Эвриптерид
| Эвриптерид | |
|---|---|
 | |
| Ископаемый экземпляр Eurypterus remipes, хранящийся в Государственном музее естественной истории Карлсруэ в Карлсруэ , Германия. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Подтип: | Хелицерата |
| Клэйд : | Склерофората |
| Заказ: | † Эвриптерида Бурмейстер , 1843 г. |
| Подзаказы | |
Неуверенное сиденье | |
| Синонимы | |
Эвриптериды , часто неофициально называемые морскими скорпионами , представляют собой группу вымерших членистоногих , образующих отряд Eurypterida . Самые ранние известные эвриптериды относятся к дарривильскому этапу ордовикского периода 467,3 миллиона лет назад . Вероятно, эта группа впервые появилась либо в раннем ордовике , либо в позднем кембрии . Eurypterida, насчитывающий около 250 видов, является наиболее разнообразным палеозоя отрядом хелицератов . После своего появления в ордовике эвриптериды стали основными компонентами морской фауны в силурийском периоде , из которого было описано большинство видов эвриптерид. Силурийский род Eurypterus составляет более 90% всех известных экземпляров эвриптерид. Хотя группа продолжала диверсифицироваться в течение последующего девонского периода, эвриптериды сильно пострадали от вымирания в позднем девоне . Их численность и разнообразие сокращались, пока они не вымерли во время пермско-триасового вымирания (или где-то незадолго до этого) 251,9 миллион лет назад.
Хотя в народе их называли «морскими скорпионами», только самые ранние эвриптериды были морскими ; многие более поздние формы жили в солоноватой или пресной воде и не были настоящими скорпионами . Некоторые исследования предполагают, что существовала двойная дыхательная система , которая позволяла находиться в земной среде в течение коротких периодов времени. Название Eurypterida происходит от древнегреческих слов εὐρύς ( eurús ), что означает «широкий» или «широкий», и πτερόν ( pterón ), что означает «крыло», имея в виду пару широких плавательных придатков, присутствующих у многих представителей группы.
Отряд эвриптерид включает самых крупных из когда-либо существовавших членистоногих. Самый крупный из них, Jaekelopterus , достигал 2,5 метра (8,2 фута) в длину. Эвриптериды не были одинаково большими, и большинство видов имели длину менее 20 сантиметров (8 дюймов); самый маленький эвриптерид, Alkenopterus , имел длину всего 2,03 сантиметра (0,80 дюйма). Окаменелости эвриптерид были обнаружены на всех континентах. Большинство окаменелостей взято из окаменелостей в Северной Америке и Европе , поскольку эта группа жила в основном в водах вокруг и внутри древнего суперконтинента Еврамерика . Лишь несколько групп эвриптерид распространились за пределы Еврамерики, а несколько родов, таких как Adelophthalmus и Pterygotus , достигли космополитического распространения , а окаменелости были обнаружены по всему миру.
Описание
[ редактировать ]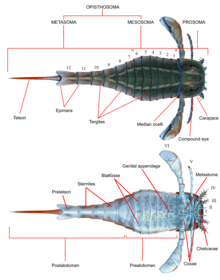
Как и все другие членистоногие , эвриптериды обладали сегментированными телами и членистыми придатками (конечностями), покрытыми кутикулой, состоящей из белков и хитина . Как и у других хелицератов , тело было разделено на две тагматы (отделения); лобная просома (голова) и задняя опистосома (брюшок). [1] Просома была покрыта панцирем ( иногда называемым «просомальным щитом»), на котором располагались как сложные глаза , так и глазки (простые глазоподобные органы чувств). [2]
Просома также имела шесть пар придатков, которые обычно называют парами придатков с I по VI. Первая пара придатков, единственная пара, располагающаяся перед ртом, называется хелицерами ( гомологичны клыкам пауков). Они были оснащены небольшими клешнями, с помощью которых манипулировали фрагментами пищи и заталкивали их в рот. [2] У одной линии, Pterygotidae , хелицеры были большими и длинными, с сильными, хорошо развитыми зубами на специализированных челах (когтях). [3] Последующие пары придатков, номера II–VI, имели гнатобазы (или «зубные пластинки») на тазиках (сегментах конечностей), используемых для питания. Эти придатки обычно представляли собой ходовые ноги цилиндрической формы, у некоторых видов покрытые шипами. В большинстве линий конечности имели тенденцию становиться больше, чем дальше назад они находились. В Eurypterina подотряде , более крупном из двух подотрядов эвриптерид, шестая пара придатков также была преобразована в весло для плавания, чтобы помочь в перемещении по водной среде. [2]
Опистосома состояла из 12 сегментов и тельсона — самого заднего отдела тела, который у большинства видов имел клиновидную форму. [2] У некоторых линий, особенно у Pterygotioidea , Hibbertopteridae и Mycteroptidae , тельсон был сплющен и, возможно, использовался в качестве руля во время плавания. Некоторые роды надсемейства Carcinosomatoidea , особенно Eusarcana , имели тельсон, похожий на тельсон современных скорпионов , и, возможно, были способны использовать его для введения яда . [4] [5] Тазики шестой пары придатков были покрыты пластинкой, называемой метастомой, первоначально полученной из полного сегмента экзоскелета. Саму опистосому можно разделить либо на « мезосому » (включающую сегменты с 1 по 6) и « метасому » (включающую сегменты с 7 по 12), либо на «преабдомен» (обычно включающий сегменты с 1 по 7) и «постабдомен» (обычно включающий сегменты с 8 по 12). [2]
Нижняя сторона опистосомы была покрыта структурами, развившимися из видоизмененных придатков опистосомы. По всей опистосоме эти структуры образовывали пластинчатые структуры, называемые Blattfüsse ( букв. « листовые ножки » на немецком языке). [6] Они создали жаберную камеру (жаберный тракт) между предшествующим Blattfüsse и вентральной поверхностью самой опистосомы, в которой находились органы дыхания. Со второго по шестой сегменты опистосом также содержали овальные или треугольные органы, которые интерпретировались как органы, способствующие дыханию. Эти органы, называемые Kiemenplatten или «жаберные тракты», потенциально помогали эвриптеридам дышать воздухом над водой, в то время как Blattfüssen , похожий на органы современных мечехвостов , покрывал бы части, служащие для подводного дыхания . [2]
Придатки сегментов 1 и 2 опистосомы (седьмой и восьмой сегменты в целом) слились в структуру, называемую половой крышкой, занимающую большую часть нижней стороны сегмента 2 опистосомы. Рядом с передним краем этой структуры находится генитальный придаток (также называемый Ципфелем или срединным брюшным придатком). Этот придаток, часто сохраняющийся очень заметно, постоянно интерпретируется как часть репродуктивной системы и встречается в двух признанных типах, которые, как предполагается, соответствуют мужскому и женскому полу. [2]
Биология
[ редактировать ]Размер
[ редактировать ]
Размер эвриптерид сильно различался в зависимости от таких факторов, как образ жизни, среда обитания и таксономическое родство . Размеры около 100 сантиметров (3,3 фута) обычны для большинства групп эвриптерид. [7] Самый маленький эвриптерид, Alkenopterus burglahrensis , имел длину всего 2,03 сантиметра (0,80 дюйма). [8]
Самый крупный эвриптерид и самое большое известное членистоногое из когда-либо живших — Jaekelopterus rhenaniae . Хелицера из эмсийской формации Клерф в Вильверате, Германия , имела длину 36,4 сантиметра (14,3 дюйма), но ей не хватало четверти ее длины, что позволяет предположить, что длина полной хелицеры должна была составлять 45,5 сантиметра (17,9 дюйма). Если бы пропорции между длиной тела и хелицерами соответствовали пропорциям его ближайших родственников, у которых соотношение размера когтей и длины тела относительно постоянное, экземпляр Jaekelopterus , обладавший рассматриваемыми хелицерами, имел бы размеры от 233 до 259 сантиметров (7,64 и 8,50). футов), в среднем 2,5 метра (8,2 фута) в длину. Если хелицеры вытянутся, к этой длине добавится еще один метр (3,28 фута). Эта оценка превышает максимальный размер тела всех других известных гигантских членистоногих почти на полметра (1,64 фута), даже если не учитывать расширенные хелицеры. [9] По оценкам, две другие эвриптериды также достигали длины 2,5 метра; Erettopterus grandis (близкий родственник Jaekelopterus ) и Hibbertopterus wittebergensis , но E. grandis очень фрагментирован, и оценка размера H. wittenbergensis основана на следах, а не на ископаемых останках. [10]
Семейство Jaekelopterus , Pterygotidae, известно несколькими необычайно крупными видами. И Acutiramus , чей самый крупный представитель A. bohemicus имел размер 2,1 метра (6,9 фута), и Pterygotus , чей самый крупный вид P. grandidentatus имел размеры 1,75 метра (5,7 фута), были гигантскими. [9] Было предложено несколько различных факторов, способствующих большому размеру птериготид, включая ухаживание, хищничество и конкуренцию за ресурсы окружающей среды. [11]
Гигантские эвриптериды не ограничивались семейством Pterygotidae. Изолированная ископаемая метастома карциносоматоидного эвриптерида Carcinosoma punctatum длиной 12,7 см (5,0 дюйма) указывает на то, что при жизни животное достигало бы длины 2,2 метра (7,2 фута), соперничая по размеру с птериготидами. [12] Другим гигантом был Pentecopterus Decorahensis , примитивный карцинозоматоид, длина которого, по оценкам, достигала 1,7 метра (5,6 футов). [13]
Для крупных эвриптерид характерно легкое телосложение. Такие факторы, как передвижение, затраты энергии на линьку и дыхание, а также фактические физические свойства экзоскелета , ограничивают размер, которого могут достичь членистоногие. Легкая конструкция существенно снижает влияние этих факторов. Птериготиды были особенно легкими: большинство окаменелых крупных сегментов тела сохранились тонкими и неминерализованными. [9] Легкие адаптации присутствуют и у других гигантских палеозойских членистоногих, таких как гигантская многоножка Arthropleura , и, возможно, имеют жизненно важное значение для эволюции гигантских размеров у членистоногих. [9] [14]
