редактировать ]RORα, RORβ и RORγ являются активаторами транскрипции, распознающими элементы ROR-ответа. [19] ROR-альфа экспрессируется в различных типах клеток и участвует в регуляции некоторых аспектов развития, воспалительных реакций и развития лимфоцитов . [20] Изоформы RORα (от RORα1 до RORα3) возникают в результате альтернативного процессинга РНК, при этом RORα2 и RORα3 имеют общую аминоконцевую область, отличную от RORα1. [5] В отличие от RORα, RORβ экспрессируется в тканях центральной нервной системы (ЦНС), участвующих в обработке сенсорной информации и генерации циркадных ритмов , тогда как RORγ имеет решающее значение в лимфатических узлов органогенезе и тимопоэзе . [20]
ДНК-связывающие домены орфанного рецептора DHR3 у дрозофилы демонстрируют особенно тесную гомологию в амино- и карбоксильных областях, прилегающих ко второму региону цинкового пальца в RORα, что позволяет предположить, что эта группа остатков важна для функциональности белков. [5]
PDP1 и VRI у дрозофилы регулируют циркадные ритмы, конкурируя за один и тот же сайт связывания, VP-бокс, аналогично тому, как ROR и REV-ERB конкурентно связываются с RRE. [17] PDP1 и VRI составляют петлю обратной связи и являются функциональными гомологами ROR и REV-ERB у млекопитающих. [17]
Прямые ортологи этого гена были идентифицированы у мышей и человека.
человека Псевдоген цитохрома c HC2 и RORα имеют перекрывающуюся геномную организацию с псевдогеном HC2, расположенным внутри транскрипционной единицы RORα2. Нуклеотидные и выведенные аминокислотные последовательности псевдогена, процессированного цитохромом с, находятся на смысловой цепи, тогда как последовательности аминоконцевого экзона RORα2 находятся на антисмысловой цепи. [5]
Взаимодействия
[ редактировать ]- ДНК: RORα связывается с P-боксом RORE. [16]
- Коактиваторы :
- SRC-1 , CBP , p300 , TRIP-1, TRIP-230 , транскрипционный промежуточный белок-1 ( TIF-1 ), белок, связывающий пролифератор пероксисомы (PBP), и GRIP-1 физически взаимодействуют с RORα. [14]
- Мотив LXXLL: ROR взаимодействует с SRC-1, GRIP-1, CBP и p300 через мотивы LXXLL (L = лейцин, X = любая аминокислота) на этих белках. [14]
- SRC-1 , CBP , p300 , TRIP-1, TRIP-230 , транскрипционный промежуточный белок-1 ( TIF-1 ), белок, связывающий пролифератор пероксисомы (PBP), и GRIP-1 физически взаимодействуют с RORα. [14]
- Убиквитинирование : RORα нацелен на протеасому посредством убиквитинирования. Ко -репрессор Hairless стабилизирует RORα, защищая его от этого процесса, который также репрессирует RORα. [21]
- Сумойлирование : UBE21/UBC9: Фермент I, конъюгирующий убиквитин, взаимодействует с ROR, но его эффект еще не известен. [16]
- Фосфорилирование :
- Фосфорилирование RORα1, которое ингибирует его транскрипционную активность, индуцируется протеинкиназой C. [14]
- ERK2 : внеклеточным сигналом, также фосфорилирует RORα. киназа-2, регулируемая [22]
- ATXN1 : ATXN1 и RORα составляют часть белкового комплекса в клетках Пуркинье . [16]
- FOXP3 : FOXP3 напрямую подавляет транскрипционную активность ROR. [16]
- NME1 : было показано, что ROR специфически взаимодействует с NME1. [23]
- NM23-2 представляет собой нуклеозиддифосфаткиназу, : NM23-2 участвующую в органогенезе и дифференцировке. [6]
- NM23-1 : NM23-1 является продуктом гена-кандидата- супрессора опухолевого метастазирования . [6]
Как мишень для наркотиков
[ редактировать ]Поскольку RORα и REV-ERBα представляют собой ядерные рецепторы, которые имеют одни и те же гены-мишени и участвуют в процессах, регулирующих метаболизм , развитие , иммунитет и циркадный ритм, они обладают потенциалом в качестве мишеней для лекарств . Синтетические лиганды имеют множество потенциальных терапевтических применений и могут использоваться для лечения таких заболеваний, как диабет , атеросклероз , аутоиммунитет и рак . Было обнаружено, что T0901317 и SR1001, два синтетических лиганда, являются RORα и RORγ, обратными агонистами которые подавляют репортерную активность и, как было показано, задерживают начало и клиническую тяжесть рассеянного склероза и других Th17 аутоиммунных заболеваний, опосредованных клетками . SR1078 был обнаружен как агонист RORα и RORγ, который увеличивает экспрессию G6PC и FGF21, что дает терапевтический потенциал для лечения ожирения и диабета, а также рака молочной железы , яичников и простаты . SR3335 также был обнаружен как обратный агонист RORα. [13]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000069667 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000032238 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Jump up to: а б с д и Жигер В., Тини М., Флок Дж., Онг Э., Эванс Р.М., Отулаковски Г. (март 1994 г.). «Специфические для изоформы аминоконцевые домены определяют ДНК-связывающие свойства ROR-альфа, нового семейства ядерных рецепторов орфанных гормонов» . Гены и развитие . 8 (5): 538–53. дои : 10.1101/gad.8.5.538 . ПМИД 7926749 .
