Развернутый белковый ответ
Эта статья включает список общих ссылок , но в ней отсутствуют достаточные соответствующие встроенные цитаты . ( сентябрь 2012 г. ) |
Реакция развернутого белка ( UPR ) представляет собой клеточную реакцию на стресс, связанную со стрессом эндоплазматической сети (ER). [1] Было обнаружено, что он сохраняется между млекопитающих , видами [2] а также дрожжи [1] [3] и червевые организмы.
UPR активируется в ответ на накопление развернутых или неправильно свернутых белков в просвете эндоплазматической сети. В этом сценарии UPR преследует три цели: сначала восстановить нормальную функцию клетки путем остановки трансляции белков , разрушения неправильно свернутых белков и активации сигнальных путей, которые приводят к увеличению выработки молекулярных шаперонов , участвующих в сворачивании белков . Если эти цели не достигаются в течение определенного периода времени или нарушение затягивается, УПО направлено на апоптоз .
Устойчивая сверхактивация UPR связана с прионовыми заболеваниями, а также с некоторыми другими нейродегенеративными заболеваниями , и ингибирование UPR может стать методом лечения этих заболеваний. [4] Заболевания, поддающиеся ингибированию UPR, включают болезнь Крейцфельдта-Якоба , болезнь Альцгеймера , болезнь Паркинсона и болезнь Хантингтона . [5] [6]
Складывание белков в эндоплазматическом ретикулуме
[ редактировать ]Синтез белка
[ редактировать ]Термин «складывание белка» включает в себя все процессы, участвующие в производстве белка после того, как образующиеся полипептиды синтезируются рибосомами . Белки, предназначенные для секреции или сортировки в другие клеточные органеллы, несут N-концевую сигнальную последовательность, которая будет взаимодействовать с частицей распознавания сигнала (SRP). SRP приведет весь комплекс ( рибосому , РНК , полипептид ) к мембране ЭР. Как только последовательность «стыковывается», белок продолжает трансляцию, при этом полученная цепь подается через полипептидный транслокатор непосредственно в ЭР. Сворачивание белка начинается, как только полипептид попадает в просветную среду, даже если трансляция оставшегося полипептида продолжается.
Сворачивание белка и контроль качества
[ редактировать ]Этапы сворачивания белка включают ряд ферментов и молекулярных шаперонов для координации и регулирования реакций, а также ряд субстратов, необходимых для протекания реакций. Наиболее важными из них, на которые следует обратить внимание, являются N-связанное гликозилирование и образование дисульфидной связи. N-связанное гликозилирование происходит, как только белковая последовательность проходит в ЭР через транслокон , где она гликозилируется с помощью молекулы сахара, образующей ключевой лиганд для молекул лектина кальретикулина (CRT; растворим в просвете ЭР) и кальнексина (CNX; мембраносвязанный). [7] которым благоприятствует сильно окислительная среда ЭР, Белок-дисульфид-изомеразы, способствуют образованию дисульфидных связей, которые придают белку структурную стабильность, позволяющую ему противостоять неблагоприятным условиям, таким как экстремальные значения pH и деградирующие ферменты .
ЭР способен распознавать неправильно свернутые белки, не вызывая нарушения функционирования ЭР. Вышеупомянутая молекула сахара остается средством, с помощью которого клетка контролирует сворачивание белка, поскольку неправильно сворачивающийся белок становится характерным образом лишенным остатков глюкозы, направляя его для идентификации и повторного гликозилирования с помощью фермента UGGT (UDP-глюкоза:гликопротеинглюкозилтрансфераза). [7] Если это не помогает восстановить нормальный процесс сворачивания, открытые гидрофобные остатки неправильно свернутого белка связываются белком 78, регулирующим глюкозу (Grp78), членом семейства белков теплового шока с массой 70 кДа. [8] что предотвращает дальнейший транзит и секрецию белка. [9]
Если обстоятельства продолжают вызывать неправильное сворачивание определенного белка, считается, что этот белок представляет угрозу правильному функционированию ЭР, поскольку они могут агрегировать друг с другом и накапливаться. В таких обстоятельствах белок проходит через деградацию, связанную с эндоплазматическим ретикулумом ( ERAD ). Шаперон EDEM направляет ретротранслокацию неправильно свернутого белка обратно в цитозоль в временных комплексах с PDI и Grp78. [10] Здесь он вступает в путь убиквитин-протеасома, поскольку он помечен множеством молекул убиквитина, направляя его на деградацию цитозольными протеасомами.