Помимо легких гигантских эвриптерид, очень крупными были и некоторые глубокотелые формы семейства Hibbertopteridae. Панцирь из каменноугольного периода Шотландии, относящийся к виду Hibbertoperus scouleri, имеет ширину 65 см (26 дюймов). Поскольку Hibbertopterus был очень широким по сравнению с его длиной, рассматриваемое животное могло иметь длину чуть меньше 2 метров (6,6 футов). Более крепкий, чем птериготиды, этот гигантский гиббертоптерус , возможно, мог бы конкурировать с самыми крупными птериготидами по весу, если не превзойти их, и, как таковой, был бы одним из самых тяжелых членистоногих. [15]
Передвижение
[ редактировать ]

Два подотряда эвриптерид, Eurypterina и Stylonurina , отличаются прежде всего морфологией последней пары придатков. У Stylonurina этот придаток имеет форму длинной и тонкой шагающей ноги, а у Eurypterina нога видоизменена и расширена в плавательное весло. [16] Если не считать весла для плавания, ноги многих эвриптеринов были слишком малы, чтобы позволить им делать что-то большее, чем просто ползать по морскому дну . Напротив, у ряда стилонуринов были удлиненные и мощные ноги, которые, возможно, позволяли им ходить по суше (по аналогии с современными крабами ). [17]
Ископаемая тропа была обнаружена в ископаемых отложениях каменноугольного возраста в Шотландии в 2005 году. Ее приписали стилонуриновому эвриптериду Hibbertopterus из-за соответствующего размера (по оценкам, длина следа составляла около 1,6 метра (5,2 фута)) и предполагаемой анатомии ног. . Это самая большая наземная тропа длиной 6 метров (20 футов) и шириной в среднем 95 сантиметров (3,12 фута), проложенная найденным на данный момент членистоногим. Это первая запись эвриптерид о передвижении по суше. Этот след предоставляет доказательства того, что некоторые эвриптериды могут выжить в наземной среде, по крайней мере, в течение коротких периодов времени, и дает информацию о стилонуриновой походке. У Hibbertopterus , как и у большинства эвриптерид, пары придатков различаются по размеру (так называемое гетероподное состояние конечностей). Эти пары разного размера двигались синхронно, а короткая длина шага указывает на то, что Hibbertopterus полз с исключительно медленной скоростью, по крайней мере, на суше. Большой тельсон тащился по земле и оставлял за животным большую центральную бороздку. Наклоны следов через случайные промежутки времени позволяют предположить, что движение было прерывистым. [18] Походка более мелких стилонуринов, таких как Parastylonurus , вероятно, была быстрее и точнее. [19]
Функциональность эвриптериновых лопаток для плавания варьировалась от группы к группе. У Eurypteroidea весла по форме были похожи на весла. Состояние суставов их придатков позволяло перемещать весла только в почти горизонтальных плоскостях, а не вверх или вниз. Некоторые другие группы, такие как Pterygotioidea, не обладали бы этим состоянием и, вероятно, могли бы плавать быстрее. [20] Принято считать, что большинство эвриптеринов использовали гребной тип движения, аналогичный тому, который используют крабы и водяные жуки . Более крупные особи, возможно, были способны к подводному полету (или подводному полету ), при котором движения и формы лопастей достаточно для создания подъемной силы , подобно плаванию морских черепах и морских львов . Этот тип движения имеет относительно более медленную скорость ускорения, чем тип гребли, тем более что у взрослых лопасти пропорционально меньше, чем у молодых. Однако, поскольку более крупные размеры взрослых особей означают более высокий коэффициент лобового сопротивления , использование этого типа движения более энергоэффективно. [21]

Некоторые эвриптерины, такие как Mixopterus (как следует из обнаруженных ископаемых следов), не обязательно были хорошими пловцами. Вероятно, он держался в основном на дне, время от времени используя лопасти для резких вертикальных движений, а четвертая и пятая пары придатков были расположены назад, чтобы производить незначительное движение вперед. При ходьбе он, вероятно, использовал походку, подобную походке большинства современных насекомых. Вес его длинного брюшка уравновешивался двумя тяжелыми специализированными передними придатками, а центр тяжести можно было регулировать, поднимая и перемещая хвост. [22]
Сохранившиеся окаменелые следы эвриптерид, как правило, большие и разноногие, и часто имеют связанный с ними след тельсона вдоль средней линии (как в случае со следом шотландского Hibbertopterus ). Такие пути были обнаружены на всех континентах, кроме Южной Америки. В некоторых местах, где ископаемые останки эвриптерид в остальном редки, например, в Южной Африке и остальной части бывшего суперконтинента Гондвана , открытия следов как предшествуют, так и превосходят численность окаменелостей тел эвриптерид. [23] Следы эвриптерид отнесены к нескольким ихнородам, в первую очередь к Palmichnium (определяемому как серия из четырех следов, часто с соответствующим знаком перетаскивания посередине), [24] при этом голотип ихновида P. kosinkiorum сохраняет самые большие из известных на сегодняшний день следов эвриптерид, каждый из которых имеет диаметр около 7,6 см (3,0 дюйма). [25] Другие ихнороды эвриптерид включают Merostomichnites (хотя вполне вероятно, что многие экземпляры на самом деле представляют собой следы ракообразных) и Arcuites (которые сохраняют бороздки, оставленные плавательными придатками). [24] [26] [27]
Дыхание
[ редактировать ]
У эвриптерид органы дыхания располагались на вентральной стенке тела (нижняя сторона опистосомы). Blattfüsse , развившийся из опистосомальных придатков, покрыл нижнюю сторону и создал жаберную камеру, где располагались «жаберные тракты». [2] В зависимости от вида жаберный тракт эвриптерид имел треугольную или овальную форму и, возможно, был приподнят в подушкообразное состояние. На поверхности этого жаберного тракта было несколько шипов (маленьких шипов), что привело к увеличению площади поверхности. Он состоял из губчатой ткани из-за множества впячиваний в структуру. [28]
Хотя Кименплатте называют «жаберным трактом», возможно, он не обязательно функционировал как настоящие жабры. У других животных жабры используются для поглощения кислорода из воды и являются выростами стенок тела. Несмотря на то, что эвриптериды явно были преимущественно водными животными, которые почти наверняка развились под водой (некоторые эвриптериды, такие как птериготиды, были бы даже физически неспособны ходить по суше), маловероятно, что «жаберный тракт» содержал функциональные жабры, если сравнивать этот орган с жабрами. у других беспозвоночных и даже рыб. Предыдущие интерпретации часто идентифицировали «жабры» эвриптерид как гомологичные жабрам других групп (отсюда и терминология), с газообменом, происходящим внутри губчатого тракта, и с рисунком жаберно-сердечных и дендритных вен (как в родственных группах), несущих насыщенную кислородом кровь в тело. Основной аналогией, использованной в предыдущих исследованиях, были мечехвосты, хотя их жаберное строение отличается от строения эвриптерид. У мечехвостов жабры более сложны и состоят из множества пластинок (пластинок), которые обеспечивают большую площадь поверхности, используемой для газообмена. Кроме того, жаберный тракт эвриптерид пропорционально слишком мал, чтобы поддерживать их, если он аналогичен жабрам других групп. Чтобы быть функциональными жабрами, они должны были быть высокоэффективными и иметь высокоэффективную систему кровообращения. Однако считается маловероятным, что этих факторов будет достаточно, чтобы объяснить большое несоответствие между размером жаберного тракта и размером тела. [29]
Вместо этого было высказано предположение, что «жаберный тракт» был органом дыхания воздухом, возможно, на самом деле являвшимся легким , пластроном или псевдотрахеей . [30] Пластроны — это органы, которые у некоторых членистоногих развились вторично, чтобы дышать воздухом под водой. Это считается маловероятным объяснением, поскольку эвриптериды с самого начала развивались в воде, и у них не было органов, развившихся из имеющихся органов воздушного дыхания. Кроме того, пластроны обычно обнажены на внешних частях тела, тогда как жаберный тракт эвриптерид расположен позади Блаттфюссена . [31] Вместо этого среди органов дыхания членистоногих жаберные тракты эвриптерид наиболее похожи на псевдотрахеи, обнаруженные у современных изопод . Эти органы, называемые псевдотрахеями, из-за некоторого сходства с трахеями (дыхательными трубками) организмов, дышащих воздухом, похожи на легкие и расположены внутри плеопод (задних ног) равноногих. Строение псевдотрахеи сравнивают с губчатой структурой жаберных трактов эвриптерид. Возможно, оба органа функционировали одинаково. [32]
Некоторые исследователи предположили, что эвриптериды могли быть адаптированы к амфибийному образу жизни, используя всю структуру жаберного тракта в качестве жабр, а впячивания внутри него - в качестве псевдотрахеи. Однако такой образ жизни мог быть физиологически невозможен, поскольку давление воды могло бы привести к попаданию воды в инвагинации, что привело бы к удушью . Более того, большинство эвриптерид всю свою жизнь вели водный образ жизни. Независимо от того, сколько времени они провели на суше, органы дыхания в подводной среде должны были присутствовать. Настоящие жабры, которые, как ожидается, располагались внутри жаберной камеры Блаттфюссена , у эвриптерид остаются неизвестными. [32]
Онтогенез
[ редактировать ]
