Циркадный ритм
| Циркадный ритм | |
|---|---|
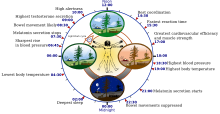 Особенности циркадных биологических часов человека | |
| Произношение | |
| Частота | Повторяется примерно каждые 24 часа |
( Циркадный ритм / s ər ˈ k eɪ d i ə n / ) , или циркадный цикл , представляет собой естественное колебание, которое повторяется примерно каждые 24 часа. Циркадные ритмы могут относиться к любому процессу, который возникает внутри организма (т. е. эндогенный ) и реагирует на окружающую среду ( увлекается окружающей средой). Циркадные ритмы регулируются циркадными часами , основная функция которых заключается в ритмической координации биологических процессов, чтобы они происходили в правильное время, чтобы максимизировать физическую форму человека. Циркадные ритмы широко наблюдались у животных , растений , грибов и цианобактерий, и есть свидетельства того, что они развивались независимо в каждом из этих царств жизни. [1] [2]
Термин «циркадный» происходит от латинского «circa », что означает «вокруг», и «dies », что означает «день». Процессы с 24-часовыми циклами чаще называют суточными ритмами ; суточные ритмы не следует называть циркадными ритмами, пока не будет подтверждено, что они являются эндогенными, а не экологическими. [3]
Хотя циркадные ритмы являются эндогенными, они приспосабливаются к местной среде с помощью внешних сигналов, называемых zeitgebers (от немецкого Zeitgeber (нем. Zeitgeber ). Немецкий: [ˈtsaɪtˌɡeːbɐ] ; горит. ' датель времени ' )), которые включают в себя световой, температурный и окислительно-восстановительный циклы. В клинических условиях аномальный циркадный ритм у человека известен как нарушение циркадного ритма сна . [4]
История [ править ]
Самый ранний зарегистрированный отчет о циркадном процессе принадлежит Теофрасту и датируется 4 веком до нашей эры, вероятно, предоставленный ему отчетом Андросфена , капитана корабля, служившего под командованием Александра Великого . В своей книге «Περὶ φυτῶν ἱστορία», или «Исследование о растениях», Теофраст описывает «дерево со множеством листьев, подобное розе , которое ночью закрывается, но открывается на восходе солнца и к полудню полностью раскрывается; вечером он снова постепенно закрывается и остается закрытым на ночь, и туземцы говорят, что он засыпает». [5] Упомянутое им дерево гораздо позже было идентифицировано как тамариндовое дерево ботаником Х. Брецлем в его книге о ботанических находках александрийских кампаний. [6]
Наблюдение циркадных или суточных процессов у людей упоминается в китайских медицинских текстах, датируемых примерно 13 веком, в том числе в «Руководстве по полудню и полуночи» и « Мнемонической рифме для помощи в выборе акупунктурных точек в соответствии с суточным циклом, дневным циклом». месяца и сезона года . [7]
В 1729 году французский учёный Жан-Жак д’Ортус де Мэран провел первый эксперимент, призванный отличить эндогенные часы от реакций на ежедневные раздражители. Он отметил, что 24-часовые закономерности в движении листьев растения Mimosa pudica сохранялись, даже когда растения находились в постоянной темноте. [8] [9]
В 1896 году Патрик и Гилберт заметили, что во время длительного периода лишения сна сонливость увеличивается и уменьшается с периодом примерно 24 часа. [10] В 1918 г. Дж. С. Шиманский показал, что животные способны поддерживать 24-часовой режим активности при отсутствии внешних сигналов, таких как свет и изменения температуры. [11]
В начале 20 века во время кормления пчел были замечены циркадные ритмы. Огюст Форель , Ингеборг Белинг и Оскар Валь провели многочисленные эксперименты, чтобы определить, обусловлен ли этот ритм эндогенными часами. [12] Существование циркадного ритма было независимо обнаружено у плодовых мух в 1935 году двумя немецкими зоологами, Гансом Калмусом и Эрвином Бюннингом . [13] [14]
В 1954 году важный эксперимент, о котором сообщил Колин Питтендри, продемонстрировал, что эклозия (процесс превращения куколки во взрослую особь) у Drosophila pseudoobscura является циркадным поведением. Он продемонстрировал, что, хотя температура играет жизненно важную роль в ритме эклозий, период эклозий задерживается, но не останавливается при понижении температуры. [15] [14]
Термин «циркадный» был придуман Францем Хальбергом в 1959 году. [16] Согласно первоначальному определению Халберга:
Термин «циркадный» произошел от слов около (около) и dies (день); это может означать, что определенные физиологические периоды длятся около 24 часов, если не совсем так. Здесь термин «циркадный» может быть применен ко всем «24-часовым» ритмам, независимо от того, отличаются ли их периоды, индивидуально или в среднем, от 24-часовых, длиннее или короче, на несколько минут или часов. [17] [18]
В 1977 году Международный комитет по номенклатуре Международного общества хронобиологии официально принял определение:
Циркадный: относящийся к биологическим вариациям или ритмам с частотой 1 цикл в 24 ± 4 часа; около (около, приблизительно) и умирает (сутки или 24 часа). Примечание: термин описывает ритмы с продолжительностью цикла около 24 часов, независимо от того, синхронизированы ли они по частоте (приемлемо), десинхронизированы или независимы от местной временной шкалы окружающей среды, с периодами, слегка, но постоянно отличающимися от 24-часовых. [19]
Рон Конопка и Сеймур Бензер идентифицировали первую часовую мутацию у дрозофилы в 1971 году, назвав ген «геном периода » ( per ), первым обнаруженным генетическим детерминантом поведенческой ритмичности. [20] Пергенен был выделен в 1984 году двумя группами исследователей. Конопка, Джеффри Холл, Майкл Рошбаш и их команда показали, что каждый локус является центром циркадного ритма, и что потеря пера останавливает циркадную активность. [21] [22] В то же время команда Майкла В. Янга сообщила о схожих эффектах per , а также о том, что этот ген покрывает интервал 7,1 тыс. оснований (т.п.н.) на Х-хромосоме и кодирует поли(А) + РНК размером 4,5 т.п.н. [23] [24] Далее они открыли ключевые гены и нейроны циркадной системы дрозофилы , за что Холл, Росбаш и Янг получили Нобелевскую премию по физиологии и медицине 2017 года . [25]
Джозеф Такахаши обнаружил первую мутацию циркадных часов млекопитающих ( clockΔ19 ), используя мышей в 1994 году. [26] [27] Однако недавние исследования показывают, что удаление часов не приводит к поведенческому фенотипу (у животных все еще сохраняются нормальные циркадные ритмы), что ставит под сомнение его важность в генерации ритмов. [28] [29]
Первая мутация человеческих часов была обнаружена в большой семье штата Юта Крисом Джонсом и генетически охарактеризована Ин-Хуэй Фу и Луи Птачеком. Пострадавшие люди представляют собой крайних « утренних жаворонков » с 4-часовым опережением сна и другими ритмами. Эта форма семейного синдрома развитой фазы сна вызвана изменением одной аминокислоты S662➔G в белке PER2 человека. [30] [31]
Критерии [ править ]
Чтобы называться циркадным, биологический ритм должен соответствовать трем общим критериям: [32]
- Ритм имеет эндогенный период свободного хода, который длится примерно 24 часа. Ритм сохраняется в постоянных условиях, т. е. в постоянной темноте, с периодом около 24 часов. Период ритма в постоянных условиях называется периодом свободного хода и обозначается греческой буквой τ (тау). Обоснованием этого критерия является отличие циркадных ритмов от простых реакций на ежедневные внешние сигналы. Ритм нельзя назвать эндогенным , если он не проверен и не сохраняется в условиях отсутствия внешнего периодического воздействия. У дневных животных (активных в дневное время) обычно τ немного превышает 24 часа, тогда как у ночных животных (активных в ночное время) обычно τ короче 24 часов.
- Ритмы легко поддаются дрессировке. Ритм можно восстановить под воздействием внешних раздражителей (таких как свет и тепло). Этот процесс называется увлечением . Внешний стимул, используемый для поддержания ритма, называется zeitgeber , или «дающий время». Путешествие через часовые пояса иллюстрирует способность биологических часов человека приспосабливаться к местному времени; человек обычно испытывает смену часовых поясов до того, как смещение его циркадных часов приведет их в синхронизацию с местным временем.
- Ритмы имеют температурную компенсацию. Другими словами, они поддерживают циркадную периодичность в диапазоне физиологических температур. Многие организмы живут в широком диапазоне температур, и различия в тепловой энергии влияют на кинетику всех молекулярных процессов в их клетках. Чтобы отслеживать время, циркадные часы организма должны поддерживать примерно 24-часовую периодичность, несмотря на меняющуюся кинетику, — свойство, известное как температурная компенсация. Q 10 является Температурный коэффициент мерой этого компенсирующего эффекта. Если коэффициент Q 10 при повышении температуры остается примерно равным 1, ритм считают температурно-компенсированным.
Происхождение [ править ]
В этом разделе отсутствует информация о четырех независимо эволюционировавших [PMID 11533719]. ( сентябрь 2021 г. ) |
Циркадные ритмы позволяют организмам предвидеть и готовиться к точным и регулярным изменениям окружающей среды. Таким образом, они позволяют организмам лучше использовать ресурсы окружающей среды (например, свет и пищу) по сравнению с теми, которые не могут предсказать такое наличие. Поэтому было высказано предположение, что циркадные ритмы дают организмам селективное преимущество с точки зрения эволюции. Однако ритмичность оказывается столь же важной для регулирования и координации внутренних метаболических процессов, как и для координации с окружающей средой . [33] Об этом свидетельствует сохранение (наследственность) циркадных ритмов у плодовых мух после нескольких сотен поколений в постоянных лабораторных условиях. [34] а также у существ, находящихся в постоянной темноте в дикой природе, и путем экспериментального устранения поведенческих, но не физиологических, циркадных ритмов у перепелов . [35] [36]
Что заставило циркадные ритмы развиваться, остается загадочным вопросом. Предыдущие гипотезы подчеркивали, что светочувствительные белки и циркадные ритмы могли возникнуть одновременно в самых ранних клетках с целью защиты реплицирующейся ДНК от высоких уровней повреждающего ультрафиолетового излучения в дневное время. В результате репликация оказалась в тени. Однако доказательств этому недостаточно: на самом деле простейшие организмы с циркадным ритмом, цианобактерии, делают противоположное: они больше делятся в дневное время. [37] Вместо этого недавние исследования подчеркивают важность совместной эволюции окислительно-восстановительных белков с циркадными осцилляторами во всех трех сферах жизни после Великого события окисления, произошедшего примерно 2,3 миллиарда лет назад. [1] [4] В настоящее время считается, что циркадные изменения уровня кислорода в окружающей среде и выработка активных форм кислорода (АФК) в присутствии дневного света, вероятно, привели к необходимости развития циркадных ритмов, чтобы упреждать и, следовательно, противодействовать разрушительным окислительно-восстановительным реакциям в повседневной жизни. основе.