- ^ Jump up to: а б с «Ген Энтрез: RORA RAR-связанный сиротский рецептор А» .
- ^ Сидман Р.Л., Лейн П.В., Дики М.М. (август 1962 г.). «Штаггерер, новая мутация у мышей, поражающая мозжечок». Наука . 137 (3530): 610–2. Бибкод : 1962Sci...137..610S . дои : 10.1126/science.137.3530.610 . ПМИД 13912552 . S2CID 30733570 .
- ^ Гамильтон Б.А., Франкель В.Н., Керреброк А.В., Хокинс Т.Л., ФитцХью В., Кусуми К. и др. (февраль 1996 г.). «Нарушение работы ядерного рецептора гормона RORalpha у мышей-шатгереров». Природа . 379 (6567): 736–9. Бибкод : 1996Natur.379..736H . дои : 10.1038/379736a0 . ПМИД 8602221 . S2CID 4318427 .
- ^ Голд Д.А., Бэк С.Х., Шорк Н.Дж., Роуз Д.В., Ларсен Д.Д., Сакс Б.Д. и др. (декабрь 2003 г.). «RORalpha координирует реципрокную передачу сигналов в развитии мозжечка посредством звукового ежа и кальций-зависимых путей» . Нейрон . 40 (6): 1119–31. дои : 10.1016/s0896-6273(03)00769-4 . ПМК 2717708 . ПМИД 14687547 .
- ^ Халим Т.Ю., Макларен А., Романиш М.Т., Голд М.Дж., МакНэгни К.М., Такей Ф. (сентябрь 2012 г.). «Сиротский ядерный рецептор альфа, связанный с рецептором ретиноевой кислоты, необходим для развития естественных хелперных клеток и аллергического воспаления» . Иммунитет . 37 (3): 463–74. doi : 10.1016/j.immuni.2012.06.012 . ПМИД 22981535 .
- ^ Голд М.Дж., Антиньяно Ф., Халим Т.Ю., Хирота Дж.А., Бланше М.Р., Заф С. и др. (апрель 2014 г.). «Врожденные лимфоидные клетки группы 2 способствуют сенсибилизации к локальному, но не системному воздействию TH2-индуцирующих аллергенов». Журнал аллергии и клинической иммунологии . 133 (4): 1142–8. дои : 10.1016/j.jaci.2014.02.033 . ПМИД 24679471 .
- ^ Ло Б.С., Голд М.Дж., Хьюз М.Р., Антиньяно Ф., Вальдес Ю., Заф С. и др. (сентябрь 2016 г.). «Осиротский ядерный рецептор ROR-альфа и врожденные лимфоидные клетки группы 3 вызывают фиброз в мышиной модели болезни Крона» . Наука Иммунология . 1 (3): eaaf8864. doi : 10.1126/sciimmunol.aaf8864 . ПМЦ 5489332 . ПМИД 28670633 .
- ^ Jump up to: а б Кожетин DJ, Беррис Т.П. (март 2014 г.). «Ядерные рецепторы REV-ERB и ROR как мишени для лекарств» . Обзоры природы. Открытие наркотиков . 13 (3): 197–216. дои : 10.1038/nrd4100 . ПМЦ 4865262 . ПМИД 24577401 .
- ^ Jump up to: а б с д и Джеттен А.М., Куребаяши С., Уэда Э. (2001). «Подсемейство ядерных рецепторов-сирот ROR: критические регуляторы множества биологических процессов» . Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии . 69 : 205–47 . дои : 10.1016/S0079-6603(01)69048-2 . ISBN 978-0-12-540069-5 . ПМИД 11550795 .
- ^ Jump up to: а б с Ко Ч., Такахаши Дж.С. (октябрь 2006 г.). «Молекулярные компоненты циркадных часов млекопитающих» . Молекулярная генетика человека . 15 Спецификация № 2 (2): R271-7. дои : 10.1093/hmg/ddl207 . ПМЦ 3762864 . ПМИД 16987893 .
