ракообразное
| Ракообразные Временной диапазон: Кембрий, чтобы представить | |
|---|---|
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Клэйд : | Панкрустацея |
| Подтип: | Ракообразные |
| Группы включены | |
| Кладистически включенные, но традиционно исключенные таксоны | |








Ракообразные — группа членистоногих , входящих в подтип Crustacea ( / k r ə ˈ s t eɪ ʃ ə / ), большую, разнообразную группу преимущественно водных членистоногих , включая десятиногих ( креветки , креветки , крабы , омары и раки ). , семенные креветки , жаберные моллюски , рыбьи вши , криль , ремипеды , изоподы , ракушки , копеподы , креветки-опоссумы , амфиподы и креветки-богомолы . [1] Группу ракообразных можно рассматривать как подтип клады Mandibulata . В настоящее время общепризнано, что шестиногие ( насекомые и энтогнатаны ) возникли глубоко в группе ракообразных, а законченная группа называется Pancrustacea . [2] Три класса Cephalocarida , Branchiopoda и Remipedia более тесно связаны с шестиногими ракообразными, чем с любыми другими ракообразными ( олигостраканами и многоракообразными ). [3]
Размер 67 000 описанных видов варьируется от Stygotantulus stocki размером 0,1 мм (0,004 дюйма) до японского краба-паука с размахом ног до 3,8 м (12,5 футов) и массой 20 кг (44 фунта). Как и другие членистоногие , ракообразные имеют экзоскелет , который они линяют, чтобы расти. От других групп членистоногих, таких как насекомые , многоножки и хелицераты , они отличаются наличием двуветвистых (двураздельных) конечностей, а также личиночными формами , такими как науплиусная стадия жаберопод и веслоногих .
Большинство ракообразных являются свободноживущими водными животными , но некоторые из них являются наземными (например, мокрицы , песчанки ), некоторые паразитическими (например, Rhizocephala , рыбьи вши , языковые черви ), а некоторые ведут сидячий образ жизни (например, ракушки ). Группа имеет обширную летопись окаменелостей , начиная с кембрия . Более 7,9 миллионов тонн ракообразных в год добывается рыболовством или сельским хозяйством для потребления человеком. [4] состоящий в основном из креветок и креветок . Криль и копеподы не так широко вылавливаются, но могут быть животными с наибольшей биомассой на планете и составлять жизненно важную часть пищевой цепи. Научное изучение ракообразных известно как карцинология (альтернативно — малакостракология , ракообразные или корталогия ), а учёный, работающий в области канцерологии, — карцинолог .
Анатомия [ править ]

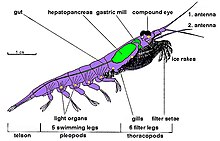
Тело ракообразного состоит из сегментов, которые сгруппированы в три отдела: головной мозг или голова , [5] переон грудная или клетка , [6] и плеон или живот . [7] Голова и грудная клетка могут сливаться вместе, образуя головогрудь . [8] который может быть покрыт одним большим панцирем . [9] Тело ракообразного защищено твердым экзоскелетом , который необходимо линять , чтобы животное могло расти. Оболочку вокруг каждого сомита можно разделить на дорсальную тергум , вентральную грудину и латеральный плевр. Различные части экзоскелета могут быть слиты вместе. [10] : 289
Каждый сомит , или сегмент тела, может нести пару придатков : на сегментах головы к ним относятся две пары усиков , мандибулы и максиллы ; [5] грудные сегменты несут ноги , которые могут специализироваться как переоподы (ходячие ноги) и максиллепеды (кормящие ноги). [6] Malacostraca и Remipedia (и шестиногие) имеют брюшные придатки. Все остальные классы ракообразных имеют брюшко без конечностей, за исключением тельсона и хвостовых ветвей , которые имеются у многих групп. [11] [12] Брюшко у малакостраканов несет плеоподы . [7] и заканчивается тельсоном, который несет анус , и часто окружен уроподами, образующими хвостовой веер . [13] Количество и разнообразие придатков у разных ракообразных могут частично отвечать за успех группы. [14]
ракообразных Придатки обычно двуветвистые , то есть разделены на две части; сюда входит вторая пара усиков, но не первая, которая обычно одноветвистая , за исключением класса Malacostraca, у которого антеннулы могут быть обычно двуветвистыми или даже трехветвистыми. [15] [16] Неясно, является ли двойственное состояние производным состоянием, развившимся у ракообразных, или вторая ветвь конечности утеряна во всех других группах. Трилобиты , например, также обладали двуветвистыми придатками. [17]
Основная полость тела представляет собой открытую систему кровообращения , где кровь перекачивается в гемоцель сердцем, расположенным вблизи спины. [18] Малакострака содержит гемоцианин в качестве пигмента, переносящего кислород, тогда как копеподы, остракоды, ракушки и жаберные моллюски содержат гемоглобины . [19] Пищеварительный канал состоит из прямой трубки, часто имеющей желудочную «желудочную мельницу» для измельчения пищи, и пары пищеварительных желез, поглощающих пищу; эта структура имеет спиральный формат. [20] Рядом с усиками расположены структуры, выполняющие функцию почек. Мозг существует в виде ганглиев, расположенных рядом с усиками, а под кишечником находится группа крупных ганглиев. [21]