Простейшими из известных циркадных часов являются бактериальные циркадные ритмы , примером которых служат прокариотные цианобактерии . Недавние исследования показали, что циркадные часы Synechococcus elongatus можно восстановить in vitro с помощью всего трех белков ( KaiA , KaiB , KaiC ). [38] своего центрального генератора. Было показано, что эти часы поддерживают 22-часовой ритм в течение нескольких дней после добавления АТФ . Предыдущие объяснения прокариотического циркадного хронометриста зависели от механизма обратной связи транскрипции/трансляции ДНК. [ нужна ссылка ]
Дефект человеческого гомолога » дрозофилы гена « периода был идентифицирован как причина расстройства сна FASPS ( семейный синдром продвинутой фазы сна ), что подчеркивает консервативную природу молекулярных циркадных часов в ходе эволюции. Сейчас известно гораздо больше генетических компонентов биологических часов. Их взаимодействие приводит к возникновению замкнутой петли обратной связи генных продуктов, приводящей к периодическим колебаниям, которые клетки организма интерпретируют как определенное время суток. [39]
Сейчас известно, что молекулярные циркадные часы могут функционировать внутри одной клетки. То есть он клеточно-автономен. [40] Это было показано с помощью Gene Block на изолированных базальных нейронах сетчатки моллюска (BRN). [41] В то же время разные ячейки могут взаимодействовать друг с другом, что приводит к синхронизированному выводу электрических сигналов. Они могут взаимодействовать с эндокринными железами головного мозга, что приводит к периодическому выделению гормонов. Рецепторы этих гормонов могут располагаться далеко по всему телу и синхронизировать периферийные часы различных органов. Таким образом, информация о времени суток, передаваемая глазами, поступает к часам в мозгу, и благодаря этому часы в остальном теле могут быть синхронизированы. Именно так биологические часы скоординированно контролируют время сна/бодрствования, температуру тела, жажду и аппетит. [42] [43]
для животных Значение
Циркадная ритмичность присутствует в режиме сна и питания животных, включая человека. Существуют также четкие закономерности внутренней температуры тела, мозговых волн активности , выработки гормонов , регенерации клеток и других биологических активностей. Кроме того, фотопериодизм , физиологическая реакция организмов на продолжительность дня или ночи, жизненно важен как для растений, так и для животных, а циркадная система играет роль в измерении и интерпретации продолжительности дня. Своевременное предсказание сезонных периодов погодных условий, наличия пищи или активности хищников имеет решающее значение для выживания многих видов. Хотя это и не единственный параметр, изменение продолжительности фотопериода (длины дня) является наиболее предсказуемым экологическим сигналом для определения сезонных сроков физиологии и поведения, особенно для определения времени миграции, спячки и размножения. [44]
Эффект циркадных ритмов нарушения
Мутации или делеции генов часовых у мышей продемонстрировали важность биологических часов для обеспечения правильного выбора времени клеточных/метаболических событий; мыши с часовой мутацией страдают гиперфагией , ожирением и имеют измененный метаболизм глюкозы. [45] У мышей удаление гена альфа-часов Rev-ErbA вызванному диетой может привести к ожирению, , и изменить баланс между использованием глюкозы и липидов, предрасполагая к диабету . [46] Однако неясно, существует ли сильная связь между полиморфизмом часовых генов у людей и предрасположенностью к развитию метаболического синдрома. [47] [48]
Эффект цикла света и темноты [ править ]
Ритм связан с циклом света и темноты. Животные, в том числе люди, находящиеся в полной темноте в течение длительного времени, в конечном итоге начинают функционировать в свободном ритме. Их цикл сна сдвигается назад или вперед каждый «день», в зависимости от того, их «день», их эндогенный короче или длиннее период, 24 часа. Сигналы окружающей среды, которые каждый день переустанавливают ритмы, называются «духами времени». [49] Полностью слепые подземные млекопитающие (например, слепой слепыш Spalax sp.) способны поддерживать свои эндогенные часы при кажущемся отсутствии внешних раздражителей. Хотя у них нет глаз, формирующих изображение, их фоторецепторы (определяющие свет) все еще функционируют; они тоже периодически всплывают на поверхность. [ нужна страница ] [50]
Свободноживущие организмы, которые обычно имеют один или два консолидированных эпизода сна, все равно будут иметь их в среде, защищенной от внешних сигналов, но ритм не подчиняется 24-часовому циклу света и темноты в природе. В этих обстоятельствах ритм сна-бодрствования может оказаться не в фазе с другими циркадными или ультрадианными ритмами, такими как метаболические, гормональные, электрические ритмы ЦНС или ритмы нейротрансмиттеров . [51]
Недавние исследования повлияли на дизайн среды космических кораблей , поскольку было обнаружено, что системы, имитирующие цикл света и темноты, очень полезны для астронавтов. [ ненадежный медицинский источник? ] [52] Светотерапия была опробована в качестве лечения нарушений сна .
Животные Арктики [ править ]
Норвежские исследователи из Университета Тромсё показали, что некоторые арктические животные (например, куропатка , северный олень ) демонстрируют циркадные ритмы только в те части года, где наблюдаются ежедневные восходы и закаты. В одном исследовании северных оленей животные на 70 градусах северной широты демонстрировали циркадные ритмы осенью, зимой и весной, но не летом. У северных оленей на Шпицбергене на 78 градусе северной широты такие ритмы наблюдались только осенью и весной. Исследователи подозревают, что и другие арктические животные могут не проявлять циркадных ритмов при постоянном свете летом и постоянной темноте зимой. [53]
Исследование, проведенное в 2006 году на севере Аляски, показало, что дневные суслики и ночные дикобразы строго поддерживают свои циркадные ритмы в течение 82 солнечных дней и ночей. Исследователи предполагают, что эти два грызуна замечают, что кажущееся расстояние между Солнцем и горизонтом становится самым коротким раз в день, и, таким образом, имеют достаточный сигнал для подхвата (приспособления). [54]
Бабочки и мотыльки [ править ]
Для навигации по осенней миграции восточно-североамериканской бабочки-монарха ( Danaus plexippus ) к местам зимовки в центральной Мексике используется солнечный компас с компенсацией времени, который зависит от циркадных часов в их усиках. [55] [56] Также известно, что циркадный ритм контролирует поведение спаривания у некоторых видов бабочек, таких как Spodopteralittoralis , у которых самки производят специфический феромон , который привлекает и сбрасывает циркадный ритм самцов, чтобы вызвать спаривание в ночное время. [57]
У растений [ править ]

Циркадные ритмы растений сообщают растению, какое сейчас время года и когда цвести, чтобы получить больше шансов привлечь опылителей. Поведение, демонстрирующее ритмы, включает движение листьев ( никтинастия ), рост, прорастание, устьичный/газовый обмен, активность ферментов , фотосинтетическую активность и выделение ароматов, среди прочего. [58] Циркадные ритмы возникают, когда растение синхронизируется со световым циклом окружающей среды. Эти ритмы генерируются эндогенно , являются самоподдерживающимися и относительно постоянны в диапазоне температур окружающей среды. Важные особенности включают две взаимодействующие петли обратной связи транскрипции-трансляции : белки, содержащие домены PAS, которые облегчают белок-белковые взаимодействия; и несколько фоторецепторов, которые точно настраивают часы на различные условия освещенности. Предвидение изменений в окружающей среде позволяет соответствующим образом изменить физиологическое состояние растения, что дает ему адаптивное преимущество. [59] Лучшее понимание циркадных ритмов растений может найти применение в сельском хозяйстве, например, помогая фермерам распределять сбор урожая, чтобы увеличить доступность урожая и защититься от огромных потерь из-за погодных условий.
Свет — это сигнал, с помощью которого растения синхронизируют свои внутренние часы с окружающей средой, и он воспринимается множеством фоторецепторов. Красный и синий свет поглощаются несколькими фитохромами и криптохромами . Фитохром А, phyA, светолабилен и обеспечивает прорастание и деэтиоляцию при недостатке света. [60] Фитохромы B–E более стабильны с phyB – основным фитохромом проростков, выращенных на свету. Ген криптохрома (cry) также является светочувствительным компонентом циркадных часов и, как полагают, участвует как в качестве фоторецептора, так и в качестве части эндогенного механизма кардиостимулятора часов. Криптохромы 1–2 (участвующие в синем спектре UVA) помогают поддерживать продолжительность периода часов в широком диапазоне условий освещенности. [58] [59]

Центральный генератор генерирует самоподдерживающийся ритм и приводится в действие двумя взаимодействующими петлями обратной связи, которые активны в разное время суток. Утренняя петля состоит из CCA1 (Циркадный и Clock-Associated 1) и LHY (Поздний удлиненный гипокотиль), которые кодируют близкородственные транскрипционные факторы MYB , регулирующие циркадные ритмы у Arabidopsis , а также PRR 7 и 9 (регуляторы псевдоответа). Вечерняя петля состоит из GI (Gigantea) и ELF4, которые участвуют в регуляции генов времени цветения. [61] [62] Когда CCA1 и LHY сверхэкспрессируются (при постоянном освещении или темноте), растения становятся аритмичными, а сигналы мРНК уменьшаются, что способствует возникновению петли отрицательной обратной связи . Экспрессия генов CCA1 и LHY колеблется и достигает максимума ранним утром, тогда как экспрессия гена TOC1 колеблется и достигает пика ранним вечером. Хотя ранее предполагалось, что эти три гена моделируют петлю отрицательной обратной связи, в которой сверхэкспрессия CCA1 и LHY репрессирует TOC1, а сверхэкспрессия TOC1 является положительным регулятором CCA1 и LHY, [59] в 2012 году Эндрю Миллар и другие показали, что TOC1 фактически служит репрессором не только CCA1, LHY и PRR7 и 9 в утренней петле, но также GI и ELF4 в вечерней петле. Это открытие и дальнейшее компьютерное моделирование функций и взаимодействий гена TOC1 предполагают переосмысление циркадных часов растений как модель репрессилятора с тройным отрицательным компонентом , а не петлю обратной связи с положительными/отрицательными элементами, характеризующую часы у млекопитающих. [63]
В 2018 году исследователи обнаружили, что экспрессия возникающих транскриптов hnRNA PRR5 и TOC1 следует той же колебательной схеме, что и процессированные транскрипты мРНК ритмично у A. thaliana . LNK связываются с 5'-областью PRR5 и TOC1 и взаимодействуют с RNAP II и другими факторами транскрипции. Более того, взаимодействие RVE8-LNKs позволяет модифицировать пермиссивный паттерн метилирования гистонов (H3K4me3), а сама модификация гистонов параллельна колебаниям экспрессии часового гена. [64]
Ранее было обнаружено, что согласование циркадного ритма растения с циклами света и темноты внешней среды может оказать положительное влияние на растение. [65] Исследователи пришли к такому выводу, проведя эксперименты на трех различных сортах Arabidopsis thaliana . У одной из этих разновидностей был нормальный 24-часовой циркадный цикл. [65] Две другие разновидности были мутированы: одна имела циркадный цикл более 27 часов, а другая имела более короткий, чем обычно, циркадный цикл - 20 часов. [65]
Арабидопсис . с 24-часовым циркадным циклом выращивали в трех разных средах [65] Одна из этих сред имела 20-часовой цикл света и темноты (10 часов света и 10 часов темноты), другая имела 24-часовой цикл света и темноты (12 часов света и 12 часов темноты), а конечная среда имела 28-часовой цикл света и темноты (14 часов света и 14 часов темноты). [65] Два мутировавших растения выращивали как в среде с 20-часовым циклом света и темноты, так и в среде с 28-часовым циклом света и темноты. [65] Было обнаружено, что сорт Arabidopsis с 24-часовым циклом циркадного ритма лучше всего рос в среде, которая также имела 24-часовой цикл света и темноты. [65] В целом было обнаружено, что все разновидности Arabidopsis thaliana имели более высокий уровень хлорофилла и ускоряли рост в средах, где циклы света и темноты соответствовали их циркадному ритму. [65]
Исследователи предположили, что причиной этого может быть то, что соответствие Arabidopsis циркадного ритма его окружающей среде может позволить растению лучше подготовиться к рассвету и закату и, таким образом, лучше синхронизировать свои процессы. [65] В этом исследовании также было обнаружено, что гены, которые помогают контролировать хлорофилл, достигают пика через несколько часов после рассвета. [65] Это, по-видимому, согласуется с предполагаемым феноменом, известным как метаболический рассвет. [66]
Согласно гипотезе метаболического рассвета, сахара, образующиеся в результате фотосинтеза, потенциально могут помочь регулировать циркадный ритм и определенные пути фотосинтеза и метаболизма. [66] [67] С восходом солнца становится доступно больше света, что обычно способствует усилению фотосинтеза. [66] Сахара, образующиеся в результате фотосинтеза, подавляют PRR7. [68] Эта репрессия PRR7 затем приводит к увеличению экспрессии CCA1. [68] С другой стороны, снижение уровня фотосинтетического сахара увеличивает экспрессию PRR7 и снижает экспрессию CCA1. [66] Предполагается, что эта петля обратной связи между CCA1 и PRR7 вызывает метаболический рассвет. [66] [69]
У дрозофилы [ править ]
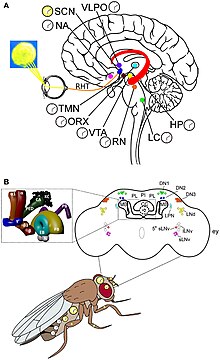
Молекулярный механизм циркадного ритма и восприятия света лучше всего понятен у дрозофилы . Гены часов обнаружены у дрозофилы и действуют вместе с часовыми нейронами. Существует два уникальных ритма: один во время процесса вылупления (так называемого вылупления ) из куколки, а другой во время спаривания. [70] Часовые нейроны расположены в отдельных кластерах в центральном мозге. Наиболее изученными часовыми нейронами являются большие и малые латеральные вентральные нейроны (l-LNvs и s-LNvs) зрительной доли . Эти нейроны вырабатывают фактор диспергирования пигмента (PDF), нейропептид, который действует как циркадный нейромодулятор между различными часовыми нейронами. [71]

Циркадный ритм дрозофилы осуществляется через петлю обратной связи транскрипции-трансляции. Основной механизм синхронизации состоит из двух взаимозависимых контуров обратной связи, а именно контура PER/TIM и контура CLK/CYC. [72] Петля CLK/CYC возникает в течение дня и инициирует транскрипцию генов per и tim . Но уровень их белков остается низким до наступления сумерек, поскольку при дневном свете также активируется ген двойного времени ( dbt ). Белок DBT вызывает фосфорилирование и оборот мономерных белков PER. [73] [74] ТИМ также фосфорилируется лохматым до захода солнца. После захода солнца ДБТ исчезает, и молекулы ПЕР стабильно связываются с ТИМ. Димер PER/TIM проникает в ядро несколько раз в ночное время и связывается с димерами CLK/CYC. Связанный PER полностью останавливает транскрипционную активность CLK и CYC. [75]
Ранним утром свет активирует ген крика , и его белок CRY вызывает разрушение ТИМ. Таким образом, димер PER/TIM диссоциирует, и несвязанный PER становится нестабильным. PER подвергается прогрессивному фосфорилированию и, в конечном итоге, деградации. Отсутствие PER и TIM позволяет активировать гены clk и cyc . Таким образом, часы сбрасываются, чтобы начать следующий циркадный цикл. [76]
Модель PER-TIM [ править ]
Эта белковая модель была разработана на основе колебаний белков PER и TIM у дрозофилы . [77] Он основан на своей предшественнице, модели PER, в которой объяснялось, как ген PER и его белок влияют на биологические часы. [78] Модель включает образование ядерного комплекса PER-TIM, который влияет на транскрипцию генов PER и TIM (обеспечивая отрицательную обратную связь) и множественное фосфорилирование этих двух белков. Циркадные колебания этих двух белков, по-видимому, синхронизируются с циклом света и темноты, даже если они не обязательно от него зависят. [79] [77] Белки PER и TIM фосфорилируются, и после того, как они образуют ядерный комплекс PER-TIM, они возвращаются внутрь ядра, чтобы остановить экспрессию мРНК PER и TIM. Это ингибирование длится до тех пор, пока белок или мРНК не разрушаются. [77] Когда это происходит, комплекс снимает торможение. Здесь также можно упомянуть, что деградация белка ТИМ ускоряется под действием света. [79]
У млекопитающих [ править ]

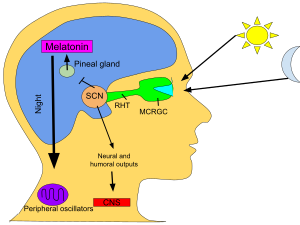
Первичные циркадные часы у млекопитающих расположены в супрахиазматическом ядре (или ядрах) (SCN), паре отдельных групп клеток , расположенных в гипоталамусе . Разрушение СХЯ приводит к полному отсутствию регулярного ритма сна-бодрствования. СХЯ получает информацию об освещенности через глаза. Сетчатка глаза содержит «классические» фоторецепторы (« палочки » и « колбочки »), которые используются для обычного зрения. Но сетчатка также содержит специализированные ганглиозные клетки , которые непосредственно светочувствительны и проецируются непосредственно на СХЯ, где они помогают вовлечь (синхронизировать) эти главные циркадные часы. Белки, участвующие в работе часов SCN, гомологичны белкам, обнаруженным у плодовой мухи. [80]
Эти клетки содержат фотопигмент меланопсин , и их сигналы следуют по пути, называемому ретиногипоталамическим трактом , ведущему к СХЯ. Если клетки из SCN удалить и культивировать, они сохранят свой собственный ритм в отсутствие внешних сигналов. [81]
СХЯ получает информацию о продолжительности дня и ночи от сетчатки, интерпретирует ее и передает в шишковидную железу — крошечную структуру, имеющую форму сосновой шишки и расположенную на эпиталамусе . В ответ шишковидная железа выделяет гормон мелатонин . [82] Пик секреции мелатонина ночью и спад в течение дня, а его присутствие дает информацию о продолжительности ночи.
