Крупнейшие доисторические животные

К крупнейшим доисторическим животным относятся как позвоночные , так и беспозвоночные виды. Многие из них описаны ниже вместе с типичным диапазоном их размеров (общие даты исчезновения см. по ссылке на каждый из них). Многие упомянутые виды на самом деле могут не быть самыми крупными представителями своей клады из-за неполноты летописи окаменелостей , а многие из приведенных размеров являются лишь приблизительными, поскольку ни один полный экземпляр не был найден. Масса их тела, в частности, во многом является лишь предположением, поскольку мягкие ткани редко окаменели. В целом размер вымерших видов зависел от энергетических [1] и биомеханические ограничения. [2]
Синапсиды немлекопитающих (Synapsida)
[ редактировать ]Казеазавры (Caeasauria)
[ редактировать ]Травоядный Алиеразавр был самым крупным казеидом и самым крупным амниотом , жившим в то время, его предполагаемая длина составляла около 6–7 м (20–23 футов). [3] Cotylorhynchus hancocki также крупный, его предполагаемая длина и вес не менее 6 м (20 футов). [4] и более 500 кг (1100 фунтов). [5]
Эдафозавриды (Edaphosauridae)
[ редактировать ]
Самыми крупными эдафозавридами были лупеозавры длиной 3 м (9,8 футов). [6] и Эдафозавр , который мог достигать даже более 3 м (9,8 футов) в длину. [7]
Сфенакодонтиды (Sphenacodontidae)
[ редактировать ]Самым крупным плотоядным синапсидом ранней перми был диметродон , который мог достигать 4,6 м (15 футов) и 250 кг (550 фунтов). [8] Крупнейшие представители рода Dimetrodon также были первыми в мире полностью наземными хищниками . [9]
Таппенозавриды
[ редактировать ]оценивалась в Длина среднепермского таппенозавра 5,5 м (18 футов), что почти такого же размера, как у крупнейших диноцефалов . [10]
Терапсиды (Therapsida)
[ редактировать ]Аномодонты (Anomodontia)
[ редактировать ]
Растительноядный дицинодонт Lisowicia bojani немлекопитающих - самый крупный известный из всех синапсидов , его длина составляет около 4,5 м (15 футов), высота 2,6 м (8 футов 6 дюймов) и масса тела 9000 кг (20 000 фунтов). [11] [12] [13]
Диноцефалы (Диноцефалия)
[ редактировать ]
Среди крупнейших плотоядных синапсидов, не относящихся к млекопитающим, был диноцефалический антеозавр , длина которого составляла 5–6 м (16–20 футов), а вес – 500–600 кг (1100–1300 фунтов). [14] [15] Взрослый Титанофонеус из того же семейства Anteosauridae, вероятно, имел череп длиной 1 м (3,3 фута). [15] Еще одним огромным диноцефалом был позднепермский Eotitanosurus (возможный синоним Biarmosuchus). [16] ). Взрослые экземпляры могли достигать 6 м (20 футов) в длину и более 600 кг (1300 фунтов) в весе. [16]
Горгонопсианы (Gorgonopsia)
[ редактировать ]
Inostrancevia latifrons — самый крупный из известных горгонопсов с длиной черепа более 60 см (24 дюйма), общей длиной около 3,5 м (11 футов) и массой 300 кг (660 фунтов). [17] Rubidgea atrox — крупнейшая африканская горгонопсия, длина черепа которой составляет почти 45 см (18 дюймов). [18] Другие крупные горгонопсии включают Диногоргона с черепом длиной около 40 см (16 дюймов), [19] Леонтозавр с черепом длиной почти 40 см (16 дюймов), [18] и сикозавр с черепом длиной около 38 см (15 дюймов). [18]
Тероцефалы (Therocephalia)
[ редактировать ]Самый крупный из тероцефалов — сцимнозавр . [20] [21] достигавший размеров современной гиены . [22]
Цинодонты немлекопитающих (Cynodontia)
[ редактировать ]- Самый крупный известный цинодонт, не относящийся к млекопитающим, — это Scalenodontoides , траверсодонтид , максимальная длина черепа которого, судя по фрагментарному экземпляру, составляла примерно 61,7 сантиметра (24,3 дюйма). [23]
- Paceyodon davidi был самым крупным из морганукодонтанов , цинодонтов, близких к млекопитающим. Он известен по правому нижнему моляру размером 3,3 мм (0,13 дюйма) в длину, что больше, чем у моляров всех других морганукодонтанов. [24]
- Самым крупным известным докодонтом был Castorocauda , длиной почти 50 см (20 дюймов). [25]
Млекопитающие (Mammalia)
[ редактировать ]Нетерианские млекопитающие
[ редактировать ]Гобиконодонты (Gobiconodonta)
[ редактировать ]
Самым крупным гобиконодонтом и самым крупным известным млекопитающим мезозоя был Repenomamus . [26] [27] [28] [29] [30] [31] Известная взрослая особь Repenomamus giganticus достигала общей длины около 1 м (3 фута 3 дюйма) и предполагаемой массы 12–14 кг (26–31 фунт). [28] При таких параметрах он превосходил по размерам нескольких мелких тероподовых динозавров раннего мела . [32] Гобиконодон также был крупным млекопитающим. [30] [31] он весил 5,4 кг (12 фунтов), [28] имел череп 10 см (3,9 дюйма) в длину и 35 см (14 дюймов) в пресакральной длине тела. [33]
Мультитуберкулезные (Multituberculata)
[ редактировать ]Самый крупный мультитуберкулезный [34] Taeniolabis taoensis — самое крупное из известных нетерийных млекопитающих , его вес, возможно, превышает 100 кг (220 фунтов). [35]
Однопроходные (Monotremata)
[ редактировать ]
- Самым крупным из когда-либо известных однопроходных (яйцекладущих млекопитающих) был вымерший вид длинноклювых ехидн , известный как Murrayglossus , известный по паре костей, найденных в Западной Австралии. Он был размером с овцу и весил, вероятно, до 30 кг (66 фунтов). [36]
- Самый крупный известный орниторинхид — Obdurodon tharalkooschild , его длина даже превышала 70 см (28 дюймов) Monotrmatum sudamericanum . [37]
- Колликодон, вероятно, был крупнейшим однопроходным в мезозое . Длина его тела могла достигать 1 м (3 фута 3 дюйма). [38]
Метатерийцы (Метатерия)
[ редактировать ]
- Самым крупным несумчатым метатерием был Thylacosmilus весом от 80 до 120 кг (от 180 до 260 фунтов). [39] [40] по одной оценке, даже 150 кг (330 фунтов). [41] По оценкам, Proborhyaenid Proborhyaena gigantea весит более 50 кг (110 фунтов) и, возможно, достигает 150 кг (330 фунтов). [42] Австралогиена — еще один крупный метатерий, весом до 70 кг (150 фунтов). [43]
- Стагодонтидное млекопитающее Дидельфодон был одним из крупнейших метатерий мезозоя и всех млекопитающих мелового периода . [44] Его череп мог достигать более 10 сантиметров (3,9 дюйма) в длину. [45] а вес всего животного составлял 5,2 кг (11 фунтов). [46]
Сумчатые (Marsupialia)
[ редактировать ]- Самым крупным известным сумчатым и самым крупным многоклеточным животным является вымерший дипротодон длиной около 3 м (9,8 футов), ростом 2 м (6 футов 7 дюймов) и весом до 2786 кг (6142 фунтов). [47] Его собратья -ворбатиформа Palorchestes azael были аналогичны по длине - около 2,5 м (8,2 фута), а по оценкам массы тела, она могла превышать 1000 кг (2200 фунтов). [48]
- Самым крупным известным хищным сумчатым животным был Thylacoleo carnifex . Измерения, проведенные у ряда экземпляров, показывают, что их средний вес составлял от 101 до 164 кг (от 223 до 362 фунтов). [49] [50]
- Самым крупным известным кенгуру был пока еще неназванный вид Macropus , весивший, по оценкам, 274 кг (604 фунта). [51] больше, чем самый крупный известный экземпляр Прокоптодона , который мог вырасти до 2 м (6 футов 7 дюймов) и весить 230 кг (510 фунтов). [52] Некоторые виды из рода Shenurus были сходны по размерам или немного крупнее современного серого кенгуру ( Macropus giganteus ). [53]
- Самым крупным потороидом, когда-либо зарегистрированным, был Борунгабуди , который был почти на 30% больше, чем самый крупный из ныне живущих видов, и весил до 10 кг (22 фунта). [54]
Неплацентарные евтерианы
[ редактировать ]
Чимолестанс (Чимолеста)
[ редактировать ]Самый крупный известный цимолестан - корифодон , высота в холке 1 м (3 фута 3 дюйма), длина 2,5 м (8 футов 2 дюйма). [55] [56] и массой до 700 кг (1500 фунтов). [57] Бариламбда также была огромным млекопитающим, ее вес составлял 650 кг (1430 фунтов). [58] Wortmania и Psittacotherium из группы Taeniodonta были одними из крупнейших млекопитающих раннего палеоцена . [59] Прожив всего через полмиллиона лет после границы K – Pg , Wortmania достигла массы тела 20 кг (44 фунта). Пситтакотерий , появившийся два миллиона лет спустя, достигал 50 кг (110 фунтов). [59]
Лептитиды
[ редактировать ]Самый крупный из -либо обнаруженных лепптид — Leptictidium tobieni из среднего эоцена Германии когда . У него был череп длиной 101 мм (4,0 дюйма), длина головы с туловищем 375 мм (14,8 дюйма) и хвост длиной 500 мм (20 дюймов). [60] Близкие европейские родственники из того же семейства Pseudorhyncocyonidae имели черепа длиной 67–101 мм (2,6–4,0 дюйма). [60]
Тенреки и союзники (Афроскида)
[ редактировать ]Самый крупный из двух видов бибималагасы ( Plesiorycteropus madagascariensis ), вымерших родственников тенреков с Мадагаскара , по оценкам, весил от 10 до 18 килограммов (от 21 до 40 фунтов). [61]
Парнокопытные (Artiodactyla)
[ редактировать ]
- Самым крупным известным наземным парнокопытным был Hippopotamus gorgops длиной 4,3 м (14 футов), высотой 2,1 м (6 футов 11 дюймов) и весом 5 т (11 000 фунтов). [62]