- ^ Jump up to: а б с д и ж Эмери П., Репперт С.М. (август 2004 г.). «Ритмичный Рор» . Нейрон . 43 (4): 443–6. дои : 10.1016/j.neuron.2004.08.009 . ПМИД 15312644 .
- ^ Лайтинен С., Стальс Б. (2003). «Потенциальная роль ROR-альфа в сердечно-сосудистой эндокринологии» . Передача сигналов ядерных рецепторов . 1 : e011. дои : 10.1621/nrs.01011 . ПМК 1402228 . ПМИД 16604183 .
- ^ Чжао X, Чо Х, Ю РТ, Аткинс А.Р., Даунс М., Эванс Р.М. (май 2014 г.). «Ядерные рецепторы работают круглосуточно» . Отчеты ЭМБО . 15 (5): 518–28. дои : 10.1002/эмбр.201338271 . ПМК 4210094 . ПМИД 24737872 .
- ^ Jump up to: а б Ду Дж, Хуан С., Чжоу Б, Зиглер С.Ф. (апрель 2008 г.). «Изоформ-специфическое ингибирование ROR-альфа-опосредованной активации транскрипции человеческим FOXP3» . Журнал иммунологии . 180 (7): 4785–92. дои : 10.4049/jimmunol.180.7.4785 . ПМИД 18354202 .
- ^ Сюн Г., Ван С., Эверс Б.М., Чжоу Б.П., Сюй Р. (апрель 2012 г.). «RORα подавляет инвазию опухоли молочной железы, индуцируя экспрессию SEMA3F» . Исследования рака . 72 (7): 1728–39. дои : 10.1158/0008-5472.CAN-11-2762 . ПМК 3319846 . ПМИД 22350413 .
- ^ Паравичини Г., Штайнмайр М., Андре Э., Беккер-Андре М. (октябрь 1996 г.). «Кандидат в супрессоры метастазирования нуклеотиддифосфаткиназа NM23 специфически взаимодействует с членами подсемейства ядерных орфанных рецепторов ROR/RZR». Связь с биохимическими и биофизическими исследованиями . 227 (1): 82–7. дои : 10.1006/bbrc.1996.1471 . ПМИД 8858107 .
Дальнейшее чтение
[ редактировать ]- Жигер В., Битти Б., Сквайр Дж., Коупленд Н.Г., Дженкинс Н.А. (август 1995 г.). «Сиротский ядерный рецептор ROR альфа (RORA) картируется с консервативной областью гомологии на хромосоме 15q21-q22 человека и хромосоме 9 мыши». Геномика . 28 (3): 596–8. дои : 10.1006/geno.1995.1197 . ПМИД 7490103 .
- Штайнхильбер Д., Брунгс М., Верц О., Визенберг И., Даниэльссон С., Кален Дж.П. и др. (март 1995 г.). «Ядерный рецептор мелатонина подавляет экспрессию гена 5-липоксигеназы в В-лимфоцитах человека» . Журнал биологической химии . 270 (13): 7037–40. дои : 10.1074/jbc.270.13.7037 . ПМИД 7706239 .
- Форман Б.М., Чен Дж., Блумберг Б., Кливер С.А., Хеншоу Р., Онг Э.С. и др. (сентябрь 1994 г.). «Взаимосвязь между ROR альфа 1 и семейством сиротских ядерных рецепторов Rev-erb» . Молекулярная эндокринология . 8 (9): 1253–61. дои : 10.1210/mend.8.9.7838158 . ПМИД 7838158 .
- Беккер-Андре М., Андре Э., ДеЛамартер Дж. Ф. (август 1993 г.). «Идентификация мРНК ядерных рецепторов путем RT-PCR-амплификации консервативных последовательностей мотива цинковых пальцев». Связь с биохимическими и биофизическими исследованиями . 194 (3): 1371–9. дои : 10.1006/bbrc.1993.1976 . ПМИД 7916608 .
- Карлберг С., Хофт ван Хейсдуйнен Р., Стейпл Дж.К., ДеЛамартер Дж.Ф., Беккер-Андре М. (июнь 1994 г.). «RZR, новое семейство связанных с ретиноидами орфанных рецепторов, которые функционируют как мономеры, так и гомодимеры» . Молекулярная эндокринология . 8 (6): 757–70. дои : 10.1210/mend.8.6.7935491 . ПМИД 7935491 . S2CID 22342101 .
- Паравичини Г., Штайнмайр М., Андре Э., Беккер-Андре М. (октябрь 1996 г.). «Кандидат в супрессоры метастазирования нуклеотиддифосфаткиназа NM23 специфически взаимодействует с членами подсемейства ядерных орфанных рецепторов ROR/RZR». Связь с биохимическими и биофизическими исследованиями . 227 (1): 82–7. дои : 10.1006/bbrc.1996.1471 . ПМИД 8858107 .