Несколько исследований показали, что мелатонин шишковидной железы влияет на ритмичность SCN, модулируя циркадные паттерны активности и другие процессы. Однако природа и значение этой обратной связи на системном уровне неизвестны. [83]
Циркадные ритмы человека можно настроить на несколько более короткие и длинные периоды, чем земные 24 часа. Исследователи из Гарварда показали, что людей можно, по крайней мере, приучить к 23,5-часовому циклу и 24,65-часовому циклу. [84]
Люди [ править ]
Ранние исследования циркадных ритмов показали, что большинство людей предпочитают день около 25 часов, когда они изолированы от внешних раздражителей, таких как дневной свет и хронометраж. Однако это исследование было ошибочным, поскольку не смогло защитить участников от искусственного света. Хотя испытуемые были защищены от сигналов времени (например, от часов) и дневного света, исследователи не знали о эффекте задержки фазы электрического освещения в помещении. [85] [ сомнительно – обсудить ] Испытуемым разрешалось включать свет, когда они бодрствовали, и выключать его, когда они хотели спать. Электрический свет вечером задержал их циркадную фазу. [86] Более строгое исследование, проведенное в 1999 году Гарвардским университетом, показало, что естественный человеческий ритм приближается к 24 часам и 11 минутам: намного ближе к солнечным суткам . [87] С этим исследованием согласуется более позднее исследование 2010 года, в котором также были выявлены половые различия: циркадный период у женщин немного короче (24,09 часа), чем у мужчин (24,19 часа). [88] В этом исследовании женщины, как правило, просыпались раньше, чем мужчины, и проявляли большее предпочтение утренним занятиям, чем мужчины, хотя биологические механизмы, лежащие в основе этих различий, неизвестны. [88]
и Биологические эффекты маркеры
Классическими фазовыми маркерами для измерения времени циркадного ритма млекопитающих являются:
- мелатонина секреция шишковидной железой , [89]
- температура тела , минимальная [89] и
- уровень кортизола в плазме . [90]
Для температурных исследований испытуемые должны оставаться бодрствующими, но спокойными и полулежащими в почти полной темноте, при этом им постоянно измеряют ректальную температуру. велики Хотя различия между нормальными хронотипами , у среднего взрослого человека температура достигает минимума примерно в 5 часов утра, примерно за два часа до обычного времени пробуждения. Бэр и др. [91] обнаружили, что у молодых людей дневной минимум температуры тела приходится примерно на 04:00 (4 часа утра) для утренних типов, но примерно в 06:00 (6 часов утра) для вечерних типов. Этот минимум наблюдался примерно в середине восьмичасового периода сна у утренних типов, но ближе к пробуждению у вечерних типов.
Мелатонин отсутствует в организме или его уровень неопределяемо низок в дневное время. Его начало при тусклом свете, начало мелатонина при тусклом свете (DLMO), примерно в 21:00 (21:00), можно измерить в крови или слюне. Его основной метаболит также можно определить в утренней моче. И DLMO, и средняя точка (по времени) присутствия гормона в крови или слюне использовались в качестве циркадных маркеров. мелатонина Однако новые исследования показывают, что смещение может быть более надежным маркером. Бенлусиф и др. [89] обнаружили, что маркеры фазы мелатонина были более стабильными и более сильно коррелировали со временем сна, чем минимум внутренней температуры. Они обнаружили, что как смещение сна, так и смещение мелатонина более сильно коррелируют с маркерами фазы, чем начало сна. Кроме того, фаза снижения уровня мелатонина более надежна и стабильна, чем прекращение синтеза мелатонина.
Другие физиологические изменения, которые происходят в соответствии с циркадным ритмом, включают частоту сердечных сокращений и многие клеточные процессы, включая окислительный стресс , клеточный метаболизм , иммунные и воспалительные реакции. [92] эпигенетическая модификация, на гипоксию / гипероксию пути реакции , эндоплазматический ретикулярный стресс , аутофагия и регуляция среды стволовых клеток ». [93] В исследовании молодых мужчин было обнаружено, что частота сердечных сокращений достигает минимального среднего значения во время сна, а самого высокого среднего значения вскоре после пробуждения. [94]
В отличие от предыдущих исследований было обнаружено, что температура тела не влияет на результаты психологических тестов. Вероятно, это связано с эволюционным давлением более высоких когнитивных функций по сравнению с другими областями функций, изученными в предыдущих исследованиях. [95]
За пределами «главных часов» [ править ]
Более или менее независимые циркадные ритмы обнаруживаются во многих органах и клетках организма за пределами супрахиазматических ядер (SCN), «главных часов». Действительно, нейробиолог Джозеф Такахаши и его коллеги заявили в статье 2013 года, что «почти каждая клетка в организме содержит циркадные часы». [96] Например, эти часы, называемые периферическими генераторами, были обнаружены в надпочечниках, пищеводе , легких , печени , поджелудочной железе , селезенке , тимусе и коже. [97] [98] [99] Есть также некоторые доказательства того, что обонятельная луковица [100] и простата [101] могут испытывать колебания, по крайней мере, при культивировании.
Хотя осцилляторы в коже реагируют на свет, системное влияние не доказано. [102] Кроме того, было показано, что многие осцилляторы, такие как клетки печени , реагируют не только на свет, но и на питание. [103]
биологические часы и Свет
Свет сбрасывает биологические часы в соответствии с кривой фазового отклика (ФХЧ). В зависимости от времени свет может ускорять или замедлять циркадный ритм. И PRC, и необходимая освещенность грызунов требуются более низкие уровни освещенности, варьируются от вида к виду, и для сброса часов у ночных чем у людей. [104]
Принудительные более длинные или короткие циклы [ править ]
В различных исследованиях на людях использовались принудительные циклы сна/бодрствования, сильно отличающиеся от 24-часовых, например, исследования, проведенные Натаниэлем Клейтманом в 1938 году (28 часов) и Дерком-Яном Дейком и Чарльзом Цейслером в 1990-х годах (20 часов). Поскольку люди с нормальными (типичными) циркадными ритмами не могут привыкнуть к таким аномальным дневным/ночным ритмам, [105] это называется протоколом принудительной десинхронизации. Согласно такому протоколу эпизоды сна и бодрствования не связаны с эндогенным циркадным периодом организма, что позволяет исследователям оценить влияние циркадной фазы (т. е. относительное время циркадного цикла) на аспекты сна и бодрствования, включая латентность сна и другие аспекты. функции - как физиологические, поведенческие, так и когнитивные. [106] [107] [108] [109] [110]
Исследования также показывают, что Cyclosa turbinata уникальна тем, что из-за ее двигательной активности и построения паутины циркадные часы имеют исключительно короткий период, около 19 часов. Когда пауков C. turbinata поместили в камеры с периодами 19, 24 или 29 часов равномерного распределения света и темноты, ни один из пауков не обнаружил снижения продолжительности жизни по своим циркадным часам. Эти данные позволяют предположить, что C. turbinata не несет таких же последствий крайней десинхронизации, как другие виды животных.