- Деодон и близкие ему по размеру и морфологии параэнтелодон. [63] были самыми крупными известными энтелодонтами , когда-либо жившими, их длина составляла 3,7 м (12 футов) и высота в плече 1,77 м (5,8 футов). [64] Огромный Эндрюсарх из эоцена Внутренней Монголии имел череп длиной 83,4 см (32,8 дюйма). [65] хотя систематика этого рода оспаривается. [66] [67]

- Самым крупным из быков , а также самым крупным быком был Bison latifrons . Он достиг веса 1250 кг (2760 фунтов). [68] [69] до 2000 кг (4400 фунтов), [70] Длина 4,75 м (15,6 футов), высота в плечах 2,31 м (7,6 футов), [71] и имели рога в размахе 2,13 м (7 футов 0 дюймов). [72] Североамериканский античный бизон достигал 4,6 м (15 футов) в длину, 2,27 м (7,4 фута) в высоту, вес 1588 кг (3501 фунт). [73] и размах рупора 1 м (3,3 фута). [71] Африканский пелоровис достигал веса 2 т (2,2 коротких тонны) и имел костные ядра рогов длиной около 1 м (3 фута 3 дюйма). [74] Другой огромный бык, африканский гигантский буйвол ( Syncerus antiquus ), достигал 3 м (9,8 фута) в длину от морды до конца хвоста, 1,85 м (6,1 фута) в высоту в холке , 1,7 м (5,6 фута) в высоту. в задней части, [75] [76] а расстояние между кончиками его рогов достигало 2,4 м (7 футов 10 дюймов). [75] Помимо местных популяций и подвидов современных видов, таких как популяция гаура на Шри-Ланке, зубра на Британских островах, кавказского зубра и карпатского зубра , самым крупным современным вымершим быком является зубр ( Bos primigenius ) со средней высотой в холке 155–180 см (61–71 дюйм) у быков и 135–155 см (53–61 дюйм) у коров, в то время как в популяциях зубров в Венгрии быки достигали 155–160 см (61–63 дюйма). [77] Купрей , ( Bos sauveli ), достигающий 1,7–1,9 м (5 футов 7 дюймов – 6 футов 3 дюйма) в высоту в плечах [78] [79] существует со среднего плейстоцена [80] и также считается, возможно, вымершим. [81] [82]
- Длинноногий Megalotragus , возможно, самый крупный из известных быков -алселафинов . [83] больше, чем нынешние антилопы гну . [84] Кончики рогов M. priscus располагались на расстоянии около 1,2 м (3 фута 11 дюймов) друг от друга. [85]

- Вымерший оленьий ирландский лось ( Megaloceros giganteus ) достигал высоты более 2,1 м (7 футов), массы 680 кг (1500 фунтов) и мог иметь рога шириной до 4,3 м (14 футов), что примерно в два раза превышало максимальный размах для рога лося. [86] [87] Гигантский лось ( Cervalces latifrons ) достигал высоты от 2,1 до 2,4 м (от 6,9 до 7,9 футов). [88] и был вдвое тяжелее ирландского лося, но размах его рогов 2,5 м (8,2 фута) был меньше, чем у Megaloceros . [89] [90] Североамериканский лось-олень ( Cervalces scotti ) достигал 2,5 метра (8,2 фута) в длину и веса 708,5 кг (1562 фунта). [91] [92]
- Самым крупным известным жирафом , не считая ныне живущих жирафов, является Sivatherium , с массой тела 1250 кг (2760 фунтов). [93]
- Самым крупным протоцератидом был Synthetoceras , он достигал 2 м (6 футов 7 дюймов) в длину и 150–200 кг (330–440 фунтов) в массе. [94] [95]
- Самым крупным из когда-либо существовавших диких свиней был Kubanochoerus gigas , его вес составлял до 500 кг (1100 фунтов), а рост в холке составлял около 1 м (3 фута 3 дюйма). [96] Megalochoerus мог быть схожим по размеру, возможно, весом 303 кг (668 фунтов) или 526 кг (1160 фунтов). [97]
- Самые крупные виды таяссуидов вымершие Platygonus были похожи по размеру на современных пекари, особенно на гигантских пекари , имели длину тела около 1 м (3,3 фута) и имели длинные ноги, что позволяло им хорошо бегать. У них также была свиная морда и длинные клыки , которые, вероятно, использовались для отпугивания хищников. [98]
- Самым крупным верблюдовым был Titanotylopus из миоцена Северной Америки. Возможно, он достигал 2485,6 кг (5480 фунтов) и высоты в плечах более 3,4 м (11 футов). [99] [100] Сирийский верблюд ( Camelus moreli ) был в два раза крупнее современных верблюдов. [101] Это было 3 м (9,8 футов) в плече. [102] и 4 м (13 футов) в высоту. [101] Ноги камелопа были на 20% длиннее, чем у дромадера , рост в холке составлял около 2,3 м (7 футов 7 дюймов), а вес - около 1000 кг (2200 фунтов). [103]
Китообразные (Cetacea)
[ редактировать ]
- Самым тяжелым археоцетом и, возможно, самым тяжелым известным млекопитающим был Perucetus , вес которого оценивается в 85–340 т (84–335 длинных тонн; 94–375 коротких тонн), а длина оценивается в 17,0–20,1 метра (55,8–65,9 футов). . [104] Самым длинным из известных китов-археоцитов эоцена был Basilosaurus, его длина составляла 17–20 м (56–66 футов). [105] [106] [107]
- Самым крупным сквалодельфинидом был Macrosqualodelphis , его длина составляла 3,5 м (11 футов). [108]
- Некоторые неогеновые рорквалы по размерам были сравнимы с современными огромными сородичами. По оценкам, Parabalaenoptera были размером с современного серого кита . [109] около 16 м (52 фута) в длину. Некоторые балаеноптериды, возможно, соперничали с синим китом . по размерам [109] хотя другие исследования не согласны с тем, что какой-либо усатый кит вырастал такими большими в миоцене . [110]
Непарнокопытные (Perissodactyla)
[ редактировать ]

- Самым крупным известным непарнокопытным и вторым по величине наземным млекопитающим (см. Palaeoloxodon namadicus ) всех времен был безрогий носорог Paraceratherium . Рост самой крупной известной особи оценивался в 4,8 м (15,7 фута) в плечах, 7,4 м (24,3 фута) в длину от носа до крупа и вес 17 т (18,7 коротких тонн). [111] [112]
- Некоторые доисторические рогатые носороги также вырастали до больших размеров. Самый большой эласмотерий достигал 5–5,2 м (16–17 футов) в длину. [113] Высота 2,5 м (8 футов 2 дюйма) [114] и весил 3,5–5 т (3,9–5,5 коротких тонн). [115] [113] [114] Такие параметры делают его крупнейшим носорогом четвертичного периода . [115] Шерстистый носорог ( Coelodonta antiquitatis ) того же времени достигал 1100–1500 кг (2400–3300 фунтов). [116] или 2000 кг (4400 фунтов), [117] [118] 1,93 м (6 футов 4 дюйма) на высоте плеч и 4,6 м (15 футов) в длину. [119]
- Метаминодон , аминодонтид , достигал 4 м (13 футов) в длину, сравнимый с гиппопотамом . по размерам и форме [120]
- Гигантский тапир ( Tapirus augustus ) был самым крупным тапиром за всю историю, его вес составлял около 623 кг (1373 фунта). [121] и рост 1 м (3,3 фута) в плечах. [122] Ранее это млекопитающее оценивалось еще крупнее, в 1,5 м (4,9 фута) в высоту, и было отнесено к отдельному роду Megatapirus . [122]
- Одним из крупнейших халикотериев был Моропус . [123] Его высота в холке составляла около 2,4 метра (8 футов). [124]
- Позднеэоценовые непарнокопытные из семейства Brontotheriidae достигали огромных размеров. Североамериканский мегацеропс (также известный как бронтотерий) . [125] ) достигал 2,5 м (8 футов 2 дюйма) в высоту в плечах, [126] 5 м (16 футов) в длину, [125] и вес 3 т (6600 фунтов). [127] Эмболотерий из Азии был такого же размера. [128]
- Самой крупной доисторической лошадью был Equus giganteus из Северной Америки. По оценкам, он вырастет до более чем 1250 кг (1,38 коротких тонны) и 2 м (6 футов 7 дюймов) в плечах. [129] Самым крупным анхитериновым непарнокопытным был Hypohippus весом от 403 до 600 кг (от 888 до 1323 фунтов), что сопоставимо с крупными современными домашними лошадьми . [130] [131] Мегагиппус — еще один крупный анхитериин. При массе тела 266,2 кг (587 фунтов) он был намного тяжелее большинства своих близких родственников. [130]
Фенакодонтиды (Phenacodontidae)
[ редактировать ]Самый крупный известный фенакодонтид — Phenacodus . Его длина составляла 1,5 м (4 фута 11 дюймов). [132] и весил до 56 кг (123 фунта). [133]
Диноцератаны (Dinocerata)
[ редактировать ]Самым крупным известным диноцератаном был Eobasileus с длиной черепа 102 см (40 дюймов), высотой 2,1 м (6 футов 11 дюймов) сзади и 1,5 м (4 фута 11 дюймов) высотой в плечах. [134] Другим огромным животным этой группы был Uintatherium с длиной черепа 76 см (30 дюймов), высотой в холке 1,5 м (4 фута 11 дюймов). [134] 4 м (13 футов) в длину и 2,25 т (2,48 коротких тонны), размер носорога . [135] Несмотря на свой большой размер, у Эобасилеуса , как и Уинтатерия, был очень маленький мозг. [134] [135]
Хищники (Carnivora)
[ редактировать ]Каниформия
[ редактировать ]


- Самым крупным наземным млекопитающим- хищником и самым большим известным медведем , а также крупнейшим известным наземным хищником всех времен был Arctotherium angustidens , южноамериканский короткомордый медведь. Плечевая кость из A. angustidens Буэнос -Айреса показывает, что самцы этого вида могли весить 1588–1749 кг (3501–3856 фунтов) и иметь рост на задних конечностях не менее 3,4 м (11 футов). [136] [137] Еще одним огромным медведем был гигантский короткомордый медведь ( Arctodus simus ), средний вес которого составлял 625 кг (1378 фунтов), а максимальный - 957 кг (2110 фунтов). [138] Есть предположение, что самые крупные особи этого вида могли достигать еще большей массы — до 1200 кг (2600 фунтов). [139] Вымерший пещерный медведь ( Ursus spelaeus ) также был тяжелее многих современных медведей. Самые крупные самцы весили до 1000 кг (2200 фунтов). [140] Ailuropoda baconi из плейстоцена была крупнее современной гигантской панды ( Ailuropoda melanoleuca ). [141]