У многих десятиногих первая (а иногда и вторая) пара плеопод специализируются у самцов на переносе спермы. Многие наземные ракообразные (например, красный краб с острова Рождества ) спариваются сезонно и возвращаются в море, чтобы выпустить икру. Другие, например мокрицы , откладывают яйца на суше, хотя и во влажных условиях. У большинства десятиногих самки сохраняют яйца до тех пор, пока из них не вылупятся свободно плавающие личинки. [22]
Экология [ править ]

Большинство ракообразных ведут водный образ жизни, обитая либо в морской, либо в пресноводной среде, но некоторые группы приспособились к жизни на суше, например, наземные крабы , наземные крабы-отшельники и мокрицы . Морские ракообразные столь же повсеместно распространены в океанах, как насекомые на суше. [23] [24] Большинство ракообразных также подвижны , передвигаются независимо, хотя некоторые таксономические единицы являются паразитами и живут прикрепленными к своим хозяевам (включая морских вшей , рыбьих вшей , китовых вшей , языковых червей и Cymothoa exigua , всех из которых можно назвать ракообразные вши»), а взрослые ракушки ведут сидячий образ жизни – прикрепляются к субстрату головой вперед и не могут передвигаться самостоятельно. Некоторые бранхиураны способны противостоять быстрым изменениям солености , а также переключают хозяев с морских видов на неморские. [25] : 672 Криль является нижним слоем и наиболее важной частью пищевой цепи в сообществах животных Антарктики . [26] : 64 Некоторые ракообразные являются значительными инвазивными видами , например, китайский краб-рукавица Eriocheir sinensis . [27] и азиатский прибрежный краб Hemigrapsus sanguineus . [28] После прорыва Суэцкого канала около 100 видов ракообразных из Красного моря и Индо-Тихоокеанского региона обосновались в суббассейне восточного Средиземноморья, часто оказывая значительное воздействие на местные экосистемы. [29]
Жизненный цикл [ править ]


Система спаривания [ править ]
Большинство ракообразных раздельнополы и размножаются половым путем . Фактически, недавнее исследование объясняет, как самцы T.californicus решают, с какими самками спариваться, исходя из различий в питании, предпочитая, когда самок кормят водорослями, а не дрожжами. [30] Небольшое количество — гермафродиты , в том числе ракушки , ремипеды , [31] и Цефалокарида . [32] Некоторые могут даже сменить пол в течение жизни. [32] Партеногенез также широко распространен среди ракообразных, где жизнеспособные яйца производятся самкой без необходимости оплодотворения самцом. [30] Это происходит у многих жаберных моллюсков , некоторых остракод , некоторых изопод и некоторых «высших» ракообразных, таких как раки Марморкребс .
Яйца [ править ]
У многих ракообразных оплодотворенная икра выбрасывается в толщу воды , а у других выработан ряд механизмов удержания икры до тех пор, пока она не будет готова к вылуплению. Большинство десятиногих несут яйца, прикрепленные к плеоподам , в то время как перакариды , нотостраки , аностраки и многие изоподы образуют выводковую сумку из панциря и грудных конечностей. [30] Самки Branchiura не вынашивают яйца во внешних яйцеклетках, а прикрепляют их рядами к камням и другим предметам. [33] : 788 Большинство лептостраканов и криля несут икру между грудными конечностями; некоторые копеподы несут яйца в специальных тонкостенных мешочках, а у других они скреплены длинными запутанными нитями. [30]
Личинки [ править ]
У ракообразных имеется ряд личиночных форм, из которых самой ранней и характерной является науплиус . У него три пары придатков , выходящих из головы молодого животного, и один науплиарный глаз. В большинстве групп существуют дополнительные личиночные стадии, включая зоэю (мн. zoeæ или zoeas). [34] ). Это название ему было дано, когда натуралисты считали его отдельным видом. [35] Он следует за стадией науплиуса и предшествует постличиночной стадии . Личинки зоеи плавают грудными придатками , в отличие от науплиусов, которые используют головные придатки, и мегалопы, которые для плавания используют брюшные придатки. у него часто есть шипы На панцире , которые могут помочь этим маленьким организмам поддерживать направленное плавание. [36] У многих десятиногих ракообразных из-за ускоренного развития зоэа является первой личиночной стадией. В некоторых случаях за стадией зоэи следует стадия мизиса, а в других — стадия мегалопы, в зависимости от задействованной группы ракообразных.