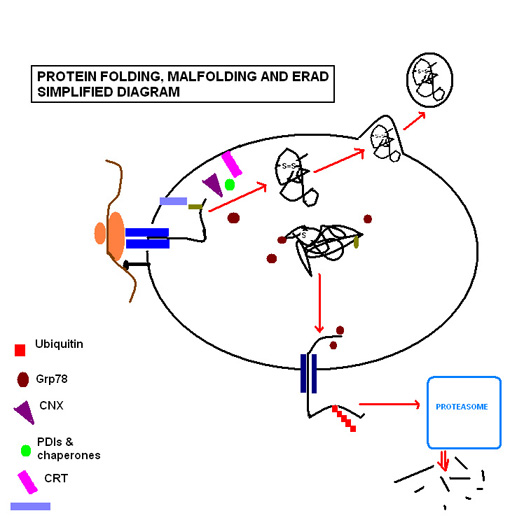
Успешный сворачивание белка требует строго контролируемой среды субстратов, включающих глюкозу, для удовлетворения метаболических энергетических потребностей функционирующих молекулярных шаперонов; кальций, который сохраняется связанным с резидентными молекулярными шаперонами; и окислительно-восстановительные буферы, которые поддерживают окислительную среду, необходимую для образования дисульфидной связи. [11]
Неудачное сворачивание белка может быть вызвано HLA-B27 , нарушающим баланс важных ( IL-10 и TNF ) сигнальных белков. По крайней мере, некоторые нарушения зависят от правильного сворачивания HLA-B27. [12]
Однако, когда обстоятельства вызывают более глобальное нарушение сворачивания белка, которое подавляет механизмы выживания ER, активируется UPR.
Молекулярный механизм
[ редактировать ]Инициация
[ редактировать ]Молекулярный шаперон BiP/Grp78 выполняет ряд функций в ЭР. Он поддерживает специфические трансмембранные рецепторные белки, участвующие в инициации последующей передачи сигналов UPR, в неактивном состоянии путем связывания с их просветными доменами. Чрезмерное количество неправильно свернутых белков или просто сверхэкспрессия белков (например, IgG) [13] требуется больше доступного BiP/Grp78 для связывания с открытыми гидрофобными областями этих белков, и, следовательно, BiP/Grp78 диссоциирует от этих рецепторных участков, чтобы удовлетворить это требование. Диссоциация от внутриклеточных доменов рецепторов позволяет им стать активными. PERK димеризуется с BiP в покоящихся клетках и олигомеризуется в клетках, подвергшихся стрессу ER.
Хотя это традиционно общепринятая модель, ее обоснованность вызывает сомнения. Утверждалось, что генетические и структурные данные, подтверждающие эту модель, просто показывают, что диссоциация BiP просто коррелирует с активацией Ire1 , а не вызывает ее конкретно. [14] Была предложена альтернативная модель, согласно которой развернутые белки взаимодействуют непосредственно с ER-люменальным доменом Ire1, вызывая олигомеризацию и трансаутофосфорилирование. [14] Однако эти модели не являются взаимоисключающими; возможно также, что как прямое взаимодействие Ire1 с развернутыми белками, так и диссоциация BiP от IRE1 способствуют активации пути Ire1.
Функции
[ редактировать ]Начальные этапы активации УПО имеют две ключевые роли:
Ослабление трансляции и остановка клеточного цикла рецептором PERK Это происходит в течение нескольких минут или часов после активации UPR, чтобы предотвратить дальнейшую трансляционную загрузку ER. PERK (протеинкиназа РНК-подобная киназа эндоплазматического ретикулума) активируется путем олигомеризации и аутофосфорилирования свободного просветного домена. Активированный цитозольный домен вызывает ослабление трансляции путем прямого фосфорилирования α-субъединицы регулирующего инициатора аппарата трансляции мРНК, eIF2. [15] Это также приводит к ослаблению трансляции белкового механизма, участвующего в управлении клеточным циклом, что приводит к остановке клеточного цикла в фазе G1. [16] Дефицит PERK может оказывать значительное влияние на физиологические состояния, связанные со стрессом ER .