- Самый большой одобенид и один из самых больших ластоногих , когда-либо существовавших, — это Pontolis magnus с длиной черепа 60 см (24 дюйма) (в два раза больше, чем черепа современных моржей -самцов ). [142] и имеющие общую длину тела более 4 м (13 футов). [143] [144] только современные самцы морских слонов ( Мирунга ). Подобных размеров достигают [143] Второе по величине доисторическое ластоногие — Gomphotaria pugnax с длиной черепа почти 47 см (19 дюймов). [142]
- Одним из крупнейших из доисторических отариид является Thalassoleon , сравнимый по размеру с крупнейшими из ныне живущих морских котиков . Ориентировочный вес T. mexicanus составляет не менее 295–318 кг (650–701 фунт). [145]
- Самым большим известным куньим , когда-либо существовавшим, вероятно, была гигантская выдра Энгидриодон . Его длина превышала 3 м (9,8 фута), а вес - около 200 кг (440 фунтов), что намного больше, чем у любого другого известного куньего, живого или вымершего. [146] [147] [148] Были и другие гигантские выдры, такие как Сиамогале , весом около 50 кг (110 фунтов). [149] и Мегаленгидрис , который был крупнее современной гигантской речной выдры. [150] Megalictis был крупнейшим чисто наземным куньим. [151] (хотя энгидриодон недавно упоминался как самый крупный куньих, который также является наземным хищником). [146] ). Похожий по размеру на ягуара , Megalictisferox имел еще более широкий череп, почти такой же широкий, как у черного медведя . [151] Мегаликтис обладал мощной силой укуса, что позволяло ему поедать крупную добычу и дробить кости, как это могут делать современные гиены и ягуары. [151] Еще одним крупным куньим был внешне кошачий Ekorus из миоцена Африки. Длинноногий Экорус весом почти 44 кг (97 фунтов) был размером с волка . [152] и заполнили экологическую нишу, аналогичную леопардам , до того, как на континент пришли большие кошки. [153] Другие огромные куньи включают Perunium. [154] и сверхплотоядные Eomellivora , оба из позднего миоцена . [155]
- Самым тяжелым проционидом , возможно, была южноамериканская Чапалмалания . Он достигал 1,5 метра (4,9 фута) в длину тела с коротким хвостом и 150 килограммов (330 фунтов), что сравнимо по размеру с американским черным медведем ( Ursus americanus ). [156] Еще одним огромным проционидом был Cyonasua , который весил около 15–25 кг (33–55 фунтов), что примерно такого же размера, как собака среднего размера. [157]
- Самой крупной собакой всех времен был Epicyon haydeni , рост которого в холке составлял 90 см (35 дюймов), длина тела составляла 2,4 м (7,9 фута), а вес - 100–125 кг (220–276 фунтов). [158] [159] [160] самый тяжелый из известных экземпляров весил до 170 кг (370 фунтов). [41] Вымерший лютоволк ( Aenocyon dirus ) достигал 1,5 м (4 фута 11 дюймов) в длину и весил от 50 до 110 кг (от 110 до 243 фунтов). [41] [161] Самый крупный подвид волка ( Canis lupus ), когда-либо существовавший в Европе, — это lupus maximus из позднего плейстоцена Франции Canis . Его длинные кости на 10% крупнее, чем у современных европейских волков , и на 20% длиннее, чем у C. l. лунелленсис . [162] Позднеплейстоценовый итальянский волк морфометрически был близок к C. l. Максимус . [163]
- Самой крупной медвежьей собакой был вид Pseudocyon весом около 773 кг (1704 фунта), что представляло собой очень крупную особь. [164]
Фелиформия
[ редактировать ]

- Самым крупным нимравидом, вероятно, был Quercylurus major , поскольку его окаменелости позволяют предположить, что он был похож по размеру на современного бурого медведя и имел сканвориальную форму . [165] В 2021 году Эусмилус был объявлен самым крупным из холплофониновых нимравидов, его вес достигал почти 111 кг (245 фунтов), что сравнимо с небольшим африканским львом. [166] Однако вес самого крупного гоплофонея оценивался в 160 кг (350 фунтов). [41]
- Самыми крупными саблезубыми кошками являются Amphimachairodus kabir и Smilodon populator , самцы, возможно, достигают веса 350–490 кг (770–1080 фунтов) и 220–450 кг (490–990 фунтов) соответственно. [41] [167] [168] Еще одним претендентом на звание крупнейшего кошачьего всех времен является Махайрод . M. horribilis из Китая оценивался в 405 кг (893 фунта). [169] в то время как североамериканский M. lahayishupup весил до 410 кг (900 фунтов). [170] [171] [172] Ксеносмилюс тоже был огромным котом. Его длина достигала около 2 м (6,6 футов). [173] и весил около 300–350 кг (660–770 фунтов). [169]
- Самыми тяжелыми из известных пантериновых кошачьих являются тигры Нгандонг ( Panthera tigris Soloensis ), вес которых, по оценкам, достигает 486 кг (1071 фунт). [168] американский лев ( Panthera atrox ) весом до 363 кг (800 фунтов), [174] [175] евразийский пещерный лев ( Panthera spelaea ) весом до 339 кг (747 фунтов), [168] и лев-натодомери из Восточной Африки, который по размеру был сопоставим с крупными представителями P. atrox. [176] Будучи предком современного ягуара , [177] Panthera gombaszoegensis была намного крупнее, максимальный вес до 150 кг (330 фунтов). [177]
- Некоторые вымершие кошачьи также превосходили своих современных сородичей по размерам. Евразийский гигантский гепард ( Acinonyx pardinensis ) достигал 60–121 кг (132–267 фунтов), что примерно в два раза больше современного гепарда . [178] Североамериканский Пратифелис был крупнее современной пумы . [179]
- Самым крупным барбурофелидом был Barbourofelis fricki с высотой плеч 90 см (35 дюймов). [180]
- Самая крупная из виверрид когда-либо существовавших — это Viverraleakeyi , которая была размером примерно с волка или маленького леопарда и весила 41 кг (90 фунтов). [181]
- Самая крупная известная ископаемая гиена - Pachycrocuta , ее длина в холке оценивается в 90–100 см (35–39 дюймов). [182] и вес 190 кг (420 фунтов). [41] Еще одна огромная гиена массой более 100 кг (220 фунтов) — пещерная гиена . На самом деле это подвид африканской пятнистой гиены , которая на 10% меньше вымершей пещерной гиены. [183]
- Перкрокутидная . кошачья, Dinocrocuta , была в два или даже в три раза крупнее современной пятнистой гиены — 160 или 240 кг (350 или 530 фунтов) [184]
- Вымершая гигантская ямка ( Cryptoprocta spelea ) имела массу тела от 17 кг (37 фунтов). [185] до 20 кг (44 фунта), [186] намного больше, чем весит современная ямка (до 8,6 кг (19 фунтов) для взрослых самцов). [187] ).
Гиенодонты (Hyaenodonta)
[ редактировать ]Самым крупным гиенодонтом был Симбакубва , его вес составлял 1500 кг (3300 фунтов). [188] Другой гигантский гиенодонт, Мегистотерий, достигал 500 кг (1100 фунтов). [41] и имел череп длиной 66,4 см (26,1 дюйма). [189]
Оксиениды (Oxyaenidae)
[ редактировать ]
Самым крупным известным оксиенидом был Саркастодон весом 800 кг (1800 фунтов). [41]
Мезонихии (Мезонихия)
[ редактировать ]Некоторые мезонихи достигали размеров медведя. Такими крупными были монголониксы из Азии. [190] и Анкалагон из Северной Америки. [191] [192] Еще один крупный мезонихий — Harpagolestes с длиной черепа у некоторых видов до полуметра. [190]
Летучие мыши (Chiroptera)
[ редактировать ]Обнаруженный в четвертичных отложениях Южной и Центральной Америки, Desmodus draculae имел размах крыльев 0,5 м (20 дюймов) и массу тела до 60 г (2,1 унции). Такие пропорции делают его самой крупной летучей мышью-вампиром , когда-либо существовавшей. [193]
Ежи, гимнуры, землеройки и кроты (Eulipotyphla)
[ редактировать ]
Самым крупным известным животным группы Eulipotyphla был дейногалерикс . [194] общая длина до 60 см (24 дюйма) и длина черепа до 21 см (8,3 дюйма). [195]
Грызуны (Rodentia)
[ редактировать ]
- Некоторые из вымерших южноамериканских диномид были намного крупнее современных грызунов. Josephoartigasia monesi был самым крупным известным грызуном всех времен, его вес составлял примерно 480–500 кг (1060–1100 фунтов). [196] Phoberomys pattersoni весил 125–150 кг (276–331 фунт). [196] И Джозефоартигасия , и Фоборомис достигали примерно 1,5 м (4 фута 11 дюймов) в высоту в холке. [197] Еще один огромный диномид, Telcomys gigantissimus, имел минимальный вес 200 кг (440 фунтов). [197]
- Amblyrhiza inundata из семейства Heptaxodontidae была крупным животным, ее вес составлял 50–200 кг (110–440 фунтов). [198] [197]
- Самым крупным бобром был гигантский бобр ( Castoroides ) Северной Америки. Он вырастал более 2 м в длину и весил примерно от 90 до 125 кг (от 198 до 276 фунтов), что также делало его одним из крупнейших когда-либо существовавших грызунов. [199]
- Самый крупный дикобраз Старого Света — Hystrix refossa — был крупнее ныне живущих дикобразов. Он был примерно на 20% крупнее своего ближайшего родственника, современного индийского дикобраза ( H. indica ), достигая длины более 115 см (45 дюймов).