Как и все членистоногие, эвриптериды созревали и росли, проходя статические стадии развития, называемые возрастами . Эти возрасты перемежались периодами, в течение которых у эвриптерид происходило шелушение (линька кутикулы), после чего они подвергались быстрому и немедленному росту. Некоторые членистоногие, например насекомые и многие ракообразные, в процессе взросления претерпевают резкие изменения. Хелицераты, включая эвриптериды, обычно считаются непосредственными разработчиками, не претерпевающими серьезных изменений после вылупления (хотя , в ходе онтогенеза у некоторых линий, таких как меченосцы и морские пауки могут появиться дополнительные сегменты тела и дополнительные конечности ). В прошлом были спорными вопросы о том, были ли эвриптериды настоящими прямыми разработчиками (вылупившиеся птенцы более или менее идентичны взрослым особям) или гемианаморфными прямыми разработчиками (с возможным добавлением дополнительных сегментов и конечностей во время онтогенеза). [33]
Гемианаморфное прямое развитие наблюдалось у многих групп членистоногих, таких как трилобиты , мегахейраны , базальные ракообразные и базальные многоножки . Настоящее прямое развитие иногда называют чертой, уникальной для паукообразных . Исследований онтогенеза эвриптерид было проведено немного, поскольку в летописи окаменелостей в целом не хватает экземпляров, которые можно с уверенностью утверждать, что они представляют собой молодь. [33] Вполне возможно, что многие виды эвриптерид, которые считались отдельными, на самом деле представляют собой молодые экземпляры других видов, при этом палеонтологи редко учитывают влияние онтогенеза при описании новых видов. [34]
Исследования хорошо сохранившегося комплекса окаменелостей эвриптерид из пражского возраста формации Медвежий Бьютт в каньоне Коттонвуд , штат Вайоминг , состоящего из множества экземпляров различных стадий развития эвриптерид Jaekelopterus и Strobilopterus , показали, что онтогенез эвриптерид был более или менее параллельным и схожим с у вымерших и современных мечевидных, за исключением того, что эвриптериды вылупились с полным набором придатков и сегментов опистосом. Таким образом, эвриптериды были не прямыми гемианаморфными разработчиками, а настоящими прямыми разработчиками, как современные паукообразные. [35]
Наиболее часто наблюдаемым изменением, происходящим в онтогенезе (за исключением некоторых родов, таких как Eurypterus , которые, по-видимому, были статичными), является пропорциональное уменьшение ширины метастомы. Это онтогенетическое изменение наблюдалось у членов нескольких надсемейств, таких как Eurypteroidea, Pterygotioidea и Moselopteroidea . [36]
Кормление
[ редактировать ]
Содержимое ископаемого кишечника эвриптерид неизвестно, поэтому прямых доказательств их диеты нет. Биология эвриптерид особенно указывает на плотоядный образ жизни. Многие из них были не только крупными (в общем, большинство хищников, как правило, крупнее своей добычи), но и обладали стереоскопическим зрением (способностью воспринимать глубину). [37] Ноги многих эвриптерид были покрыты тонкими шипами, которые использовались как для передвижения, так и для сбора пищи. В некоторых группах эти колючие придатки стали сильно специализированными. У некоторых эвриптерид Carcinosomatoidea обращенные вперед придатки были большими и имели чрезвычайно удлиненные шипы (как у Mixopterus и Megalograptus ). У производных представителей Pterygotioidea придатки были полностью лишены шипов, а вместо этого имели специальные когти. [38] Другие эвриптериды, лишенные этих специализированных придатков, вероятно, питались аналогично современным мечехвостам, хватая и измельчая пищу придатками, а затем заталкивая ее в рот с помощью хелицер. [39]
Окаменелости, сохраняющие пищеварительный тракт, были обнаружены в окаменелостях различных эвриптерид, в том числе Carcinosoma , Acutiramus и Eurypterus . Хотя сообщалось о потенциальном анальном отверстии в тельсоне экземпляра Buffalopterus , более вероятно, что анус открывался через тонкую кутикулу между последним сегментом перед тельсоном и самим тельсоном, как у современных мечехвостов. [37]
Копролиты эвриптерид, обнаруженные в отложениях ордовикского возраста в Огайо, содержащие фрагменты трилобита и эвриптерида Megalograptus ohioensis в сочетании с полными экземплярами того же вида эвриптерид, как предполагается, представляют собой свидетельство каннибализма . Подобные копролиты, относящиеся к виду Lanarkopterus dolichoschelus из ордовика штата Огайо, содержат фрагменты бесчелюстных рыб и фрагменты более мелких экземпляров самого Lanarkopterus . [37]
Хотя роль высших хищников была бы ограничена самыми крупными эвриптеридами, более мелкие эвриптериды, вероятно, сами по себе были грозными хищниками, как и их более крупные родственники. [7]
Репродуктивная биология
[ редактировать ]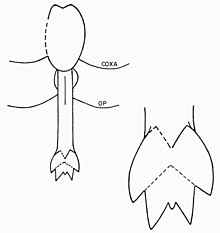

Как и во многих других полностью вымерших группах, понимание и исследование размножения и полового диморфизма эвриптерид затруднено, поскольку они известны только по окаменевшим панцирям и панцирям. В некоторых случаях очевидных различий может быть недостаточно для разделения полов только на основе морфологии. [17] Иногда два пола одного и того же вида интерпретировались как два разных вида, как это было в случае с двумя видами Drepanopterus ( D. bembycoides и D. lobatus ). [40]
Просома эвриптерид состоит из первых шести сегментов экзоскелета, слитых вместе в более крупную структуру. Седьмой сегмент (таким образом, первый сегмент опистосомы) называется метастомой , а восьмой сегмент (отчетливо пластинчатый) называется жаберной крышкой и содержит половое отверстие. Нижнюю сторону этого сегмента занимает половая крышка — структура, первоначально развившаяся из наследственных седьмой и восьмой пары придатков. В его центре, как и у современных мечехвостов, находится половой придаток. Этот придаток, удлиненный стержень с внутренним протоком, встречается в двух различных морфах, обычно называемых «типом А» и «типом B». [17] Эти генитальные придатки часто сохраняются в окаменелостях и были предметом различных интерпретаций размножения эвриптерид и полового диморфизма. [41]
Придатки типа A обычно длиннее, чем придатки типа B. У некоторых родов они разделены на разное количество секций, например, у Eurypterus , где придаток типа A разделен на три, а придаток типа B только на две. [42] Такое деление полового придатка часто встречается у эвриптерид, но их численность не универсальна; например, придатки обоих типов в семействе Pterygotidae неразделены. [43] Придаток типа А также вооружен двумя изогнутыми шипами, называемыми фурка (буквально «вилка» на латыни). Также возможно наличие фурки в придатке типа Б, структура может представлять собой несросшиеся кончики придатков. Между дорсальной и вентральной поверхностями Blattfüsse , связанным с придатками типа А, расположен набор органов, традиционно описываемых как «трубчатые органы» или «роговые органы». Эти органы чаще всего интерпретируются как сперматеки (органы хранения спермы ), хотя эта функция еще окончательно не доказана. [44] У членистоногих сперматеки используются для хранения сперматофора, полученного от самцов. Это означало бы, что придаток типа А является женской морфой, а придаток типа В — мужской. [17] Дополнительным доказательством того, что придатки типа А представляют женскую морфию генитальных придатков, является их более сложная конструкция (общая тенденция для гениталий самок членистоногих). Возможно, большая длина придатка типа А означает, что он использовался как яйцеклад (служил для откладывания яиц). [45] Различные типы генитальных придатков не обязательно являются единственной особенностью, которая различает полы эвриптерид. В зависимости от рассматриваемого рода и вида другие особенности, такие как размер, количество орнаментов и пропорциональная ширина тела, могут быть результатом полового диморфизма. [2] В целом эвриптериды с придатками типа B (самцы), по-видимому, были пропорционально шире, чем эвриптериды с придатками типа A (самки) того же рода. [46]

Основная функция длинных, предполагаемых женских придатков типа А, вероятно, заключалась в переносе сперматофора из субстрата в репродуктивный тракт, а не в том, чтобы служить яйцекладом, поскольку яйцеклады членистоногих обычно длиннее, чем придатки эвриптерид типа А. Вращая стороны жаберной крышки, можно было бы опустить придаток от тела. Из-за того, что в месте его расположения накладываются разные пластинки, придаток был бы невозможен для перемещения без мышечных сокращений, движущихся вокруг жаберной крышки. Он бы оставался на месте, когда не использовался. Вилка на придатках типа А могла способствовать раскрытию сперматофора и высвобождению свободных сперматозоидов внутри для поглощения. Считается, что «роговые органы», возможно, сперматеки, были связаны непосредственно с придатком через тракты, но эти предполагаемые тракты остаются несохранившимися в доступном ископаемом материале. [47]
Придатки типа B, предположительно мужские, должны были производить, хранить и, возможно, формировать сперматофор в форме сердца на дорсальной поверхности придатка. Широкое генитальное отверстие позволило бы высвободить большое количество сперматофоров за один раз. Длинная фурка, связанная с придатками типа B, возможно, способная опускаться, как придаток типа A, могла быть использована для определения того, подходит ли субстрат для отложения сперматофоров. [48]
Эволюционная история
[ редактировать ]Происхождение
[ редактировать ]
До 1882 года не было известно никаких эвриптерид до силура. Современные открытия, сделанные с 1880-х годов, расширили знания о ранних эвриптеридах ордовикского периода . [49] Самые ранние эвриптериды, известные сегодня, мегалограптид Pentecopterus , датируются дарривильским этапом среднего ордовика, 467,3 миллиона лет назад . [50] Есть также сообщения о еще более ранних ископаемых эвриптеридах в биоте Фезуаты позднетремадокийского ( раннего ордовика) возраста в Марокко , но они еще не тщательно изучены. [51] и, вероятно, представляют собой придатки пейтоид .