- Лау П., Бейли П., Дохан Д.Х., Маскат Дж.Е. (январь 1999 г.). «Экзогенная экспрессия доминантно-негативного вектора RORalpha1 в мышечных клетках нарушает дифференцировку: RORalpha1 напрямую взаимодействует с p300 и myoD» . Исследования нуклеиновых кислот . 27 (2): 411–20. дои : 10.1093/нар/27.2.411 . ПМК 148194 . ПМИД 9862959 .
- Аткинс ГБ, Ху X, Гюнтер М.Г., Рачес С., Фридман Л.П., Лазар М.А. (сентябрь 1999 г.). «Коактиваторы сиротского ядерного рецептора RORalpha» . Молекулярная эндокринология . 13 (9): 1550–7. дои : 10.1210/mend.13.9.0343 . ПМИД 10478845 .
- Мейер Т., Кнайссель М., Мариани Дж., Фурнье Б. (август 2000 г.). «In vitro и in vivo доказательства функции сиротского ядерного рецептора ROR-альфа в костном метаболизме» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (16): 9197–202. Бибкод : 2000PNAS...97.9197M . дои : 10.1073/pnas.150246097 . ПМК 16845 . ПМИД 10900268 .
- Гавлас К., Станненберг Х.Г. (декабрь 2000 г.). «Дифференциальное связывание и транскрипционное поведение двух очень родственных рецепторов-сирот, ROR альфа (4) и ROR бета (1)». Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1494 (3): 236–41. дои : 10.1016/s0167-4781(00)00237-2 . ПМИД 11121580 .
- Делерив П., Чин В.В., Суен К.С. (сентябрь 2002 г.). «Идентификация Reverb(alpha) как нового целевого гена ROR(alpha)» . Журнал биологической химии . 277 (38): 35013–8. дои : 10.1074/jbc.M202979200 . ПМИД 12114512 .
- Моретти Р.М., Монтаньани Марелли М., Мотта М., Лимонта П. (2003). «Роль сиротского ядерного рецептора ROR альфа в контроле метастатического поведения андроген-независимых клеток рака простаты». Отчеты онкологии . 9 (5): 1139–43. дои : 10.3892/или.9.5.1139 . ПМИД 12168086 .
- Распе Э., Маутино Г., Дюваль С., Фонтен С., Дуэз Х., Барбье О. и др. (декабрь 2002 г.). «Транкрипционная регуляция экспрессии гена Rev-erbalpha человека с помощью сиротского ядерного рецептора ретиноевой кислоты, связанного с орфанным рецептором альфа» . Журнал биологической химии . 277 (51): 49275–81. дои : 10.1074/jbc.M206215200 . ПМИД 12377782 .
- Каллен Дж., Шлеппи Дж.М., Битш Ф., Делон И., Фурнье Б. (апрель 2004 г.). «Кристаллическая структура связывающего домена ROR-альфа-лиганда человека в комплексе с сульфатом холестерина при 2,2 А» . Журнал биологической химии . 279 (14): 14033–8. дои : 10.1074/jbc.M400302200 . ПМИД 14722075 .
- Мигита Х, Сатозава Н, Лин Дж. Х., Морсер Дж., Каваи К. (январь 2004 г.). «RORalpha1 и RORalpha4 подавляют индуцированную TNF-альфа экспрессию VCAM-1 и ICAM-1 в эндотелиальных клетках человека» . Письма ФЭБС . 557 (1–3): 269–74. дои : 10.1016/S0014-5793(03)01502-3 . ПМИД 14741380 . S2CID 24388280 .
- Мики Н., Икута М., Мацуи Т. (апрель 2004 г.). «Индуцированная гипоксией активация гена орфанного рецептора альфа4, связанного с рецептором ретиноевой кислоты, путем взаимодействия между индуцируемым гипоксией фактором-1 и Sp1» . Журнал биологической химии . 279 (15): 15025–31. дои : 10.1074/jbc.M313186200 . ПМИД 14742449 .
- Мигита Х., Морсер Дж., Каваи К. (март 2004 г.). «Rev-erbalpha активирует гены, реагирующие на NF-kappaB, в гладкомышечных клетках сосудов» . Письма ФЭБС . 561 (1–3): 69–74. дои : 10.1016/S0014-5793(04)00118-8 . ПМИД 15013753 . S2CID 84456190 .
Внешние ссылки
[ редактировать ]- сирота + ядерный + рецептор + ROR-гамма в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
галерея PDB |
|---|

