дрозофилы Циркадный ритм

Циркадный ритм дрозофилы — это суточный 24-часовой цикл отдыха и активности у плодовых мушек рода Drosophila . Биологический процесс был открыт и лучше всего изучен у вида Drosophila melanogaster . Многие виды поведения находятся под циркадным контролем, включая экклозию, двигательную активность, питание и спаривание. Двигательная активность максимальна на рассвете и в сумерках, эклозия – на рассвете. [ 1 ]
Биологические ритмы впервые были изучены у Drosophila pseudoobscura . Циркадный ритм дрозофилы проложил путь к пониманию циркадного поведения и заболеваний, связанных с условиями сна и бодрствования, у других животных, включая человека. Это потому, что циркадные часы фундаментально схожи. [ 2 ] Циркадный ритм дрозофилы был открыт в 1935 году немецкими зоологами Гансом Кальмусом и Эрвином Бюннингом . Американский биолог Колин С. Питтендри в 1954 году провел важный эксперимент, который установил, что циркадный ритм управляется биологическими часами. Генетика была впервые понята в 1971 году, когда Сеймур Бензер и Рональд Дж. Конопка сообщили, что мутация в определенных генах изменяет или останавливает циркадное поведение. Они обнаружили ген периода ( per ), мутации которого изменяют циркадный ритм. Это был первый известный ген, контролирующий поведение. Спустя десятилетие Конопка, Джеффри С. Холл , Майкл Росбаш и Майкл В. Янг открыли новые гены, включая timeless ( tim ), Clock ( Clk ), цикл ( cyc ), крик . Эти гены и белки-продукты играют ключевую роль в циркадных часах. Исследование, проведенное в лаборатории Бензера, описано книге «Время, любовь, память» в Джонатаном Вайнером .
За свой вклад Холл, Росбаш и Янг получили Нобелевскую премию по физиологии и медицине в 2017 году. [ 3 ]
История
[ редактировать ]В процессе выхода взрослой мухи из куколки дрозофила проявляет регулярную двигательную активность (за счет вибрации), которая происходит в течение 8-10-часовых интервалов, начиная непосредственно перед рассветом. Существование этого циркадного ритма было независимо обнаружено у D. melanogaster в 1935 году двумя немецкими зоологами, Гансом Кальмусом из Зоологического института Немецкого университета в Праге (ныне Карлов университет ) и Эрвином Бюннингом из Ботанического института Йенского университета. . [ 4 ] [ 5 ] В 1938 году Калмус обнаружил, что за циркадную активность отвечает область мозга. [ 6 ] Кальмус и Бюннинг считали, что главным фактором была температура. Но вскоре выяснилось, что даже при разной температуре циркадный ритм может оставаться неизменным. [ 7 ] В 1954 году Колин С. Питтендри из Принстонского университета обнаружил важность условий света и темноты для D. pseudoobscura . Он продемонстрировал, что ритм эклозии задерживался, но не прекращался при снижении температуры. Он пришел к выводу, что температура влияет только на час пик ритма и не является основным фактором. [ 8 ] Тогда было известно, что циркадный ритм контролируется биологическими часами. Но природа часов тогда была загадкой. [ 5 ]
Спустя почти два десятилетия существование циркадных часов было обнаружено Сеймуром Бензером и его студентом Рональдом Дж. Конопкой в Калифорнийском технологическом институте . Они обнаружили, что мутации в Х-хромосоме D. melanogaster могут вызывать аномальную циркадную активность. Когда определенная часть хромосомы отсутствовала (инактивировалась), циркадный ритм отсутствовал; в одной мутации (называемой per С , «S» сокращенно или укороченно) ритм укорочен до ~19 часов; тогда как в другой мутации ( по л , «L» — длинный или удлиненный) ритм удлинялся до ~29 часов, в отличие от нормального 24-часового ритма. Они опубликовали свое открытие в 1971 году. [ 9 ] Они назвали местоположение гена ( локус ) периодом ( для краткости), поскольку он контролирует период ритма. В противоположность этому были и другие ученые, которые утверждали, что гены не могут контролировать такое сложное поведение, как циркадная активность. [ 10 ]