Кролики, зайцы и пищухи (Lagomorpha)
[ редактировать ]Самый крупный известный доисторический зайцеобразный - гигантский минорский зайцеобразный Nuralagus rex весом 12 кг (26 фунтов). [200]
Панголины (Фолидота)
[ редактировать ]Самым крупным панголином был вымерший Manis palaeojavanica. [201] Его общая длина составляет до 2,5 м (8,2 фута). [202]
Приматы (Приматы)
[ редактировать ]
- Самым крупным известным приматом , а также самым крупным гоминидом всех времен был Gigantopithecus blackii , ростом 3 м (9,8 фута) и весом 540 кг (1200 фунтов). [203] [204] Однако в 2017 году новые исследования показали, что масса тела этого примата составляет 200–300 кг (440–660 фунтов). [205] Другим гигантским гоминидом был Meganthropus palaeojavanicus , ростом 2,4 м (7 футов 10 дюймов). [206] хотя это известно по очень плохим останкам. [207]
- В плейстоцене некоторые архаичные люди были близки по размерам или даже больше, чем ранние современные люди . Неандертальцы ( Homo neanderthalensis ) достигали 77,6 кг (171 фунт) и 66,4 кг (146 фунтов) в среднем весе для мужчин и женщин соответственно, что превышало параметры современного человека ( Homo sapiens ) (68,5 кг (151 фунт) и 59,2 кг). (131 фунт) для мужчин и женщин соответственно). [208] Большеберцовая кость из Кабве ( Замбия ) указывает на неопределённого человека , возможно, ростом 181,2 см (71,3 дюйма). Это был один из самых высоких людей среднего плейстоцена , заметно крупный даже по сравнению с современными людьми. [209] Самые высокие особи Homo sapiens из среднего плейстоцена Испании достигали 194 см (76 дюймов) и 174 см (69 дюймов) для мужчин и женщин соответственно. [209] Некоторые Homo erectus могли достигать 185 см (73 дюйма) в высоту и 68 кг (150 фунтов) в весе. [210] [211]
- Самая крупная известная обезьяна Старого Света - доисторический бабуин, самец динопитека , по прогнозам, будет весить в среднем от 46 кг (101 фунт) до 57 кг (126 фунтов). [212] Он превышает рекорд максимального веса павиана чакма , самого крупного из ныне живущих павианов. По оценкам одного источника, экземпляр Theropithecus oswaldi весил 72 кг (159 фунтов). [213]
- Самой крупной известной обезьяной Нового Света была Cartelles , которая изучалась как образец Protopithecus , весом до 34,27 кг (75,6 фунта). Caipora bambuiorum — еще один крупный вид, его вес достигает 27,74 кг (61,2 фунта). [214]
- Крупнейшими омомидами были Macrotarsius и Ourayia из среднего эоцена . Оба достигли веса 1,5–2 кг (3,3–4,4 фунта). [215]
- Некоторые доисторические лемуриформные приматы также вырастали до огромных размеров. Археоиндрис длиной 1,5 метра (4,9 фута) был лемуром-ленивцем , который жил на Мадагаскаре и весил 150–187,8 кг (331–414 фунтов). [216] размером со взрослый самец гориллы . [217] Палеопропитек из того же семейства также был тяжелее большинства современных лемуров - 25,8–45,8 кг (57–101 фунт). [218] Мегаладапис - еще один крупный вымерший лемур длиной от 1,3 до 1,5 м (от 4 футов 3 дюйма до 4 футов 11 дюймов). [ нужна ссылка ] и средняя масса тела около 140 кг (310 фунтов). [219] По другим оценкам, его вес составляет 46,5–85,1 кг (103–188 фунтов), но он все же намного крупнее любого существующего лемура. [218]
Слоны, мамонты и мастодонты (Proboscidea)
[ редактировать ]

- Самым крупным известным наземным млекопитающим когда-либо был хобот по имени Palaeoloxodon namadicus , который весил около 22 т (24,3 коротких тонны) и имел высоту около 5,2 м (17,1 фута) в холке. [111] самые крупные особи степного мамонта Евразии По оценкам , ( Mammuthus trogontherii ) достигают 4,5 м (14,8 футов) в плечах и 14,3 т (15,8 коротких тонн) в весе. [111] [220] Stegodon zdanskyi , самый крупный вид стегодонов , имел массу тела 13 т (14,3 коротких тонны). [111] Еще один огромный хоботок — Stegotetrabelodon syrticus , высотой более 4 м (13 футов) и весом от 11 до 12 т (от 12,1 до 13,2 коротких тонн). [111] Колумбийский мамонт ( Mammuthus columbi ) имел высоту в холке около 4 м (13,1 фута), но весил не так много, как другие огромные мамонты. Его средняя масса составляла 9,5 т (10,5 коротких тонн), а один необычно крупный экземпляр - около 12,5 т (13,8 коротких тонн). [111] Колумбийские мамонты имели очень длинные бивни. Самый большой известный бивень мамонта длиной 4,9 м (16 футов) принадлежал этому виду. [221]
- Самым крупным маммутидом был неогеновый Mammut borsoni . Самый крупный экземпляр достигал 4,1 м (13 футов) в высоту и весил около 16 т (17,6 коротких тонн). [111] У этого вида также были самые длинные бивни: 5,02 м (16,5 футов) в длину от основания до кончика по изгибу. [222]
- Дейнотерий был самым крупным хоботком семейства Deinotheriidae . Кости, найденные на Крите, подтверждают существование экземпляра высотой 4,1 м (13 футов) в плечах и весом более 14 т (15,4 коротких тонны). [111]
Морские коровы (Сирения)
[ редактировать ]По имеющимся данным, стеллеровы морские коровы во взрослом состоянии вырастают до 8–9 м (26–30 футов) в длину, что намного крупнее любых современных сирен . [223] Вес стеллеровых коров оценивается в 8–10 т (8,8–11,0 коротких тонн). [224]
Арсиноитерес (Arsinoitheriidae)
[ редактировать ]
Самым крупным известным арсинотериидом был Arsinoitherium . А. Зиттели имел рост 1,75 м (5 футов 9 дюймов) в плечах и 3 м (9,8 футов) в длину. [225] [226] A. giganteum достигал даже больших размеров, чем A. zitteli. [227]
Даманы (Hyracoidea)
[ редактировать ]Некоторые из доисторических даманов были чрезвычайно большими по сравнению с современными маленькими родственниками. Самый крупный гиракоид из когда-либо существовавших — Titanohyrax ultimus . [228] При оценке массы от 600 кг (1300 фунтов) до более 1300 кг (2900 фунтов) он был близок по размеру к суматранскому носорогу . [229] Еще один огромный даман - Мегалогиракс , череп которого имел длину 391 мм (15,4 дюйма). [230] и достиг размеров тапира. [231] [228] Более поздний Gigantohyrax был в три раза крупнее современного родственника Procavia capensis . [232] хотя он заметно меньше более ранних Megalohyrax и Titanohyrax . [233]
Демостилийцы (Desmostylia)
[ редактировать ]
Самым крупным из известных десмостилей был вид Desmostylus с длиной черепа 81,8 см (32,2 дюйма) и сравнимый по размеру с морской коровой Стеллера . [234]
Палеопарадоксия также известна как один из крупнейших десмостилей с длиной тела 3,03 м (9,9 футов). [235]
Броненосцы, глиптодонты и пампатеры (Cingulata)
[ редактировать ]Самая крупная поясная извилина известная — Doedicurus , длиной 4 м (13 футов) и высотой 1,5 м (4 фута 11 дюймов). [135] и достигая массы примерно от 1910 до 2370 кг (от 2,11 до 2,61 коротких тонны). [ нужна ссылка ] Самый крупный вид глиптодонов , Glyptodon clavipes , достигал 3–3,3 м (9,8–10,8 футов) в длину. [236] [135] и весом 2 т (2,2 коротких тонны). [ нужна ссылка ]
Муравьеды и ленивцы (Pilosa)
[ редактировать ]

Самый крупный из известных пилосанов — Eremotherium , наземный ленивец с предполагаемым весом до 6,55 т (7,22 коротких тонны) и длиной до 6 м (20 футов). [237] размером с быка африканского слона . Близкий родственник наземного ленивца Megatherium достиг таких же больших размеров. [238]
Астрапотерии (Astrapotheria)
[ редактировать ]Некоторые из крупнейших известных астрапотерий весили около 3–4 т (3,3–4,4 коротких тонны), включая род Granastrapotherium. [239] и некоторые виды Parastrapotherium ( P. Martiale ). [240] Остатки скелета позволяют предположить, что вид Hilarcotherium miyou был еще крупнее: его вес составлял 6,456 т (7,117 коротких тонн). [241]
Литоптерны (Литоптерны)
[ редактировать ]Самым крупным известным литоптерном была Macrauchenia , имевшая по три копыта на фут. Это было относительно крупное животное с длиной тела около 3 м (9,8 футов). [242]
Нотоунгуляты (Notoungulata)
[ редактировать ]Самым крупным неопытного животного из известных полных останков является токсодон . Он имел длину тела около 2,7 м (8 футов 10 дюймов) и высоту в плечах около 1,5 м (4 фута 11 дюймов) и напоминал тяжелого носорога. Сохранившиеся окаменелости, хотя и неполные, позволяют предположить, что Mixotoxodon был самым массивным представителем группы, его вес составлял около 3,8 т (4,2 коротких тонны). [243]
Пиротерийцы (Пиротерия)
[ редактировать ]Самым крупным млекопитающим южноамериканского отряда Pyrotheria был Pyrotherium , его длина составляла 2,9–3,6 м (9 футов 6 дюймов – 11 футов 10 дюймов), а вес - 1,8–3,5 т (4 000–7 700 фунтов). [244]
Рептилии
[ редактировать ]Ящерицы и змеи (Squamata)
[ редактировать ]
- Мозазавры — самые крупные из известных чешуйчатых животных . Самым крупным известным мозазавром, вероятно, является Mosasaurus hoffmanni , длина которого оценивается более чем в 17 м (56 футов). [245] [246] однако эти оценки основаны на соотношении длины головы и тела 1:10, что маловероятно для мозазавра , и, вероятно, это соотношение составляет около 1:7. [247] Другой гигантский мозазавр — Тилозавр , длина которого оценивается в 10–14 м (33–46 футов). [248] [249] Другой мозазавр, прогнатодон, может достигать подобных размеров.