Здоровье человека [ править ]

Фонд циркадной медицины
Передовым направлением исследований в области циркадной биологии является перевод основных механизмов биологических часов в клинические инструменты, и это особенно актуально для лечения сердечно-сосудистых заболеваний. [111] [112] [113] [114] Выбор времени лечения в соответствии с биологическими часами, хронотерапия , также может принести пользу пациентам с гипертонией (высоким кровяным давлением) за счет значительного повышения эффективности и снижения токсичности лекарств или побочных реакций. [115] 3) «Циркадная фармакология» или препараты, воздействующие на механизм циркадных часов, как было экспериментально показано на моделях грызунов, значительно уменьшают ущерб от сердечных приступов и предотвращают сердечную недостаточность. [116] Важно отметить, что для рационального внедрения наиболее многообещающих методов циркадной медицины в клиническую практику крайне важно понимать, как они помогают лечить заболевания у обоих биологических полов. [117] [118] [119] [120]
Причины нарушения циркадных ритмов [ править ]
Внутреннее освещение [ править ]
Требования к освещению для циркадной регуляции не просто такие же, как для зрения; это начинает учитываться при планировании внутреннего освещения в офисах и учреждениях. [121] В исследованиях на животных по воздействию света в лабораторных условиях до недавнего времени учитывалась интенсивность света ( облученность ), а не цвет, который, как можно показать, «действует как важный регулятор биологического времени в более естественных условиях». [122]
Синее светодиодное освещение подавляет выработку мелатонина в пять раз больше, чем оранжево-желтое натриевое освещение высокого давления (HPS) ; Металлогалогенная лампа белого света подавляет мелатонин со скоростью более чем в три раза большей, чем HPS. [123] Симптомы депрессии, вызванные длительным воздействием света в ночное время, можно устранить, вернувшись к нормальному циклу. [124]
Пилоты авиакомпаний и бортпроводники [ править ]
Из-за характера работы пилотов авиакомпаний, которые часто пересекают несколько часовых поясов и областей солнечного света и темноты за один день и проводят много часов бодрствуя как днем, так и ночью, они часто не могут поддерживать режим сна, соответствующий естественному циркадному ритму человека. ритм; эта ситуация может легко привести к усталости . NTSB считает , что это способствовало множеству несчастных случаев. [125] и провел несколько научных исследований с целью найти методы борьбы с утомляемостью пилотов. [126]
Эффект наркотиков [ править ]
Исследования, проведенные как на животных, так и на людях, показывают основные двунаправленные связи между циркадной системой и злоупотреблением наркотиками. Указывается, что эти злоупотребляющие препараты влияют на центральный циркадный водитель ритма. У людей с расстройствами, вызванными употреблением психоактивных веществ, наблюдаются нарушения ритма. Эти нарушенные ритмы могут увеличить риск злоупотребления психоактивными веществами и рецидива. Вполне возможно, что генетические и/или экологические нарушения нормального цикла сна и бодрствования могут повысить восприимчивость к зависимости. [127]
Трудно определить, является ли нарушение циркадного ритма причиной увеличения распространенности злоупотребления психоактивными веществами или же виноваты другие факторы окружающей среды, такие как стресс.Изменения циркадного ритма и сна происходят, когда человек начинает злоупотреблять наркотиками и алкоголем. Как только человек прекращает употреблять наркотики и алкоголь, циркадный ритм продолжает нарушаться. [127]
Стабилизация сна и циркадного ритма, возможно, поможет снизить уязвимость к зависимости и снизить вероятность рецидива. [127]
Циркадные ритмы и часовые гены, экспрессируемые в областях мозга за пределами супрахиазматического ядра, могут существенно влиять на эффекты, вызываемые такими наркотиками, как кокаин . [ нужна ссылка ] Более того, генетические манипуляции с часовыми генами глубоко влияют на действие кокаина. [128]
циркадных нарушения Последствия ритмов
Разрушение [ править ]
Нарушение ритма обычно имеет негативные последствия. Многие путешественники сталкивались с состоянием, известным как смена часовых поясов , с сопутствующими ему симптомами усталости , дезориентации и бессонницы . [129]
Ряд других расстройств, таких как биполярное расстройство и некоторые расстройства сна , такие как расстройство задержки фазы сна (DSPD), связаны с нерегулярным или патологическим функционированием циркадных ритмов. [130] [131]
Считается, что нарушение ритмов в долгосрочной перспективе имеет серьезные неблагоприятные последствия для здоровья периферических органов за пределами головного мозга, в частности, в развитии или обострении сердечно-сосудистых заболеваний. [132] [133]
Исследования показали, что поддержание нормального сна и циркадных ритмов важно для многих аспектов работы мозга и здоровья. [132] Ряд исследований также показал, что короткий период сна в течение дня может снизить стресс и повысить продуктивность без какого-либо заметного влияния на нормальные циркадные ритмы. [134] [135] [136] Циркадные ритмы также играют роль в ретикулярной активирующей системе , которая имеет решающее значение для поддержания состояния сознания. Разворот [ нужны разъяснения ] в цикле сон-бодрствование может быть признаком или осложнением уремии , [137] азотемия или острое повреждение почек . [138] [139] Исследования также помогли выяснить, как свет оказывает прямое влияние на здоровье человека через его влияние на циркадную биологию. [140]
Связь с сердечно заболеваниями - сосудистыми
Одно из первых исследований, призванных определить, как нарушение циркадных ритмов вызывает сердечно-сосудистые заболевания, было проведено на хомяках Тау, у которых имеется генетический дефект в механизме циркадных часов. [141] При поддержании 24-часового цикла света и темноты, который был «не синхронизирован» с их нормальным циркадным механизмом, у них развились серьезные сердечно-сосудистые и почечные заболевания; однако, когда животные Тау в течение всей своей жизни выращивались при 22-часовом ежедневном цикле света и темноты, у них была здоровая сердечно-сосудистая система. [141] Неблагоприятное воздействие нарушения циркадных ритмов на физиологию человека изучалось в лаборатории с использованием протокола смещения. [142] [143] и изучая сменных рабочих. [111] [144] [145] Циркадное смещение связано со многими факторами риска сердечно-сосудистых заболеваний. Сообщалось о высоких уровнях биомаркера атеросклероза, резистина, у посменных рабочих, что указывает на связь между циркадным нарушением и накоплением бляшек в артериях. [145] Кроме того, наблюдались повышенные уровни триацилглицеридов (молекул, используемых для хранения избыточных жирных кислот), которые способствуют затвердеванию артерий, что связано с сердечно-сосудистыми заболеваниями, включая сердечный приступ, инсульт и болезни сердца. [145] [146] Сменная работа и возникающее в результате этого смещение циркадных ритмов также связано с гипертонией. [147]
Ожирение и диабет [ править ]
Ожирение и диабет связаны с образом жизни и генетическими факторами. Среди этих факторов в развитии метаболических нарушений могут играть роль нарушение циркадного механизма и/или несогласованность системы циркадного времени с внешней средой (например, циклом света и темноты). [132]
Сменная работа или хроническая смена часовых поясов имеют серьезные последствия для циркадных и метаболических процессов в организме. Животные, которых заставляют есть во время периода отдыха, демонстрируют увеличение массы тела и измененную экспрессию часовых и метаболических генов. [148] [146] У людей сменная работа, которая способствует нерегулярному питанию, связана с изменением чувствительности к инсулину, диабетом и увеличением массы тела. [147] [146] [149]
Рак [ править ]
Смещение циркадных ритмов также связано с повышенным риском развития рака. У мышей было обнаружено, что нарушение основных часовых генов, генов периода (Per2, Per1), вызванное смещением циркадных ритмов, ускоряет рост раковых клеток у мышей. Однако связь между этими генами и раком зависит от типа путей и задействованных генов. [150] [151] Существуют значительные доказательства того, что сменная работа и, следовательно, циркадное смещение коррелируют с раком молочной железы и простаты у людей. [152] [153] [154] [155] [151]
эффекты Когнитивные
Снижение когнитивной функции связано с нарушением циркадного ритма. У хронически сменных работников наблюдается повышенный уровень операционных ошибок, нарушение зрительно-моторных функций и эффективности обработки информации, что может привести как к снижению производительности, так и к потенциальным проблемам с безопасностью. [156] Повышенный риск деменции связан с хроническими работниками ночной смены по сравнению с работниками дневной смены, особенно для людей старше 50 лет. [157] [158] [159]
и культура Общество
В 2017 году Джеффри К. Холл , Майкл В. Янг и Майкл Росбаш были удостоены Нобелевской премии по физиологии и медицине «за открытия молекулярных механизмов, контролирующих циркадный ритм». [160] [161]
Циркадные ритмы были взяты в качестве примера переноса научных знаний в общественную сферу. [162]
См. также [ править ]
- Актиграфия (также известная как актиметрия)
- АРНТЛ
- АРНТЛ2
- Бактериальные циркадные ритмы
- Нарушения циркадного ритма сна , такие как
- Хронобиология
- Хроноразрушение
- ЧАСЫ
- Цирказемидный ритм
- Цирказептан , 7-дневный биологический цикл
- Криптохром
- CRY1 и CRY2 : гены семейства криптохромов.
- Суточный цикл
- Влияние света на циркадный ритм
- Свет в школьных зданиях
- PER1 , PER2 и PER3 : гены семейства периодов.
- Фоточувствительная ганглиозная клетка : часть глаза, которая участвует в регуляции циркадного ритма.
- Полифазный сон
- Рев-Эрба альфа
- Сегментированный сон
- Архитектура сна (сон у человека)
- Сон у животных, не являющихся людьми
- Стефания Фоллини
- Ультрадианный ритм
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б Эдгар Р.С., Грин Э.В., Чжао Ю., ван Оойен Г., Олмедо М., Цинь Х. и др. (май 2012 г.). «Пероксиредоксины являются консервативными маркерами циркадных ритмов» . Природа . 485 (7399): 459–464. Бибкод : 2012Natur.485..459E . дои : 10.1038/nature11088 . ПМЦ 3398137 . ПМИД 22622569 .
- ^ Янг М.В., Кей С.А. (сентябрь 2001 г.). «Часовые пояса: сравнительная генетика циркадных часов». Обзоры природы. Генетика . 2 (9): 702–715. дои : 10.1038/35088576 . ПМИД 11533719 . S2CID 13286388 .
- ^ Витатерна М.Х., Такахаши Дж.С., Турек Ф.В. (2001). «Обзор циркадных ритмов» . Исследования алкоголя и здоровье . 25 (2): 85–93. ПМК 6707128 . ПМИД 11584554 .
- ↑ Перейти обратно: Перейти обратно: а б Басс Дж. (ноябрь 2012 г.). «Циркадная топология метаболизма». Природа . 491 (7424): 348–356. Бибкод : 2012Natur.491..348B . дои : 10.1038/nature11704 . ПМИД 23151577 . S2CID 27778254 .
- ^ Теофраст, «Περὶ φυτῶν ἱστορία». «Исследование растений и небольшие работы по запахам и погодным признакам, с английским переводом сэра Артура Хорта, барт, 1916 год» . Архивировано из оригинала 13 апреля 2022 г.
- ^ Брецль Х (1903). Ботаническое исследование Александровского крестного хода . Лейпциг: Тойбнер. [ нужна страница ]
- ^ Лу Г.Д. (25 октября 2002 г.). Небесные Ланцеты . Психология Пресс. стр. 137–140. ISBN 978-0-7007-1458-2 .
- ^ Майран Дж. Дж. (1729 г.). «Ботаническое наблюдение». История Королевской академии наук : 35–36.
- ^ Гарднер М.Дж., Хаббард К.Е., Хотта К.Т., Додд А.Н., Уэбб А.А. (июль 2006 г.). «Как растения определяют время» . Биохимический журнал . 397 (1): 15–24. дои : 10.1042/BJ20060484 . ПМК 1479754 . ПМИД 16761955 .
- ^ Дейк DJ, фон Шанц М (август 2005 г.). «Время и консолидация человеческого сна, бодрствования и производительности с помощью симфонии осцилляторов» . Журнал биологических ритмов . 20 (4): 279–290. дои : 10.1177/0748730405278292 . ПМИД 16077148 . S2CID 13538323 .
- ^ Данчин А. «Важные даты 1900–1919 годов» . Исследовательский центр ХКУ-Пастер . Архивировано из оригинала 20 октября 2003 г. Проверено 12 января 2008 г.
- ^ Antle MC, Silver R (ноябрь 2009 г.). «Нейронная основа выбора времени и упреждающего поведения» . Европейский журнал неврологии . 30 (9): 1643–1649. дои : 10.1111/j.1460-9568.2009.06959.x . ПМЦ 2929840 . ПМИД 19878281 .
- ^ Брюс В.Г., Питтендри CS (1957). «Эндогенные ритмы у насекомых и микроорганизмов». Американский натуралист . 91 (858): 179–195. дои : 10.1086/281977 . S2CID 83886607 .
- ↑ Перейти обратно: Перейти обратно: а б Питтендри CS (1993). «Временная организация: размышления дарвинистского часовщика». Ежегодный обзор физиологии . 55 (1): 16–54. дои : 10.1146/annurev.ph.55.030193.000313 . ПМИД 8466172 . S2CID 45054898 .
- ^ Питтендри CS (октябрь 1954 г.). «О независимости от температуры в часовой системе, контролирующей время вылета у дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 40 (10): 1018–1029. Бибкод : 1954PNAS...40.1018P . дои : 10.1073/pnas.40.10.1018 . ПМК 534216 . ПМИД 16589583 .
- ^ Хальберг Ф., Корнелиссен Г., Катинас Г., Сюткина Е.В., Сотерн Р.Б., Заславская Р. и др. (октябрь 2003 г.). «Трансдисциплинарное объединяющее значение циркадных открытий 1950-х годов» . Журнал циркадных ритмов . 1 (1): 2. дои : 10.1186/1740-3391-1-2 . ПМК 317388 . ПМИД 14728726 .
В конце концов, по той же причине, я вернулся к «циркадному режиму»…
- ^ Хальберг Ф (1959). «[Физиологическая 24-часовая периодичность; общие и процедурные соображения в отношении цикла надпочечников]». Международный журнал исследований витаминов. Добавка . 10 :225-296. ПМИД 14398945 .
- ^ Куккари В.Л., Южный РБ (2006 г.). Знакомство с биологическими ритмами: учебник временной организации жизни с последствиями для здоровья, общества, воспроизводства и окружающей среды . Нью-Йорк: Спрингер. п. 23. ISBN 978-1-4020-3691-0 .
- ^ Хальберг Ф., Каранденте Ф., Корнелиссен Г., Катинас Г.С. (1977). «[Глоссарий хронобиологии (перевод автора)]». Хронобиология . 4 (Приложение 1): 1–189. ПМИД 352650 .
- ^ Конопка Р.Дж., Бензер С. (сентябрь 1971 г.). «Часовые мутанты Drosophila melanogaster» . Труды Национальной академии наук Соединенных Штатов Америки . 68 (9): 2112–2116. Бибкод : 1971PNAS...68.2112K . дои : 10.1073/pnas.68.9.2112 . ПМЦ 389363 . ПМИД 5002428 .
- ^ Редди П., Церинг В.А., Уилер Д.А., Пирротта В., Хэдфилд С., Холл Дж.К. и др. (октябрь 1984 г.). «Молекулярный анализ локуса периода у Drosophila melanogaster и идентификация транскрипта, участвующего в биологических ритмах». Клетка . 38 (3): 701–710. дои : 10.1016/0092-8674(84)90265-4 . ПМИД 6435882 . S2CID 316424 .
- ^ Церинг В.А., Уиллер Д.А., Редди П., Конопка Р.Дж., Кириаку К.П., Росбаш М. и др. (декабрь 1984 г.). «Трансформация P-элемента с помощью ДНК локуса периода восстанавливает ритмичность мутантной аритмичной Drosophila melanogaster» . Клетка . 39 (2, часть 1): 369–376. дои : 10.1016/0092-8674(84)90015-1 . ПМИД 6094014 .