Обеспечивая маскировку от хищников, черные глаза у некоторых форм плавающих личинок покрыты тонким слоем кристаллического изоксантоптерина , который придает их глазам тот же цвет, что и окружающая вода, а крошечные отверстия в этом слое позволяют свету достигать сетчатки. [37] По мере того, как личинки становятся взрослыми особями, этот слой мигрирует в новое положение за сетчаткой, где он работает как зеркало обратного рассеяния, увеличивающее интенсивность света, проходящего через глаза, как это наблюдается у многих ночных животных. [38]
восстановление ДНК [ править ]
Чтобы понять, могут ли процессы репарации ДНК защитить ракообразных от повреждения ДНК , были проведены фундаментальные исследования с целью выяснения механизмов репарации, используемых Penaeus monodon (черной тигровой креветкой). [39] Было обнаружено, что репарация двухцепочечных разрывов ДНК преимущественно осуществляется путем точной гомологичной рекомбинационной репарации. Другой, менее точный процесс, соединение концов, опосредованное микрогомологией , также используется для восстановления таких разрывов. Характер экспрессии генов, связанных с репарацией ДНК, и генов, отвечающих за повреждение ДНК, у литорального копепода Tigriopus japonicus был проанализирован после ультрафиолетового облучения. [40] Это исследование выявило повышенную экспрессию белков, связанных с процессами репарации ДНК, такими как соединение негомологичных концов , гомологичная рекомбинация , репарация вырезания оснований и репарация несоответствия ДНК .
и филогения Классификация


Название «ракообразное» восходит к самым ранним работам по описанию животных, в том числе работам Пьера Белона и Гийома Ронделе , но это название не использовалось некоторыми более поздними авторами, включая Карла Линнея , который включил ракообразных в число « аптер » в своей «Системе ». Природа . [41] Самая ранняя номенклатурная действующая работа, в которой использовалось название «Ракообразные», была » Мортена Трейна Брюнниха в «Zoologiæ Fundamenta 1772 году. [42] хотя он включил и хелицераты . в группу [41]
Подтип Crustacea включает почти 67 000 описанных видов . [43] что считается справедливым 1 ⁄ 10 до 1/100 общего числа , от поскольку большинство видов еще не открыты . [44] Хотя большинство ракообразных небольшие, их морфология сильно различается и включает в себя как самого крупного членистоногого в мире – японского краба-паука с размахом ног 3,7 метра (12 футов), так и самого крупного членистоногого в мире. [45] – и самый маленький, длиной 100 микрометров (0,004 дюйма) Stygotantulus stocki . [46] Несмотря на разнообразие форм, ракообразных объединяет особая личиночная форма, известная как науплиус .
Точные взаимоотношения ракообразных с другими таксонами по состоянию на апрель 2012 г. полностью не установлены. [update]. Исследования, основанные на морфологии, привели к гипотезе Pancrustacea . [47] в котором ракообразные и шестиногие ( насекомые и союзники) являются сестринскими группами . Более поздние исследования с использованием последовательностей ДНК предполагают, что ракообразные являются парафилетическими , с гексаподами, вложенными в более крупную кладу Pancrustacea . [48] [49]
Традиционная классификация ракообразных, основанная на морфологии, выделяла от четырех до шести классов. [50] Боуман и Абеле (1982) выделили 652 сохранившихся семейства и 38 отрядов, объединенных в шесть классов: Branchiopoda , Remipedia , Cephalocarida , Maxillopoda , Ostracoda и Malacostraca . [50] Мартин и Дэвис (2001) обновили эту классификацию, сохранив шесть классов, но включив 849 существующих семейств в 42 отрядах. Несмотря на изложение доказательств того, что Maxillopoda не является монофилетическим, они сохранили его как один из шести классов, хотя и предположили, что Maxillipoda можно заменить, возведя его подклассы в классы. [51] С тех пор филогенетические исследования подтвердили полифилию Maxillipoda и парафилетическую природу ракообразных по отношению к Hexapoda. [52] [53] [54] [55] Недавние классификации выделяют от десяти до двенадцати классов Crustacea или Pancrustacea, при этом несколько бывших подклассов челюстноногих моллюсков теперь признаны классами (например, Thecostraca , Tantulocarida , Mystacocarida , Copepoda , Branchiura и Pentastomida ). [56] [57]










Следующая кладограмма показывает обновленные отношения между различными существующими группами парафилетических ракообразных по отношению к классу Hexapoda . [53]
| Панкрустацея | Ракообразные | |
Согласно этой диаграмме, Hexapoda находятся глубоко в дереве ракообразных, и любой из Hexapoda явно ближе, например, к Multicrustacean, чем к Oligostracan.