- Самая крупная известная доисторическая змея — Titanoboa cerrejonensis , длина которой оценивается в 12,8 м (42 фута) или даже 14,3 м (47 футов). [250] в длину и 1135 кг (2502 фунта) в весе, [251] и мадцоид Vasuki indicus , высота которого, по оценкам, составляет 11–15 м (36–49 футов). [252] Близким соперником этих змей по размеру является палеофидная морская змея Palaeophis colossaeus , длина которой могла достигать около 9 м (30 футов). [251] [253] [254] или даже до 12,3 м (40 футов). [255] Другая известная очень крупная ископаемая змея — Gigantophis garstini , длина которой оценивается в 9,3–10,7 м (31–35 футов). [256] [257] хотя более позднее исследование показывает меньшую оценку - около 6,6–7,2 м (22–24 фута). [258] Самый крупный ископаемый питон — Liasis dubudingala , его длина составляет около 9 м (30 футов). [259] Самая большая гадюка , а также самая большая ядовитая змея, когда-либо зарегистрированная, — это crotaloides из раннего плиоцена Греции Laophis . Эта змея достигала более 3 м (9,8 футов) в длину и 26 кг (57 фунтов) в весе. [260] [261] Еще одна огромная ископаемая гадюка — неопределенный вид Vipera . При длине около 2 м (6 футов 7 дюймов) он был одним из крупнейших хищников Майорки в раннем плиоцене. [262] Самая крупная известная слепая змея - Boipeba tayasuensis, общая длина которой оценивается в 1,1 м (3 фута 7 дюймов). [263]
- Самая крупная из известных наземных ящериц — это, вероятно, мегалания ( Varanus priscus ) длиной 7 м (23 фута). [264] Будучи современными родственниками, мегалания могла быть ядовитой, и в этом случае эта ящерица также была самым крупным ядовитым позвоночным, когда-либо существовавшим. [265] Однако максимальный размер этого животного является предметом споров. [266]
Черепахи, черепахи и их близкие родственники (Pantestudines)
[ редактировать ]Криптодира
[ редактировать ]- Самой крупной известной черепахой когда-либо была Archelon ischyros, ее длина составляла 5 м (16 футов), а вес - 2200 кг (4900 фунтов). [267] Возможно, второй по величине морской черепахой была Protostega с общей длиной тела 3,9 м (13 футов). [268] [269] Существует даже более крупный экземпляр этого рода из Техаса, общая длина которого оценивается в 4,2 м (14 футов). [270] [268] По оценкам, частично известный кратохелон достигает 4 м (13 футов) в длину. [271] Еще одна огромная доисторическая морская черепаха — позднемеловой гигантатипус , длина которого оценивается более чем в 3,5 м (11 футов). [272] Psephophorus terrypratchetti из эоцена достигал 2,3–2,5 м (7,5–8,2 футов) в длину. [273]
- Самой крупной черепахой была Megalochelys atlas с длиной панциря до 2 м (6,6 футов). [274] и весом 0,8–1,0 т (1800–2200 фунтов). [127] M. margae имел панцирь длиной 1,4–2 м (4,6–6,6 футов); безымянный вид с Явы достигал длины панциря не менее 1,75 м (5,7 футов). [275] Кайнозойские . титанохелоны также были крупнее современных гигантских черепах , их длина панциря достигала 2 м (6 футов 7 дюймов) [276] [277] Другие гигантские черепахи включают Centrochelys marocana длиной панциря 1,8–2 м (5,9–6,6 футов) и мезоамериканскую Hesperotestudo sp. при длине панциря 1,5 м (4,9 фута). [275]
- Самая крупная из когда-либо зарегистрированных трионихид — неопределенный экземпляр GSP-UM 3019 из эоцена Пакистана среднего . Костный панцирь GSP-UM 3019 имеет длину 120 см (3,9 фута) и ширину 110 см (3,6 фута), что означает, что общий диаметр панциря (с мягким краем) составляет около 2 м (6,6 фута). [278] Drazinderetes tethyensis из той же формации имел костный панцирь длиной 80 см (2,6 фута) и шириной 70 см (2,3 фута). [278] Еще один огромный трионихид — североамериканский Axestemys byssinus, общая длина которого превышает 2 м (6,6 фута). [279]
Бокошейные черепахи (Pleurodira)
[ редактировать ]
Самой крупной пресноводной черепахой всех времен была миоценовая подокнемида Stupendemys с предполагаемой длиной парасагиттального панциря 2,86 м (9 футов 5 дюймов) и весом до 1145 кг (2524 фунта). [280] Carbonemys cofrinii из того же семейства имел раковину размером около 1,72 м (5 футов 8 дюймов). [281] [282] [283] полная оболочка оценивалась в 1,8 м (5,9 футов). [284]
Макробениды (Macrobaenidae)
[ редактировать ]Крупнейшими макробенидами были раннемеловой Yakemys , позднемеловой Anatolemys и палеоценовый Judithemys . Все достигали 70 см (2,3 фута) в длину панциря. [285]
Мейоланообразные
[ редактировать ]
Самым крупным мейоланиидом была Meiolania . Meiolania platyceps имела панцирь длиной 100 см (3,3 фута). [275] и, вероятно, достигал общей длины тела более 3 м (9,8 футов). [286] Безымянный вид позднего плейстоцена из Квинсленда был еще крупнее, до 200 см (6,6 футов) в длину панциря. [275] Ninjemys oweni достигал 100 см (3,3 фута) в длину панциря. [275] и вес 200 кг (440 фунтов). [287]
Зауроптеригии (Sauropterygia)
[ редактировать ]Плакодонты и близкие родственники (Placodontiformes)
[ редактировать ]Плакодус был одним из крупнейших плакодонтов , его длина достигала 3 м (9,8 футов). [288]
Нотозавры и близкие родственники (Nothosauroidea)
[ редактировать ]Самым крупным нотозавром , а также самым крупным триаса зауроптеригом был Nothosaurus giganteus длиной 7 м (23 фута). [289]
Плезиозавры (Plesiosauria)
[ редактировать ]- Самым крупным известным плезиозавроидом был неопределенный экземпляр, возможно, принадлежащий Aristonectes (идентифицированный как Aristonectes sp.), с длиной тела 11–11,9 метров (36–39 футов) и массой тела 10,7–13,5 метрических тонн (11,8–14,9 футов). короткие тонны). [290] Еще одним длинным плезиозавроидом был Albertonectes, его рост составлял 11,2–11,6 метров (37–38 футов). [291] Талассомедон конкурировал с ним по размеру: его длина составляла 10,86–11,6 м (35,6–38,1 фута). [292] Другими крупными плезиозавроидами являются Стиксозавр и Эласмозавр . Оба достигали более 10 м (33 футов) в длину. [293] [294] Гидралмозавр (ранее известный как эласмозавр и стиксозавр ) достигал 9,44 м (31,0 фута) в общей длине тела. [294] Раньше считалось, что Мауизавр имел длину более 8 м (26 футов). [295] [294] но позже его определили как nomen dubium . [296]

- Существует много споров по поводу самого крупного из известных представителей Pliosauroidea . Pliosaurus funkei (также известный как « Хищник X ») — это вид большого плиозавра, известный по останкам, обнаруженным в Норвегии в 2008 году. Длина этого плиозавра оценивается в 10–13 м (33–43 фута). [297] Однако в 2002 году группа палеонтологов в Мексике обнаружила останки плиозавра, получившего прозвище « Монстр Арамберри », длина которого также оценивается в 15 м (49 футов). [298] с более короткой оценкой около 11,5 м (38 футов). [299] Однако этот вид считается молодым и подвергся нападению более крупного плиозавра. [300] Некоторые источники СМИ утверждали, что Монстр Арамберри был лиоплевродоном, но его вид пока не подтвержден. [298] Еще одним очень крупным плиозавром был Pliosaurus macromerus , известный по единственной неполной нижней челюсти длиной 2,8 метра (9,2 фута). [301] Раннемеловой . Kronosaurus queenslandicus оценивается в 9–10,9 м (30–36 футов) в длину и 10,6–12,1 т (11,7–13,3 коротких тонны) в весе [302] [303] Позднеюрский мегальнейзавр рекс мог достигать длины 7,6–9,1 метра (25–30 футов). [304] [305] Близким соперником по размеру был позднемеловой Megacephalosaurus eulerti длиной 6–9 м (20–30 футов). [306]
Протерозухиды (Proterosuchidae)
[ редактировать ]Proterosuchusfergusi — самый крупный известный протерозухид с длиной черепа 47,7 см (18,8 дюйма) и возможной длиной тела 3,5–4 м (11–13 футов). [307]
Эритрозухиды (Erythrosuchidae)
[ редактировать ]
Самым крупным эритрозухидом был Erythrosuchus africanus с максимальной длиной 4,75–5 м (15,6–16,4 фута). [308]
Фитозавры (Phytosauria)
[ редактировать ]Некоторые из самых крупных известных фитозавров включают редондазавра длиной 6,4 м (21 фут). [309] и Smilosurus длиной более 7 м (23 фута). [310]
Некрокодиломорфные псевдозухи (Pseudosuria)
[ редактировать ]
- Крупнейшим шувосавридом и одним из крупнейших псевдозухий триасового периода был Sillosuchus . Самые крупные экземпляры могли достигать 9–10 м (30–33 футов) в длину. [311] [312]
- Самый крупный известный плотоядный псевдозух триасового периода — лорикатан Fasolasuchus tenax , размер которого оценивается от 8 до 10 м (от 26 до 33 футов). [313] [311] [312] Это одновременно самый крупный « рауизух », известный науке, и самый крупный из когда-либо обнаруженных наземных хищников, не являющихся динозаврами. [ нужна ссылка ] Крупнейшие особи Postosuchus [314] и Заурозух [315] имел длину тела около 7 м (23 фута). Образец Prestosuchus, обнаруженный в 2010 году, позволяет предположить, что это животное также достигало длины почти 7 м (23 фута), что делало его одним из крупнейших псевдозухов триаса. [316]
- Десматозух , вероятно, был одним из крупнейших известных этозавров , около 4–6 м (13–20 футов) в длину и 280 кг (620 фунтов) в весе. [317] [318] [319]
Крокодилы и близкие родственники (Crocodylomorpha)
[ редактировать ]

Эгиптозухиды (Aegyptosuridae)
[ редактировать ]Позднемеловой либо зарегистрированного крокодиломорфа Aegisuchus — главный претендент на звание крупнейшего когда - . По нижней оценке он достигал 15 м (49 футов) в длину и 22 м (72 фута) по верхней оценке. [320] хотя длина более 15 м, скорее всего, завышена. [320]
Крокодилы (Crocodylia)
