Якелоптерус
| Якелоптерус | |
|---|---|

| |
| Ископаемое J. rhenaniae , Музей естественной истории, Майнц. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Подтип: | Хелицерата |
| Заказ: | † Эвриптерида |
| Суперсемейство: | † Птериготиоидеа |
| Семья: | † Птериготиды |
| Род: | † Якелоптерус Уотерстон, 1964 год. |
| Типовой вид | |
| † Jaekelopterus rhenianii ( Джекель , 1914)
| |
| Разновидность | |
| |
| Синонимы | |
| |
Jaekelopterus — род хищных эвриптерид , группы вымерших водных членистоногих . Окаменелости Jaekelopterus были обнаружены в отложениях раннего девона , пражского и эмсского ярусов. Известны два вида: типовой вид J. rhenaniae из солоновато - пресноводных слоев в Рейнской области и J. Howelli из устьевых слоев в Вайоминге . Родовое название сочетает в себе имя немецкого палеонтолога Отто Якеля , описавшего типовой вид, и греческое слово πτερόν ( птерон ), означающее «крыло».
Судя по изолированным ископаемым останкам большой хелицеры (коготь) из формации Клерф в Германии, было подсчитано, что J. rhenaniae достиг размера около 2,3–2,6 метра (7,5–8,5 футов), что делает его самым крупным членистоногим из когда-либо существовавших. обнаружен, превосходя других крупных членистоногих, таких как собратья-эвриптериды Acutiramus и Pterygotus ; многоножка Артроплевра . J. Howelli был намного меньше, достигая 80 сантиметров (2,6 фута) в длину.
По внешнему виду Jaekelopterus похож на других крыловидных эвриптерид, обладая большим расширенным тельсоном (самым задним сегментом тела) и увеличенными клешнями и передними конечностями. Оба вида Jaekelopterus были впервые описаны как виды близкородственного Pterygotus , но были выделены в отдельный род на основании наблюдаемых различий в генитальных придатках. Хотя с тех пор эта особенность оказалась ошибочной, были идентифицированы и другие особенности, отличающие этот род от его родственников, в том числе тельсон треугольной формы и другой наклон зубцов когтей.
Хелицеры и сложные глаза Jaekelopterus указывают на то, что он был активным и мощным, с высокой остротой зрения и, скорее всего, был высшим хищником в экосистемах раннего девона Еврамерики . Хотя эвриптерид, таких как Jaekelopterus, часто называют «морскими скорпионами», слои, в которых были обнаружены окаменелости Jaekelopterus , позволяют предположить, что они жили в пресноводной среде.
Описание
[ редактировать ]

Jaekelopterus — самый крупный из известных эвриптерид и самое большое известное членистоногое из когда-либо существовавших. Это было определено на основе хелицер (коготь) из эмсской формации Клерф в Вильверате, Германия , длина которой составляет 36,4 сантиметра (14,3 дюйма), но в которой отсутствует четверть ее длины, что позволяет предположить, что полная хелицера составляла бы 45,5 сантиметра ( 17,9 дюйма) в длину. Если соотношение длины тела к длине хелицер совпадает с таковым у других гигантских птериготид , таких как Acutiramus и Pterygotus , у которых соотношение между размером когтей и длиной тела относительно постоянное, организм, обладавший хелицерами, имел бы размеры от 233 до 259 сантиметров ( 7,64 и 8,50 футов) в длину. Если хелицеры вытянутся, к этой длине прибавится еще один метр. Эта оценка превышает максимальный размер тела всех других известных гигантских членистоногих почти на полметра, даже если не учитывать расширенные хелицеры. [ 1 ]