Pentecopterus был относительно производным эвриптеридом, частью семейства мегалограптид внутри надсемейства карцинозоматоидных. Его производное положение предполагает, что большинство клад эвриптерид, по крайней мере, в пределах подотряда эвриптеринов, уже сформировались к этому моменту в среднем ордовике. [52] Самая ранняя из известных стилонуриновых эвриптерид Brachyopterus . [7] По возрасту также относится к среднему ордовику. Присутствие представителей обоих подотрядов указывает на то, что им предшествовали примитивные стеблевые эвриптериды, хотя они до сих пор неизвестны в летописи окаменелостей. Присутствие нескольких клад эвриптерид в среднем ордовике позволяет предположить, что эвриптериды либо возникли в раннем ордовике и испытали быструю и взрывную радиацию и диверсификацию вскоре после того, как появились первые формы, либо что группа возникла намного раньше, возможно, в кембрийский период. [52]
Таким образом, точное время происхождения эвриптерид остается неизвестным. Хотя окаменелости, называемые «примитивными эвриптеридами», иногда описывались из отложений кембрийского или даже докембрийского периода, [53] сегодня они не признаются эвриптеридами, а иногда даже как родственные формы. Некоторые животные, ранее считавшиеся примитивными эвриптеридами, например, род Strabops из кембрия штата Миссури , [54] теперь классифицируются как агласпидиды или страбопиды . Агласпидиды, когда-то считавшиеся примитивными хелицератами, теперь рассматриваются как группа, более тесно связанная с трилобитами. [55]
Летопись окаменелостей ордовикских эвриптерид весьма скудна. Большинство эвриптерид, когда-то известных из ордовика, с тех пор оказались ошибочными идентификациями или псевдоокаменелостями . Сегодня только 11 видов можно с уверенностью отнести к ордовикским эвриптеридам. Эти таксоны делятся на две отдельные экологические категории; крупные и активные хищники с древнего континента Лаврентия , а также демерсальные (живущие на морском дне ) и базальные животные с континентов Авалония и Гондвана. [50] Лаврентийские хищники, отнесенные к семейству Megalograptidae (включающее роды Echinognathus , Megalograptus и Pentecopterus ), вероятно, представляют собой первую по-настоящему успешную группу эвриптерид, испытавшую небольшую радиацию в позднем ордовике. [56]
силурийский
[ редактировать ]
Эвриптериды были наиболее разнообразны и многочисленны между средним силуром и ранним девоном, с абсолютным пиком разнообразия в эпоху Придоли , от 423 до 419,2 миллионов лет назад, самого позднего силура. [16] Этот пик разнообразия был признан с начала двадцатого века; из примерно 150 видов эвриптерид, известных в 1916 г., более половины были из силура, а треть - только из позднего силура. [49]
Хотя стилонуриновые эвриптериды в целом оставались редкими и малочисленными, как это было в предшествующий ордовик, эвриптериновые эвриптериды испытали быстрый рост разнообразия и численности. [57] В большинстве силурийских ископаемых пластов эвриптериновые эвриптериды составляют 90% всех присутствующих эвриптерид. [58] Хотя некоторые из них, вероятно, уже присутствовали в позднем ордовике (пока просто отсутствуют в летописи окаменелостей), [52] подавляющее большинство групп эвриптерид впервые отмечается в слоях силурийского возраста. К ним относятся как группы стилонуринов, такие как Stylonuroidea , Kokomopteroidea и Mycteropoidea , так и группы эвриптеринов, такие как Pterygotioidea, Eurypteroidea и Waeringopteroidea . [59]
Самым успешным эвриптеридом на сегодняшний день был Eurypterus среднего и позднего силура , универсал , который с одинаковой вероятностью занимался хищничеством или падальщиком . Считается, что они охотились в основном на мелких и мягкотелых беспозвоночных, таких как черви . [60] Виды этого рода (из которых наиболее распространенным является типовой вид E. remipes ) составляют более 90% (возможно, до 95%) всех известных ископаемых экземпляров эвриптерид. [58] Несмотря на свою огромную численность, Eurypterus известны только в относительно коротком временном диапазоне: впервые они появились в эпоху позднего Лландовери (около 432 миллионов лет назад) и вымерли к концу эпохи Придоли. [61] Eurypterus также был ограничен континентом Еврамерика (состоящим из экваториальных континентов Авалония, Балтика и Лаврентия), который был полностью колонизирован этим родом во время его слияния и не мог пересекать обширные океанские просторы, отделявшие этот континент от других частей континента. мире, например, южный суперконтинент Гондвана. Таким образом, Eurypterus был географически ограничен береговой линией и мелководными внутренними морями Еврамерики. [58] [62]
В позднем силуре появились крыловидные эвриптериды, крупные и специализированные формы с несколькими новыми приспособлениями, такими как большие и уплощенные тельсоны, способные использоваться в качестве рулей, и крупные и специализированные хелицеры с увеличенными клешнями для захвата (а в некоторых случаях и убийства) добычи. . [3] [4] Хотя самые крупные представители семейства появились в девоне, крупные двухметровые (6,5+ футов) птериготиды, такие как Acutiramus, уже присутствовали в позднем силуре. [10] Их экология варьировалась от общего хищнического поведения до нападения хищников из засад , а некоторые, такие как сам Pterygotus , были активными высшими хищниками в морских экосистемах позднего силура. [63] Птериготиды, очевидно, также были способны пересекать океаны, став одной из двух групп эвриптерид, достигших космополитического распространения . [64]
девонский период
[ редактировать ]
Хотя эвриптериды продолжали быть многочисленными и разнообразными в течение раннего девона (например, что привело к эволюции крыловидного Jaekelopterus , крупнейшего из всех членистоногих), эта группа была одной из многих, сильно пострадавших от позднего девонского вымирания . Вымирание, о котором известно, что оно затронуло только морскую жизнь (особенно трилобитов, брахиопод и организмы, строящие рифы ), фактически подорвало численность и разнообразие, ранее наблюдавшиеся среди эвриптерид. [65]
Значительное снижение разнообразия началось уже в раннем девоне, а к позднему девону эвриптериды стали редко встречаться в морской среде. На франском этапе вымерли четыре семьи, а на более позднем фаменском этапе вымерли еще пять семей. [65] Поскольку больше всего пострадали морские группы, эвриптериды в первую очередь пострадали в пределах подотряда эвриптеринов. Лишь одна группа стилонуринов (семейство Parastylonuridae ) вымерла в раннем девоне. До позднего девона вообще сохранились только два семейства эвриптерин ( Adelophthalmidae и Waeringopteridae). Наибольшее сокращение эвриптеринов произошло в раннем девоне, во время которого более 50% их разнообразия было потеряно всего за 10 миллионов лет. Стилонурины, с другой стороны, сохранялись в течение всего периода с более или менее постоянным разнообразием и численностью, но были затронуты в позднем девоне, когда многие из более старых групп были заменены новыми формами в семействах Mycteroptidae и Hibbertopteridae. [66]
Вполне возможно, что катастрофические закономерности вымирания, наблюдаемые в подотряде эвриптеринов, были связаны с появлением более производных рыб. Снижение эвриптерина началось в тот момент, когда бесчелюстные рыбы впервые стали более развитыми, и это совпадает с появлением плакодерм (бронированных рыб) как в Северной Америке, так и в Европе. [67]
Стилонурины выживших семейств гиббертоптеридов и миктероптид полностью избежали конкуренции с рыбами, эволюционировав в сторону новой и отдельной экологической ниши. Эти семейства пережили радиацию и диверсификацию в позднем девоне и раннем каменноугольном периоде, последнюю радиацию среди эвриптерид, которая привела к появлению нескольких новых форм, способных «подметать» (рыскать по субстрату в поисках добычи). [68]
Карбон и Пермь
[ редактировать ]
Только три семейства эвриптерид — Adelophthalmidae, Hibbertopteridae и Mycteroptidae — полностью пережили вымирание. Раньше мы думали, что все это пресноводные животные, что привело бы к исчезновению эвриптерид в морской среде. [65] а с исчезновением морских хищников-эвриптерид саркоптериги , такие как ризодонты , стали новыми высшими хищниками в морской среде. [67] Однако различные недавние находки вызывают сомнения по этому поводу и позволяют предположить, что эти эвриптериды были эвригалинными формами, обитавшими в окраинных морских средах, таких как устья рек, дельты, лагуны и прибрежные пруды. Один аргумент палеобиогеографический; Распространение крыловидных особей, по-видимому, требует распространения в океане. [69] В недавнем обзоре Adelophthalmoidea было признано, что «во многих секциях, дающих Adelophthalmus, наблюдается гораздо большее морское влияние , чем считалось ранее». [70] Аналогичным образом, исследование эвриптерид Hibbertopterus из каменноугольного периода Нью-Мексико пришло к выводу, что среда обитания некоторых эвриптерид «возможно, потребует переоценки». [71] Единственное сохранившееся семейство эвриптеринов, Adelophthalmidae, было представлено только одним родом, Adelophthalmus . Гиббертоптериды, миктероптиды и аделофтальмусы дожили до перми. [72]