- ^ Барджиелло Т.А., Джексон Ф.Р., Янг М.В. (1984). «Восстановление циркадных поведенческих ритмов путем переноса генов у дрозофилы». Природа . 312 (5996): 752–754. Бибкод : 1984Natur.312..752B . дои : 10.1038/312752a0 . ПМИД 6440029 . S2CID 4259316 .
- ^ Барджиелло Т.А., Янг М.В. (апрель 1984 г.). «Молекулярная генетика биологических часов у дрозофилы» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (7): 2142–2146. Бибкод : 1984Natur.312..752B . дои : 10.1038/312752a0 . ПМЦ 345453 . ПМИД 16593450 .
- ^ «Нобелевская премия по физиологии и медицине 2017» . www.nobelprize.org . Проверено 6 октября 2017 г.
- ^ [ ненадежный медицинский источник? ] «У мышей обнаружен ген, регулирующий биологические часы». Чикаго Трибьюн . 29 апреля 1994 г.
- ^ [ нужен неосновной источник ] Витатерна М.Х., Кинг Д.П., Чанг А.М., Корнхаузер Дж.М., Лоури П.Л., Макдональд Дж.Д. и др. (апрель 1994 г.). «Мутагенез и картирование мышиного гена Clock, необходимого для циркадного поведения» . Наука . 264 (5159): 719–725. Бибкод : 1994Sci...264..719H . дои : 10.1126/science.8171325 . ПМЦ 3839659 . ПМИД 8171325 .
- ^ Дебрюйн Дж.П., Нотон Э., Ламберт К.М., Мэйвуд Э.С., Уивер Д.Р., Репперт С.М. (май 2006 г.). «Потрясение часов: ЧАСЫ мыши не требуются для функции циркадного осциллятора» . Нейрон . 50 (3): 465–477. дои : 10.1016/j.neuron.2006.03.041 . ПМИД 16675400 . S2CID 19028601 .
- ^ Коллинз Б., Блау Дж. (май 2006 г.). «Определение времени без часов» . Нейрон . 50 (3): 348–350. дои : 10.1016/j.neuron.2006.04.022 . ПМИД 16675389 .
- ^ Тох К.Л., Джонс С.Р., Хе Ю., Эйде Э.Дж., Хинц В.А., Виршуп Д.М. и др. (февраль 2001 г.). «Мутация сайта фосфорилирования hPer2 при семейном синдроме развитой фазы сна». Наука . 291 (5506): 1040–1043. Бибкод : 2001Sci...291.1040T . дои : 10.1126/science.1057499 . ПМИД 11232563 . S2CID 1848310 .
- ^ Джонс С.Р., Кэмпбелл С.С., Зона С.В., Купер Ф., ДеСано А., Мерфи П.Дж. и др. (сентябрь 1999 г.). «Семейный синдром продвинутой фазы сна: вариант короткопериодного циркадного ритма у человека». Природная медицина . 5 (9): 1062–1065. дои : 10.1038/12502 . ПМИД 10470086 . S2CID 14809619 .
- ^ Джонсон С. (2004). Хронобиология: биологический хронометраж . Сандерленд, Массачусетс, США: Sinauer Associates, Inc., стр. 67–105.
- ^ Шарма В.К. (ноябрь 2003 г.). «Адаптационное значение циркадных часов». Хронобиология Интернэшнл . 20 (6): 901–919. дои : 10.1081/CBI-120026099 . ПМИД 14680135 . S2CID 10899279 .
- ^ [ нужен неосновной источник ] Шиба В., Шарма В.К., Чандрашекаран М.К., Джоши А. (сентябрь 1999 г.). «Сохранение ритма эклозии у Drosophila melanogaster после 600 поколений в апериодической среде». Die Naturwissenschaften . 86 (9): 448–449. Бибкод : 1999NW.....86..448S . дои : 10.1007/s001140050651 . ПМИД 10501695 . S2CID 13401297 .
- ^ [ нужен неосновной источник ] Гайомарх С., Люмино С., Ричард Дж. П. (май 1998 г.). «Циркадный ритм активности японских перепелов в постоянной темноте: изменчивость ясности и возможность отбора». Хронобиология Интернэшнл . 15 (3): 219–230. дои : 10.3109/07420529808998685 . ПМИД 9653576 .
- ^ [ нужен неосновной источник ] Живкович Б.Д., Андервуд Х., Стил Коннектикут, Эдмондс К. (октябрь 1999 г.). «Формальные свойства циркадной и фотопериодической систем японского перепела: кривая фазового отклика и эффекты Т-циклов» . Журнал биологических ритмов . 14 (5): 378–390. дои : 10.1177/074873099129000786 . ПМИД 10511005 . S2CID 13390422 .
- ^ Мори Т., Джонсон CH (апрель 2001 г.). «Независимость циркадного времени от деления клеток у цианобактерий» . Журнал бактериологии . 183 (8): 2439–2444. дои : 10.1128/JB.183.8.2439-2444.2001 . ПМК 95159 . ПМИД 11274102 .
- ^ Хат РА, Беерсма ГД (июль 2011 г.). «Эволюция механизмов хронометража: раннее возникновение и адаптация к фотопериоду» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 366 (1574): 2141–2154. дои : 10.1098/rstb.2010.0409 . ПМК 3130368 . ПМИД 21690131 .
- ^ Дубовый С., Сегал А. (апрель 2017 г.). «Циркадные ритмы и сон у Drosophila melanogaster » . Генетика . 205 (4): 1373–1397. дои : 10.1534/genetics.115.185157 . ПМЦ 5378101 . ПМИД 28360128 .
- ^ [ ненадежный медицинский источник? ] Нагоши Э., Сайни С., Бауэр С., Ларош Т., Наеф Ф., Шиблер У. (ноябрь 2004 г.). «Циркадная экспрессия генов в отдельных фибробластах: клеточно-автономные и автономные осцилляторы передают время дочерним клеткам» . Клетка . 119 (5): 693–705. дои : 10.1016/j.cell.2004.11.015 . ПМИД 15550250 . S2CID 15633902 .
- ^ [ нужен неосновной источник ] Мишель С., Гойш М.Е., Зарицкий Дж.Дж., Блок Г.Д. (январь 1993 г.). «Циркадный ритм мембранной проводимости, выраженный в изолированных нейронах» . Наука . 259 (5092): 239–241. Бибкод : 1993Sci...259..239M . дои : 10.1126/science.8421785 . ПМИД 8421785 .
- ^ Рефинетти Р. (январь 2010 г.). «Циркадный ритм температуры тела» . Границы бионауки . 15 (2): 564–594. дои : 10.2741/3634 . ПМИД 20036834 . S2CID 36170900 .
- ^ Шеер Ф.А., Моррис С.Дж., Ши С.А. (март 2013 г.). «Внутренние циркадные часы усиливают голод и аппетит вечером независимо от приема пищи и другого поведения» . Ожирение . 21 (3): 421–423. дои : 10.1002/oby.20351 . ПМЦ 3655529 . ПМИД 23456944 .
- ^ [ ненадежный медицинский источник? ] Живкович БК (25 июля 2007 г.). «Урок по часам № 16: Фотопериодизм – модели и экспериментальные подходы (оригинальная работа от 13 августа 2005 г.)» . Блог круглосуточно . Научные блоги. Архивировано из оригинала 1 января 2008 г. Проверено 9 декабря 2007 г.
- ^ [ нужен неосновной источник ] Турек Ф.В., Джошу С., Косака А., Лин Э., Иванова Г., МакДирмон Э. и др. (май 2005 г.). «Ожирение и метаболический синдром у мышей-мутантов по циркадным часам» . Наука . 308 (5724): 1043–1045. Бибкод : 2005Sci...308.1043T . дои : 10.1126/science.1108750 . ПМЦ 3764501 . ПМИД 15845877 .
- ^ Делези Дж., Дюмон С., Дарденте Х., Удар Х., Греше-Кассио А., Клозен П. и др. (август 2012 г.). «Ядерный рецептор REV-ERBα необходим для ежедневного баланса углеводного и липидного обмена» . Журнал ФАСЭБ . 26 (8): 3321–3335. дои : 10.1096/fj.12-208751 . ПМИД 22562834 . S2CID 31204290 .
- ^ [ нужен неосновной источник ] Делези Дж., Дюмон С., Дарденте Х., Удар Х., Греше-Кассио А., Клозен П. и др. (август 2012 г.). «Ядерный рецептор REV-ERBα необходим для ежедневного баланса углеводного и липидного обмена» . Журнал ФАСЭБ . 26 (8): 3321–3335. дои : 10.1096/fj.12-208751 . ПМИД 22562834 . S2CID 31204290 .
- ^ [ нужен неосновной источник ] Скотт Э.М., Картер А.М., Грант П.Дж. (апрель 2008 г.). «Связь между полиморфизмом гена Clock, ожирением и метаболическим синдромом у человека» . Международный журнал ожирения . 32 (4): 658–662. дои : 10.1038/sj.ijo.0803778 . ПМИД 18071340 .
- ^ [ ненадежный медицинский источник? ] Шнеерсон Дж. М., Охайон М. М., Карскадон М. А. (2007). «Циркадные ритмы» . Сон с быстрым движением глаз (БДГ) . Армянская медицинская сеть . Проверено 19 сентября 2007 г.
- ^ «Ритмы жизни: биологические часы, которые контролируют повседневную жизнь каждого живого существа» Рассел Фостер и Леон Крейцман, Издатель: Profile Books Ltd.
- ^ [ ненадежный медицинский источник? ] Регештейн QR, Павлова М (сентябрь 1995 г.). «Лечение синдрома задержки фазы сна». Общая больничная психиатрия . 17 (5): 335–345. дои : 10.1016/0163-8343(95)00062-В . ПМИД 8522148 .
- ^ Хауэлл Э. (14 декабря 2012 г.). «Космическая станция получит новые лампочки для борьбы с бессонницей» . Space.com . Проверено 17 декабря 2012 г.
- ^ [ нужен неосновной источник ] Пролитый I (декабрь 2005 г.). «Северный олень без циркадного ритма» (на норвежском букмоле). forskning.no Архивировано из оригинала 3 декабря 2007 г. Проверено 24 ноября 2007 г.
...не похоже, что у оленей летом отсутствовал какой-либо циркадный ритм. Зимой его не было и у шпицбергенских оленей.
- ^ Фолк GE, Thrift DL, Циммерман МБ, Рейманн П (01 декабря 2006 г.). «Активность млекопитающих – ритмы отдыха при непрерывном дневном свете в Арктике». Исследование биологического ритма . 37 (6): 455–469. Бибкод : 2006BioRR..37..455F . дои : 10.1080/09291010600738551 . S2CID 84625255 .
Могут ли местные животные, содержащиеся при естественном постоянном дневном освещении, демонстрировать эффект Ашоффа, описанный в ранее опубликованных лабораторных экспериментах с использованием непрерывного света, в которых циркадная активность крыс систематически менялась на более длительный период, выражающий 26-часовой день активности и отдыха?
- ^ [ нужен неосновной источник ] Мерлин С., Гегир Р.Дж., Репперт С.М. (сентябрь 2009 г.). «Усиковые циркадные часы координируют ориентацию по солнечному компасу у мигрирующих бабочек-монархов» . Наука . 325 (5948): 1700–1704. Бибкод : 2009Sci...325.1700M . дои : 10.1126/science.1176221 . ПМЦ 2754321 . ПМИД 19779201 .
- ^ [ нужен неосновной источник ] Кириаку КП (сентябрь 2009 г.). «Физиология. Разгадка путешествия». Наука . 325 (5948): 1629–1630. дои : 10.1126/science.1178935 . ПМИД 19779177 . S2CID 206522416 .
- ^ Сильвегрен Г., Лёфстедт С., Ци Розен В. (март 2005 г.). «Циркадная брачная активность и влияние предварительного воздействия феромонов на ритмы реакции феромонов у бабочки Spodopteralittoralis». Журнал физиологии насекомых . 51 (3): 277–286. Бибкод : 2005JInsP..51..277S . дои : 10.1016/j.jinsphys.2004.11.013 . ПМИД 15749110 .
- ↑ Перейти обратно: Перейти обратно: а б Уэбб А.А. (ноябрь 2003 г.). «Физиология циркадных ритмов у растений» . Новый фитолог . 160 (2): 281–303. дои : 10.1046/j.1469-8137.2003.00895.x . JSTOR 1514280 . ПМИД 33832173 . S2CID 15688409 .
- ↑ Перейти обратно: Перейти обратно: а б с МакКлунг CR (апрель 2006 г.). «Циркадные ритмы растений» . Растительная клетка . 18 (4): 792–803. дои : 10.1105/tpc.106.040980 . ПМЦ 1425852 . ПМИД 16595397 .
- ^ Легрис М., Инс Ю., Фанкхаузер С. (ноябрь 2019 г.). «Молекулярные механизмы, лежащие в основе фитохром-контролируемого морфогенеза у растений» . Природные коммуникации . 10 (1): 5219. Бибкод : 2019NatCo..10.5219L . дои : 10.1038/s41467-019-13045-0 . ПМК 6864062 . ПМИД 31745087 .