Jaekelopterus похож на другие крыловидные эвриптериды по своей общей морфологии . [ 2 ] отличается треугольным тельсоном (самый задний сегмент тела) и наклоненными главными зубцами на ветвях хелицер (подвижная часть клешней). [ 3 ] Птериготиды, группа высокоразвитых ( «продвинутых») эвриптерид, отличаются от других групп некоторыми особенностями, особенно хелицерами и тельсоном. Хелицеры Pterygotidae увеличены и мощные, явно приспособлены к активному захвату добычи, с хелами (клешнями), более похожими на клешни некоторых современных ракообразных , с хорошо развитыми зубцами на клешнях по сравнению с хелицерами других групп эвриптерид. [ 4 ] Еще одной особенностью, отличающей эту группу от других групп эвриптерид, являются их уплощенные и расширенные тельсоны, которые, вероятно, используются в качестве рулей при плавании. [ 5 ]
J. Howelli , известный по более чем 30 экземплярам, имеет почти такой же рисунок зубцов на хелицерах, как и J. rhenaniae , а также сохраняет уплощенный задний край тельсона, что приводит к треугольной форме, как у J. rhenaniae . Зубчатый край тельсона и массивное удлинение второго промежуточного зубца четко отличают его от J. rhenaniae . Кроме того, генитальный придаток типа А не раздваивается на конце. [ 3 ] J. Howelli намного меньше J. rhenaniae , достигая 80 сантиметров (2,6 фута) в длину. [ 6 ]
История исследований
[ редактировать ]
Jaekelopterus был первоначально описан как вид Pterygotus , P. rhenaniae , в 1914 году немецким палеонтологом Отто Йекелем на основе полученного им изолированного ископаемого претельсона (сегмента, непосредственно предшествующего тельсону), который был обнаружен в Алькене в нижнего девона отложениях Рейнской области. в Германии. Якель считал претельсон характерным для Pterygotus , другие обнаруженные элементы мало отличались от ранее известных видов этого рода, таких как P. buffaloensis , а длину животного при жизни он оценил примерно в 1 метр (1,5 метра, если хелицеры включены, 3,3 и 4,9 фута). [ 7 ]
Основываясь на более полном материале, включая генитальные придатки, хелицеры и фрагменты метастомы ( большая пластинка, являющаяся частью брюшка) и тельсона, обнаруженных немецким палеонтологом Вальтером Р. Гроссом недалеко от Оверата , Германия, норвежский палеонтолог Лейф Стёрмер предоставил более полную картину. и подробное описание вида в 1936 г. [ 8 ] Штёрмер интерпретировал генитальные придатки как сегментированные, в отличие от других видов Pterygotus . [ 9 ]
Британский палеонтолог Чарльз Д. Уотерстон выделил род Jaekelopterus в 1964 году для включения Pterygotus rhenaniae , который он считал достаточно отличным от других видов Pterygotus , чтобы гарантировать отдельный род, в первую очередь из-за того, что брюшные придатки Jaekelopterus были сегментированы в отличие от таковых у Pterygotus . [ 10 ] Уотерстон диагностировал Jaekelopterus как крыловидное крыло с сегментированными половыми придатками, трапециевидной просомой , узкими и длинными хелицерами с концевыми зубцами почти под прямым углом к ветвям и молочными зубами, слегка наклоненными вперед, а также с тельсоном с расширенным концевым шипом и дорсальным килем. [ 10 ] Родовое название дано в честь Отто Якеля; греческое слово πτερόν ( птерон ), означающее «крыло», является распространенным эпитетом в названиях эвриптерид. [ 10 ]
В 1974 году Стёрмер выделил новое семейство для рода Jaekelopteridae из-за предполагаемых значительных различий между генитальными придатками Jaekelopterus и других птериготид. [ 9 ] С тех пор было доказано, что эта расходящаяся особенность просто представляет собой неправильную интерпретацию Стёрмера в 1936 году: генитальный придаток Jaekelopterus на самом деле не сегментирован, как и у Pterygotus . [ 1 ] Таким образом, семейство Jaekelopteridae впоследствии было отвергнуто и рассматривалось как синоним семейства Pterygotidae. [ 9 ]