Еще одно циркадное поведение дрозофилы — ухаживание самца и самки во время спаривания. Ухаживание включает в себя песню, сопровождаемую ритуальным двигательным танцем у самцов. Основная полетная активность обычно происходит утром, а еще один пик приходится на перед закатом. Брачная песня создается вибрацией крыльев самца и состоит из тональных импульсов, производимых с интервалом примерно 34 мс у D. melanogaster (48 мс у D. simulans ). В 1980 году Джеффри К. Холл и его студент Хараламбос П. Кириаку из Университета Брандейса в Уолтеме обнаружили, что активность ухаживания также контролируется каждым геном. [ 11 ] В 1984 году Конопка, Холл, Майкл Рошбаш и их команда сообщили в двух статьях, что каждый локус является центром циркадного ритма и что потеря пера останавливает циркадную активность. [ 12 ] [ 13 ] В то же время команда Майкла В. Янга из Университета Рокфеллера сообщила об аналогичных эффектах per , а также о том, что этот ген покрывает интервал длиной 7,1 килобаз (т.п.н.) на Х-хромосоме и кодирует поли(А)+ РНК размером 4,5 т.п.н. [ 14 ] [ 15 ] В 1986 году они секвенировали весь фрагмент ДНК и обнаружили, что ген кодирует РНК размером 4,5 т.п.н., которая продуцирует белок протеогликан , состоящий из 1127 аминокислот. [ 16 ] В то же время команда Рошбаша показала, что белок PER отсутствует у мутанта per . [ 17 ] В 1994 году Янг и его команда обнаружили ген timeless ( tim ), влияющий на активность человека . [ 18 ] В 1998 году они открыли даблтаймы ( dbt ), которые регулируют количество белка PER. [ 19 ]
В 1990 году Конопка и Росбаш идентифицировали новый ген под названием Clock ( Clk ), жизненно важный для циркадного периода. [ 20 ] В 1998 году они обнаружили новый генный цикл ( cyc ), который действует совместно с Clk . [ 21 ] В конце 1998 года команда Холла и Рошбаша обнаружила Cryb , ген чувствительности к синему свету. [ 22 ] Одновременно они определили белок CRY как основную светочувствительную ( фоторецепторную ) систему. Активность плача находится под циркадной регуляцией и находится под влиянием других генов, таких как per , tim , clk и cyc . [ 23 ] Генный продукт CRY представляет собой основной фоторецепторный белок, принадлежащий к классу флавопротеинов , называемых криптохромами . Они также присутствуют в бактериях и растениях. [ 24 ] В 1998 году Холл и Джэ Х. Парк выделили ген, кодирующий нейропептид, названный фактором диспергирования пигмента (PDF), исходя из одной из ролей, которую он играет у ракообразных . [ 25 ] В 1999 году они обнаружили, что pdf экспрессируется вентральными кластерами латеральных нейронов (LNv), что указывает на то, что белок PDF является основным циркадным нейротрансмиттером и что нейроны LNv являются основными циркадными водителями ритма. [ 26 ] В 2001 году Янг и его команда продемонстрировали, что ортолог лохматой киназы-3 гликогенсинтазы (GSK-3) (SGG) представляет собой фермент, который регулирует созревание и накопление ТИМ ранней ночью, вызывая фосфорилирование . [ 27 ]