Adelophthalmus стал наиболее распространенным из всех позднепалеозойских эвриптерид, существовавшим в большем количестве и разнообразии, чем сохранившиеся стилонурины, и диверсифицированным в отсутствие других эвриптеринов. [73] Из 33 видов, отнесенных к Adelophthalmus , 23 (69%) происходят только из каменноугольного периода. [74] [75] Пика разнообразия род достиг в позднем карбоне. Хотя Adelophthalmus уже был относительно широко распространен и представлен на всех основных участках суши в позднем девоне, объединение Пангеи в глобальный суперконтинент в течение последних двух периодов палеозоя позволило Adelophthalmus получить почти всемирное распространение. [58]
В позднем карбоне и ранней перми Adelophthalmus был широко распространен и обитал преимущественно в солоноватых и пресноводных средах, прилегающих к прибрежным равнинам. Эти условия поддерживались благоприятными климатическими условиями. Они не сохранились, поскольку изменения климата, вызванные образованием Пангеи, изменили модели осадконакопления и растительности по всему миру. С исчезновением среды обитания численность Adelophthalmus сократилась, и они уже вымерли к леонардскому этапу ранней перми. [76]
Миктероптиды и гиббертоптериды продолжали выживать в течение некоторого времени, при этом из пермских слоев был известен по одному роду каждой группы: Hastimima и Campylocephalus соответственно. [77] Хастимима вымерла в ранней перми. [78] как это было у Adelophthalmus , тогда как Campylocephalus сохранялся дольше. Массивный неполный панцирь из пермских отложений в России представляет собой единственные ископаемые останки вида C. permianus , длина которого могла достигать 1,4 метра (4,6 фута), хотя первоначально считалось, что это позднепермский возраст. [10] другие источники предполагают кунгурско-роудский возраст экземпляра. [79] Фрагмент кутикулы эвриптерид, получивший видовое название Woodwardopterus freemanorum, известен из конечной перми Австралии и представляет собой самую молодую из известных эвриптерид. [79] Эвриптериды из ископаемых слоев выше перми не известны. Это указывает на то, что последние эвриптериды погибли либо в результате катастрофического вымирания в его конце, либо в какой-то момент незадолго до него. Это событие вымирания, пермско-триасовое вымирание , является самым разрушительным из зарегистрированных массовых вымираний и привело к вымиранию многих других успешных палеозойских групп, таких как трилобиты. [80]
История обучения
[ редактировать ]
Первый известный экземпляр эвриптерид был обнаружен в силурийских породах Нью-Йорка , которые по сей день являются одним из самых богатых мест ископаемых эвриптерид. Сэмюэл Л. Митчилл описал экземпляр, обнаруженный недалеко от Уэстморленда в графстве Онейда в 1818 году. Он ошибочно определил ископаемое как образец рыбы Silurus , вероятно, из-за странного, похожего на сома внешнего вида панциря. Семь лет спустя, в 1825 году, Джеймс Э. ДеКей исследовал окаменелость и признал, что она явно принадлежит членистоногому. Он считал, что ископаемое, которое он назвал Eurypterus remipes , представляет собой ракообразного отряда Branchiopoda , и предположил, что оно может представлять собой недостающее звено между трилобитами и более развитыми жабероподами. [81] Название Eurypterus происходит от греческого eurús ( εὐρύς «широкий, широкий») и птерона ( πτερόν «крыло»). [82]
В 1843 году Герман Бурмейстер опубликовал свой взгляд на таксономию трилобитов и на то, как эта группа связана с другими организмами, живыми и вымершими, в работе Die Organization der Trilobiten aus ihren lebenden Verwandten entwickelt . Он считал трилобитов ракообразными, как и предыдущие авторы, и классифицировал их вместе с тем, что он считал их ближайшими родственниками, Eurypterus и родом Cytherina , в кладе, которую он назвал «Palaeadae». В Палеадах Бурмейстер основал три семьи; «Trilobitae» (состоящие из всех трилобитов), «Cytherinidae» (состоящие только из Cytherina , животного, которое сегодня рассматривается как остракод ) и Eurypteridae (состоящие из Eurypterus , включавшие в то время три вида). [83]
Четвертым описанным родом эвриптерид (после Hibbertopterus в 1836 году и Campylocephalus в 1838 году, который был идентифицирован как эвриптериды лишь позднее), из тех, которые все еще считаются таксономически действительными в наше время, был Pterygotus ( букв. « крылатый » ), описанный Луи Агассис в 1839 году. [84] Pterygotus был значительно больше по размеру, чем Eurypterus , и когда каменоломни в Шотландии прозвали их « серафимами обнаружили первые окаменелости , рабочие ». Агассис сначала подумал, что окаменелости представляют собой останки рыб, но признал их природу членистоногих только пять лет спустя, в 1844 году. [85]
В 1849 году Фредерик Маккой классифицировал Pterygotus вместе с Eurypterus и Belinurus (род, который сегодня рассматривается как xiphosuran) в Eurypteridae Бурмейстера. Маккой считал Eurypteridae группой ракообразных отряда Entomostraca , тесно связанных с мечехвостами. [86] Четвертый род, Slimonia , основанный на ископаемых останках, ранее отнесенных к новому виду Pterygotus , был отнесен к Eurypteridae в 1856 году Дэвидом Пейджем . [87]

Яна Нешковского В книге «De Euryptero Remipede» (1858 г.) содержится подробное описание Eurypterus fischeri (теперь рассматриваемого как синоним другого вида Eurypterus , E. Tetragonophthalmus ), которое, наряду с монографией «О роде Pterygotus» Томаса Генри Хаксли и Джона Уильяма Солтера , и исчерпывающее описание различных эвриптерид Нью-Йорка в третьем томе «Палеонтологии Нью-Йорка » (1859 г.), сделанное Джеймсом Холлом , внесло огромный вклад в понимание разнообразия и биологии эвриптерид. Эти публикации были первыми, кто полностью описал всю анатомию эвриптерид, признав полное количество просомальных придатков, а также количество преабдоминальных и постабдоминальных сегментов. И Нешковский, и Холл признали, что эвриптериды были тесно связаны с современными хелицератами, такими как мечехвосты. [88]
В 1865 году Генри Вудворд описал род Stylonurus (названный и изображенный, но не полностью описанный Дэвидом Пейджем в 1856 году) и повысил ранг Eurypteridae до порядка, фактически создав Eurypterida как таксономическую единицу, которую он воспринимает сегодня. . [89] В работе «Анатомия и взаимоотношения эвриптерид» (1893) Малкольм Лори значительно расширил знания и обсуждение анатомии и взаимоотношений эвриптерид. Он сосредоточился на том, как эвриптериды связаны друг с другом, а также с трилобитами, ракообразными, скорпионами, другими паукообразными и мечехвостами. Описание Eurypterus fischeri Герхардом Хольмом в 1896 году было настолько подробным, что этот вид стал одним из наиболее полностью известных из всех вымерших животных, настолько, что знания об E. fischeri были сопоставимы со знаниями о его современных родственниках (таких как Атлантический мечехвост ). Описание также помогло укрепить тесную связь между эвриптеридами и другими хелицератами, продемонстрировав многочисленные гомологии между двумя группами. [90]
В 1912 году Джон Мейсон Кларк и Рудольф Рюдеманн опубликовали книгу «Эвриптериды Нью-Йорка» , в которой обсуждались все виды эвриптерид, обнаруженные на данный момент из ископаемых отложений. Кларк и Рюдеман создали одно из первых филогенетических деревьев эвриптерид, разделив отряд на два семейства; Eurypteridae (отличаются гладкими глазами и включают Eurypterus , Anthraconectes , Stylonurus , Eusarcus , Dolichopterus , Onychopterus и Drepanopterus ) и Pterygotidae (отличаются фасеточными глазами, включая Pterygotus , Erettopterus , Slimonia и Hughmilleria ). Считалось, что обе семьи произошли от общего предка Страбопса . [91] Как и более ранние авторы, Кларк и Рюдеманн также поддержали тесную связь между эвриптеридами и мечехвостами (объединенными в класс Merostomata), но также обсудили альтернативные гипотезы, такие как более близкое родство с паукообразными. [92]
Классификация
[ редактировать ]

Исторически сложилось так, что большинство исследователей предполагали тесную связь между эвриптеридами и мечехвостами (такими как современный атлантический мечехвост). Некоторые гомологии подтверждают эту точку зрения, например, корреляция сегментов придатков и просомы. Кроме того, наличие пластинчатых придатков, несущих «жаберные тракты» на придатках опистосомы ( Blattfüssen ), ранее упоминалось как важная гомология. В последние несколько десятилетий девятнадцатого века были установлены дальнейшие гомологии, такие как схожие структуры сложных глаз Pterygotus и мечехвостов (что считалось особенно важным, поскольку считалось, что глаз мечехвоста обладает почти уникальной структурой). и сходства в онтогенезе внутри обеих групп. [93] Это онтогенетическое сходство было наиболее очевидным при изучении непионических стадий (стадия развития, следующая сразу за эмбриональной стадией) в обеих группах, во время которых как меченосцы, так и эвриптериды имеют пропорционально больший панцирь, чем взрослые особи, обычно шире, обладают отчетливым гребнем вниз. средние имеют меньшее количество сегментов, лишенных дифференцировки, со слаборазвитым тельсоном. [94]