Другой вид Pterygotus , P.howelli , был назван американскими палеонтологами Эриком Кьеллесвиг-Варингом и Стормером в 1952 году на основе ископаемых тельсона и тергита ( дорсальная часть сегмента тела) из отложений нижнего девона формации Беартут-Бьютт в Вайоминге . Видовое название Howelli дано в честь доктора Бенджамина Хауэлла из Принстонского университета, который одолжил исследованные в описании ископаемые образцы Кьеллесвиг-Варингу и Стормеру. [ 11 ] Этот вид был отнесен к Jaekelopterus как Jaekelopterus Howelli норвежским палеонтологом О. Эриком Тетли в 2007 году. [ 4 ]
Классификация
[ редактировать ]

Jaekelopterus относится к семейству Pterygotidae в надсемействе Pterygotioidea . [ 3 ] [ 1 ] [ 12 ] Jaekelopterus похож на Pterygotus , практически отличается только особенностями генитального придатка и, возможно, тельсона. Близкое сходство между этими двумя родами побудило некоторых исследователей задаться вопросом, не являются ли птериготиды чрезмерно разделенными на родовом уровне. Основываясь на некотором сходстве генитального придатка, американские палеонтологи Джеймс К. Ламсделл и Дэвид А. Легг в 2010 году предположили, что Jaekelopterus , Pterygotus и даже Acutiramus могут быть синонимами друг друга. [ 2 ] Хотя различия были отмечены в хелицерах, эти структуры были поставлены под сомнение Чарльзом Д. Уотерстоном в 1964 году как основа родовых различий эвриптерид, поскольку их морфология зависит от образа жизни и варьируется на протяжении онтогенеза (развития организма после его рождения). Хотя морфологию тельсонов можно использовать для различения родов у эвриптерид, Ламсделл и Легг отметили, что треугольный тельсон Jaekelopterus все еще может находиться в пределах морфологического диапазона лопастных тельсонов, присутствующих у Pterygotus и Acutiramus . [ 2 ] Половые придатки могут различаться даже внутри рода; например, генитальный придаток Acutiramus меняется от вида к виду: у более ранних видов он имеет форму ложки, а затем становится двулопастным и в конечном итоге начинает выглядеть похожим на придаток Jaekelopterus . Ламсделл и Легг пришли к выводу, что необходим инклюзивный филогенетический анализ с участием нескольких видов Acutiramus , Pterygotus и Jaekelopterus , чтобы решить, являются ли эти роды синонимами друг друга. [ 2 ]
основана Приведенная ниже кладограмма на девяти наиболее известных видах птериготид и двух таксонах внешней группы ( Slimonia acuminata и Hughmilleria Socialis ). Jaekelopterus ранее классифицировался как базальный сестринский таксон по отношению к остальным Pterygotidae с момента его описания Уотерстоном в 1964 году как отдельный род из-за его предположительно сегментированных генитальных придатков (слитых и неразделенных у других птериготид), но повторное исследование рассматриваемых экземпляров выявили, что генитальный придаток Jaekelopterus также не был разделен. Материал изучен и филогенетический анализ проведен британским палеонтологом Саймоном Дж. Брэдди, немецким палеонтологом Маркусом Пошманном и О. Эриком Тетли в 2007 г. [ 1 ] выявили, что Jaekelopterus не был базальным крыловидным, а одним из наиболее производных таксонов в группе. [ 1 ] Кладограмма также содержит максимальные размеры, достигнутые рассматриваемым видом, что, возможно, было эволюционной чертой группы согласно правилу Коупа («филетический гигантизм») Брэдди, Пошманном и Тетли. [ 1 ] [ 13 ]
| Птериготиоидеа |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]Гигантизм
[ редактировать ]
К крыловидным эвриптеридам относятся многие из крупнейших известных эвриптерид, такие как Pterygotus и Acutiramus . Было высказано предположение о нескольких факторах, которые могли способствовать беспрецедентно большому размеру Jaekelopterus , его родственников и других крупных палеозойских беспозвоночных, таких как хищничество, ухаживание, конкуренция и ресурсы окружающей среды. [ 14 ]