[ редактировать ]- Самым крупным кайманом и, вероятно, самым крупным крокодилом был Purussaurus brasiliensis, рост которого оценивался в 11–13 м (36–43 фута). [321] По другой информации, максимальная оценка составляет 11,4 м (37 футов) и почти 7,8 т (8,6 коротких тонн) в длину и вес соответственно. [322] Еще одним гигантским кайманом был Муразух . По разным оценкам, самые большие экземпляры достигали 9,47 м (31,1 фута) в длину и 8,5 т (9,4 коротких тонны) в весе. [323] или 4,7–5,98 м (15,4–19,6 футов) в длину тела. [324]
- Самым крупным аллигатороидом, вероятно, является Deinosuchus riograndensis, его длина составляет 12 м (39 футов), а вес - 8,5 т (9,4 коротких тонны). [325] [326]
- Самым крупным вымершим видом рода Аллигатор был аллигатор Хайле ( Alligator hailensis ), который имел череп длиной 52,5 см (20,7 дюйма) и был похож по размеру на современного американского аллигатора ( Alligator Mississippiensis ). [327]
- Самыми крупными гавиалидами были азиатские рамфосухи высотой 8–11 м (26–36 футов). [328] [329] [322] и южноамериканский Gryposuchus длиной 10,15 м (33,3 фута). [330] [329]
- Базальный крокодилоид Astorgosuchus bugtiensis из олигоцена был крупным. Его длина оценивается в 8 м (26 футов). [329]
- Самым крупным известным настоящим крокодилом был Euthecodon , который, по оценкам, достигал 6,4–8,6 м (21–28 футов) или даже 10 м (33 фута) в длину. [331] [322] Самым крупным видом современного Crocodylus был кенийский Crocodylus thorbjarnarsoni длиной 7,56 м (24,8 фута). [332] [322] Танзанийский Crocodylus anthropophagus длиной 7,5 м (25 футов) [333] [334] и неопределенные виды из Кали-Геде ( Ява ) длиной 6–7 м (20–23 фута). [335]
- Самый крупный из известных мекозухий — Paludirex vincenti , длина которого, по оценкам, достигает 5 м (16 футов). [336] [337] [264] Частичный образец челюсти из плиоцена , приписываемый Кинкане, предполагает, что это особь длиной около 6–7 м (20–23 футов). [264] хотя другие виды (известные от олигоцена до плейстоцена ) меньше, их длина составляет около 1,5–3 м (4 фута 11 дюймов – 9 футов 10 дюймов). [338] [339] [340]
Параллигаториды (Paralligatoridae)
[ редактировать ]Самым крупным параллигаторидом , вероятно, был Kansajsuchus, длина которого оценивалась до 8 м (26 футов). [341]
Тетисухи (Tethysuria)
[ редактировать ]- Некоторые вымершие фолидозавриды достигали гигантских размеров. В прошлом саркозух-император считался крупнейшим крокодиломорфом: по первоначальным оценкам его длина составляла 12 м (39 футов), а вес - 8 т (8,8 коротких тонны). [342] Однако, по последним оценкам, длина теперь сократилась до 9–9,5 м (от 29,5 до 31,2 футов) и веса от 3,5 до 4,3 метрических тонн (от 3,9 до 4,7 коротких тонн). [343] Родственный саркозуху , Chalawan thailandicus мог достигать более 10 м (33 футов) в длину. [344] хотя по другим оценкам 7–8 м (23–26 футов). [329]
- Самым крупным дирозавридом был Phosphatosaurus gavialoides, длина которого оценивалась в 9 м (30 футов). [345] [329]
Стоматозухиды (Stomatosuridae)
[ редактировать ]Stomatosurus , стоматозухид , длина которого оценивается в 10 м (33 фута). [346]
Нотосухии (Notosuria)
[ редактировать ]- Одними из крупнейших наземных нотозуховых крокодиломорфов были миоценовый себецид Barinasurus с черепом длиной 95–110 см (37–43 дюйма) и эоценовый себецид Dentaneosurus с предполагаемой длиной нижней челюсти 1 м (3,3 фута). [347] [348] По разным оценкам, возможная длина этих животных составляет 3–10 м (9,8–32,8 футов). Используя часть Stratiotosurus , которая также большая и имеет череп длиной 47 см (19 дюймов), [349] По оценкам, бариназух имеет длину не менее 6,3 м (21 фут). [347] [348]
- Другой огромный нотозух, хотя и известный только по фрагментарному материалу, — это ранний представитель Razanandrongobe , размер черепа которого может превышать размер черепа Barinasuchus , а общая длина может составлять около 7 м (23 фута). [350] [351]
Талаттозухи (Thalattosuria)
[ редактировать ]
- Самый крупный талаттозух , а также самый крупный телеозавроид был безымянным ископаемым остатком из формации Паджа , который может принадлежать животному длиной 9,6 м (31 фут). [352] Это столько же, сколько устаревшая оценка длины раннемелового Machimosaurus rex , длина которой, по последним оценкам, составляла 7,15 м (23,5 фута). [353] Неостенеозавр Эдвардси (ранее известный как Стенеозавр Эдвардси) [354] ) был самым крупным среднеюрским крокодиломорфом, его длина достигала 6,6 м (22 фута). [353]
- Плезиозух был очень крупным метриоринхидом . При длине 6,83 м (22,4 фута) он превосходил даже некоторых плиозавридов того же времени и местности, таких как лиоплевродон . [355] Среди других огромных метриоринхидов - Tyrannoneustes длиной 5 м (16 футов). [356] и Торвонеустес длиной 4,7 м (15 футов). [357]
Базальные крокодиломорфы
[ редактировать ]Редондавенатор был крупнейшим из триасового периода . когда-либо зарегистрированных крокодиломорфов [358] с черепом длиной не менее 60 см (2,0 фута). [359] [360] Еще одним огромным базальным крокодиломорфом был Карнуфекс. [358] длиной 3 м (9,8 футов), хотя он еще незрелый. [361]
Птерозавры (Птерозаврия)
[ редактировать ]
- Самым крупным известным птерозавром был Quetzalcoatlus Northropi , его вес составлял 127 кг (280 фунтов) и размах крыльев 10–12 м (33–39 футов). [362] Еще один близкий соперник — Hatzegopteryx , также с размахом крыльев 12 м (39 футов) и более. [362] Эта оценка основана на черепе длиной 3 м (9,8 фута). [363] Еще одним возможным претендентом на титул является Криодракон , имевший размах крыльев 10 метров (33 фута). [364] Безымянный птеродактилоидный птерозавр из формации Немегт мог достигать размаха крыльев почти 10 м (33 фута). [365] [366] По разным предположениям, размах крыльев Arambourgiania philadelphiae достигал от 8 м (26 футов) до более 10 м (33 фута). [365] [364] Южноамериканский тропеогнат достиг максимального размаха крыльев 8,7 м (29 футов). [367] [368]
- Самый крупный из нептеродактилоидных птерозавров, а также самый крупный юрский птерозавр. [369] был Дирк с предполагаемым размахом крыльев от 2,2 м (7 футов 3 дюйма) до 3,8 м (12 футов). [370] Только фрагментарный экземпляр рамфоринхида из Германии мог быть больше (184% размера самого большого рамфоринха ). [371] Другими крупными нептеродактилоидными птерозаврами были Sericipterus , Campylognathoides и Harpactognathus с размахом крыльев 1,73 м (5 футов 8 дюймов), [372] 1,75 м (5 футов 9 дюймов), [372] и 2,5 м (8 футов 2 дюйма), [371] соответственно.
Хористодеры (Choristodera)
[ редактировать ]Самый крупный известный хористодер , Kosmodraco dakotensis (ранее известный как Simoedosaurus dakotensis). [373] ), по оценкам, имел общую длину около 5 м (16 футов). [374] [373]
Танистрофеиды (Tanystropheidae)
[ редактировать ]
Танистрофей , самый крупный из всех танистрофеидов , достигал до 5 м (16 футов) в длину. [375]
Талаттозавры (Thalattosauria)
[ редактировать ]Самый крупный вид талаттозавра , Miodentosaurus brevis, вырастал до более чем 4 м (13 футов) в длину. [376] Вторым по величине представителем этой группы является Concavispina длиной 3,64 м (11,9 футов). [377]
Ихтиозавры (Ichthyosauria)
[ редактировать ]
В апреле 2018 года палеонтологи объявили об открытии ранее неизвестного ихтиозавра , длина которого могла достигать 26 м (85 футов), что делало его одним из крупнейших известных животных, соперничая с некоторыми синими китами . по размеру [378] [379] Эти останки позже были названы Ихтиотитаном , и, по оценкам, они достигают 25 м (82 фута), что делает его крупнейшим ихтиозавром и самой крупной морской рептилией за всю историю. [380] Еще одним крупным ихтиозавром был позднетриасовый Shastasaurus sikanniensis длиной 21 м (69 футов). [381] [382] и вес 81,5 т (180 000 фунтов). [383] Другой, более крупный ихтиозавр был найден в 1850 году в Аусте . [384] Его останки, казалось, превосходили размеры другого ихтиозавра, но исследователи отметили, что останки были слишком фрагментарными, чтобы можно было оценить их размер. [384] Еще одним огромным ихтиозавром был Shonisaurus Popularis, его длина составляла 15 м (49 футов), а вес - 29,7 т (65 000 фунтов). [382] Самым крупным ихтиозавром среднего триаса, а также самым крупным животным того времени был Cymbospondylus youngorum длиной 17,65 м (57,9 футов). [383] и вес 44,7 т (99 000 фунтов). [383]
Тангасавриды (Tangasauridae)
[ редактировать ]Самым крупным тангасавридом был Ховазавр с предполагаемой длиной морды 30–35 см (12–14 дюймов) и хвостом 60 см (24 дюйма). [385]
Парейазавры (Pareiasauria)
[ редактировать ]Крупнейшие парейазавры достигали 3 м (9,8 футов) в длину. Такие размеры имели среднепермский брадизавр , эмбритозавр и нохелезавр из Южной Африки , [386] и позднепермский скутозавр из России . [386] Самый крепкий скутозавр имел массу тела 1,16 т (2600 фунтов). [386]
Капториниды (Captorhinidae)
[ редактировать ]Тяжелый Moradisaurus grandis длиной 2 м (6 футов 7 дюймов). [387] является крупнейшим известным капторинидом . [388] Вторым по величине капторинидом был лабидозаврикос с самым большим экземпляром черепа взрослой особи длиной 28 см (11 дюймов). [389]
Нептичьи динозавры (Динозаврия)
[ редактировать ]Зауроподоморфы (Sauropodomorpha)