- ^ Мидзогучи Т., Райт Л., Фудзивара С., Кремер Ф., Ли К., Оноучи Х. и др. (август 2005 г.). «Особая роль GIGANTEA в содействии цветению и регулировании циркадных ритмов Arabidopsis» . Растительная клетка . 17 (8): 2255–2270. дои : 10.1105/tpc.105.033464 . ПМЦ 1182487 . ПМИД 16006578 .
- ^ Колмос Э., Дэвис С.Дж. (сентябрь 2007 г.). «ELF4 как центральный ген циркадных часов» . Сигнализация и поведение растений . 2 (5): 370–372. Бибкод : 2007ПлСиБ...2..370К . дои : 10.4161/psb.2.5.4463 . ПМК 2634215 . ПМИД 19704602 .
- ^ Похилко А., Фернандес А.П., Эдвардс К.Д., Саузерн ММ, Холлидей К.Дж., Миллар А.Дж. (март 2012 г.). «Цепь часового гена Arabidopsis включает в себя репрессилятор с дополнительными петлями обратной связи» . Молекулярная системная биология . 8 : 574. дои : 10.1038/msb.2012.6 . ПМК 3321525 . ПМИД 22395476 .
- ^ Ма Ю, Гил С., Грассер К.Д., Мас П. (апрель 2018 г.). «Направленное задействование базального транскрипционного механизма с помощью компонентов часов LNK контролирует циркадные ритмы возникающих РНК у Arabidopsis» . Растительная клетка . 30 (4): 907–924. дои : 10.1105/tpc.18.00052 . ПМЦ 5973845 . ПМИД 29618629 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж Додд А.Н., Салатия Н., Холл А., Кевей Э., Тот Р., Надь Ф. и др. (июль 2005 г.). «Циркадные часы растений увеличивают фотосинтез, рост, выживаемость и конкурентное преимущество». Наука . 309 (5734): 630–633. Бибкод : 2005Sci...309..630D . дои : 10.1126/science.1115581 . ПМИД 16040710 . S2CID 25739247 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Додд А.Н., Белбин Ф.Е., Фрэнк А., Уэбб А.А. (2015). «Взаимодействие между циркадными часами и фотосинтезом для временной и пространственной координации метаболизма» . Границы в науке о растениях . 6 : 245. doi : 10.3389/fpls.2015.00245 . ПМЦ 4391236 . ПМИД 25914715 .
- ^ Уэбб А.А., Секи М., Сатаке А., Калдана С. (февраль 2019 г.). «Непрерывная динамическая настройка циркадного осциллятора растений» . Природные коммуникации . 10 (1): 550. Бибкод : 2019NatCo..10..550W . дои : 10.1038/s41467-019-08398-5 . ПМК 6358598 . ПМИД 30710080 .
- ↑ Перейти обратно: Перейти обратно: а б Хейдон М.Дж., Мельчарек О., Робертсон Ф.К., Хаббард К.Е., Уэбб А.А. (октябрь 2013 г.). «Фотосинтетическое увлечение циркадных часов Arabidopsis thaliana» . Природа . 502 (7473): 689–692. Бибкод : 2013Natur.502..689H . дои : 10.1038/nature12603 . ПМЦ 3827739 . ПМИД 24153186 .
- ^ Фарре Э.М., Кей С.А. (ноябрь 2007 г.). «Уровень белка PRR7 регулируется светом и циркадными часами у арабидопсиса» . Заводской журнал . 52 (3): 548–560. дои : 10.1111/j.1365-313X.2007.03258.x . ПМИД 17877705 .
- ^ Велери С., Вюльбек С. (май 2004 г.). «Уникальные автономные циркадные осцилляторы в мозгу Drosophila melanogaster». Хронобиология Интернэшнл . 21 (3): 329–342. дои : 10.1081/CBI-120038597 . ПМИД 15332440 . S2CID 15099796 .
- ^ Йоши Т., Герман-Луибль С., Хелфрих-Фёрстер С. (2015). «Циркадные пути поступления света у дрозофилы» . Коммуникативная и интегративная биология . 9 (1): e1102805. дои : 10.1080/19420889.2015.1102805 . ПМЦ 4802797 . ПМИД 27066180 .
- ^ Бутройд CE, Янг М.В. (2008). «Входы и выходы циркадных часов дрозофилы». Анналы Нью-Йоркской академии наук . 1129 (1): 350–357. Бибкод : 2008NYASA1129..350B . дои : 10.1196/анналы.1417.006 . ПМИД 18591494 . S2CID 2639040 .
- ^ Грима Б, Ламуру А, Шело Э, Папен С, Лимбург-Бушон Б, Руйе Ф (ноябрь 2002 г.). «Протеин F-box slimm контролирует уровни часовых белков периодического и вневременного». Природа . 420 (6912): 178–182. Бибкод : 2002Natur.420..178G . дои : 10.1038/nature01122 . ПМИД 12432393 . S2CID 4428779 .
- ^ Ко Х.В., Цзян Дж., Эдери И. (декабрь 2002 г.). «Роль Slimb в деградации белка периода дрозофилы, фосфорилированного Doubletime». Природа . 420 (6916): 673–678. Бибкод : 2002Natur.420..673K . дои : 10.1038/nature01272 . ПМИД 12442174 . S2CID 4414176 .
- ^ Хелфрих-Фёрстер С (март 2005 г.). «Нейробиология циркадных часов плодовой мухи» . Гены, мозг и поведение . 4 (2): 65–76. дои : 10.1111/j.1601-183X.2004.00092.x . ПМИД 15720403 . S2CID 26099539 .
- ^ Лалчхандама К. (2017). «Путь к Нобелевской премии по физиологии и медицине 2017 года» . Научное видение . 3 (Приложение): 1–13.
- ↑ Перейти обратно: Перейти обратно: а б с Лелуп Дж.К., Голдбетер А. (февраль 1998 г.). «Модель циркадных ритмов у дрозофилы, включающая образование комплекса между белками PER и TIM». Журнал биологических ритмов . 13 (1): 70–87. дои : 10.1177/074873098128999934 . ПМИД 9486845 . S2CID 17944849 .
- ^ Гольдбетер А (сентябрь 1995 г.). «Модель циркадных колебаний белка периода дрозофилы (PER)». Слушания. Биологические науки . 261 (1362): 319–324. Бибкод : 1995РСПСБ.261..319Г . дои : 10.1098/rspb.1995.0153 . ПМИД 8587874 . S2CID 7024361 .
- ↑ Перейти обратно: Перейти обратно: а б Гольдбетер А. (ноябрь 2002 г.). «Вычислительные подходы к клеточным ритмам». Природа . 420 (6912): 238–245. Бибкод : 2002Natur.420..238G . дои : 10.1038/nature01259 . ПМИД 12432409 . S2CID 452149 .
- ^ «Биологические часы у млекопитающих» . БиоИнтерактив . Медицинский институт Говарда Хьюза. 4 февраля 2000 г. Архивировано из оригинала 5 мая 2015 г. Проверено 5 мая 2015 г.
- ^ Уэльс Д.К., Такахаши Дж.С., Кей С.А. (март 2010 г.). «Супрахиазматическое ядро: клеточная автономия и сетевые свойства» . Ежегодный обзор физиологии . 72 : 551–77. doi : 10.1146/annurev-psyol-021909-135919 . ПМЦ 3758475 . ПМИД 20148688 .
- ^ Пфеффер М., Корф Х.В., Вихт Х. (март 2018 г.). «Синхронизация воздействия мелатонина на суточные и циркадные ритмы». Общая и сравнительная эндокринология . 258 : 215–221. дои : 10.1016/j.ygcen.2017.05.013 . ПМИД 28533170 .
- ^ Калпеш Дж. «Оздоровление при искусственном свете» . Проверено 11 января 2016 г.
- ^ [ ненадежный медицинский источник? ] Шеер Ф.А., Райт К.П., Кронауер Р.Е., Чейслер К.А. (август 2007 г.). «Пластичность внутреннего периода циркадной системы времени человека» . ПЛОС ОДИН . 2 (8): е721. Бибкод : 2007PLoSO...2..721S . дои : 10.1371/journal.pone.0000721 . ЧВК 1934931 . ПМИД 17684566 .
- ^ [ ненадежный медицинский источник? ] Даффи Дж. Ф., Райт К. П. (август 2005 г.). «Увлечение циркадной системы человека светом». Журнал биологических ритмов . 20 (4): 326–38. дои : 10.1177/0748730405277983 . ПМИД 16077152 . S2CID 20140030 .
- ^ Хальса С.Б., Джуэтт М.Э., Кайохен С., Чейслер Калифорния (июнь 2003 г.). «Кривая фазового отклика на одиночные яркие световые импульсы у людей» . Журнал физиологии . 549 (Часть 3): 945–52. дои : 10.1113/jphysicalol.2003.040477 . ПМК 2342968 . ПМИД 12717008 .
- ^ Кроми В (15 июля 1999 г.). «Биологические часы человека переведены на час назад» . Гарвардская газета . Проверено 4 июля 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б Даффи Дж.Ф., Кейн С.В., Чанг А.М., Филлипс А.Дж., Мюнх М.Ю., Гронфье С. и др. (сентябрь 2011 г.). «Половые различия в почти 24-часовом внутреннем периоде циркадной системы времени человека» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (Дополнение_3): 15602–8. Бибкод : 2011PNAS..10815602D . дои : 10.1073/pnas.1010666108 . ПМК 3176605 . ПМИД 21536890 .
- ↑ Перейти обратно: Перейти обратно: а б с Бенлусиф С., Гико М.Дж., Рид К.Дж., Вулф Л.Ф., Л'Эрмит-Балерио М., Зи ПК (апрель 2005 г.). «Стабильность мелатонина и температуры как маркеров циркадной фазы и их связь со временем сна у людей» . Журнал биологических ритмов . 20 (2): 178–88. дои : 10.1177/0748730404273983 . ПМИД 15834114 . S2CID 36360463 .
- ^ Адам Е.К., Куинн М.Э., Тавернье Р., Маккуиллан М.Т., Дальке К.А., Гилберт К.Е. (сентябрь 2017 г.). «Суточные наклоны кортизола и последствия для психического и физического здоровья: систематический обзор и метаанализ» . Психонейроэндокринология . 83 : 25–41. doi : 10.1016/j.psyneuen.2017.05.018 . ПМЦ 5568897 . ПМИД 28578301 .
- ^ Баер Э.К., Ревелл В., Истман К.И. (июнь 2000 г.). «Индивидуальные различия фазы и амплитуды циркадного температурного ритма человека: с акцентом на утро-вечер» . Журнал исследований сна . 9 (2): 117–27. дои : 10.1046/j.1365-2869.2000.00196.x . ПМИД 10849238 . S2CID 6104127 .
- ^ Зейзер Л., Корнелиссен-Гийом Г., Шипек Г.К., Чамсон Э., Блим Х.Р. и Шуберт С. (2022) Примерно еженедельная закономерность в динамической сложности клеточной иммунной активности здорового субъекта: биопсихосоциальный анализ. Передний. Психиатрия 13:799214. дои: 10.3389/fpsyt.2022.799214
- ^ «Семинар NHLBI: «Циркадные часы на взаимосвязи здоровья легких и заболеваний» 28-29 апреля 2014 г. Краткое содержание» . Национальный институт сердца, легких и крови. Сентябрь 2014 г. Архивировано из оригинала 04 октября 2014 г. Проверено 20 сентября 2014 г.
- ^ Дегауте Дж. П., ван де Борн П., Линковски П., Ван Каутер Э. (август 1991 г.). «Количественный анализ суточных показателей артериального давления и сердечного ритма у юношей» . Гипертония . 18 (2): 199–210. дои : 10.1161/01.hyp.18.2.199 . ПМИД 1885228 .
- ^ Квартель Л (2014). «Влияние циркадного ритма температуры тела на результаты экзамена A-level» . Бакалавриат по психологии . 27 (1).
- ^ Ирокез Дж.А., Грин CB, Такахаши Дж.С. (14 июля 2013 г.). «Центральные и периферические циркадные часы млекопитающих» . Ежегодный обзор неврологии . 35 : 445–62. doi : 10.1146/annurev-neuro-060909-153128 . ПМЦ 3710582 . ПМИД 22483041 .
- ^ Там же.
- ^ Пендергаст Дж. С., Нисвендер К. Д., Ямазаки С. (11 января 2012 г.). «Тканеспецифическая функция Period3 в циркадной ритмичности» . ПЛОС ОДИН . 7 (1): e30254. Бибкод : 2012PLoSO...730254P . дои : 10.1371/journal.pone.0030254 . ПМК 3256228 . ПМИД 22253927 .
- ^ Сингх М. (10 октября 2013 г.). «Чувство времени нашей кожи помогает защититься от ультрафиолетового излучения» . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . Проверено 19 февраля 2019 г.