Увеличение производства белков, участвующих в функциях УПР. Активация UPR также приводит к усилению регуляции белков, участвующих в шаперонировании белков мальфолдинга, сворачивании белков и ERAD, включая дальнейшее производство Grp78. В конечном итоге это увеличивает молекулярные механизмы клетки, с помощью которых она может справиться с неправильно свернутой белковой нагрузкой. Эти рецепторные белки были идентифицированы как:
- Инозитол-требующая киназа 1, [17] чей свободный просветный домен активируется путем гомодимеризации и трансаутофосфорилирования. [18] Активированный домен способен активировать мРНК транскрипционного фактора XBP1 (Xbox-связывающий белок) (эквивалент мРНК дрожжей Hac1 у млекопитающих) путем расщепления и удаления интрона длиной 26 п.о. Активированный фактор транскрипции усиливает «гены стресса» UPR путем прямого связывания с промоторами стрессовых элементов в ядре. [19]
- ATF6 (активирующий фактор транскрипции 6) является основным фактором транскрипции лейциновой молнии. [20] При диссоциации Grp78 весь белок массой 90 кДа перемещается в аппарат Гольджи, где расщепляется протеазами с образованием активного транскрипционного фактора массой 50 кДа. [21] который перемещается в ядро. Он связывается с промоторами стрессовых элементов выше генов, которые активируются в UPR. [22]
Целью этих реакций является устранение накопленной белковой нагрузки, предотвращая при этом любое дальнейшее усиление стресса, чтобы нормальная функция ЭР могла быть восстановлена как можно скорее.
Если путь UPR активируется аномальным образом, например, когда ожирение вызывает хронический стресс ER , и этот путь конститутивно активен, это может привести к нечувствительности к передаче сигналов инсулина и, следовательно, к резистентности к инсулину. Лица, страдающие ожирением, предъявляют повышенные требования к секреторной и синтезирующей системам своих клеток. Это активирует передачу сигналов клеточного стресса и воспалительные пути из-за аномальных условий, нарушающих гомеостаз ER.
Последующим эффектом стресса ER является значительное снижение инсулин-стимулированного фосфорилирования тирозиновых остатков субстрата инсулинового рецептора (IRS-1), который является субстратом инсулин-тирозинкиназы (инсулинового рецептора). N-концевая киназа C-Jun (JNK) также активируется на высоких уровнях с помощью IRE-1α, который сам фосфорилируется и активируется в присутствии стресса ER. Впоследствии JNK фосфорилирует остатки серина IRS-1 и, таким образом, ингибирует передачу сигналов инсулинового рецептора. IRE-1α также рекрутирует фактор 2, связанный с рецептором фактора некроза опухоли ( TRAF2 ). Этот киназный каскад, который зависит от IRE-1α и JNK, опосредует индуцированное стрессом ER ингибирование действия инсулина. [23]
Ожирение обеспечивает хронические клеточные стимулы для пути UPR в результате стрессов и перенапряжений, воздействующих на ER, и, не позволяя восстановить нормальную клеточную реакцию на передачу сигналов гормона инсулина, у человека с высокой вероятностью развивается диабет 2 типа.
Скелетные мышцы чувствительны к физиологическому стрессу, поскольку физические упражнения могут нарушить гомеостаз ЭР. Это приводит к тому, что экспрессия шаперонов ER индуцируется UPR в ответ на стресс ER , вызванный физической нагрузкой . Мышечное сокращение во время тренировки вызывает высвобождение кальция из саркоплазматического ретикулума (СР), специализированной сети ЭР в скелетных мышцах. Затем этот кальций взаимодействует с кальциневрином и кальций/кальмодулин-зависимыми киназами, которые, в свою очередь, активируют факторы транскрипции. Эти транскрипционные факторы затем начинают изменять экспрессию мышечных генов, регулируемых физическими упражнениями. PGC-1alpha , коактиватор транскрипции, является ключевым фактором транскрипции, участвующим в опосредовании UPR тканеспецифичным образом в скелетных мышцах путем коактивации ATF6alpha. Таким образом, PGC-1альфа экспрессируется в мышцах после острых и длительных тренировок. Функция этого транскрипционного фактора заключается в увеличении количества и функции митохондрий, а также в индуцировании переключения скелетных волокон на медленные окислительные мышечные волокна, поскольку они устойчивы к утомлению. Таким образом, этот путь UPR опосредует изменения в мышцах, прошедших тренировку на выносливость, делая их более устойчивыми к утомлению и защищая их от будущего стресса. [24]
Инициация апоптоза
[ редактировать ]В условиях длительного стресса цель UPR меняется с цели, способствующей выживанию клеток, на цель, которая направляет клетку на путь апоптоза. Было идентифицировано, что белки, расположенные ниже всех трех путей рецептора UPR, играют проапоптотическую роль. Однако момент, в котором активируется «апоптотический переключатель», еще не определен, но логично предположить, что это должно происходить за пределами определенного периода времени, в течение которого не было достигнуто разрешение стресса. Двумя основными задействованными рецепторами UPR являются Ire1 и PERK.