Ископаемая запись [ править ]

Ракообразные имеют богатую и обширную летопись окаменелостей , которая начинается с таких животных, как Canadaspis и Perspicaris из среднего кембрия сланцев Бёрджесс . [58] [59] Большинство основных групп ракообразных появляются в летописи окаменелостей до конца кембрия, а именно Branchiopoda , Maxillopoda (включая ракушек и языковых червей ) и Malacostraca ; Есть некоторые дебаты относительно того, являются ли кембрийские животные, отнесенные к остракодам, действительно остракодами , которые в противном случае возникли бы в ордовике . [60] Единственные классы, которые появятся позже, — это Cephalocarida , [61] у которых нет летописи окаменелостей, и Remipedia , которые были впервые описаны по ископаемым Tesnusocaris goldichi , но не появлялись до каменноугольного периода . [62] Большинство ранних ракообразных редки, но начиная с каменноугольного периода ископаемые ракообразные становятся многочисленными . [58]

В Малакостраке окаменелости криля не известны . [63] в то время как и Hoplocarida , и Phyllopoda содержат важные группы, которые сейчас вымерли, а также существующие представители (Hoplocarida: креветки-богомолы сохранились, а Aeschronectida вымерли; [64] Phyllopoda: Canadaspidida вымерли, а Leptostraca сохранились. [59] ). Cumacea и Isopoda известны из каменноугольного периода . [65] [66] как и первые настоящие креветки-богомолы. [67] У Decapoda , креветки и полихелиды появляются в триасе [68] [69] а креветки и крабы появляются в юрском периоде . [70] [71] Ископаемая нора Ophiomorpha принадлежит креветкам-призракам, тогда как ископаемая нора Camborygma принадлежит ракам. В пермско-триасовых отложениях Нурры сохранились древнейшие (пермь: Roadian) речные норы, приписываемые креветкам-призракам (Decapoda: Axiidea, Gebiidea) и ракам (Decapoda: Astacidea, Parastacidea) соответственно. [72]
Однако большая радиация ракообразных произошла в меловой период , особенно у крабов, и, возможно, была вызвана адаптивной радиацией их основных хищников, костистых рыб . [71] Первые настоящие омары также появляются в меловом периоде. [73]
Потребление людьми [ править ]
Многие ракообразные потребляются человеком: почти 10 700 000 тонн в 2007 году было выловлено ; Подавляющую часть этой продукции составляют десятиногие ракообразные : крабы , омары , креветки , раки и креветки . [74] Более 60% по весу всех ракообразных, выловленных для потребления, составляют креветки и креветки, и почти 80% производятся в Азии, причем только Китай производит почти половину мирового объема. [74] Недесятиногие ракообразные не потребляются широко: всего 118 000 тонн криля . вылавливается [74] несмотря на то, что криль имеет одну из самых больших биомасс на планете. [75]
См. также [ править ]
Ссылки [ править ]
- ^ Калман, Уильям Томас (1911). . В Чисхолме, Хью (ред.). Британская энциклопедия . Том. 7 (11-е изд.). Издательство Кембриджского университета. п. 552.
- ^ Рота-Стабелли, Омар; Каял, Эхсан; Глисон, Дайанна; и др. (2010). «Экдизозойная митогеномика: доказательства общего происхождения четвероногих беспозвоночных, панартропод» . Геномная биология и эволюция . 2 : 425–440. дои : 10.1093/gbe/evq030 . ПМК 2998192 . ПМИД 20624745 .
- ^ Кенеманн, Стефан; Дженнер, Рональд А.; Хёнеманн, Марио; и др. (01.03.2010). «Возврат к филогении членистоногих с акцентом на взаимоотношения ракообразных» . Строение и развитие членистоногих . 39 (2–3): 88–110. Бибкод : 2010АртСД..39...88К . дои : 10.1016/j.asd.2009.10.003 . ISSN 1467-8039 . ПМИД 19854296 .
- ^ «Состояние мирового рыболовства и аквакультуры в 2018 году – достижение целей устойчивого развития» (PDF) . Фао.орг . Рим: Продовольственная и сельскохозяйственная организация Объединенных Наций . 2018.
- ↑ Перейти обратно: Перейти обратно: а б «Цефалон» . Словарь ракообразных . Музей естественной истории округа Лос-Анджелес . Архивировано из оригинала 27 июля 2011 г. Проверено 10 сентября 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б «Торакс» . Словарь ракообразных . Музей естественной истории округа Лос-Анджелес . Архивировано из оригинала 27 июля 2011 г. Проверено 10 сентября 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б «Живот» . Словарь ракообразных . Музей естественной истории округа Лос-Анджелес . Архивировано из оригинала 27 июля 2011 г. Проверено 10 сентября 2016 г.
- ^ «Цефалоторакс» . Словарь ракообразных . Музей естественной истории округа Лос-Анджелес . Архивировано из оригинала 27 июля 2011 г. Проверено 10 сентября 2016 г.
- ^ «Панцирь» . Словарь ракообразных . Музей естественной истории округа Лос-Анджелес . Архивировано из оригинала 27 июля 2011 г. Проверено 10 сентября 2016 г.
- ^ Пи Джей Хейворд; Дж. С. Райланд (1995). Справочник морской фауны Северо-Западной Европы . Издательство Оксфордского университета . ISBN 978-0-19-854055-7 . Проверено 10 сентября 2016 г.
- ^ Фрич, Мартин; Рихтер, Стефан (5 сентября 2022 г.). «Как паттерн тела мог повлиять на эволюцию членистоногих — на примере мистакокариды Derocheilocaris remanei (Crustacea, Oligostraca)» . Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 338 (6): 342–359. Бибкод : 2022JEZB..338..342F . дои : 10.1002/jez.b.23140 . ПМИД 35486026 . S2CID 248430846 .
- ^ Морфология мозга Hutchinsoniella macracantha (Cephalocarida, Crustacea) - стр. 290.
- ^ «Тельсон» . Словарь ракообразных . Музей естественной истории округа Лос-Анджелес . Архивировано из оригинала 27 июля 2011 г. Проверено 10 сентября 2016 г.