- ^ Абрахам У., Гранада А.Е., Вестермарк П.О., Хейне М., Крамер А., Герцель Х. (ноябрь 2010 г.). «Связь управляет диапазоном увлечения циркадных часов» . Молекулярная системная биология . 6 : 438. дои : 10.1038/msb.2010.92 . ПМК 3010105 . ПМИД 21119632 .
- ^ Цао К., Гери С., Дашти А., Инь Д., Чжоу Ю., Гу Дж. и др. (октябрь 2009 г.). «Роль часового гена per1 при раке простаты» . Исследования рака . 69 (19): 7619–25. doi : 10.1158/0008-5472.CAN-08-4199 . ПМЦ 2756309 . ПМИД 19752089 .
- ^ Кавара С., Мидларски Р., Мамелак А.Дж., Фрид И., Ван Б., Ватанабэ Х. и др. (декабрь 2002 г.). «Низкие дозы ультрафиолетовых лучей B изменяют экспрессию мРНК генов циркадных часов в культивируемых кератиноцитах человека» . Журнал исследовательской дерматологии . 119 (6): 1220–3. дои : 10.1046/j.1523-1747.2002.19619.x . ПМИД 12485420 .
- ^ Дамиола Ф, Ле Мин Н, Прейтнер Н, Корнманн Б, Флери-Олела Ф, Шиблер Ю (декабрь 2000 г.). «Ограниченное питание разъединяет циркадные осцилляторы в периферических тканях от центрального водителя ритма в супрахиазматическом ядре» . Гены и развитие . 14 (23): 2950–61. дои : 10.1101/gad.183500 . ПМК 317100 . ПМИД 11114885 .
- ^ Даффи Дж. Ф., Чейслер, Калифорния (июнь 2009 г.). «Влияние света на циркадную физиологию человека» . Клиники медицины сна . 4 (2): 165–177. дои : 10.1016/j.jsmc.2009.01.004 . ПМЦ 2717723 . ПМИД 20161220 .
- ^ Цейслер К.А., Даффи Дж.Ф., Шанахан Т.Л., Браун Э.Н., Митчелл Дж.Ф., Риммер Д.В. и др. (июнь 1999 г.). «Стабильность, точность и почти 24-часовой период работы циркадного водителя ритма человека». Наука . 284 (5423): 2177–81. дои : 10.1126/science.284.5423.2177 . ПМИД 10381883 . S2CID 8516106 .
- ^ Олдрич М.С. (1999). Лекарство для сна . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-512957-1 .
- ^ Вятт Дж.К., Ритц-Де Чекко А., Цейслер Калифорния, Дейк DJ (октябрь 1999 г.). «Циркадная температура и ритмы мелатонина, сон и нейроповеденческие функции у людей, живущих с 20-часовым днем». Американский журнал физиологии . 277 (4 ч. 2): R1152-63. дои : 10.1152/ajpregu.1999.277.4.R1152 . ПМИД 10516257 . S2CID 4474347 .
- ^ Райт К.П., Халл Дж.Т., Чейслер Калифорния (декабрь 2002 г.). «Взаимосвязь между бдительностью, работоспособностью и температурой тела у людей». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 283 (6): Р1370-7. CiteSeerX 10.1.1.1030.9291 . дои : 10.1152/ajpregu.00205.2002 . ПМИД 12388468 .
- ^ Чжоу X, Фергюсон С.А., Мэтьюз Р.В., Сарджент С., Дарвент Д., Кеннауэй DJ и др. (июль 2011 г.). «Сон, бодрствование и фазозависимые изменения нейроповеденческих функций в условиях вынужденной десинхронии» . Спать . 34 (7): 931–41. дои : 10.5665/SLEEP.1130 . ПМК 3119835 . ПМИД 21731143 .
- ^ Космадопулос А., Сарджент С., Дарвент Д., Чжоу Х., Доусон Д., Роуч Г.Д. (декабрь 2014 г.). «Влияние разделенного графика сна и бодрствования на нейроповеденческие функции и прогнозы производительности в условиях вынужденной десинхронии». Хронобиология Интернэшнл . 31 (10): 1209–17. дои : 10.3109/07420528.2014.957763 . ПМИД 25222348 . S2CID 11643058 .
- ↑ Перейти обратно: Перейти обратно: а б Алибхай Ф.Дж., Цимакуридзе Е.В., Рейтц К.Дж., Пайл В.Г., Мартино Т.А. (июль 2015 г.). «Последствия циркадных нарушений и нарушений сна для сердечно-сосудистой системы». Канадский журнал кардиологии . 31 (7): 860–872. дои : 10.1016/j.cjca.2015.01.015 . ПМИД 26031297 .
- ^ Мартино Т.А., Young ME (июнь 2015 г.). «Влияние циркадных часов кардиомиоцитов на физиологию и патофизиологию сердца» . Журнал биологических ритмов . 30 (3): 183–205. дои : 10.1177/0748730415575246 . ПМИД 25800587 . S2CID 21868234 .
- ^ Мистри П., Дуонг А., Киршенбаум Л., Мартино Т.А. (октябрь 2017 г.). «Сердечные часы и доклинический перевод» . Клиники сердечной недостаточности . 13 (4): 657–672. дои : 10.1016/j.hfc.2017.05.002 . ПМИД 28865775 .
- ^ Азиз И.С., МакМахон А.М., Фридман Д., Рабинович-Никитин И., Киршенбаум Л.А., Мартино Т.А. (апрель 2021 г.). «Циркадное влияние на воспалительную реакцию при сердечно-сосудистых заболеваниях» . Современное мнение в фармакологии . 57 : 60–70. дои : 10.1016/j.coph.2020.11.007 . ПМИД 33340915 . S2CID 229332749 .
- ^ Гроте Л., Майер Дж., Пензель Т., Кассель В., Кржизанек Э., Питер Дж.Х. и др. (1994). «Ночная гипертония и сердечно-сосудистый риск: последствия для диагностики и лечения». Журнал сердечно-сосудистой фармакологии . 24 (Приложение 2): С26–С38. дои : 10.1097/00005344-199412001-00006 . ПМИД 7898092 .
- ^ Рейтц С.Дж., Алибхай Ф.Дж., Хатуа Т.Н., Расули М., Бридл Б.В., Беррис Т.П. и др. (2019). «SR9009, вводимый в течение одного дня после ишемии-реперфузии миокарда, предотвращает сердечную недостаточность у мышей, воздействуя на сердечную инфламмасому» . Коммуникационная биология . 2 : 353. doi : 10.1038/s42003-019-0595-z . ПМК 6776554 . ПМИД 31602405 .
- ^ Алибхай Ф.Дж., Рейтц С.Дж., Пепплер В.Т., Басу П., Шеппард П., Холерис Э. и др. (февраль 2018 г.). «Самки мышей ClockΔ19/Δ19 защищены от развития возрастной кардиомиопатии» . Сердечно-сосудистые исследования . 114 (2): 259–271. дои : 10.1093/cvr/cvx185 . ПМИД 28927226 .
- ^ Алибхай Ф.Дж., ЛаМарр Дж., Рейтц К.Дж., Цимакуридзе Е.В., Кроетч Дж.Т., Больц С.С. и др. (апрель 2017 г.). «Нарушение ключевого циркадного регулятора ЧАСОВ приводит к возрастным сердечно-сосудистым заболеваниям» . Журнал молекулярной и клеточной кардиологии . 105 : 24–37. дои : 10.1016/j.yjmcc.2017.01.008 . ПМИД 28223222 .
- ^ Беннардо М., Алибхай Ф., Цимакуридзе Е., Чиннапаредди Н., Подобед П., Рейтц С. и др. (декабрь 2016 г.). «Зависимость экспрессии генов и воспалительных реакций от дня и ночи при ремоделировании мышиного сердца после инфаркта миокарда». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 311 (6): R1243–R1254. дои : 10.1152/ajpregu.00200.2016 . ПМИД 27733386 . S2CID 36325095 .
- ^ Пайл В.Г., Мартино Т.А. (октябрь 2018 г.). «Циркадные ритмы по-разному влияют на сердечно-сосудистые заболевания у мужчин и женщин: роль пола и гендера» . Современное мнение в физиологии . 5 : 30–37. doi : 10.1016/j.cophys.2018.05.003 . ISSN 2468-8673 . S2CID 80632426 .
- ^ Ри М.С., Фигейро М., Буллоу Дж. (май 2002 г.). «Циркадная фотобиология: новая основа практики и исследований освещения». Световые исследования и технологии . 34 (3): 177–187. дои : 10.1191/1365782802lt057oa . S2CID 109776194 .
- ^ Уолмсли Л., Ханна Л., Муланд Дж., Марсьяль Ф., Уэст А., Смедли А.Р. и др. (апрель 2015 г.). «Цвет как сигнал для включения циркадных часов млекопитающих» . ПЛОС Биология . 13 (4): e1002127. дои : 10.1371/journal.pbio.1002127 . ПМЦ 4401556 . ПМИД 25884537 .
- ^ Хардт Р. (1 января 1970 г.). «Опасности светодиодного синего света. Подавление мелатонина, приводящее к бессоннице и раку | Роберт Хардт» . Академия.edu . Проверено 24 декабря 2016 г. [ постоянная мертвая ссылка ]
- ^ Бедросян Т.А., Нельсон Р.Дж. (январь 2017 г.). «Время воздействия света влияет на настроение и работу мозга» . Трансляционная психиатрия . 7 (1): e1017. дои : 10.1038/tp.2016.262 . ПМЦ 5299389 . ПМИД 28140399 .
- ^ [1] [ мертвая ссылка ]
- ^ Нарушение циркадного ритма и полеты. ФАУ на https://www.faa.gov/pilots/safety/pilotsafetybrochures/media/Circadian_Rhythm.pdf. Архивировано 1 мая 2017 г. в Wayback Machine.
- ↑ Перейти обратно: Перейти обратно: а б с Логан Р.В., Уильямс В.П., МакКлунг, Калифорния (июнь 2014 г.). «Циркадные ритмы и зависимость: механистические идеи и будущие направления» . Поведенческая нейронаука . 128 (3): 387–412. дои : 10.1037/a0036268 . ПМК 4041815 . ПМИД 24731209 .
- ^ Проссер Р.А., Гласс JD (июнь 2015 г.). «Оценка действия этанола на супрахиазматические циркадные часы с использованием подходов in vivo и in vitro» . Алкоголь . 49 (4): 321–339. дои : 10.1016/j.alcohol.2014.07.016 . ПМК 4402095 . ПМИД 25457753 .
- ^ «Наука о смене часовых поясов» . Таймшифтер . Проверено 03 января 2023 г.
- ^ Голд АК, Кинрис Г (март 2019 г.). «Лечение нарушения циркадного ритма при биполярном расстройстве» . Текущие отчеты психиатрии . 21 (3): 14. дои : 10.1007/s11920-019-1001-8 . ПМК 6812517 . ПМИД 30826893 .
- ^ Чжу Л, Zee PC (ноябрь 2012 г.). «Нарушения циркадного ритма сна» . Неврологические клиники . 30 (4): 1167–1191. дои : 10.1016/j.ncl.2012.08.011 . ПМК 3523094 . ПМИД 23099133 .
- ↑ Перейти обратно: Перейти обратно: а б с Зелински Э.Л., Дейбель С.Х., Макдональд Р.Дж. (март 2014 г.). «Проблема с дисфункцией циркадных часов: множественные вредные воздействия на мозг и тело». Неврологические и биоповеденческие обзоры . 40 (40): 80–101. doi : 10.1016/j.neubiorev.2014.01.007 . ПМИД 24468109 . S2CID 6809964 .
- ^ Ориц-Тульдела Э, Мартинес-Николас А, Диас-Мардоминго С, Гарсия-Эрранс С, Переда-Перес I, Валенсия А, Пераита Х, Венеро С, Мадрид Дж, Рол М. 2014. Характеристика биологических ритмов при легкой форме Когнитивные нарушения. БиоМед Исследования Интернэшнл.
- ^ Хершнер С.Д., Червин Р.Д. (23 июня 2014 г.). «Причины и последствия сонливости у студентов» . Природа и наука сна . 6 : 73–84. дои : 10.2147/NSS.S62907 . ПМК 4075951 . ПМИД 25018659 .
- ^ Милнер CE, Кот, штат Калифорния (июнь 2009 г.). «Преимущества дневного сна у здоровых взрослых: влияние продолжительности сна, времени суток, возраста и опыта дневного сна» . Журнал исследований сна . 18 (2): 272–281. дои : 10.1111/j.1365-2869.2008.00718.x . ПМИД 19645971 . S2CID 22815227 .
- ^ Ловато Н., Лак Л. (2010). Влияние дневного сна на когнитивные функции . Прогресс в исследованиях мозга. Том. 185. стр. 155–166. дои : 10.1016/B978-0-444-53702-7.00009-9 . ISBN 978-0-444-53702-7 . ПМИД 21075238 .
- ^ Синерт Т., Павлин PR (10 мая 2006 г.). «Почечная недостаточность, острая» . Электронная медицина от WebMD . Проверено 3 августа 2008 г.