Такие факторы, как дыхание, энергетические затраты на линьку , передвижение и фактические свойства экзоскелета, ограничивают размер членистоногих. [ 1 ] За исключением мощных и сильно склеротизированных когтей, большинство сохранившихся крупных сегментов тела птериготид тонкие и неминерализованные. Даже тергиты и стерниты (пластины, образующие поверхности брюшных сегментов) обычно сохраняются в виде сжатий толщиной с бумагу, что позволяет предположить, что птериготиды были очень легкими по конструкции. [ 1 ] Подобные адаптации к легкому весу можно наблюдать и у других гигантских членистоногих палеозоя, таких как гигантская многоножка Arthropleura , и было высказано предположение, что это жизненно важно для эволюции размеров гигантских членистоногих. [ 15 ] Легкая конструкция снижает влияние факторов, ограничивающих размер тела. [ 1 ]
и другие гигантские крыловидные эвриптериды были самыми крупными членистоногими, легкое телосложение Несмотря на то, что Jaekelopterus означало, что они, вероятно, не были самыми тяжелыми. Другие гигантские эвриптериды, особенно глубокотелые шагающие формы Hibbertopteridae , длиной почти 2 метра такие как Hibbertopterus , возможно, соперничали по весу с птериготидами и другими гигантскими членистоногими, а то и превосходили их. [ 16 ]
Американский палеонтолог Александр Кайзер и южноафриканский палеонтолог Жако Клок в 2008 году предположили, что оценки огромных размеров Jaekelopterus преувеличены, отметив, что оценки размеров предполагают, что относительные пропорции между хелицерами и длиной тела останутся такими же по мере взросления животного. Зубцы (зубцы когтей) имели положительную аллометрию (они были пропорционально больше у более крупных экземпляров), что, как предполагают Кайзер и Клок, могло произойти в хелицерах в целом. Кроме того, самые большие тазики (сегменты конечностей), обнаруженные у того же вида, шириной 27 сантиметров (11 дюймов), [ 1 ] предполагают, что общая максимальная длина тела составляет всего 180 сантиметров (5,9 футов). [ 17 ] Положительная аллометрия не была продемонстрирована в хелицерах эвриптерид в целом ни у одного другого рода эвриптерид, в том числе у ближайших родственников Jaekelopterus . Есть также несколько неописанных экземпляров J. rhenaniae, сходных по пропорциям с большими хелицерами, в том числе еще одна клешня, найденная в тех же слоях, что и первоначальная находка. По мнению Брэдди, Пошмана и Тетли, которые ответили Кайзеру и Клоку в том же году, оценки размера около 2,5 метров (8,2 фута) остаются наиболее точными оценками максимального размера этого вида на данный момент. [ 18 ]
Онтогенез
[ редактировать ]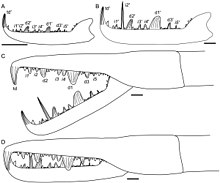
Как и все другие членистоногие, эвриптериды созревают через последовательность стадий, называемых « возрастами », состоящих из периодов шелушения (линьки), за которыми следует быстрый рост. В отличие от многих членистоногих, таких как насекомые и ракообразные, хелицераты (группа, к которой принадлежат эвриптериды, такие как Jaekelopterus , наряду с другими организмами, такими как мечехвосты , морские пауки и паукообразные ), как правило, являются непосредственными разработчиками, а это означает, что после их появления не происходит резких морфологических изменений. вылупился. Существующие меченосцы вылупляются без полного набора взрослых опистосомальных придатков (придатков, прикрепленных к опистосоме , задним сегментам тела), но современные пауки являются полностью прямыми разработчиками. Исследования ископаемых образцов Strobilopterus и Jaekelopterus позволяют предположить, что онтогенез эвриптерид во многом аналогичен таковому современных мечехвостов, но что эвриптериды (как и паукообразные) были настоящими прямыми разработчиками, вылупляясь с тем же количеством придатков и сегментов, что и взрослые особи. [ 3 ]