Холл, Росбаш и Янг разделили Нобелевскую премию по физиологии и медицине 2017 года «за открытие молекулярных механизмов, контролирующих циркадный ритм». [ 3 ]
Механизм
[ редактировать ]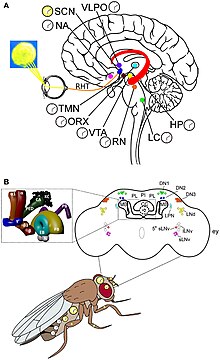
У дрозофилы есть две отдельные группы циркадных часов: часовые нейроны и часовые гены. Они действуют согласованно, создавая 24-часовой цикл отдыха и активности. Свет является источником активации часов. , сложные глаза глазки и Непосредственными наружными фоторецепторными органами являются глазки Гофбауэра-Бюхнера (петельки НВ). Но циркадные часы могут работать и в постоянной темноте. [ 28 ] Тем не менее, фоторецепторы необходимы для измерения продолжительности дня и обнаружения лунного света. Сложные глаза важны для того, чтобы отличать долгие дни от постоянного света. Для обычных маскирующих эффектов света, таких как стимулирование активности светом и торможение темнотой. [ 29 ] Есть два отчетливых пика активности, называемых пиком М (утренний), который происходит на рассвете, и пиком Е (вечерний), который приходится на сумерки. Они следят за разной продолжительностью дня в разные сезоны года. [ 30 ] Светочувствительные белки глаза, называемые родопсинами (родопсин 1 и 6), играют решающую роль в активации колебаний M и E. [ 31 ] около 100 000 нейронов При обнаружении окружающего света около 150 нейронов (в мозге дрозофилы ) в мозге регулируют циркадный ритм. [ 32 ] Часовые нейроны расположены в отдельных кластерах в центральном мозге. Наиболее изученными часовыми нейронами являются большие и малые латеральные вентральные нейроны (l-LNvs и s-LNvs) в основании зрительной доли . Эти нейроны вырабатывают фактор диспергирования пигмента (PDF), нейропептид, который действует как циркадный нейромодулятор между различными часовыми нейронами. [ 33 ]

Циркадный ритм дрозофилы отслеживает время посредством ежедневных колебаний белков, связанных с часами, которые взаимодействуют в петле обратной связи транскрипции-трансляции. Основной механизм синхронизации состоит из двух взаимозависимых контуров обратной связи, а именно контура PER/TIM и контура CLK/CYC. [ 34 ] Петля CLK/CYC возникает в течение дня, в течение которого вырабатываются как часовой белок, так и циклический белок. Гетеродимеры CLK/CYC действуют как факторы транскрипции и связываются вместе, инициируя транскрипцию генов per и tim , путем связывания с промоторным элементом, называемым E-боксом, около полудня. ДНК транскрибируется с образованием мРНК PER и мРНК TIM. Белки PER и TIM синтезируются в цитоплазме и демонстрируют плавное увеличение уровня в течение дня. Уровни РНК в них достигают максимума рано вечером, а уровни белка достигают пика ближе к рассвету. [ 32 ] Но уровень белка в них поддерживается на постоянно низком уровне до наступления сумерек, поскольку дневной свет также активирует ген двойного времени ( dbt ). Белок DBT индуцирует посттрансляционные модификации , то есть фосфорилирование и оборот мономерных белков PER. Поскольку PER транслируется в цитоплазме, он активно фосфорилируется с помощью DBT ( казеинкиназа 1 ε) и казеинкиназа 2 (синтезируется And и Tik ), что является прелюдией к преждевременной деградации. Фактическая деградация происходит по пути убиквитин-протеасома и осуществляется убиквитинлигазой, называемой Slimb (сверхкомплектные конечности). [ 35 ] [ 36 ] При этом сам ТИМ фосфорилируется лохматыми, активность которых снижается после захода солнца. ДБТ постепенно исчезает, а удаление ДБТ способствует стабилизации молекул PER за счет физической