- ^ Маунг С.К., Эль Сара А., Чепмен С., Коэн Д., Кьюкор Д. (май 2016 г.). «Нарушения сна и хроническая болезнь почек» . Всемирный журнал нефрологии . 5 (3): 224–232. дои : 10.5527/wjn.v5.i3.224 . ПМЦ 4848147 . ПМИД 27152260 .
- ^ Накано С., Учида К., Кигоши Т., Азукизава С., Ивасаки Р., Канеко М. и др. (август 1991 г.). «Циркадный ритм артериального давления у нормотензивных пациентов с ИНСД. Его связь с микрососудистыми осложнениями». Уход при диабете . 14 (8): 707–711. дои : 10.2337/diacare.14.8.707 . ПМИД 1954805 . S2CID 12489921 .
- ^ Фигейро М.Г., Ри М.С., Буллоу Дж.Д. (август 2006 г.). «Способствует ли архитектурное освещение развитию рака молочной железы?» . Журнал канцерогенеза . 5:20 . дои : 10.1186/1477-3163-5-20 . ПМЦ 1557490 . ПМИД 16901343 .
- ↑ Перейти обратно: Перейти обратно: а б Мартино Т.А., Оудит Г.Ю., Герценберг А.М., Тата Н., Колетар М.М., Кабир Г.М. и др. (май 2008 г.). «Деорганизация циркадного ритма приводит к тяжелым сердечно-сосудистым и почечным заболеваниям у хомяков». Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 294 (5): R1675–R1683. дои : 10.1152/ajpregu.00829.2007 . ПМИД 18272659 . S2CID 13356393 .
- ^ Шеер Ф.А., Хилтон М.Ф., Манцорос К.С., Ши С.А. (март 2009 г.). «Неблагоприятные метаболические и сердечно-сосудистые последствия нарушения циркадных ритмов» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (11): 4453–4458. дои : 10.1073/pnas.0808180106 . ПМЦ 2657421 . ПМИД 19255424 .
- ^ Шеер Ф.А., Майкельсон А.Д., Фрелингер А.Л., Эвонюк Х., Келли Э.Э., Маккарти М. и др. (2011). «Эндогенная циркадная система человека вызывает наибольшую активацию тромбоцитов в течение биологического утра, независимо от поведения» . ПЛОС ОДИН . 6 (9): e24549. Бибкод : 2011PLoSO...624549S . дои : 10.1371/journal.pone.0024549 . ПМК 3169622 . ПМИД 21931750 .
- ^ Рабинович-Никитин И., Либерман Б., Мартино Т.А., Киршенбаум Л.А. (февраль 2019 г.). «Циркадно-регулируемая гибель клеток при сердечно-сосудистых заболеваниях» . Тираж . 139 (7): 965–980. doi : 10.1161/CIRCULATIONAHA.118.036550 . ПМИД 30742538 . S2CID 73436800 .
- ↑ Перейти обратно: Перейти обратно: а б с Кервези Л., Космадопулос А., Бойвин Д.Б. (январь 2020 г.). «Метаболические и сердечно-сосудистые последствия сменной работы: роль циркадных нарушений и нарушений сна» . Европейский журнал неврологии . 51 (1): 396–412. дои : 10.1111/ejn.14216 . ПМИД 30357975 . S2CID 53031343 .
- ↑ Перейти обратно: Перейти обратно: а б с Пропер К.И., ван де Лангенберг Д., Роденбург В., Вермюлен Р.К., ван дер Бик А.Дж., ван Стеег Х. и др. (май 2016 г.). «Взаимосвязь между сменной работой и метаболическими факторами риска: систематический обзор продольных исследований». Американский журнал профилактической медицины . 50 (5): е147–е157. дои : 10.1016/j.amepre.2015.11.013 . ПМИД 26810355 .
- ↑ Перейти обратно: Перейти обратно: а б Делези Дж., Шале Э. (декабрь 2011 г.). «Взаимодействие между метаболизмом и циркадными часами: взаимные нарушения» . Анналы Нью-Йоркской академии наук . 1243 (1): 30–46. Бибкод : 2011NYASA1243...30D . дои : 10.1111/j.1749-6632.2011.06246.x . ПМИД 22211891 . S2CID 43621902 .
- ^ Джонстон Джей Ди (июнь 2014 г.). «Физиологические реакции на прием пищи в течение дня» . Обзоры исследований в области питания . 27 (1): 107–18. дои : 10.1017/S0954422414000055 . ПМК 4078443 . ПМИД 24666537 .
- ^ ван Дронгелен А., Бут Ч.Р., Меркус С.Л., Смид Т., ван дер Бик А.Дж. (июль 2011 г.). «Влияние сменной работы на изменение массы тела – систематический обзор продольных исследований» . Скандинавский журнал труда, окружающей среды и здоровья . 37 (4): 263–275. дои : 10.5271/sjweh.3143 . ПМИД 21243319 . S2CID 35457083 .
- ^ Вуд П.А., Ян X, Грушески В.Дж. (декабрь 2009 г.). «Часовые гены и рак» . Интегративная терапия рака . 8 (4): 303–308. дои : 10.1177/1534735409355292 . ПМИД 20042409 . S2CID 29808156 .
- ↑ Перейти обратно: Перейти обратно: а б Масри С., Сассоне-Корси П. (декабрь 2018 г.). «Новая связь между раком, обменом веществ и циркадными ритмами» . Природная медицина . 24 (12): 1795–1803. дои : 10.1038/s41591-018-0271-8 . ПМК 6535395 . ПМИД 30523327 .
- ^ Саламанка-Фернандес Э, Родригес-Барранко М, Гевара М, Арданас Э, Олри де Лабри Лима А, Санчес МХ (август 2018 г.). «Работа в ночную смену и риск рака груди и простаты: обновление данных эпидемиологических исследований» . Анналы санитарной системы Наварры . 41 (2): 211–226. дои : 10.23938/ASSN.0307 . ПМИД 30063040 . S2CID 51874242 .
- ^ Шернхаммер Э.С., Ладен Ф., Спейзер Ф.Е. , Уиллетт В.К., Хантер Д.Д., Кавачи И. и др. (октябрь 2001 г.). «Смена ночных смен и риск рака молочной железы у женщин, участвовавших в исследовании здоровья медсестер» . Журнал Национального института рака . 93 (20): 1563–1568. дои : 10.1093/jnci/93.20.1563 . ПМИД 11604480 .
- ^ Папантониу К., Кастаньо-Виньялс Г., Эспиноза А., Арагонес Н., Перес-Гомес Б., Бургос Дж. и др. (сентябрь 2015 г.). «Работа в ночную смену, хронотип и риск рака простаты в исследовании случай-контроль MCC-Испания». Международный журнал рака . 137 (5): 1147–1157. дои : 10.1002/ijc.29400 . ПМИД 25530021 . S2CID 19665388 .
- ^ Ли Дж. А., Россинк Дж., Кьерхайм К. (февраль 2006 г.). «Рак молочной железы и ночная работа норвежских медсестер». Причины рака и борьба с ним . 17 (1): 39–44. дои : 10.1007/s10552-005-3639-2 . ПМИД 16411051 . S2CID 6281682 .
- ^ Челлаппа С.Л., Моррис СиДжей, Шеер Ф.А. (январь 2019 г.). «Влияние циркадного дисбаланса на когнитивные функции у хронических посменных работников» . Научные отчеты . 9 (1): 699. Бибкод : 2019НатСР...9..699С . doi : 10.1038/s41598-018-36762-w . ПМК 6346005 . ПМИД 30679522 .
- ^ Ленг Й., Мусик Э.С., Ху К., Капучио Ф.П., Яффе К. (март 2019 г.). «Связь между циркадными ритмами и нейродегенеративными заболеваниями» . «Ланцет». Неврология . 18 (3): 307–318. дои : 10.1016/S1474-4422(18)30461-7 . ПМК 6426656 . ПМИД 30784558 .
- ^ Ван ZZ, Сунь Z, Чжан МЛ, Сюн К., Чжоу Ф (07.11.2022). «Связь между сменной работой, ночной работой и последующим слабоумием: систематическая оценка и метаанализ» . Границы в неврологии . 13 : 997181. doi : 10.3389/fneur.2022.997181 . ПМЦ 9677942 . ПМИД 36419534 .
- ^ Лесо V, Катурано А, Ветрани I, Явиколи I (январь 2021 г.). «Сменная или ночная смена и риск деменции: систематический обзор». Европейский обзор медицинских и фармакологических наук . 25 (1): 222–232. дои : 10.26355/eurrev_202101_24388 . ПМИД 33506911 .
- ^ Ча А.Е. (2 октября 2017 г.). «Нобелевская премия по физиологии и медицине присуждена трем американцам за открытие «часовых генов» » . Вашингтон Пост . Проверено 2 октября 2017 г.
- ^ «Нобелевская премия по физиологии и медицине 2017 года – пресс-релиз» . Нобелевский фонд. 2 октября 2017 г. Проверено 2 октября 2017 г.
- ^ Бенджакоб О., Авирам Р. (июнь 2018 г.). «Заводная Arc.Ask3.Ru: от широкой перспективы к тематическому исследованию» . Журнал биологических ритмов . 33 (3): 233–244. дои : 10.1177/0748730418768120 . ПМИД 29665713 . S2CID 4933390 .
Дальнейшее чтение [ править ]
- Ашофф Дж, изд. (1965). Циркадные часы . Амстердам: Северная Голландия Пресс.
- Авиви А., Альбрехт У., Остер Х., Джоэл А., Бейлес А., Нево Э. (ноябрь 2001 г.). «Биологические часы в полной темноте: циркадная система Clock/MOP3 слепого подземного землекопа» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (24): 13751–6. Бибкод : 2001PNAS...9813751A . дои : 10.1073/pnas.181484498 . ПМК 61113 . ПМИД 11707566 .
- Авиви А., Остер Х., Джоэл А., Бейлес А., Альбрехт У., Нево Э. (сентябрь 2002 г.). «Циркадные гены слепого подземного млекопитающего II: сохранение и уникальность трех гомологов Периода у слепого подземного землекопа супервида Spalax ehrenbergi» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (18): 11718–23. Бибкод : 2002PNAS...9911718A . дои : 10.1073/pnas.182423299 . ПМК 129335 . ПМИД 12193657 .
- Ли Д., Ма С., Го Д., Ченг Т., Ли Х., Тянь Ю. и др. (октябрь 2016 г.). «Нарушение циркадных ритмов окружающей среды ухудшает неврологические нарушения и подавляет нейрогенез гиппокампа у взрослых крыс после черепно-мозговой травмы» . Клеточная и молекулярная нейробиология . 36 (7): 1045–55. дои : 10.1007/s10571-015-0295-2 . ПМК 4967018 . ПМИД 26886755 .
- Дитти Дж.Л., Уильямс С.Б., Голден СС (2003). «Цианобактериальный циркадный механизм синхронизации». Ежегодный обзор генетики . 37 : 513–43. дои : 10.1146/annurev.genet.37.110801.142716 . ПМИД 14616072 . S2CID 36703896 .
- Данлэп Дж.С., Лорос Дж., ДеКурси П.Дж. (2003). Хронобиология: биологический хронометраж . Сандерленд: Синауэр. ISBN 978-0-87893-149-1 .
- Дворник В, Виноградова О, Нево Е (март 2003 г.). «Происхождение и эволюция генов циркадных часов у прокариот» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (5): 2495–500. Бибкод : 2003PNAS..100.2495D . дои : 10.1073/pnas.0130099100 . ПМК 151369 . ПМИД 12604787 .
- Куккари В.Л., Южный РБ (2006 г.). Знакомство с биологическими ритмами . Нью-Йорк: Спрингер.
- Мартино Т., Араб С., Страуме М., Белшам Д.Д., Тата Н., Кай Ф. и др. (апрель 2004 г.). «Дневные/ночные ритмы в экспрессии генов нормального мышиного сердца». Журнал молекулярной медицины . 82 (4): 256–64. дои : 10.1007/s00109-003-0520-1 . ПМИД 14985853 . S2CID 871822 .
- Рефинетти Р. (2006). Циркадная физиология (2-е изд.). Бока-Ратон: CRC Press.
- Такахаши Дж. С., Зац М. (сентябрь 1982 г.). «Регуляция циркадной ритмичности». Наука . 217 (4565): 1104–11. Бибкод : 1982Sci...217.1104T . дои : 10.1126/science.6287576 . ПМИД 6287576 .
- Томита Дж., Накадзима М., Кондо Т., Ивасаки Х. (январь 2005 г.). «Отсутствует обратная связь транскрипции-трансляции в циркадном ритме фосфорилирования KaiC» . Наука . 307 (5707): 251–4. Бибкод : 2005Sci...307..251T . дои : 10.1126/science.1102540 . ПМИД 15550625 . S2CID 9447128 .
- Мур-Эде MC, Сульцман FM, Фуллер Калифорния (1982). Часы, которые определяют наше время: физиология циркадной системы времени . Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-13581-9 .
Внешние ссылки [ править ]
| Внутренние ритмы |  | |
|---|---|---|
| Внешние циклы | ||
| Поля | ||
| См. также | ||