Из-за этого сходства мечевидные и эвриптериды часто объединялись в один класс или подкласс, называемый Merostomata (выведенный Генри Вудвордом в 1866 году для размещения обеих групп). Хотя меченосцы (как и эвриптериды) исторически считались ракообразными из-за их дыхательной системы и водного образа жизни, эта гипотеза была дискредитирована после того, как были обнаружены многочисленные сходства между мечехвостами и паукообразными. [94] Некоторые авторы, такие как Джон Стерлинг Кингсли в 1894 году, классифицировали Merostomata как сестринскую группу паукообразных в классе «Acerata» в подтипе «Branchiata». Другие, такие как Рэй Ланкестер в 1909 году, пошли еще дальше и классифицировали Merostomata как подкласс паукообразных, возведенный в ранг класса. [95]
В 1866 году Эрнст Геккель классифицировал Merostomata (содержащий практически только Eurypterida) и Xiphosura в группу ракообразных, которую он назвал Gigantostraca. Хотя Геккель не обозначил никакого таксономического ранга для этой клады, более поздние исследователи, такие как Джон Стерлинг Кинсгсли, интерпретировали его как эквивалент ранга подкласса, такого как Malacostraca и Entomostraca. [96] В последующих исследованиях Gigantostraca рассматривалась как синоним Merostomata (редко) и самой Eurypterida (чаще). [97] [98] Филогенетический анализ (результаты представлены на кладограмме ниже), проведенный Джеймсом Ламсделлом в 2013 году, по взаимоотношениям внутри Xiphosura и отношениям с другими близкородственными группами (включая эвриптериды, которые были представлены в анализе родами Eurypterus , Parastylonurus , Rhenopterus) . и Stoermeropterus ) пришли к выводу, что Xiphosura, как это понимается в настоящее время, была парафилетической (группа, имеющая последнего общего предка , но не включающая всех потомков этого предка) и, следовательно, не являющаяся действительной филогенетической группой. [99] Эвриптериды были обнаружены как тесно связанные с паукообразными, а не с ксифосуранами, образуя группу Sclerophorata в кладе Dekatriata (состоящей из склерофоратов и хасмаспидид ). Ламсделл отметил, что вполне возможно, что Dekatriata является синонимом Sclerophorata, поскольку репродуктивная система, основная определяющая особенность склерофоратов, не была тщательно изучена у хазматаспидид. Dekatriata, в свою очередь, является частью Prosomapoda , группы, включающей Xiphosurida (единственная монофилетическая группа ксифосуранов) и других стеблевых родов. [100]
| Членистоногие |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Внутренние отношения
[ редактировать ]

Внутренняя классификация эвриптерид внутри Eurypterida основана в основном на одиннадцати установленных признаках. Они использовались на протяжении всей истории исследований эвриптерид для установления клад и родов. К этим признакам относятся: форма просомы, форма метастомы, форма и положение глаз, типы просомальных придатков, типы плавательных ластов ног, строение дублюры (бахромы спинного экзоскелета). , строение опифосомы, строение половых придатков, форма тельсона и тип орнаментации. Стоит отметить, что не все эти признаки имеют одинаковое таксономическое значение. [101] Они также применимы не ко всем эвриптеридам; стилонуриновые эвриптериды полностью лишены лопастей для плавательных ног. [16] Некоторые признаки, в том числе форма просомы и метастомы, а также положение и форма глаз, считаются важными только для различия между разными родами. [102] Большинство надсемейств и семей определяются на основе морфологии придатков. [103]
Наиболее важным признаком, используемым в систематике эвриптерид, является тип просомальных придатков, поскольку этот признак используется для определения целых подотрядов. Общая анатомия ног также может использоваться для определения суперсемейств и семейств. Исторически хелицеры считались наиболее важными придатками с таксономической точки зрения, поскольку они встречались только в двух основных типах: эвриптеридном типе с маленькими и беззубыми клешнями и крыловидном типе с большими клешнями и зубами. Это различие исторически использовалось для разделения Eurypterida на два подотряда Eurypterina (маленькие хелицеры) и «Pterygotina» (большие и мощные хелицеры). [104] Эта классификационная схема не лишена проблем. В таксономическом пересмотре Eurypterida, проведенном Виктором Толлертоном в 1989 году, с признанием подотрядов Eurypterina и Pterygotina, несколько клад эвриптерид, сегодня признанных стилонуринами (включая гиббертоптериды и миктероптиды), были переклассифицированы как неэвриптериды в новый отдельный отряд «Cyrtoctenida» на основании предполагаемого несоответствия в просомальных придатках. [105]
Современные исследования отдают предпочтение классификации на подотряды Eurypterina и Stylonurina, что подтверждается филогенетическим анализом. [106] [36] В частности, птериготидные эвриптериды имеют ряд гомологий с производными эвриптериновыми эвриптеридами, такими как аделофтальмиды, и поэтому их лучше всего классифицировать как производных членов того же подотряда. [107] У Stylonurina шестая пара придатков представлена длинными и тонкими ходильными ногами и не имеет видоизмененного позвоночника (называемого подомером 7а). У большинства эвриптерид Eurypterina шестая пара придатков расширена в плавательные лопасти и всегда имеет подомер 7а. 75% видов эвриптерид являются эвриптеринами, и они составляют 99% всех ископаемых экземпляров эвриптерид. [16] Из всех клад эвриптерид Pterygotioidea является наиболее богатой видами: насчитывает более 50 видов. Второй по богатству видами кладой является Adelophthalmoidea, насчитывающая более 40 видов. [58]
Представленная ниже кладограмма, охватывающая все признанные в настоящее время семейства эвриптерид, основана на исследовании О. Эрика Тетли, проведенном в 2007 году. [108] Подотряд стилонуринов основан на исследовании 2010 года, проведенном Джеймсом Ламсделлом, Саймоном Дж. Брэдди и Тетли. [109] Надсемейство «Megalograptoidea», признанное Тетли в 2007 году и затем помещенное между Onychopterelloidea и Eurypteroidea, было опущено, поскольку более поздние исследования показывают, что мегалограптиды были членами надсемейства Carcinosomatoidea. Таким образом, филогения Carcinosomatoidea следует за исследованием Ламсделла и его коллег, проведенным в 2015 году. [110]
| Эвриптерида | |





См. также
[ редактировать ]- Список родов эвриптерид
- Nepidae — неродственное семейство насекомых, широко известное как «водяные скорпионы».
- Cottidae — семейство рыб, в общем названии некоторых представителей которого содержится слово «морской скорпион».
- История жизни
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ Стёрмер 1955 , стр. 23.
- ^ Jump up to: а б с д и ж г час я Брэдди и Данлоп 1997 , стр. 437–439.
- ^ Jump up to: а б Тетли и Бриггс 2009 , с. 1141.
- ^ Jump up to: а б Плотник и Баумиллер 1988 , с. 22.
- ^ Кларк и Рюдеманн 1912 , с. 244.
- ^ «Селективность в эволюции палеозойских групп членистоногих с акцентом на массовое вымирание и радиацию: филогенетический подход» .
- ^ Jump up to: а б с Тетли 2007 , стр. 557.
- ^ Пошманн и Тетли 2004 , стр. 189.
- ^ Jump up to: а б с д Брэдди, Пошманн и Тетли 2008 , стр. 107.
- ^ Jump up to: а б с Ламсделл и Брэдди 2009 , Дополнительная информация .
- ^ Бриггс 1985 , стр. 157–158.
- ^ Кьеллесвиг-Варинг 1961 , стр. 830.
- ^ Ламсделл и др. 2015 , с. 15.
- ^ Краус и Браукманн 2003 , стр. 5–50.
- ^ Тетли 2008 , стр. 19.
- ^ Jump up to: а б с д Тетли 2007 , стр. 559.
- ^ Jump up to: а б с д Палеос .
- ^ Уайт 2005 , с. 576.
- ^ Селден 1999 , с. 43.
- ^ Селден 1999 , с. 45.
- ^ Селден 1999 , стр. 44–46.
- ^ Ханкен и Стермер 1975 , стр. 262–267.
- ^ Брэдди и Алмонд 1999 , с. 166.
- ^ Jump up to: а б Брэдди и Алмонд 1999 , стр. 168–170.
- ^ Брезински и Коллар 2016 , с. 39.
- ^ Ханкен и Стермер 1975 , стр. 255.
- ^ Вразо и Чурка 2017 , с. 235.
- ^ Селден 1985 , с. 219.
- ^ Селден 1985 , стр. 220–221.
- ^ Селден 1985 , с. 221.
- ^ Селден 1985 , с. 222.
- ^ Jump up to: а б Селден 1985 , стр. 222–223.
- ^ Jump up to: а б Ламсделл и Селден 2013 , с. 32.
- ^ Ламсделл и Селден 2013 , с. 33.
- ^ Ламсделл и Селден 2013 , с. 44.
- ^ Jump up to: а б Ламсделл и Селден 2013 , с. 34.
- ^ Jump up to: а б с Селден 1999 , с. 46.
- ^ Селден 1999 , с. 47.
- ^ Хембри, Платт и Смит, 2014 , с. 77.
- ^ Ламсделл, Брэдди и Тетли 2009 , стр. 1119.
- ^ Брэдди и Данлоп 1997 , с. 436.
- ^ Брэдди и Данлоп 1997 , с. 438.
- ^ Брэдди, Пошманн и Тетли 2008 , стр. 108.
- ^ Брэдди и Данлоп 1997 , с. 439.
- ^ Брэдди и Данлоп 1997 , с. 449.
- ^ Ламсделл 2014 , стр. 175–177.
- ^ Брэдди и Данлоп 1997 , стр. 450–452.
- ^ Брэдди и Данлоп 1997 , стр. 454–455.
- ^ Jump up to: а б О'Коннелл 1916 , с. 11.
- ^ Jump up to: а б Ламсделл и др. 2015 , с. 1.
- ^ Ван Рой, Бриггс и Гейнс, 2015 , стр. 6.
- ^ Jump up to: а б с Ламсделл и др. 2015 , с. 29.
- ^ О'Коннелл 1916 , с. 12.
- ^ О'Коннелл 1916 , с. 13.
- ^ Ортега-Эрнандес, Легг и Брэдди, 2012 , с. 15.
- ^ Тетли 2007 , стр. 569.
- ^ Тетли 2007 , стр. 567.
- ^ Jump up to: а б с д и Тетли 2007 , стр. 570.
- ^ Данлоп, Пенни и Джекель, 2018 , стр. 17-30.
- ^ Селден 1999 , с. 44.
- ^ Тетли 2006 , стр. 410.
- ^ Тетли и Рабано 2007 , с. 124.
- ^ Маккой и др. 2015 , с. 3.
- ^ Тетли 2007 , стр. 571.
- ^ Jump up to: а б с Халлам и Виньял 1997 , с. 70.
- ^ Ламсделл и Брэдди 2009 , с. 265.