ассоциации с ТИМ. Следовательно, максимальное производство PER и TIM происходит в сумерках. В то же время CLK/CYC также напрямую активирует vri и Pdp1 (ген белка 1 домена PAR). VRI накапливается первым, на 3-6 часов раньше, и начинает репрессировать Clk ; но входящий PDP1 создает конкуренцию, активируя Clk . Димеры PER/TIM накапливаются ранней ночью, через несколько часов организованно перемещаются в ядро и связываются с димерами CLK/CYC. Связанный PER полностью останавливает транскрипционную активность CLK и CYC. [ 37 ]
Ранним утром появление света приводит к разрушению белков PER и TIM в сети активации и репрессии транскрипции. Во-первых, свет активирует ген крика в часовых нейронах. Хотя CRY вырабатывается глубоко внутри мозга, он чувствителен к ультрафиолетовому и синему свету и поэтому легко сигнализирует клеткам мозга о появлении света. Он необратимо и напрямую связывается с TIM, вызывая его разрушение посредством протеосомно-зависимой деградации, опосредованной убиквитином. CRY Домен гомологии фотолиазы используется для обнаружения света и фототрансдукции , тогда как карбоксил-концевой домен регулирует стабильность CRY, взаимодействие CRY-TIM и циркадную фоточувствительность. [ 38 ] Убиквитинированию и последующей деградации способствует другой белок JET. [ 39 ] Таким образом, димер PER/TIM диссоциирует, и несвязанный PER становится нестабильным. PER подвергается прогрессивному фосфорилированию и, в конечном итоге, деградации. Отсутствие PER и TIM позволяет активировать гены clk и cyc . Таким образом, часы сбрасываются, чтобы начать следующий циркадный цикл. [ 10 ]
Ссылки
[ редактировать ]- ^ Дубовой, Кристина; Сегал, Амита (2017). «Циркадные ритмы и сон» . Генетика . 205 (4): 1373–1397. дои : 10.1534/genetics.115.185157 . ПМЦ 5378101 . ПМИД 28360128 .
- ^ Розато, Эцио; Таубер, Эран; Кириаку, Хараламбос П. (2006). «Молекулярная генетика циркадных часов плодовых мушек» . Европейский журнал генетики человека . 14 (6): 729–738. дои : 10.1038/sj.ejhg.5201547 . ПМИД 16721409 . S2CID 12775655 .
- ^ Перейти обратно: а б Нобелевский фонд (2017). «Нобелевская премия по физиологии и медицине 2017» . www.nobelprize.org . Нобель Медиа АБ . Проверено 28 декабря 2017 г.
- ^ Брюс, Виктор Г.; Питтендри, Колин С. (1957). «Эндогенные ритмы у насекомых и микроорганизмов». Американский натуралист . 91 (858): 179–195. дои : 10.1086/281977 . S2CID 83886607 .
- ^ Перейти обратно: а б Питтендри, CS (1993). «Временная организация: размышления дарвинистского часовщика». Ежегодный обзор физиологии . 55 (1): 17–54. дои : 10.1146/annurev.ph.55.030193.000313 . ПМИД 8466172 .
- ^ Калмус, Х. (1938). «Расположение принимающего органа в период вылупления дрозофилы » . Журнал сравнительной физиологии . 26 (3): 362–365. дои : 10.1007/BF00338939 . S2CID 28171026 .
- ^ Уэлш, Дж. Х. (1938). «Суточные ритмы». Ежеквартальный обзор биологии . 13 (2): 123–139. дои : 10.1086/394554 . S2CID 222425940 .
- ^ Питтендри, CS (1954). «О температурной независимости в системе часов, контролирующей время вылета дрозофилы » . Труды Национальной академии наук Соединенных Штатов Америки . 40 (10): 1018–1029. Бибкод : 1954PNAS...40.1018P . дои : 10.1073/pnas.40.10.1018 . ПМК 534216 . ПМИД 16589583 .
- ^ Конопка, Р.Дж.; Бензер, С. (1971). «Часовые мутанты Drosophila melanogaster » . Труды Национальной академии наук Соединенных Штатов Америки . 68 (9): 2112–2116. Бибкод : 1971ПНАС...68.2112К . дои : 10.1073/pnas.68.9.2112 . ПМЦ 389363 . ПМИД 5002428 .