Связываясь с белком TRAF2, Ire1 активирует сигнальный путь JNK, [25] Считается, что в этот момент человеческая прокаспаза 4 вызывает апоптоз, активируя нижестоящие каспазы.
Хотя известно, что PERK вызывает блокировку трансляции, некоторые гены могут обходить этот блок. Важным примером является то, что проапоптотический белок CHOP ( белок, гомологичный CCAAT/-энхансер-связывающему белку ) активируется ниже транскрипционного фактора bZIP ATF4 (активирующего транскрипционный фактор 4) и уникальным образом реагирует на стресс ER. [26] CHOP вызывает подавление антиапоптотического митохондриального белка Bcl-2, [27] благоприятствуя проапоптотическому стимулированию митохондрий белками, которые вызывают повреждение митохондрий, высвобождение цитохрома С и активацию каспазы 3.
Болезни
Заболевания, поддающиеся ингибированию UPR, включают болезнь Крейцфельдта-Якоба , болезнь Альцгеймера , болезнь Паркинсона и болезнь Хантингтона . [28]
Сообщалось, что стресс эндоплазматической сети играет важную роль в индукции и прогрессировании неалкогольной жировой болезни печени (НАЖБП). У крыс, получавших диету с высоким содержанием жиров, наблюдалось увеличение маркеров стресса ER CHOP , XBP1 и GRP78 . Известно, что стресс ЭР активирует липогенез в печени de novo, ингибирует секрецию ЛПОНП, способствует резистентности к инсулину и воспалительному процессу, а также способствует апоптозу клеток. Таким образом, это увеличивает уровень накопления жира и усугубляет НАЖБП до более серьезного состояния печени. [29] экстракт Zingiber officinale (имбирь) и жирные кислоты омега-3 уменьшают стресс эндоплазматического ретикулума на модели крыс с неалкогольным ожирением печени. Сообщалось, что [29]
Как указано выше, UPR также может быть активирован в качестве компенсаторного механизма при болезненных состояниях. Например, UPR усиливается при наследственной форме дилатационной кардиомиопатии из-за мутации в гене, кодирующем белок фосфоламбан. [30] Дальнейшая активация оказалась терапевтической на модели плюрипотентных стволовых клеток человека при мутантной дилатационной кардиомиопатии PLN. [30]
Химические индукторы
[ редактировать ]- Брефельдин А является очень распространенным индуктором реакции развернутого белка или реакции на стресс эндоплазматического ретикулума (стресс ER) .
- тапсигаргин [31] ведет в отделение скорой помощи 2+ истощение из-за ингибирования сарко/эндоплазматического ретикулума Ca 2+ -АТФаза (СЕРДЦЕ).
- А23187 [31] усиливает экспрессию стрессовых белков ER
- 2-дезоксиглюкоза [31]
- дитиотреитол [31] уменьшает дисульфидные мостики белков. Денатурированные белки накапливались внутри ЭР.
- фенретинид и бортезомиб (Велкейд), каждый из которых действует посредством разных клеточных механизмов, вызывают стресс ЭР, что приводит к апоптозу в клетках меланомы.
- туникомицин ингибирует N-связанное гликозилирование.
Биологические индукторы
[ редактировать ]- Вирус денге вызывает PERK-зависимый стресс ER как часть вызванной вирусом реакции в инфицированных клетках, способствующей репликации. [32]
- Вирус гриппа требует белка эндоплазматического ретикулума 57-кДа (ERp57) для репликации и индукции апоптоза в инфицированных клетках. [33]
См. также
[ редактировать ]- Реакция на стресс эндоплазматического ретикулума (стресс ER)
- Реакция митохондриального развернутого белка
- агрессивный
- Ингибиторы PERK
Ссылки
[ редактировать ]- ^ Jump up to: а б Hetz C, Papa FR (январь 2018 г.). «Развернутый белковый ответ и контроль клеточной судьбы» . Молекулярная клетка . 69 (2): 169–181. doi : 10.1016/j.molcel.2017.06.017 . ПМИД 29107536 .