Хотя известно несколько окаменевших возрастов Jaekelopterus Howelli , фрагментарный и неполный статус экземпляров затрудняет детальное изучение его онтогенеза. Несмотря на это, в хелицерах, тельсоне и метастоме происходят заметные изменения. Четыре экземпляра J. Howelli , изученные Ламсделлом и Селденом (2013), сохранили хелицеры достаточно подробно, чтобы можно было изучить зубчики. Предполагалось, что две из этих хелицер произошли от молодых особей, а две - от взрослых особей. Морфология хелицер одинакова во всех возрастах, с одинаковым расположением и количеством зубчиков, но имелись и некоторые заметные различия. В частности, главные зубцы увеличивались в размерах относительно промежуточных: у молодых особей они в 1,5 раза превышали размеры промежуточных, а у взрослых - до 3,5 раз. Более того, у взрослых особей концевой зубец был намного крупнее и прочнее, чем у молодых. Возможно, самое крайнее из всех, то, что второй промежуточный зубец не отличается по размеру от других промежуточных зубцов у молодых особей, но у взрослых он значительно удлинен, где его длина более чем в два раза превышает длину любого основного зубца. [ 3 ] Хотя такой рост зубцов птериготид был описан и у других родов, массовое удлинение второго промежуточного зубца в процессе онтогенеза уникально для Jaekelopterus , особенно для J. Howelli . [ 3 ]
Метастома Jaekelopterus также меняла свои размеры по мере взросления животного. У J. rhenaniae относительная ширина метастомы уменьшалась в процессе онтогенеза. Метастома у J. Howelli также шире у молодых особей, чем у взрослых, хотя соотношение длины и ширины, измеренное у молодых и взрослых особей, не было таким разным, как предполагалось, и составляло 1,43 у молодых особей и 1,46 у взрослых. [ 3 ] Такое изменение размеров метастомы отмечено и у других родов эвриптерид, таких как Stoermeropterus , Moselopterus и Strobilopterus . [ 3 ]
Визуальная система
[ редактировать ]
Морфология хелицер и острота зрения крыловидных эвриптерид разделяют их на отдельные экологические группы. Основным методом определения остроты зрения у членистоногих является определение количества линз в их сложных глазах и межоматидиального угла (ИОА), который представляет собой угол между оптическими осями соседних линз. IOA особенно важен, поскольку его можно использовать для различения различных экологических ролей у членистоногих, поскольку у современных активных членистоногих хищников он низкий. [ 19 ]
И Jaekelopterus rhenaniae , и Pterygotus anglicus имели высокую остроту зрения, о чем свидетельствует низкий IOA и множество линз в их сложных глазах. [ 20 ] Дальнейшие исследования сложных глаз окаменелых экземпляров J. rhenaniae , включая крупный экземпляр с правым глазом, сохранившимся с самого верхнего зигена , и небольшой и, вероятно, ювенильный экземпляр, подтвердили высокую остроту зрения этого рода. Общий средний IOA Jaekelopterus (0,87°) сопоставим с таковым у современных хищных членистоногих. Острота зрения Jaekelopterus увеличивалась с возрастом, у более мелких экземпляров зрение было относительно хуже. [ 21 ] Это согласуется с другими птериготидами, такими как Acutiramus , и было интерпретировано как указание на то, что взрослые особи Jaekelopterus жили в более темной среде, например, на более глубокой воде. Следы ископаемых свидетельств существования эвриптерид также подтверждают такой вывод, указывая на то, что эвриптериды мигрировали в прибрежную среду для спаривания и нереста. [ 21 ]