- ^ Элизабет Пенниси (4 июля 1997 г.). «Крабовые ножки и клешни омара». Наука . 277 (5322): 36. дои : 10.1126/science.277.5322.36 . S2CID 83148200 .
- ^ «Антеннуля» . Словарь ракообразных . Музей естественной истории округа Лос-Анджелес . Архивировано из оригинала 5 ноября 2013 г. Проверено 10 сентября 2016 г.
- ^ «Ракообразные: придатки» . Калифорнийский университет в Беркли . Проверено 10 сентября 2016 г.
- ^ NC Хьюз (февраль 2003 г.). «Тагмоз трилобитов и рисунок тела с точки зрения морфологии и развития» . Интегративная и сравнительная биология . 43 (1): 185–206. дои : 10.1093/icb/43.1.185 . ПМИД 21680423 .
- ^ Акира Сакурай. «Закрытая и открытая система кровообращения» . Государственный университет Джорджии . Архивировано из оригинала 17 сентября 2016 г. Проверено 10 сентября 2016 г.
- ^ Клаус Урих (1994). «Дыхательные пигменты» . Сравнительная биохимия животных . Спрингер . стр. 249–287. ISBN 978-3-540-57420-0 .
- ^ Х. Дж. Чекальди. Анатомия и физиология пищеварительного тракта десятиногих ракообразных, выращиваемых в аквакультуре (PDF) . АКВАКОП, ИФРЕМЕР. Actes de Colloque 9. стр. 243–259.
{{cite book}}:|work=игнорируется ( помогите ) [ постоянная мертвая ссылка ] - ^ Гизелин, Майкл Т. (2005). «Рачок». Энкарта . Майкрософт .
- ^ Беркенроуд, доктор медицины (1963). «Эволюция Eucarida (Crustacea, Eumalacostraca) по отношению к летописи окаменелостей». Тулейнские исследования в области геологии . 2 (1): 1–17.
- ^ «Крабы, омары, креветки и другие ракообразные» . Австралийский музей . 5 января 2010 г. Проверено 10 сентября 2016 г.
- ^ «Бентосные животные» . Министерство рыболовства и сельского хозяйства Исландии . Архивировано из оригинала 11 мая 2014 г. Проверено 10 сентября 2016 г.
- ^ Алан П. Кович; Джеймс Х. Торп (1991). «Рачки: интродукция и перакариды» . У Джеймса Х. Торпа; Алан П. Кович (ред.). Экология и классификация пресноводных беспозвоночных Северной Америки (1-е изд.). Академическая пресса . стр. 665–722. ISBN 978-0-12-690645-5 . Проверено 10 сентября 2016 г.
- ^ Добродетель, ПД; Николс, доктор медицинских наук; Николс, С. (1997). «Механизмы выживания Euphasia superba , связанные с диетой : биохимические изменения во время длительного голодания и бактерии как возможный источник питания». . В Бруно Батталье; Валенсия, Хосе; Уолтон, DWH (ред.). Антарктические сообщества: виды, структура и выживание . Издательство Кембриджского университета . ISBN 978-0-521-48033-8 . Проверено 10 сентября 2016 г.
- ^ Голлаш, Стефан (30 октября 2006 г.). « Эриохейр китайский » (PDF) . Глобальная база данных инвазивных видов . Группа специалистов по инвазивным видам . Архивировано из оригинала (PDF) 24 декабря 2017 г. Проверено 10 сентября 2016 г.
- ^ Джон Дж. Макдермотт (1999). «Западно-тихоокеанский брахиуран Hemigrapsus sanguineus (Grapsidae) в новой среде обитания вдоль атлантического побережья США: питание, морфология и рост хелипедов» . В Шраме, Фредерик Р .; Кляйн, Дж. К. фон Ваупель (ред.). Ракообразные и кризис биоразнообразия: материалы Четвертого Международного конгресса по ракообразным, Амстердам, Нидерланды, 20–24 июля 1998 г. Конинклийке Брилл . стр. 425–444. ISBN 978-90-04-11387-9 . Проверено 10 сентября 2016 г.
- ^ Галил, Белла; Фрогья, Карло; Ноэль, Пьер (2002). Бриан, Фредерик (ред.). CIESM Атлас экзотических видов Средиземноморья: Том 2 Ракообразные . Париж, Монако: Издательство CIESM. п. 192. ИСБН 92-990003-2-8 .
- ↑ Перейти обратно: Перейти обратно: а б с д «Ракообразное (членистоногое)» . Британская энциклопедия . 5 мая 2023 г.
- ^ ГЛ Пеше. «Ремипедия Ягер, 1981» .
- ↑ Перейти обратно: Перейти обратно: а б Д.Э. Эйкен; В. Танниклифф; КТ Ши; Л. Д. Делорм. «Рачок» . Канадская энциклопедия . Архивировано из оригинала 7 июня 2011 г. Проверено 10 сентября 2016 г.
- ^ Алан П. Кович; Джеймс Х. Торп (2001). «Знакомство с подтипом ракообразных» . У Джеймса Х. Торпа; Алан П. Кович (ред.). Экология и классификация пресноводных беспозвоночных Северной Америки (2-е изд.). Академическая пресса . стр. 777–798. ISBN 978-0-12-690647-9 . Проверено 10 сентября 2016 г.