- ^ «Краткий доклад Питера Уолтера: развитие УПО» . Архивировано из оригинала 12 июля 2017 г. Проверено 24 октября 2013 г.
- ^ Каннан М., Сивапракасам К., Принц В.А., Начиаппан В. (декабрь 2016 г.). «Стресс эндоплазматической сети влияет на транспорт фосфатидилэтаноламина из митохондрий в эндоплазматический ретикулум у S.cerevisiae» . Biochimica et Biophysical Acta (BBA) - Молекулярная и клеточная биология липидов . 1861 (12 ч. А): 1959–1967. дои : 10.1016/j.bbalip.2016.09.015 . ПМК 6322925 . ПМИД 27678054 .
- ^ Морено Дж.А., Холлидей М., Моллой С., Рэдфорд Х., Верити Н., Экстен Дж.М. и др. (октябрь 2013 г.). «Пероральное лечение, направленное на развернутый белковый ответ, предотвращает нейродегенерацию и клинические заболевания у мышей, инфицированных прионами». Наука трансляционной медицины . 5 (206): 206ра138. doi : 10.1126/scitranslmed.3006767 . ПМИД 24107777 . S2CID 25570626 .
- ^ Шепер В., Хуземанс Дж. Дж. (сентябрь 2015 г.). «Развернутый белковый ответ при нейродегенеративных заболеваниях: невропатологический взгляд» . Акта Нейропатологика . 130 (3): 315–31. дои : 10.1007/s00401-015-1462-8 . ПМК 4541706 . ПМИД 26210990 .
- ^ Лаккараджу А.К., Фронцек К., Лемес Э., Херрманн У., Лоса М., Марпаквар Р., Агуцци А. (сентябрь 2021 г.). «Потеря PIKfyve приводит к губчатой дегенерации прионных заболеваний» . ЭМБО Молекулярная медицина . 13 (9): e14714. дои : 10.15252/emmm.202114714 . ПМЦ 8518562 . ПМИД 34291577 .
- ^ Jump up to: а б Блонд-Элгинди С., Квирла С.Э., Дауэр В.Дж., Липшуц Р.Дж., Спранг С.Р., Сэмбрук Дж.Ф., Гетинг М.Дж. (ноябрь 1993 г.). «Аффинное панорамирование библиотеки пептидов, представленных на бактериофагах, выявляет специфичность связывания BiP» . Клетка . 75 (4): 717–28. дои : 10.1016/0092-8674(93)90492-9 . ПМИД 7902213 .
- ^ Брюэр Дж. В., Диль Дж. А. (ноябрь 2000 г.). «PERK опосредует выход из клеточного цикла во время ответа развернутого белка у млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (23): 12625–30. Бибкод : 2000PNAS...9712625B . дои : 10.1073/pnas.220247197 . ЧВК 18814 . ПМИД 11035797 .
- ^ Чен X, Шен Дж, Прайвес Р. (апрель 2002 г.). «Люминальный домен ATF6 воспринимает стресс эндоплазматического ретикулума (ЭР) и вызывает транслокацию ATF6 из ЭР в Гольджи» . Журнал биологической химии . 277 (15): 13045–52. дои : 10.1074/jbc.M110636200 . ПМИД 11821395 .
- ^ Кокс Дж.С., Шаму CE, Уолтер П. (июнь 1993 г.). «Для индукции транскрипции генов, кодирующих резидентные белки эндоплазматического ретикулума, требуется трансмембранная протеинкиназа». Клетка . 73 (6): 1197–206. дои : 10.1016/0092-8674(93)90648-А . ПМИД 8513503 . S2CID 16065404 .
- ^ Хаммонд С., Браакман И., Хелениус А. (февраль 1994 г.). «Роль распознавания N-связанных олигосахаридов, обрезки глюкозы и кальнексина в сворачивании гликопротеинов и контроле качества» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (3): 913–7. Бибкод : 1994PNAS...91..913H . дои : 10.1073/pnas.91.3.913 . ПМК 521423 . ПМИД 8302866 .
- ^ LL Маркус Пенттинен (10 января 2004 г.). HLA-B27 связан со снижением устойчивости бактерий сальмонеллы (на финском языке). Библиотека Университета Турку: Анн. унив. Туркуенсис Д 619. ISBN 951-29-2742-Х . Архивировано из оригинала 6 января 2013 года . Проверено 9 октября 2012 г.