[ редактировать ]Самым крупным из не-зауроподов -зауроподоморфов (« просауроподов ») был Euskelosaurus . Он достигал 12,2 м (40 футов) в длину и 2 т (2,2 коротких тонны) в весе. [390] Другой огромный зауроподоморф Yunnanosaurus youngi достигал 13 м (43 фута) в длину. [391]
Зауроподы (Sauropoda)
[ редактировать ]
- Мега- зауропод Maraapunisaurus fragillimus (ранее известный как Amphicoelias fragillimus ) является претендентом на звание самого крупного известного динозавра в истории. Его максимальная длина оценивается в 58–60 м (190–197 футов) и вес 122 400 кг (269 800 фунтов). [392] К сожалению, ископаемые останки этого динозавра были утеряны. [392] Совсем недавно его длина оценивалась в 35–40 м (115–131 фут) и вес 80–120 т (180 000–260 000 фунтов). [393]
- Известный по неполным и ныне распавшимся останкам, позднемеловой Bruhathkayosaurus matleyi был аномально крупным зауроподом. [394] По неофициальным оценкам, это огромные параметры: 45 м (148 футов) в длину и 139–220 т (306 000–485 000 фунтов) в весе. [395] Однако по некоторым оценкам, его вес составляет 37 м (121 фут) и 95 т (209 000 фунтов), но он все же намного тяжелее, чем большинство других зауроподов. [395] По более поздним оценкам Грегори Пола, сделанным в 2023 году, диапазон его веса составляет от 110 т (240 000 фунтов) до 170 т (370 000 фунтов). Если бы это было правдой, то Брюхаткайозавр стал бы самым крупным наземным животным, когда-либо ходившим по Земле, и мог бы соперничать с самым большим зарегистрированным синим китом. [396]
- BYU 9024, массивный шейный позвонок, найденный в Юте , [397] может принадлежать Barosaurus lentus [398] [399] или Supersaurus vivianae [400] огромного размера, возможно, 45–48 м (148–157 футов) в длину и 60–66 т (132 000–146 000 фунтов) по массе тела. [401] [399] Сам Supersaurus vivianae , возможно, был самым длинным динозавром, когда-либо обнаруженным, поскольку исследование трех экземпляров показало, что длина составляла 39 м (128 футов) или более 40 м (130 футов). [400]

- Mamenchisaurus sinocanadorum, вероятно, был самым крупным маменчизавридом , достигая почти 35 м (115 футов) в длину и 60–80 т (130 000–180 000 фунтов) в весе. [393] Синьцзянтитанские шаньшанеси из той же семьи имели шею длиной 15 м (49 футов), что составляло около 55% ее общей длины, которая могла составлять не менее 27 м (89 футов). [402]
- Среднеюрский был Breviparopus taghbaloutensis упомянут в Книге рекордов Гиннеса как самый длинный динозавр длиной 48 м (157 футов), хотя это животное известно только по ископаемым следам . [403] [404] Первоначально считавшийся брахиозавридом , позже он был идентифицирован как огромный диплодокоид , возможно, 33,5 м (110 футов) в длину и 62 т (137 000 фунтов) в весе. [405]

- Самым высоким зауроподом был Sauroposeidon proteles , предполагаемая высота которого составляла 16,5–18 м (54–59 футов). [406] [407] [408] Азиатозавр мог достигать 17,5 м (57 футов) в высоту, но это животное известно только по зубам. [408] Высота жирафа оценивалась в 16 м (52 фута). [409]
Другие огромные зауроподы включают Аргентинозавра , Аламозавра и Пуэртазавра с предполагаемой длиной 30–33 м (98–108 футов) и весом 50–80 т (55–88 коротких тонн). [410] Длина Патаготитана оценивалась в 37 м (121 фут). [411] и 57 т (63 коротких тонны) в среднем весе, [412] и был похож по размеру на аргентинозавра и пуэртазавра . [413] Гигантские зауроподы, такие как Суперзавр , Зауропосейдон и Диплодок , вероятно, соперничали с ними по длине, но не по весу. [392] Вес Дредноута оценивался в 49 т (108 000 фунтов). [412] и 26 м (85 футов) в длину, но самая полная особь на момент смерти была неполовозрелой. [414] Туриазавр считается крупнейшим динозавром Европы. [415] [416] ориентировочной длиной 30 м (98 футов) и массой 50 т (55 коротких тонн). [410] [416] Однако при более низкой оценке в 21 м (69 футов) и 30 т (66 000 фунтов) он был меньше португальского лусотитана , который достигал 24 м (79 футов) в длину и 34 т (75 000 фунтов) в весе. [417]
Многие крупные зауроподы до сих пор не названы и могут соперничать с нынешними рекордсменами:
- « Архиепископ », крупный брахиозавр, обнаруженный в 1930 году. По состоянию на октябрь 2023 года. [update], научная работа по этому образцу все еще находится в стадии разработки. [418]
- Brachiosaurus nougaredi — еще один крупный брахиозавр из раннемеловой Северной Африки. Останки утеряны, но рисунок крестца остался. Это предполагает крестец длиной почти 1,3 м (4,3 фута). [419] что делает его самым большим крестцом динозавра, обнаруженным до сих пор, за исключением крестца аргентинозавра и апатозавра . [420]
- В 2010 году во Франции была обнаружена бедренная кость крупного зауропода. Бедренная кость предполагает животное, которое выросло до огромных размеров. [421]
Нептичьи тероподы (Theropoda)
[ редактировать ]
- Самый крупный теропод , а также самый крупный наземный (или, возможно, полуводный) [422] Известный на данный момент хищник - Spinosaurus aegyptiacus , самый крупный из известных экземпляров оценивается в 12,6–18 м (41–59 футов) в длину и около 7–20,9 т (8–23 коротких тонны) в весе. [423] [424] Новые оценки, опубликованные в 2014 и 2018 годах, основанные на более полном образце, подтвердили, что спинозавр мог достигать длины от 15 до 16 метров (от 49 до 52 футов). [425] [426] [427] По последним оценкам, вес составляет от 6,4 до 7,5 метрических тонн (от 7,1 до 8,3 коротких тонны). [426] [427] Спинозавр Уайт Рок имел позвонки, по размерам сравнимые со спинозавром , вероятно, это был огромный теропод длиной более 10 м (33 фута). [428]
- Другими крупными тероподами были Giganotosaurus carolinii и Tyrannosaurus rex , чьи самые крупные известные экземпляры оценивались в 13,2 м (43 фута). [429] и 12,3 м (40 футов) [430] в длину и весить от 4,2 до 13,8 т (от 4,6 до 15,2 коротких тонн) [431] [432] [433] [423] и 4,5 метрических тонны (5,0 коротких тонн) [434] [435] до более 7,2 метрических тонн (7,9 коротких тонн), [430] соответственно. Некоторые другие известные гигантские тероподы (например, Кархародонтозавр , Акрокантозавр и Мапузавр ), возможно, также соперничали с ними по размеру.
- Macroelongatoolithus длиной от 34 до 61 см (1,12–2,00 фута), [436] Это самый крупный из известных типов яиц динозавров . [437] Его относят к овирапторозаврам, таким как Бэйбейлонг . [437]
Бронированные динозавры (Thyreophora)
[ редактировать ]Самым крупным из известных тиреофоров был анкилозавр, его длина составляла 9 м (30 футов), а вес - 6 тонн (6,6 коротких тонн). [438] [439] Стегозавр также имел длину 9 м (30 футов). [416] но весит около 5 тонн (5,5 коротких тонн). [ нужна ссылка ]
Пахицефалозавры (Pachycephalosauria)
[ редактировать ]Самым крупным пахицефалозавром был одноименный Pachycephalosaurus . Ранее утверждалось, что его длина составляет 7 м (23 фута), [416] Позже было оценено, что его длина составляет около 4,5 метров (14,8 футов), а вес - около 450 кг (990 фунтов). [440]
Цератопсы (Ceratopsia)
[ редактировать ]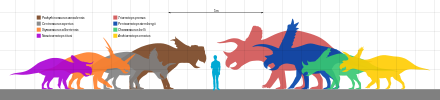
Самый крупный из цератопсов известных — Triceratops horridus , а также близкородственный ему Eotriceratops xerinsularis , длина каждого из которых оценивается в 9 м (30 футов). Пентацератопс и некоторые другие цератопсы соперничают с ними по размеру. [441] Титаноцератопс имел один из самых длинных черепов среди наземных животных - 2,65 м (8,7 футов) в длину. [442]
Орнитоподы (Ornithopoda)
[ редактировать ]
- Самые крупные известные орнитоподы , такие как шантунгозавр, были такими же тяжелыми, как и зауроподы среднего размера, до 23 т (25 коротких тонн). [443] [444] и 16,6 м (54 фута) в длину. [443] Магнапаулия достигала 12,5 м (41 фут) в длину. [445] или, согласно оригинальному описанию, даже 15 м (49 футов). [446] [416] Монгольский . зауролоф , S. angustirostris , достигал 13 м (43 фута) в длину и, возможно, больше [447] Такое животное могло весить до 11 т (12 коротких тонн). [447] Самый крупный эдмонтозавр достигал 12 м (39 футов) в длину и около 6 т (6,6 коротких тонн) массы тела. [448] Предполагаемая максимальная длина брахилофозавра составляет 11 м (36 футов), а вес - 7 т (7,7 коротких тонн). [449] PASAC-1, неофициальное название « Сабинозавр » , является крупнейшим известным североамериканским зауролофином. [450] около 11 м (36 футов) в длину, что примерно на 20% больше, чем у других известных экземпляров. [451] Hypsibema Missouriensis имела длину до 10,7 м (35 футов). [452] [453] Позднемеловой харонозавр . имел длину около 10 м (33 фута) и вес 5 т (5,5 коротких тонн) [416] [454]
- Самым крупным орнитоподом за пределами Hadrosauroidea, вероятно, был игуанодон . Самые крупные экземпляры достигали 11 м (36 футов) в длину. [455] [456] и весил около 4,5 т (5,0 коротких тонн). [457] Еще один крупный орнитопод — Iguanacolossus , длиной 9 м (30 футов) и весом 5 т (5,5 коротких тонн). [458] [459]
- Самым крупным рабдодонтидом был Matheronodon , длина которого оценивалась в 4,8 м (16 футов). [460] рабдодон достигал примерно 4 м (13 футов) и 250 кг (550 фунтов). По оценкам 2016 года, [461]
Птицы (Авес)
[ редактировать ]
Самой крупной птицей в летописи окаменелостей, возможно, является вымерший слоновых птиц вид Aepyornis maximus с Мадагаскара , ближайшим живым родственником которого является киви . Гигантские птицы-слоны превышали 2,3 метра (7,5 футов) в высоту и в среднем весили 850 кг (1870 фунтов). [462]