- ^ «Зоя» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Калман, Уильям Томас (1911). . В Чисхолме, Хью (ред.). Британская энциклопедия . Том. 7 (11-е изд.). Издательство Кембриджского университета. п. 356.
- ^ WFR Велдон (июль 1889 г.). «Заметка о функции шипов ракообразных zoœa» (PDF) . Журнал Морской биологической ассоциации Соединенного Королевства . 1 (2): 169–172. Бибкод : 1889JMBUK...1..169W . дои : 10.1017/S0025315400057994 . S2CID 54759780 . Архивировано из оригинала (PDF) 17 июля 2011 г.
- ^ Шавит, Кешет и др., Настраиваемый отражатель, позволяющий ракообразным видеть, но не быть видимым , Science , 16 февраля 2023 г., опубликовано в томе 379, выпуск 6633, 17 февраля 2023 г.
- ^ Дафф, Мэг (16 февраля 2023 г.). « Блеск для глаз Disco делает детенышей ракообразных невидимыми» . Сланец – через slate.com.
- ^ Шривастава, Шикха; Дахал, Сумедха; Найду, Шаранья Дж.; Ананд, Дипика; Гопалакришнан, Видья; Кулот Валаппил, Раджендран; Рагхаван, Сатис К. (24 января 2017 г.). «Репарация двухцепочечного разрыва ДНК у Penaeus monodon преимущественно зависит от гомологичной рекомбинации» . Исследование ДНК . 24 (2): 117–128. дои : 10.1093/dnares/dsw059 . ПМК 5397610 . ПМИД 28431013 .
- ^ Ри, Дж. С.; Ким, Б.М.; Чой, бакалавр наук; Ли, Дж. С. (2012). «Анализ характера экспрессии генов, связанных с репарацией ДНК и реакцией на повреждение ДНК, выявленных с помощью олигомикрочипа 55K при облучении УФ-В у приливных веслоногих ракообразных Tigriopus japonicus». Сравнительная биохимия и физиология. Токсикология и фармакология . 155 (2): 359–368. дои : 10.1016/j.cbpc.2011.10.005 . ПМИД 22051804 .
- ↑ Перейти обратно: Перейти обратно: а б Липке Б. Холтуис (1991). "Введение" . Морские омары мира . Каталог видов ФАО, том 13. Продовольственная и сельскохозяйственная организация . стр. 1–2. ISBN 978-92-5-103027-1 . [ постоянная мертвая ссылка ]
- ^ М. Т. Брюнних (1772). Основы зоологии в сопровождении академических лекций. Grunde i Dyrelaeren (на латыни и датском языке). Копенгаген и Лейпциг: Фридрих Кристиан Пелт. стр. 1 -254.
- ^ Чжи-Цян Чжан (2011). З.-К. Чжан (ред.). «Биоразнообразие животных: план классификации более высокого уровня и обзор таксономического богатства - Тип Arthropoda von Siebold, 1848» (PDF) . Зоотакса . 4138 : 99–103.
- ^ «Рачки — морские клопы» . Те Ара: Энциклопедия Новой Зеландии . Министерство культуры и наследия . Проверено 10 сентября 2016 г.
- ^ «Японские крабы-пауки прибыли в аквариум» . Аквариум побережья Орегона . Архивировано из оригинала 23 марта 2010 г. Проверено 10 сентября 2016 г.
- ^ Крейг Р. Макклейн; Элисон Г. Бойер (22 июня 2009 г.). «Биоразнообразие и размер тела у многоклеточных животных связаны» . Труды Королевского общества B: Биологические науки . 276 (1665): 2209–2215. дои : 10.1098/rspb.2009.0245 . ПМК 2677615 . ПМИД 19324730 .
- ^ Я. Зрзавы; П. Штис (май 1997 г.). «Основное строение тела членистоногих: данные эволюционной морфологии и биологии развития» . Журнал эволюционной биологии . 10 (3): 353–367. дои : 10.1046/j.1420-9101.1997.10030353.x . S2CID 84906139 .
- ^ Джером К. Режье; Джеффри В. Шульц; Андреас Цвик; Эйприл Хасси; Бернард Болл; Регина Ветцер; Джоэл В. Мартин; Клиффорд В. Каннингем (25 февраля 2010 г.). «Отношения членистоногих, выявленные с помощью филогеномного анализа последовательностей, кодирующих ядерные белки». Природа . 463 (7284): 1079–1083. Бибкод : 2010Natur.463.1079R . дои : 10.1038/nature08742 . ПМИД 20147900 . S2CID 4427443 .
- ^ Бьорн М. фон Роймонт; Рональд А. Дженнер; Мэтью А. Уиллс; Эмилиано Делл'Ампио; Гюнтер-Пасс; Инго Эберсбергер; Бенджамин Мейер; Стефан Кенеманн; Томас М. Илифф; Александрос Стаматакис; Оливер Нихейс; Карен Мейсеманн; Бернхард Мисоф (март 2012 г.). «Филогения панкрустообразных в свете новых филогеномических данных: поддержка Remipedia как возможной сестринской группы Hexapoda» . Молекулярная биология и эволюция . 29 (3): 1031–1045. дои : 10.1093/molbev/msr270 . ПМИД 22049065 .