- ^ Перейти обратно: а б Лалчхандама, К. (2017). «Путь к Нобелевской премии по физиологии и медицине 2017 года» . Научное видение . 3 (Приложение): 1–13.
- ^ Кириаку, КП; Холл, Дж. К. (1980). «Мутации циркадного ритма у Drosophila melanogaster влияют на кратковременные колебания брачной песни самца» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (11): 6729–6733. Бибкод : 1980PNAS...77.6729K . дои : 10.1073/pnas.77.11.6729 . ПМК 350362 . ПМИД 6779281 .
- ^ Редди, П.; Церинг, Вашингтон; Уилер, округ Колумбия; Пирротта, В.; Хэдфилд, К.; Холл, Дж. К.; Росбаш, М. (1984). «Молекулярный анализ локуса периода у Drosophila melanogaster и идентификация транскрипта, участвующего в биологических ритмах». Клетка . 38 (3): 701–710. дои : 10.1016/0092-8674(84)90265-4 . ПМИД 6435882 . S2CID 316424 .
- ^ Церинг, Вашингтон; Уилер, округ Колумбия; Редди, П.; Конопка, Р.Дж.; Кириаку, КП; Росбаш, М.; Холл, Дж. К. (1984). «Трансформация P-элемента с помощью ДНК локуса периода восстанавливает ритмичность мутантной аритмичной Drosophila melanogaster » . Клетка . 39 (2, часть 1): 369–376. дои : 10.1016/0092-8674(84)90015-1 . ПМИД 6094014 . S2CID 9762751 .
- ^ Барджиелло, штат Калифорния; Джексон, Франция; Янг, М.В. (1984). «Восстановление циркадных поведенческих ритмов путем переноса генов у дрозофилы ». Природа . 312 (5996): 752–754. Бибкод : 1984Natur.312..752B . дои : 10.1038/312752a0 . ПМИД 6440029 . S2CID 4259316 .
- ^ Барджиелло, штат Калифорния; Янг, М.В. (1984). «Молекулярная генетика биологических часов у дрозофилы » . Труды Национальной академии наук Соединенных Штатов Америки . 81 (7): 2142–2146. дои : 10.1038/312752a0 . ПМЦ 345453 . ПМИД 16593450 .
- ^ Джексон, Франция; Барджиелло, штат Калифорния; Юн, С.Х.; Янг, М.В. (1986). «Продукт каждого локуса дрозофилы имеет гомологию с протеогликанами». Природа . 320 (6058): 185–188. Бибкод : 1986Natur.320..185J . дои : 10.1038/320185a0 . ПМИД 3081818 . S2CID 4305720 .
- ^ Редди, П.; Жакье, AC; Абович, Н.; Петерсен, Г.; Росбаш, М. (1986). «Локус часов периода D. melanogaster кодирует протеогликан». Клетка . 46 (1): 53–61. дои : 10.1016/0092-8674(86)90859-7 . ПМИД 3087625 . S2CID 10514568 .
- ^ Сегал, А.; Цена, JL; Ман, Б.; Янг, М.В. (1994). «Потеря циркадных поведенческих ритмов и колебаний РНК у вневременного мутанта дрозофилы ». Наука . 263 (5153): 1603–1606. Бибкод : 1994Sci...263.1603S . дои : 10.1126/science.8128246 . ПМИД 8128246 .
- ^ Цена, JL; Блау, Дж.; Ротенфлю, А.; Абодили, М.; Клосс, Б.; Янг, М.В. (1998). «двойное время — это новый часовой ген дрозофилы , который регулирует накопление белка PERIOD» . Клетка . 94 (1): 83–95. дои : 10.1016/S0092-8674(00)81224-6 . ПМИД 9674430 . S2CID 14764407 .