- ^ Jump up to: а б Ламсделл и Брэдди 2009 , с. 266.
- ^ Ламсделл и Брэдди 2009 , с. 268.
- ^ Тетли 2007 .
- ^ Тетли, О. Эрик; Пошманн, Маркус (1 января 2008 г.). «Филогения и палеоэкология Adelophthalmoidea (Arthropoda; Chelicerata; Eurypterida)» . Журнал систематической палеонтологии . 6 (2): 237–249. Бибкод : 2008JSPal...6..237T . дои : 10.1017/S1477201907002416 . ISSN 1477-2019 . S2CID 59488956 .
- ^ Брэдди, Саймон Дж.; Лернер, Аллан Дж; Лукас, Спенсер Г. (1 февраля 2023 г.). «Новый вид эвриптерид Hibbertopterus из каменноугольного периода Нью-Мексико и обзор Hibbertopteridae» . Историческая биология . 35 (2): 257–263. Бибкод : 2023HBio...35..257B . дои : 10.1080/08912963.2022.2032690 . ISSN 0891-2963 . S2CID 246819305 .
- ^ Данлоп, Пенни и Джекель, 2018 , стр. 19 и 24.
- ^ Тетли и Ван Рой 2006 , стр. 79.
- ^ Данлоп, Пенни и Джекель, 2018 , стр. 24.
- ^ Ламсделл и др. 2020 , с. 1.
- ^ Куес и Кицке 1981 , с. 727.
- ^ Данлоп, Пенни и Джекель, 2018 , стр. 19.
- ^ Уайт 1927 , с. 575.
- ^ Jump up to: а б Пошманн и Розефельдс 2021 .
- ^ Бергстром и Дугаткин 2012 , с. 515.
- ^ Кларк и Рюдеманн 1912 , с. 13.
- ^ Наддс и Селден 2008 , стр. 78–82.
- ^ Бурмейстер 1843 , стр. 62–64.
- ^ Данлоп, Пенни и Джекель, 2018 , стр. 27.
- ^ Кьеллесвиг-Варинг 1964 , стр. 331.
- ^ Маккой 1849 , с. 393.
- ^ Хендерсон 1866 , с. 18.
- ^ Кларк и Рюдеманн 1912 , с. 14.
- ^ Вудворд 1865 , стр. 484–486.
- ^ Кларк и Рюдеманн 1912 , с. 19.
- ^ Кларк и Рюдеманн 1912 , стр. 124–125.
- ^ Кларк и Рюдеманн 1912 , с. 135 и 137.
- ^ Кларк и Рюдеманн 1912 , с. 135.
- ^ Jump up to: а б Кларк и Рюдеманн 1912 , с. 136.
- ^ Кларк и Рюдеманн 1912 , с. 137.
- ^ Кингсли 1894 , с. 119.
- ^ Ланкестер 1886 , с. 366.
- ^ Данлоп, Пенни и Джекель, 2018 , стр. 17.
- ^ Ламсделл 2012 , с. 19.
- ^ Ламсделл 2012 , стр. 20–21.
- ^ Толлертон 1989 , с. 642.
- ^ Толлертон 1989 , стр. 642–644.
- ^ Толлертон 1989 , с. 649.
- ^ Толлертон 1989 , с. 646.
- ^ Толлертон 1989 , с. 650.
- ^ Ламсделл и др. 2015 , с. 25.
- ^ Тетли и Кагги 2007 , стр. 350.
- ^ Тетли 2007 , стр. 565.
- ^ Ламсделл, Брэдди и Тетли 2010 , стр. 56.
- ^ Ламсделл и др. 2015 , с. 3.
Библиография
[ редактировать ]- Бергстром, Карл Т.; Дугаткин, Ли Алан (2012). Эволюция . Нортон. ISBN 978-0393913415 .
- Брэдди, Саймон Дж.; Данлоп, Джейсон А. (1997). «Функциональная морфология спаривания силурийского эвриптерида Baltoeurypterus тетрагонофтальмус (Фишер, 1839)» . Зоологический журнал Линнеевского общества . 120 (4): 435–461. дои : 10.1111/j.1096-3642.1997.tb01282.x . ISSN 0024-4082 .
- Брэдди, Саймон Дж.; Алмонд, Джон Э. (1999). «Следы эвриптерид из группы Столовых гор (ордовик) Южной Африки». Журнал африканских наук о Земле . 29 (1): 165–177. Бибкод : 1999JAfES..29..165B . дои : 10.1016/S0899-5362(99)00087-1 .
- Брэдди, Саймон Дж.; Пошманн, Маркус; Тетли, О. Эрик (2008). «Гигантский коготь обнаружил самое большое членистоногое в истории » Письма по биологии . 4 (1): 106–109. дои : 10.1098/rsbl.2007.0491 . ПМЦ 2412931 . ПМИД 18029297 .
- Брезински, Дэвид К.; Коллар, Альберт Д. (2016). «Переоценка возраста и происхождения гигантского Palmichnium kosinskiorum Eurypterid Trackway из округа Элк, штат Пенсильвания». Анналы музея Карнеги . 84 (1): 39–45. дои : 10.2992/007.084.0105 . S2CID 90662577 .
- Бриггс, Дерек Э.Г. (1985). «Гигантизм палеозойских членистоногих» . Специальные статьи по палеонтологии . 33 : 157–158.
- Бурмейстер, Герман (1843). Организация трилобитов произошла от их ныне живущих родственников . Джордж Реймер.
- Кларк, Джон Мейсон; Рюдеманн, Рудольф (1912). Эвриптерида Нью-Йорка . Библиотеки Калифорнийского университета. ISBN 978-1125460221 .
- Данлоп, Джейсон А.; Пенни, Дэвид; Джекель, Дениз (2018). «Сводный список ископаемых пауков и их родственников» (PDF) . Мировой каталог пауков . Музей естественной истории Берна.
- Халлам, Энтони; Виналл, Пол Б. (1997). Массовые вымирания и их последствия . Издательство Оксфордского университета. ISBN 978-0198549161 .
- Ханкен, Нильс-Мартин; Стёрмер, Лейф (1975). «След большого силурийского эвриптерида» (PDF) . Окаменелости и слои . 4 : 255–270. дои : 10.18261/8200049639-1975-16 . ISBN 82-00-04963-9 .
- Хембри, Дэниел И.; Платт, Брайан Ф.; Смит, Джон Дж. (2014). Экспериментальные подходы к пониманию ископаемых организмов: уроки жизни . Спрингер Наука и бизнес. ISBN 978-9401787208 .
- Хендерсон, Джон (1866). «IV. Уведомление о Slimonia Acuminata из силура Пентлендских холмов» . Труды Эдинбургского геологического общества . 1 (1): 15–18. дои : 10.1144/transed.1.1.15 . S2CID 131539776 .
- Кингсли, Джон Стерлинг (1894). «Классификация членистоногих». Американский натуралист . 28 (326): 118–135. дои : 10.1086/275878 . JSTOR 2452113 . S2CID 83826460 .
- Кьеллесвиг-Варинг, Эрик Н. (1961). «Силурийские Eurypterida валлийского пограничья». Журнал палеонтологии . 35 (4): 789–835. JSTOR 1301214 .
- Кьеллесвиг-Варинг, Эрик Н. (1964). «Краткий обзор семейства Pterygotidae Clarke и Ruedemann, 1912 (Eurypterida)». Журнал палеонтологии . 38 (2): 331–361. JSTOR 1301554 .
- Краус, Отто; Браукманн, Карстен (2003). «Ископаемые гиганты и сохранившиеся карлики. Arthropleurida и Pselaphognatha (Atelocerata, Diplopoda): признаки, филогенетические связи и строение» . Переговоры Ассоциации естественных наук в Гамбурге . 40 :5-50.
- Куес, Барри С.; Кицке, Кеннет К. (1981). «Большое собрание новых эвриптерид из члена Red Tanks, формация Мадера (поздний пенсильванский период - ранняя пермь) Нью-Мексико». Журнал палеонтологии . 55 (4): 709–729. JSTOR 1304420 .
- Ламсделл, Джеймс К.; Брэдди, Саймон Дж. (2009). «Правило Коупа и теория Ромера: закономерности разнообразия и гигантизма у эвриптерид и палеозойских позвоночных» . Письма по биологии . 6 (2): 265–269. дои : 10.1098/rsbl.2009.0700 . ПМК 2865068 . ПМИД 19828493 .
- Ламсделл, Джеймс К.; Брэдди, Саймон Дж.; Тетли, О. Эрик (2009). «Переописание Drepanopterus abonensis (Chelicerata: Eurypterida: Stylonurina) из позднего девона Портисхеда, Великобритания». Палеонтология . 52 (5): 1113–1139. Бибкод : 2009Palgy..52.1113L . дои : 10.1111/j.1475-4983.2009.00902.x . ISSN 1475-4983 . S2CID 129243775 .
- Ламсделл, Джеймс К.; Брэдди, Саймон Дж.; Тетли, О. Эрик (2010). «Систематика и филогения Stylonurina (Arthropoda: Chelicerata: Eurypterida)» . Журнал систематической палеонтологии . 8 (1): 49–61. Бибкод : 2010JSPal...8...49L . дои : 10.1080/14772011003603564 . ISSN 1478-0941 . S2CID 85398946 .
- Ламсделл, Джеймс К.; Маккой, Виктория Э.; Перрон-Феллер, Опал А.; Хопкинс, Мелани Дж. (2020). «Дыхание воздухом прекрасно сохранившегося морского скорпиона возрастом 340 миллионов лет» . Современная биология . 30 (21): 4316–4321. Бибкод : 2020CBio...30E4316L . дои : 10.1016/j.cub.2020.08.034 . ПМИД 32916114 . S2CID 221590821 .