- ↑ Перейти обратно: Перейти обратно: а б Джоэл В. Мартин; Джордж Э. Дэвис (2001). Обновленная классификация современных ракообразных (PDF) . Музей естественной истории округа Лос-Анджелес . стр. 1–132. Архивировано из оригинала (PDF) 12 мая 2013 г. Проверено 14 декабря 2009 г.
- ^ Хайс, Рони (2003). «Обновленная классификация современных ракообразных» . обзор. Журнал биологии ракообразных . 23 (2): 495–497. дои : 10.1163/20021975-99990355 .
- ^ Окли, Тодд Х.; Вулф, Джоанна М.; Линдгрен, Энни Р.; Захаров, Александр К. (январь 2013 г.). «Филотранскриптомика, чтобы привлечь к себе малоизученных: монофилетические остракоды, размещение окаменелостей и филогения панракообразных» . Молекулярная биология и эволюция . 30 (1): 215–233. дои : 10.1093/molbev/mss216 . ПМИД 22977117 .
- ↑ Перейти обратно: Перейти обратно: а б Швентнер, М; Комбош, диджей; Нельсон, JP; Гирибет, Г (2017). «Филогеномное решение происхождения насекомых путем разрешения взаимоотношений ракообразных и шестиногих» . Современная биология . 27 (12): 1818–1824.e5. Бибкод : 2017CBio...27E1818S . дои : 10.1016/j.cub.2017.05.040 . ПМИД 28602656 .
- ^ Лосано-Фернандес, Хесус; Джакомелли, Маттиа; Флеминг, Джеймс Ф.; Чен, Альберт; Винтер, Якоб; Томсен, Филип Фрэнсис; Гленнер, Хенрик; Палеро, Ферран; Легг, Дэвид А.; Илифф, Томас М.; Пизани, Давиде; Олесен, Йорген (2019). «Эволюция панкрустообразных, освещенная богатыми таксонами наборами данных геномного масштаба с расширенной выборкой реминожек» . Геномная биология и эволюция . 11 (8): 2055–2070. дои : 10.1093/gbe/evz097 . ПМК 6684935 . ПМИД 31270537 .
- ^ Бернот, Джеймс П.; Оуэн, Кристофер Л.; Вулф, Джоанна М.; Меланд, Кеннет; Олесен, Йорген; Крэндалл, Кейт А. (2023). «Основные изменения в филогении панкрустообразных и доказательства чувствительности к выборке таксонов». Молекулярная биология и эволюция . 40 (8): msad175. дои : 10.1093/molbev/msad175 . PMC 10414812. PMID 37552897 .
- ^ Бруска, Ричард К. (2016). Беспозвоночные (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. п. 222. ИСБН 9781605353753 .
- ^ Гирибет, Г.; Эджкомб, Джорджия (2020). Древо жизни беспозвоночных . Издательство Принстонского университета . п. 21. ISBN 978-0-6911-7025-1 . Проверено 27 мая 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б «Ископаемый рекорд» . Ископаемые группы: Ракообразные . Бристольский университет . Архивировано из оригинала 7 сентября 2016 г. Проверено 10 сентября 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б Бриггс, Дерек (23 января 1978 г.). «Морфология, образ жизни и сходство Canadaspis perfecta (Crustacea: Phyllocarida), средний кембрий, сланцы Берджесс, Британская Колумбия». Философские труды Королевского общества Б. 281 (984): 439–487. Бибкод : 1978RSPTB.281..439B . дои : 10.1098/rstb.1978.0005 .
- ^ Олни, Мэтью. «Остракоды» . Знакомство с микропалеонтологией . Университетский колледж, Лондон . Проверено 10 сентября 2016 г.
- ^ Хесслер, Р.Р. (1984). «Цефалокарида: живое ископаемое без летописи окаменелостей» . У Н. Элдриджа; С.М. Стэнли (ред.). Живые ископаемые . Нью-Йорк: Springer Verlag. стр. 181–186. ISBN 978-3-540-90957-6 .
- ^ Кенеманн, Стефан ; Шрам, Фредерик Р.; Хёнеманн, Марио; Илифф, Томас М. (12 апреля 2007 г.). «Филогенетический анализ Remipedia (Crustacea)». Разнообразие и эволюция организмов . 7 (1): 33–51. Бибкод : 2007ODivE...7...33K . дои : 10.1016/j.ode.2006.07.001 .
- ^ «Антарктическая предыстория» . Австралийский антарктический отдел . 29 июля 2008 года. Архивировано из оригинала 30 сентября 2009 года . Проверено 25 февраля 2010 г.
- ^ Дженнер, Рональд А.; Хоф, Сис Х.Дж.; Шрам, Фредерик Р. (1998). «Палео- и археостоматоподы (Hoplocarida: Crustacea) из известняка Медвежьего ущелья, штат Миссисипи (Намюр), центральной Монтаны» . Вклад в зоологию . 67 (3): 155–186. дои : 10.1163/18759866-06703001 .