- ^ Душай, М.С.; Конопка, Р.Дж.; Орр, Д.; Гринакр, ML; Кириаку, КП; Росбаш, М.; Холл, Дж. К. (1990). «Фенотипический и генетический анализ Clock, нового мутанта циркадного ритма у Drosophila melanogaster » . Генетика . 125 (3): 557–578. дои : 10.1093/генетика/125.3.557 . ПМК 1204083 . ПМИД 2116357 .
- ^ Рутила, Дж. Э.; Сури, В.; Ле, М.; Итак, ВВ; Росбаш, М.; Холл, Дж. К. (1998). «CYCLE — это второй часовой белок bHLH-PAS, необходимый для циркадной ритмичности и транскрипции периода и вневременности дрозофилы » . Клетка . 93 (5): 805–814. дои : 10.1016/S0092-8674(00)81441-5 . ПМИД 9630224 . S2CID 18175560 .
- ^ Станевский, Р.; Канеко, М.; Эмери, П.; Беретта, Б.; Вагер-Смит, К.; Кей, ЮАР; Росбаш, М.; Холл, Дж. К. (1998). « Мутация crib идентифицирует криптохром как циркадный фоторецептор у дрозофилы » . Клетка . 95 (5): 681–692. дои : 10.1016/s0092-8674(00)81638-4 . ПМИД 9845370 . S2CID 6996815 .
- ^ Эмери, П.; Итак, ВВ; Канеко, М.; Холл, Дж. К.; Росбаш, М. (1998). «CRY, часы дрозофилы и светорегулируемый криптохром, вносят основной вклад в сброс циркадных ритмов и фоточувствительность» . Клетка . 95 (5): 669–679. дои : 10.1016/S0092-8674(00)81637-2 . ПМИД 9845369 . S2CID 15629055 .
- ^ Мэй, К.; Дворник, В. (2015). «Эволюционная история суперсемейства фотолиаз/криптохромов у эукариот» . ПЛОС ОДИН . 10 (9): e0135940. Бибкод : 2015PLoSO..1035940M . дои : 10.1371/journal.pone.0135940 . ПМЦ 4564169 . ПМИД 26352435 .
- ^ Парк, Дж. Х.; Холл, Дж. К. (1998). «Выделение и хронобиологический анализ гена нейропептидного фактора диспергирования пигмента у Drosophila melanogaster » . Журнал биологических ритмов . 13 (3): 219–228. дои : 10.1177/074873098129000066 . ПМИД 9615286 . S2CID 20190155 .
- ^ Ренн, Южная Каролина; Парк, Дж. Х.; Росбаш, М.; Холл, Дж. К.; Тагерт, PH (1999). «Мутация гена нейропептида PDF и абляция нейронов PDF вызывают серьезные нарушения поведенческих циркадных ритмов у дрозофилы » . Клетка . 99 (7): 791–802. дои : 10.1016/s0092-8674(00)81676-1 . ПМИД 10619432 . S2CID 62796150 .
- ^ Мартинек, С.; Иноног, С.; Манукян, А.С.; Янг, М.В. (2001). «Роль гена полярности сегмента shaggy / GSK-3 в циркадных часах дрозофилы » . Клетка . 105 (6): 769–779. дои : 10.1016/S0092-8674(01)00383-X . ПМИД 11440719 . S2CID 17434240 .
- ^ Велери, С.; Вюльбек, К. (2004). «Уникальные автономные циркадные осцилляторы в мозгу Drosophila melanogaster ». Хронобиология Интернэшнл . 21 (3): 329–342. дои : 10.1081/CBI-120038597 . ПМИД 15332440 . S2CID 15099796 .