Jaekelopterus обладал фронтально перекрывающимся полем зрения, т. е. стереоскопическим зрением , типичным для хищных животных. По строению глаза эвриптерид были практически идентичны глазам мечехвоста. Квадратный рисунок рецепторных клеток в сложных глазах Jaekelopterus также похож, но не идентичен, на рисунок мечехвостов, что позволяет предположить наличие специализированной зрительной системы. Фоторецепторы у Jaekelopterus необычно велики . Их размер составляет около 70 мкм, что намного больше, чем у человека (1–2 мкм) и большинства членистоногих (также 1–2 мкм), но по размеру они соответствуют размерам современных мечехвостов. [ 22 ]
Уникальные глаза современных мечехвостов сильно отличаются от глаз других современных членистоногих и позволяют улучшить восприятие краев и усилить контрасты, что важно для животных в условиях низкой и рассеянной освещенности. Поскольку глаза Jaekelopterus были очень похожи, вероятно, у него были такие же приспособления. Благодаря своим узкоспециализированным глазам Jaekelopterus был очень хорошо приспособлен к хищническому образу жизни. [ 22 ]
Палеоэкология
[ редактировать ]
Морфология и строение тела Jaekelopterus и других эвриптерид Pterygotidae позволяют предположить, что они были адаптированы к полностью водному образу жизни. Брэдди, Пошманн и Тетли в исследовании 2007 года пришли к выводу, что маловероятно, что членистоногое размером и телосложением Jaekelopterus сможет ходить по суше. [ 1 ] Эвриптерид, таких как Jaekelopterus, часто называют «морскими скорпионами», но отложения, в которых были обнаружены окаменелости Jaekelopterus, позволяют предположить, что они жили в неморской водной среде. Формация Беартут-Бьютт в Вайоминге, где были обнаружены окаменелости J. Howelli , интерпретируется как тихая мелководная среда устья . Этот вид был обнаружен вместе с двумя другими видами эвриптерид: Dorfopterus angusticollis и Strobilopterus Princetonii . [ 3 ] Местонахождения ископаемых месторождений J. rhenaniae в Рейнской области также интерпретируются как часть мелководной водной среды с солоноватой или пресной водой . [ 9 ]
Хелицеры Jaekelopterus увеличены, крепкие, имеют изогнутую свободную ветвь и зубцы разной длины и размера - все приспособления, которые соответствуют сильным прокалывающим и хватательным способностям современных скорпионов и ракообразных. Некоторые колотые раны на окаменелостях пораспидной бесчелюстной рыбы Lechriaspis patula из девона штата Юта , вероятно, были вызваны Jaekelopterus Howelli . [ 20 ] Последние исследования показывают, что Jaekelopterus был активным и визуальным хищником. [ 19 ] Взрослые Jaekelopterus были бы высшими хищниками в своей среде обитания и, вероятно, охотились на более мелких членистоногих (в том числе прибегая к каннибализму ) и ранних позвоночных. [ 1 ]
Могучий и активный хищник, Jaekelopterus , вероятно, был очень проворным и обладал высокой маневренностью. Гидромеханика и других птериготид позволяет предположить , плавательных лопастей и тельсонов Jaekelopterus что все представители группы были способны к зависанию, передвижению вперед и быстрым поворотам. Хотя они не обязательно были быстро плавающими животными, они, вероятно, были способны преследовать добычу в таких местах обитания, как лагуны и устья рек . [ 5 ] [ 21 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л Брэдди, Саймон Дж.; Пошманн, Маркус; Тетли, О. Эрик (2007). «Гигантский коготь обнаружил самое большое членистоногое в истории » Письма по биологии . 4 (1): 106–109. дои : 10.1098/rsbl.2007.0491 . ПМЦ 2412931 . ПМИД 18029297 .