- ^ Кобер Л., Зехе С., Боде Дж. (октябрь 2012 г.). «Разработка новой системы селекции на основе стресса ER для выделения высокопродуктивных клонов». Биотехнология и биоинженерия . 109 (10): 2599–611. дои : 10.1002/бит.24527 . ПМИД 22510960 . S2CID 25858120 .
- ^ Jump up to: а б Берналес С., Папа Ф.Р., Уолтер П. (2006). «Внутриклеточная передача сигналов посредством ответа развернутого белка». Ежегодный обзор клеточной биологии и биологии развития . 22 : 487–508. doi : 10.1146/annurev.cellbio.21.122303.120200 . ПМИД 16822172 .
- ^ Хардинг Х.П. , Чжан Ю., Рон Д. (январь 1999 г.). «Трансляция и сворачивание белка связаны с киназой, резидентной эндоплазматической сети». Природа . 397 (6716): 271–4. Бибкод : 1999Natur.397..271H . дои : 10.1038/16729 . ПМИД 9930704 . S2CID 4416662 .
- ^ Ли А.Х., Ивакоши Н.Н., Андерсон К.К., Глимчер Л.Х. (август 2003 г.). «Ингибиторы протеасом нарушают развернутый белковый ответ в клетках миеломы» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (17): 9946–51. Бибкод : 2003PNAS..100.9946L . дои : 10.1073/pnas.1334037100 . ЧВК 187896 . ПМИД 12902539 .
- ^ Ли А.С. (январь 1987 г.). «Координированная регуляция набора генов с помощью ионофоров глюкозы и кальция в клетках млекопитающих». Тенденции биохимических наук . 12 :20–3. дои : 10.1016/0968-0004(87)90011-9 .
- ^ Мачамер С.Э., Домс Р.В., Боле Д.Г., Хелениус А., Роуз Дж.К. (апрель 1990 г.). «Белок, связывающий тяжелые цепи, распознает формы G-белка вируса везикулярного стоматита с неполными дисульфидными связями» . Журнал биологической химии . 265 (12): 6879–83. дои : 10.1016/S0021-9258(19)39231-2 . ПМИД 2157712 .
- ^ Стерба О (1975). «Пренатальный рост крота Talpa europaea Linn., 1758». Фолиа Морфологическая . 23 (3): 282–5. ПМИД 1158311 .
- ^ Молинари М., Галли С., Пиккалуга В., Пирен М., Паганетти П. (июль 2002 г.). «Последовательная помощь молекулярных шаперонов и временное образование ковалентных комплексов во время деградации белка из ЭР» . Журнал клеточной биологии . 158 (2): 247–57. дои : 10.1083/jcb.200204122 . ПМК 2173128 . ПМИД 12119363 .
- ^ Мори К., Огава Н., Кавахара Т., Янаги Х., Юра Т. (апрель 2000 г.). «Опосредованная сплайсингом мРНК С-концевая замена транскрипционного фактора Hac1p необходима для эффективной активации развернутого белкового ответа» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (9): 4660–5. Бибкод : 2000PNAS...97.4660M . дои : 10.1073/pnas.050010197 . ЧВК 18289 . ПМИД 10781071 .
- ^ Урано Ф., Ван Х., Бертолотти А., Чжан Ю., Чунг П., Хардинг Х.П., Рон Д. (январь 2000 г.). «Связь стресса в ЭР с активацией протеинкиназ JNK трансмембранной протеинкиназой IRE1». Наука . 287 (5453): 664–6. Бибкод : 2000Sci...287..664U . дои : 10.1126/science.287.5453.664 . ПМИД 10650002 .
- ^ Озджан У, Цао К, Йилмаз Э, Ли А.Х., Ивакоши Н.Н., Озделен Э. и др. (октябрь 2004 г.). «Стресс эндоплазматической сети связывает ожирение, действие инсулина и диабет 2 типа». Наука . 306 (5695): 457–61. Бибкод : 2004Sci...306..457O . дои : 10.1126/science.1103160 . ПМИД 15486293 . S2CID 22517395 .
- ^ Ву Дж., Руас Дж.Л., Эсталл Дж.Л., Расбах К.А., Чой Дж.Х., Йе Л. и др. (февраль 2011 г.). «Развернутый белковый ответ опосредует адаптацию скелетных мышц к физическим упражнениям через комплекс PGC-1α/ATF6α» . Клеточный метаболизм . 13 (2): 160–9. дои : 10.1016/j.cmet.2011.01.003 . ПМК 3057411 . ПМИД 21284983 .