- ^ Ригер, Д.; Станевский, Р.; Хелфрих-Фёрстер, К. (2003). «Криптохром, сложные глаза, глазки Гофбауэра-Бюхнера и глазки играют разную роль в захвате и маскировке ритма двигательной активности у плодовой мухи Drosophila melanogaster ». Журнал биологических ритмов . 18 (5): 377–391. дои : 10.1177/0748730403256997 . ПМИД 14582854 . S2CID 15292555 .
- ^ Ёси, Т.; Ригер, Д.; Хелфрих-Фёрстер, К. (2012). Два часа в мозгу: обновление модели утреннего и вечернего осциллятора у дрозофилы . Прогресс в исследованиях мозга. Том. 199. стр. 59–82. дои : 10.1016/B978-0-444-59427-3.00027-7 . ПМИД 22877659 .
- ^ Шлихтинг, М.; Греблер, Р.; Пешель, Н.; Ёси, Т.; Хелфрих-Фёрстер, К. (2014). «Обнаружение лунного света эндогенными часами дрозофилы зависит от множества фотопигментов в сложных глазах». Журнал биологических ритмов . 29 (2): 75–86. дои : 10.1177/0748730413520428 . ПМИД 24682202 . S2CID 6759377 .
- ^ Перейти обратно: а б Нитабах, Миннесота; Тагерт, PH (2008). «Организация циркадного контура управления дрозофилой » . Современная биология . 18 (2): 84–93. дои : 10.1016/j.cub.2007.11.061 . ПМИД 18211849 . S2CID 9321488 .
- ^ Ёси, Т.; Герман-Луибль, К.; Хелфрих-Фёрстер, К. (2015). «Циркадные пути поступления света у дрозофилы » . Коммуникативная и интегративная биология . 9 (1): e1102805. дои : 10.1080/19420889.2015.1102805 . ПМЦ 4802797 . ПМИД 27066180 .
- ^ Бутройд, CE; Янг, М.В. (2008). «Входы и выходы циркадных часов дрозофилы ». Анналы Нью-Йоркской академии наук . 1129 (1): 350–357. Бибкод : 2008NYASA1129..350B . дои : 10.1196/анналы.1417.006 . ПМИД 18591494 . S2CID 2639040 .
- ^ Грима, Б.; Ламуру, А.; Чело, Э.; Папен, К.; Лимбург-Бушон, Б.; Руйер, Ф. (2002). «Протеин F-box slimm контролирует уровни часовых белков периода и вне времени». Природа . 420 (6912): 178–182. Бибкод : 2002Natur.420..178G . дои : 10.1038/nature01122 . ПМИД 12432393 . S2CID 4428779 .
- ^ Ко, Х.В.; Цзян, Дж.; Эдери, И. (2002). «Роль Slimb в деградации белка периода дрозофилы , фосфорилированного Doubletime». Природа . 420 (6916): 673–678. Бибкод : 2002Natur.420..673K . дои : 10.1038/nature01272 . ПМИД 12442174 . S2CID 4414176 .
- ^ Хелфрих-Фёрстер, К. (2005). «Нейробиология циркадных часов плодовой мухи» . Гены, мозг и поведение . 4 (2): 65–76. дои : 10.1111/j.1601-183X.2004.00092.x . ПМИД 15720403 . S2CID 26099539 .
- ^ Буша, А.; Эмери-Ле, М.; Росбаш, М.; Эмери, П. (2004). «Роль двух структурных доменов КРИПТОХРОМА дрозофилы в циркадной фоторецепции». Наука . 304 (5676): 1503–1506. Бибкод : 2004Sci...304.1503B . дои : 10.1126/science.1096973 . ПМИД 15178801 . S2CID 18388605 .
- ^ Кох, К.; Чжэн, X.; Сегал, А. (2006). «JETLAG сбрасывает циркадные часы дрозофилы , способствуя вызванной светом деградации TIMELESS» . Наука . 312 (5781): 1809–1812. Бибкод : 2006Sci...312.1809K . дои : 10.1126/science.1124951 . ПМЦ 2767177 . ПМИД 16794082 .