- ^ Шрам, Фредерик; Хоф, Сис Х.Дж.; Мейпс, Ройал Х. и Сноудон, Полли (2003). «Палеозойские кумовые (Crustacea, Malacostraca, Peracarida) Северной Америки» . Вклад в зоологию . 72 (1): 1–16. дои : 10.1163/18759866-07201001 .
- ^ Шрам, Фредерик Р. (28 августа 1970 г.). «Изопода из Пенсильвании, штат Иллинойс». Наука . 169 (3948): 854–855. Бибкод : 1970Sci...169..854S . дои : 10.1126/science.169.3948.854 . ПМИД 5432581 . S2CID 31851291 .
- ^ Хоф, Cees HJ (1998). «Ископаемые ротоногие моллюски (Crustacea: Malacostraca) и их филогенетическое влияние». Журнал естественной истории . 32 (10 и 11): 1567–1576. Бибкод : 1998JNatH..32.1567H . дои : 10.1080/00222939800771101 .
- ^ Крин, Роберт П.Д. (14 ноября 2004 г.). «Дендробранхиата» . Заказать Декаподу . Бристольский университет . Архивировано из оригинала 29 февраля 2012 года . Проверено 25 февраля 2010 г.
- ^ Карасава, Хироаки; Такахаши, Фумио; Дои, Эйдзи; Исида, Хидео (2003). «Первое упоминание о семействе Coleiidae Van Straelen (Crustacea: Decapoda: Eryonoides) из верхнего триаса Японии» . Палеонтологические исследования . 7 (4): 357–362. дои : 10.2517/prpsj.7.357 . S2CID 129330859 .
- ^ Чейс, Феннер А. младший; Мэннинг, Раймонд Б. (1972). «Две новые каридовые креветки, одна из которых представляет новое семейство, из морских водоемов на острове Вознесения (Crustacea: Decapoda: Natantia)». Смитсоновский вклад в зоологию . 131 (131): 1–18. дои : 10.5479/si.00810282.131 . S2CID 53067015 .
- ↑ Перейти обратно: Перейти обратно: а б Вегеле, JW (декабрь 1989 г.). «О влиянии рыб на эволюцию донных ракообразных» . Журнал зоологической систематики и эволюционных исследований . 27 (4): 297–309. дои : 10.1111/j.1439-0469.1989.tb00352.x .
- ^ Бокон, А.; Рончи, А.; Феллетти, Ф.; Нето де Карвалью, К. (2014). «Эволюция ракообразных на пороге конца пермского кризиса: ихносетевой анализ речной последовательности Нурры (пермь-триас, Сардиния, Италия)» . Палеогеография, Палеоклиматология, Палеоэкология . 410 : 74. Бибкод : 2014PPP...410...74B . дои : 10.1016/j.palaeo.2014.05.034 . Проверено 19 мая 2022 г.
- ^ Чуди, Дейл; Дональдсон, В. Стивен; Коллом, Кристофер; и др. (2005). « Hoploparia albertaensis , новый вид когтистых омаров (Nephropidae) из позднекониаковой мелководной формации Bad Heart на северо-западе Альберты, Канада». Журнал палеонтологии . 79 (5): 961–968. doi : 10.1666/0022-3360(2005)079[0961:HAANSO]2.0.CO;2 . S2CID 131067067 .
- ↑ Перейти обратно: Перейти обратно: а б с «ФИГИС: Глобальная статистика производства 1950–2007 гг.» . Продовольственная и сельскохозяйственная организация . Проверено 10 сентября 2016 г.
- ^ Никол, Стивен; Эндо, Ёсинари (1997). Мировой промысел криля . Технический документ по рыболовству. Том. 367. Продовольственная и сельскохозяйственная организация . ISBN 978-92-5-104012-6 .
Источники [ править ]
- Шрам, Фредерик (1986). Ракообразные . Издательство Оксфордского университета . ISBN 978-0-19-503742-5 .
- Пауэрс М., Хилл Г., Уивер Р. и Гойманн В. (2020). Экспериментальный тест выбора партнера по окраске красных каротиноидов у морского копепода Tigriopus Californicus. Этология., 126(3), 344–352. Экспериментальный тест выбора партнера по окраске красных каротиноидов у морского копепода Tigriopus Californicus.
Внешние ссылки [ править ]
 Тексты в Wikisource:
Тексты в Wikisource: - «Американская энциклопедия - Arc.Ask3.Ru». Американская энциклопедия . Том. 8. 1920 год.
- Кларк, Хьюберт Лайман ; Ингерсолл, Эрнест (1905). « Ракоцвет ». Новая международная энциклопедия . Том. 5.
- Crustacea.net, интернет-ресурс по биологии ракообразных.
- Ракообразные. Архивировано 9 октября 2007 года в Wayback Machine : Музей естественной истории округа Лос-Анджелес.
- Ракообразные : веб-проект «Древо жизни»
- Общество ракообразных. Архивировано 10 ноября 2011 г. в Wayback Machine.
- Коллекции естествознания: ракообразные : Эдинбургский университет
- Ракообразные (Crustacea) на берегу Сингапура
- Ракообразные (крабы, омары, креветки, креветки, ракушки). Архивировано 11 января 2012 г. в Wayback Machine : Biodiversity Explorer.