- Ламсделл, Джеймс К. (2012). «Пересмотренная систематика палеозойских мечехвостов и миф о монофилетической мечехвосте» . Зоологический журнал Линнеевского общества . 167 : 1–27. дои : 10.1111/j.1096-3642.2012.00874.x .
- Ламсделл, Джеймс К.; Селден, Пол (2013). «Дети в лесу – уникальное окно в онтогенез морских скорпионов» . Эволюционная биология BMC . 13 (98): 98. Бибкод : 2013BMCEE..13...98L . дои : 10.1186/1471-2148-13-98 . ПМЦ 3679797 . ПМИД 23663507 .
- Ламсделл, Джеймс К. (2014). Селективность в эволюции палеозойских групп членистоногих с акцентом на массовое вымирание и радиацию: филогенетический подход . Университет Канзаса.
- Ламсделл, Джеймс К.; Бриггс, Дерек Э.Г.; Лю, Хуайбао; Витцке, Брайан Дж.; Маккей, Роберт М. (2015). «Самый старый описанный эвриптерид: гигантский мегалограптид среднего ордовика (дарривилиана) из Виннешик-Лагерштетте в Айове» . Эволюционная биология BMC . 15 (169): 169. Бибкод : 2015BMCEE..15..169L . дои : 10.1186/s12862-015-0443-9 . ПМК 4556007 . ПМИД 26324341 .
- Ланкестер, Э. Рэй (1886). «Профессор Клаус и классификация членистоногих» . Анналы и журнал естественной истории . 17 (100): 364–372. дои : 10.1080/00222938609460154 .
- Маккой, Фредерик (1849). «XLI.—О классификации некоторых британских ископаемых ракообразных с уведомлениями о новых формах в Университетской коллекции в Кембридже» . Анналы и журнал естественной истории . 4 (24): 392–414. дои : 10.1080/03745486009494858 .
- Маккой, Виктория Э.; Ламсделл, Джеймс К.; Пошманн, Маркус; Андерсон, Росс П.; Бриггс, Дерек Э.Г. (2015). «Тем лучше видеть вас: глаза и когти раскрывают эволюцию различных экологических ролей гигантских крыловидных эвриптерид» . Письма по биологии . 11 (8): 20150564. doi : 10.1098/rsbl.2015.0564 . ПМЦ 4571687 . ПМИД 26289442 .
- Наддс, Джон Р.; Селден, Пол (2008). Ископаемые экосистемы Северной Америки: Путеводитель по местам и их необычной биоте . Издательство Мэнсон. ISBN 978-1-84076-088-0 .
- О'Коннелл, Марджори (1916). «Среда обитания эвриптериды» . Бюллетень Общества естественных наук о буйволах . 11 (3): 1–278.
- Ортега-Эрнандес, Хавьер; Легг, Дэвид А.; Брэдди, Саймон Дж. (2012). «Филогения агласпидовых членистоногих и внутренние взаимоотношения внутри Artiopoda». Кладистика . 29 (1): 15–45. дои : 10.1111/j.1096-0031.2012.00413.x . ISSN 1502-3931 . ПМИД 34814371 . S2CID 85744103 .
- Плотник, Рой Э.; Баумиллер, Томаш К. (1988). «Крыловидный тельсон как биологический руль». Летайя . 21 (1): 13–27. Бибкод : 1988Лета..21...13П . дои : 10.1111/j.1502-3931.1988.tb01746.x . ISSN 1502-3931 .
- Пошманн, Маркус; Розефельдс, Эндрю (30 ноября 2021 г.). «Последний эвриптерид - запись о питающихся морских скорпионах из Австралии в южных высоких широтах ограничивает сроки их исчезновения». Историческая биология . 34 (10): 2020–2030. дои : 10.1080/08912963.2021.1998033 . ISSN 0891-2963 . S2CID 252467508 .
- Пошманн, Маркус; Тетли, О. Эрик (2004). «Об эмсских (раннедевонских) членистоногих Рейнских Сланцевых гор: 4. Эвриптериды Alkenopterus и Vinetopterus n. gen. (Arthropoda: Chelicerata)». Сенкенбергиана Летея . 84 (1–2): 173–193. дои : 10.1007/BF03043470 . S2CID 127313845 .
- Селден, Пол (1985). «Дыхание эвриптерид» (PDF) . Философские труды Королевского общества B: Биологические науки . 309 (1138): 219–226. Бибкод : 1985РСТБ.309..219С . дои : 10.1098/rstb.1985.0081 .
- Селден, Пол (1999). «Автэкология силурийских эвриптерид» (PDF) . Специальные статьи по палеонтологии . 32 : 39–54. ISSN 0038-6804 . Архивировано из оригинала (PDF) 3 августа 2011 г.
- Стёрмер, Лейф (1955). «Меростоматы». Трактат по палеонтологии беспозвоночных, часть P Arthropoda 2, Chelicerata . Университет Канзаса Пресс. АСИН B0043KRIVC .
- Тетли, О. Эрик (2006). «Два новых силурийских вида Eurypterus (Chelicerata: Eurypterida) из Норвегии и Канады и филогения рода» (PDF) . Журнал систематической палеонтологии . 4 (4): 397–412. Бибкод : 2006JSPal...4..397T . дои : 10.1017/S1477201906001921 . ISSN 1478-0941 . S2CID 83519549 . Архивировано из оригинала (PDF) 6 июня 2020 г. Проверено 9 октября 2018 г.
- Тетли, О. Эрик; Ван Рой, Питер (2006). «Переоценка Eurypterus dumonti Stainier, 1917 и его положения в составе Adelophthalmidae Tollerton, 1989» (PDF) . Бюллетень Королевского института естественных наук Бельгии . 76 : 79–90.
- Тетли, О. Эрик (2007). «Распространение и история расселения Eurypterida (Chelicerata)». Палеогеография, Палеоклиматология, Палеоэкология . 252 (3–4): 557–574. Бибкод : 2007PPP...252..557T . дои : 10.1016/j.palaeo.2007.05.011 . ISSN 0031-0182 .
- Тетли, О. Эрик; Кагги, Майкл Б. (2007). «Филогения базальных плавающих эвриптерид (Chelicerata; Eurypterida; Eurypterina)». Журнал систематической палеонтологии . 5 (3): 345–356. Бибкод : 2007JSPal...5..345T . дои : 10.1017/S1477201907002131 . S2CID 88413267 .
- Тетли, О. Эрик; Рабано, Изабель (2007). «Образцы Eurypterus (Chelicerata, Eurypterida) в коллекциях Museo Geominero (Геологическая служба Испании), Мадрид» (PDF) . Геологический и горный вестник . 118 (1): 117–126. ISSN 0366-0176 . Архивировано из оригинала (PDF) 22 июля 2011 года.
- Тетли, О. Эрик (2008). « Hallipterus excelsior , стилонурид (Chelicerata: Eurypterida) из позднедевонского комплекса дельты Катскилл, и его филогенетическое положение среди Hardieopteridae». Бюллетень Музея естественной истории Пибоди . 49 (1): 19–30. doi : 10.3374/0079-032X(2008)49[19:HEASCE]2.0.CO;2 . S2CID 85862868 .
- Тетли, О. Эрик; Бриггс, Дерек Э.Г. (2009). «Происхождение крыловидных эвриптерид (Chelicerata: Eurypterida)». Палеонтология . 52 (5): 1141–1148. Бибкод : 2009Palgy..52.1141T . дои : 10.1111/j.1475-4983.2009.00907.x . ISSN 0024-4082 . S2CID 84268058 .
- Толлертон, Виктор П. (1989). «Морфология, таксономия и классификация отряда Eurypterida Burmeister, 1843». Журнал палеонтологии . 63 (5): 642–657. Бибкод : 1989JPal...63..642T . дои : 10.1017/S0022336000041275 . JSTOR 1305624 . S2CID 46953627 .
- Ван Рой, Питер; Бриггс, Дерек Э.Г.; Гейнс, Роберт Р. (2015). «Окаменелости Фезуаты в Марокко: необычайные свидетельства морской жизни в раннем ордовике» . Журнал Геологического общества . 172 (5): 541–549. Бибкод : 2015JGSoc.172..541V . дои : 10.1144/jgs2015-017 . hdl : 1854/LU-8714212 . ISSN 0016-7649 . S2CID 129319753 .
- Вразо, Мэтью Б.; Чурка, Сэмюэл младший (2017). «Новые ископаемые свидетельства плавательного поведения эвриптерид» . Палеонтология . 61 (2): 235–252. дои : 10.1111/пала.12336 . S2CID 133765946 .
- Уайт, Дэвид (1927). «Флора сланцев Эрмит, Гранд-Каньон, Аризона» . Труды Национальной академии наук Соединенных Штатов Америки . 13 (8): 574–575. дои : 10.1073/pnas.13.8.574 . ПМЦ 1085121 . ПМИД 16587225 .
- Уайт, Мартин А. (2005). «Гигантская тропа ископаемых членистоногих» . Природа . 438 (7068): 576. Бибкод : 2005Natur.438..576W . дои : 10.1038/438576a . ПМИД 16319874 . S2CID 4422644 .
- Вудворд, Генри (1865). «О некоторых новых видах ракообразных отряда Eurypterida». Ежеквартальный журнал Геологического общества . 21 (1–2): 484–486. дои : 10.1144/GSL.JGS.1865.021.01-02.52 . S2CID 129427356 .
Веб-сайты
[ редактировать ]- Казлев, М. Алан (2002). «Палеос – Эвриптерида» . www.palaeos.com . Архивировано из оригинала 13 августа 2007 года.
Внешние ссылки
[ редактировать ]- Eurypterids.co.uk - онлайн-ресурс данных и исследований по эвриптеридам.
- eurypterid.net

| Базы данных органов управления : Национальные |
|---|