- ^ Ван XZ, Лоусон Б., Брюэр Дж.В., Зинзнер Х., Санджай А., Ми Л.Дж., Бурштейн Р., Крейбих Г., Хендершот Л.М., Рон Д. (август 1996 г.). «Сигналы от напряженного эндоплазматического ретикулума индуцируют гомологичный C/EBP белок (CHOP/GADD153)» . Молекулярная и клеточная биология . 16 (8): 4273–80. дои : 10.1128/mcb.16.8.4273 . ПМК 231426 . PMID 8754828 .
- ^ Велихинда А.А., Кауфман Р.Дж. (июль 1996 г.). «Путь развернутого белкового ответа у Saccharomyces cerevisiae. Для активации киназы необходимы олигомеризация и трансфосфорилирование Ire1p (Ern1p)» . Журнал биологической химии . 271 (30): 18181–7. дои : 10.1074/jbc.271.30.18181 . ПМИД 8663458 .
- ^ Ёсида Х., Хейз К., Янаги Х., Юра Т., Мори К. (декабрь 1998 г.). «Идентификация цис-действующего элемента ответа на стресс эндоплазматического ретикулума, ответственного за индукцию транскрипции белков, регулируемых глюкозой у млекопитающих. Участие основных факторов транскрипции лейциновой молнии» . Журнал биологической химии . 273 (50): 33741–9. дои : 10.1074/jbc.273.50.33741 . ПМИД 9837962 .
- ^ Новости здоровья BBC (10 октября 2013 г.). «Прорыв в борьбе с болезнью Альцгеймера был назван «поворотным моментом» » . Британская радиовещательная компания . Проверено 10 октября 2013 г.
- ^ Jump up to: а б Кандейл, Мохамед А.; Хашем, Рим М.; Махмуд, Мохамед О.; Хетта, Мона Х.; Тохами, Мохамед А. (2019). «Экстракт Zingiber officinale и жирные кислоты омега-3 уменьшают стресс эндоплазматического ретикулума на модели крыс с неалкогольным ожирением печени» . Журнал пищевой биохимии . 43 (12): e13076. дои : 10.1111/jfbc.13076 . hdl : 2027.42/152724 . ISSN 1745-4514 . ПМИД 31608477 . S2CID 204544806 .
- ^ Jump up to: а б Фейен, Дрис AM; Переа-Гил, Исаак; Маас, Рене Г.К.; Харакалова, Магдалена; Гавидия, Александра А.; Артур Адам, Дженнифер; У, Тин-Сюань; Винк, Ариан; Пей, Цзяи; Вадгама, Нирмал; Суурмейер, Альберт Дж. (3 августа 2021 г.). «Развернутый белковый ответ как компенсаторный механизм и потенциальная терапевтическая мишень при кардиомиопатии PLN R14del» . Тираж . 144 (5): 382–392. дои : 10.1161/тираж.120.049844 . ISSN 1524-4539 . ПМЦ 8667423 . ПМИД 33928785 .
- ^ Jump up to: а б с д «Китамура, М» . Архивировано из оригинала 10 февраля 2012 г. Проверено 6 февраля 2008 г.
- ^ Датан Э., Рой С.Г., Жермен Г., Зали Н., Маклин Дж.Э., Гольшан Г. и др. (март 2016 г.). «Аутофагия, вызванная денге, репликация вируса и защита от гибели клеток требуют активации пути стресса ER (PERK)» . Смерть клеток и болезни . 7 (е2127): е2127. дои : 10.1038/cddis.2015.409 . ПМЦ 4823927 . ПМИД 26938301 .
- ^ Роберсон Э.К., Талли Дж.Э., Гуала А.С., Рейсс Дж.Н., Годберн К.Е., Поциаск Д.А. и др. (май 2012 г.). «Грипп вызывает стресс эндоплазматического ретикулума, каспаза-12-зависимый апоптоз и опосредованное N-концевой киназой c-Jun высвобождение трансформирующего фактора роста-β в эпителиальных клетках легких» . Американский журнал респираторной клеточной и молекулярной биологии . 46 (5): 573–81. дои : 10.1165/rcmb.2010-0460OC . ПМК 3359902 . ПМИД 21799120 .