- ^ Jump up to: а б с д Ламсделл, Джеймс К.; Легг, Дэвид А. (2010). «Изолированная ветвь крыловидной кости (Chelicerata: Eurypterida) из девонской формации Медвежий Зуб, Вайоминг» . Журнал палеонтологии . 84 (6): 1206–1208. дои : 10.1666/10-040.1 . S2CID 129807060 .
- ^ Jump up to: а б с д и ж г час я Ламсделл, Джеймс К.; Селден, Пол (2013). «Дети в лесу – уникальное окно в онтогенез морских скорпионов» . Эволюционная биология BMC . 13 (98): 1–46. дои : 10.1186/1471-2148-13-98 . ПМЦ 3679797 . ПМИД 23663507 .
- ^ Jump up to: а б Тетли, О. Эрик (2007). «Распространение и история расселения Eurypterida (Chelicerata)» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 252 (3–4): 557–574. дои : 10.1016/j.palaeo.2007.05.011 . Архивировано из оригинала (PDF) 18 июля 2011 г.
- ^ Jump up to: а б Плотник, Рой Э.; Баумиллер, Томаш К. (1 января 1988 г.). «Крыловидный тельсон как биологический руль». Летайя . 21 (1): 13–27. дои : 10.1111/j.1502-3931.1988.tb01746.x .
- ^ Ламсделл, Джеймс К.; Брэдди, Саймон Дж. (2009). «Правило Коупа и теория Ромера: закономерности разнообразия и гигантизма у эвриптерид и палеозойских позвоночных» . Письма по биологии . 6 (2): 265–9. дои : 10.1098/rsbl.2009.0700 . ПМК 2865068 . ПМИД 19828493 . Дополнительная информация .
- ^ Джаекель, Отто (1914). «Крупный птериготус из рейнского нижнего девона» . Палеонтологический журнал . 1 :379-382. дои : 10.1007/BF03160341 . S2CID 129100799 . Архивировано из оригинала 12 января 2018 г. Проверено 12 января 2018 г.
- ^ Стёрмер, Лейф (1936). «Эвриптериды рейнского нижнего девона» . Трактаты Прусского государственного геологического института . 175 .
- ^ Jump up to: а б с д Пошманн, Маркус; Тетли, О. Эрик (1 декабря 2006 г.). «Об эмсских (нижнедевонских) членистоногих Рейнских Сланцевых гор: 5. Редкие и малоизвестные эвриптериды из Вильверата, Германия». Палеонтологическая газета . 80 (4): 325–343. дои : 10.1007/BF02990208 . S2CID 129716740 .
- ^ Jump up to: а б с Д. Уотерстон, Чарльз (1 января 1964 г.). «II. Наблюдения за птериготидными эвриптеридами» . Труды Королевского общества Эдинбурга . 66 (2): 9–33. дои : 10.1017/S0080456800023309 . S2CID 130261793 .
- ^ Кьеллесвиг-Варинг, Эрик Н.; Штёрмер, Лейф (1952). «Нижнедевонский птериготус из Вайоминга». Журнал палеонтологии . 26 (6): 997–998. JSTOR 1299790 .
- ^ Данлоп, Джейсон А.; Пенни, Дэвид; Джекель, Дениз (2015). «Сводный список ископаемых пауков и их родственников (версия 16.0)» (PDF) . Мировой каталог пауков . Архивировано (PDF) из оригинала 29 ноября 2015 г. Проверено 7 декабря 2018 г.
- ^ Гулд, Джина С.; Макфадден, Брюс Дж. (1 июня 2004 г.). «Глава 17: Гигантизм, карликовость и правило Коупа: «ничто в эволюции не имеет смысла без филогении» ». Бюллетень Американского музея естественной истории . 285 : 219–237. doi : 10.1206/0003-0090(2004)285<0219:C>2.0.CO;2 . S2CID 73556985 .
- ^ Бриггс, Дерек (1985). «Гигантизм палеозойских членистоногих». Специальные статьи по палеонтологии . 33 : 157.
- ^ Краус, О., Браукманн, К. (26 августа 2003 г.). «Ископаемые гиганты и выжившие карлики. Arthropleurida и Pselaphognatha (Atelocerata, Diplopoda): характеры, филогенетические взаимоотношения и конструкция». Архивировано 9 сентября 2018 г. на Wayback Machine . Переговоры Ассоциации естественных наук в Гамбурге . 40 .
- ^ Тетли, О.Э. (2008). « Hallipterus excelsior , стилонурид (Chelicerata: Eurypterida) из позднедевонского комплекса дельты Катскилл, и его филогенетическое положение в Hardieopteridae». Бюллетень Музея естественной истории Пибоди . 49 : 19–99. doi : 10.3374/0079-032X(2008)49[19:HEASCE]2.0.CO;2 . S2CID 85862868 .
- ^ Кайзер, Александр; Клок, Жако (23 июня 2008 г.). «Означают ли гигантские когти гигантские тела? Альтернативный взгляд на преувеличенные масштабные отношения» . Письма по биологии . 4 (3): 279–280. дои : 10.1098/rsbl.2008.0015 . ПМК 2610042 . ПМИД 18353748 .
- ^ Брэдди, Саймон Дж.; Пошманн, Маркус; Тетли, О. Эрик (23 июня 2008 г.). «Ответ: гигантские когти и большие тела» . Письма по биологии . 4 (3): 281. doi : 10.1098/rsbl.2008.0116 . ПМК 2610059 .
- ^ Jump up to: а б Маккой, Виктория Э.; Ламсделл, Джеймс К.; Пошманн, Маркус; Андерсон, Росс П.; Бриггс, Дерек Э.Г. (01 августа 2015 г.). «Тем лучше видеть вас: глаза и когти раскрывают эволюцию различных экологических ролей гигантских крыловидных эвриптерид» . Письма по биологии . 11 (8): 20150564. doi : 10.1098/rsbl.2015.0564 . ПМЦ 4571687 . ПМИД 26289442 .
- ^ Jump up to: а б Эллиотт, Дэвид К.; Петриелло, Майкл А. (2011). «Новые пораспиды (Agnatha, Heterostraci) из раннего девона запада США». Журнал палеонтологии позвоночных . 31 (3): 518–530. дои : 10.1080/02724634.2011.557113 . S2CID 130564395 .
- ^ Jump up to: а б с Пошманн, Маркус; Шенеманн, Бриджит; Маккой, Виктория Э. (01 марта 2016 г.). «Яркие глаза: боковые зрительные системы рейнских эвриптерид нижнего девона (Arthropoda, Chelicerata) и их палеобиологические последствия» . Палеонтология . 59 (2): 295–304. дои : 10.1111/пала.12228 . ISSN 1475-4983 . S2CID 87690133 .
- ^ Jump up to: а б Шенеманн, Бриджит; Пошманн, Маркус; Кларксон, Юан НК (28 ноября 2019 г.). «Изучение глаз гигантских морских скорпионов (Eurypterida) возрастом 400 миллионов лет позволяет предположить структуру палеозойских сложных глаз» . Научные отчеты . 9 (1): 17797. Бибкод : 2019NatSR...917797S . дои : 10.1038/s41598-019-53590-8 . ISSN 2045-2322 . ПМЦ 6882788 . ПМИД 31780700 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Jaekelopterus, на Викискладе?
СМИ, связанные с Jaekelopterus, на Викискладе?

- Птериготиоидеа
- Животные раннего девона
- Девонские животные Европы
- Девонские животные Северной Америки
- Первые появления рода в раннем девоне
- Вымирание родов в раннем девоне
- Девонские эвриптериды
- Эвриптериды Европы
- Эвриптериды Северной Америки
- Ископаемые таксоны описаны в 1964 г.
- Таксоны, названные Отто Якелем