Самой крупной птицей был михирунг Dromornis stirtoni. Австралии . Его высота превышала 2,7 м (8,9 футов), а средняя масса составляла 500 кг (1100 фунтов). [463]
Другим претендентом является Brontornis burmeisteri , вымершая нелетающая птица из Южной Америки , достигшая веса 319 кг (703 фунта) и высоты примерно 2,8 метра (9,2 фута). [464]
Самой высокой зарегистрированной птицей был Pachystruthio dmanisensis , родственник страуса . Этот конкретный вид птиц имел рост 3,5 метра (11,5 футов) и среднюю массу 450 кг (990 фунтов). [465]
Самым крупным известным нелетающим неоаве была птица-ужас Paraphysornis brasiliensis. Южной Америки,Бразильская птица - террор весила более 240 кг (530 фунтов), [466]
Таблица самых тяжелых вымерших видов птиц
[ редактировать ]Энантиорниты (Enantiornithes)
[ редактировать ]Одним из крупнейших энантиорнитов был Энантиорнис . [487] при жизни около 78,5 см (30,9 дюйма), высоте бедер 34 см (13 дюймов), весе 6,75 кг (14,9 фунта), [488] и размах крыльев сравним с некоторыми современными чайками, около 1,2 м (3 фута 11 дюймов). [487] Гурилиния была самой крупной мезозойской птицей из Монголии : ее длина составляла 53 см (21 дюйм), высота бедер 23,2 см (9,1 дюйма) и вес 2,1 кг (4,6 фунта). [488]
Авизавриды
[ редактировать ]
Позднемеловой был почти такого же размера , авизавр как энантиорнис . У него был размах крыльев около 1,2 м (3 фута 11 дюймов). [487] длина 72 см (28 дюймов), высота бедер 31,5 см (12,4 дюйма) и вес 5,1 кг (11 фунтов). [488] Еще более крупным мог быть Сороавизавр . Одна большеберцовая кость (PVL-4033) указывает на животное длиной 80 см (31 дюйм), высотой бедра 35 см (14 дюймов) и весом 7,25 кг (16,0 фунтов). [488] Однако, по данным Уокера и Дайка (2009), которые считали PVL-4033 Мартинависом sp., длина его большеберцовой кости составляет 85,6 мм (3,37 дюйма), [489] намного короче, чем у Lectavis (156 мм (6,1 дюйма) большеберцовой кости) [490] длина которого в той же книге оценивается в 41 см (16 дюймов), высота бедер 30 см (12 дюймов) и вес 1,15 кг (2,5 фунта) . [488] Мирарс по размеру был сравним с индейкой , намного крупнее большинства других энантиорнитов. [491]
Пенгорнитиды
[ редактировать ]Одной из крупнейших энантиорнитиновых птиц раннего мела был Пенгорнис, его длина составляла 50 см (1,6 фута). [416] и длина черепа 54,7 мм (2,15 дюйма). [492]
Гаргантуавииды
[ редактировать ]Гаргантуавис — крупнейшая известная птица мезозоя , размером от казуара до страуса и массой 140 кг (310 фунтов), как у современных страусов. [493] В 2019 году образцы MDE A-08 и IVPP-V12325 имели длину 1,8 м (5 футов 11 дюймов), высоту бедер 1,3 м (4 фута 3 дюйма) и вес 120 кг (260 фунтов). [473]
Дроморнитиформные
[ редактировать ]
Самым крупным дроморнитидом был Dromornis stirtoni высотой более 3 м (9,8 футов). [494] и масса 528–584 кг (1164–1287 фунтов) для мужчин. [495]
Гасторнид (Gastornithiformes)
[ редактировать ]Крупные особи Гасторниса достигали 2 м (6 футов 7 дюймов) в высоту. [496] Вес гасторниса колеблется от 100 кг (220 фунтов) до 156 кг (344 фунтов), а иногда и до 180 кг (400 фунтов) для европейских экземпляров и от 160 кг (350 фунтов) до 229 кг (505 фунтов) для североамериканских экземпляров. [497] [471] [498]
Водоплавающие птицы (Anseriformes)
[ редактировать ]
Вероятно, нелетающий, миоценовый Garganornis ballmanni был крупнее любых ныне живущих представителей Anseriformes , с массой тела 15,3–22,3 кг (34–49 фунтов). [499] Еще одним огромным гусеобразным был нелетающий новозеландский гусь ( Cnemiornis ). Он достигал 15–18 кг (33–40 фунтов), приближаясь по размеру к мелким видам моа. [500]
Лебеди (Цигнини)
[ редактировать ]Самым крупным известным лебедем был плейстоцена гигантский лебедь ( Cygnus falconeri ), длина которого от клюва до хвоста достигала около 190–210 см (75–83 дюйма). [501] вес около 16 кг (35 фунтов) и размах крыльев 3 м (9,8 футов). [502] [503] [504] Новозеландский лебедь ( Cygnus sumnerensis ) весил до 10 кг (22 фунта), по сравнению с родственным ему современным черным лебедем , который весил всего 6 кг (13 фунтов). [505] Крупный морской лебедь Annakacygna yoshiiensis из миоцена Японии как по размеру , намного превосходил современного лебедя-шипуна так и по весу. [506]
Анатина
[ редактировать ]Утка Финша ( Chenonetta finschi ) достигала веса 1–2 кг (2,2–4,4 фунта), превосходя родственную современную австралийскую древесную утку (800 г (1,8 фунта)). [507]
Пеликаны, ибисы и их союзники (Pelecaniformes)
[ редактировать ]- Раннеплиоценовый Pelecanus schreiberi был крупнее большинства современных пеликанов. Pelecanus odessanus из позднего миоцена, вероятно, был того же размера, что и P. schreiberi , его предплюсна имеет длину 150 мм (5,9 дюйма). [508]
- Самая крупная цапля Цапля Бенну ( Ardea bennuides ). Судя по обнаруженным останкам, она имела рост примерно 2 м (6,6 фута) и размах крыльев до 2,7 м (8,9 фута), что превосходило по размерам самый крупный из ныне живущих видов семейства цапельных. , цапля-голиаф . [509]
- Самый крупный ибис Ямайский ибис ( Xenicibis xympithecus ) в виде большого ибиса весом около 2 кг (70 унций).
Аисты и союзники (Ciconiiformes)
[ редактировать ]
Самым крупным из известных представителей Ciconiiformes был Leptoptilosrobustus , ростом 1,8 м (5 футов 11 дюймов) и весом около 16 кг (35 фунтов). [510] [483] Ciconia Maltha — относительно крупный вид Ciconia , высотой более 5 футов (1,5 метра) и размахом крыльев до 10 футов (3,0 метра) в поперечнике. [511]
Журавли (Gruiformes)
[ редактировать ]Крупный настоящий журавль ( Gruinae ) из позднего миоцена ( тортонского века ) Германии был равен по размерам самым крупным из ныне живущих журавлей и напоминал длинноклювого стерха ( Leucogeranus leucogeranus ). [512]
Береговые птицы (Ржакообразные)
[ редактировать ]Miomancalla Howardi была крупнейшей известной ржанкообразной всех времен, ее вес был примерно на 0,6 кг (1,3 фунта) больше, чем у второй по величине особи, большой гагарки ( Pinguinus impennis ). [513]
Гесперорнитины (Hesperornithes)
[ редактировать ]Самым крупным из известных гесперорнитинов был Canadaga arctica , его длина составляла 2,2 м (7 футов 3 дюйма). [514]
Стервятники Нового Света (Cathartiformes)
[ редактировать ]
Одной из самых тяжелых летающих птиц всех времен был Аргентавис миоцена , тераторнитид . Огромная птица имела размах крыльев примерно 5,09–6,5 м (16,7–21,3 фута). [475] [515] и вес от 70 до 72 кг (от 154 до 159 фунтов). [516] [475] была Аргентависа Плечевая кость лишь немного короче всей человеческой руки. [517] Еще одним огромным тераторном был Айолорнис с размахом крыльев около 5 м (16 футов). [518] Плейстоценовый ) Teratornis merriami достигал 13,7 кг (30 фунтов) и 2,94–3,38 м (9,6–11,1 фута) в размахе крыльев, при этом меньшие оценки размера все еще превышали самые крупные экземпляры калифорнийского кондора ( Gymnogyps Californianus . [519]
Сериемы и союзники (Cariamiformes)
[ редактировать ]
Самым крупным из известных когда-либо Cariamiforme и крупнейшим форусрацидом или «ужасной птицей» (очень хищными, нелетающими птицами Америки) был Brontornis , который имел высоту около 175 см (69 дюймов) в холке и мог поднимать голову на 2,8 м (9 футов 2 дюйма). in) над землей и мог весить до 400 кг (880 фунтов). [520] Огромный форусрацид Келенкен имел высоту 3 м (9,8 футов). [521] [522] с черепом длиной 716 мм (28,2 дюйма) (460 мм (18 дюймов) из которых составлял клюв), имел самую большую голову среди всех известных птиц. [521] Южноамериканский Форусракос имел рост 2,4–2,7 м (7,9–8,8 футов) и весил почти 130 кг (290 фунтов), столько же, сколько самец страуса . [523] [524] Самым крупным североамериканским форусрацидом был Титанис , достигавший высоты примерно 2,5 м (8,2 фута). [525] немного выше африканского лесного слона .
Ястребообразные (Accipitriformes)
[ редактировать ]
Самой крупной известной хищной птицей когда-либо был огромный орел Хааста ( Hieraaetus moorei ) с размахом крыльев от 2,6 до 3 м (от 8 футов 6 дюймов до 9 футов 10 дюймов), что было относительно коротким для своего размера. [526] [527] Общая длина, вероятно, составляла до 1,4 м (4 фута 7 дюймов) у самок. [528] и они весили от 10 до 15 кг (от 22 до 33 фунтов). [529] Еще одним гигантским вымершим ястребом был Titanohierax весом около 7,3 кг (16 фунтов), который обитал на Антильских островах и Багамских островах , где он был одним из главных хищников. [530] Безымянный орел позднечетвертичного периода из Эспаньолы мог быть на 15–30% крупнее современного беркута ( Aquila chrysaetos ). [531] Некоторые вымершие виды Buteogallus превосходили по размерам своих ныне живущих сородичей. Buteogallus borrasi был примерно на 33% крупнее современного большого черного ястреба ( B. urubitinga ). [532] B. daggetti , также известный как «ходячий орел», был примерно на 40% крупнее саванного ястреба ( B. meridionalis ). [533] Лунь Эйлса ( Circus eylesi ) из плейстоцена - голоцена Новой Зеландии был более чем в два раза тяжелее современного C. Approsimans . [534]
Моа (Dinornithiformes)
[ редактировать ]Самой высокой известной птицей был гигантский моа Южного острова ( Dinornisrobustus ), представитель семейства моа Новой Зеландии , вымершего около 500 лет назад. Его высота достигала 3,7 м (12 футов). [535] и весил примерно вдвое меньше большой птицы-слона из-за своего сравнительно стройного телосложения. [536]
Тинамус (Tinamiforms)
[ редактировать ]MPLK-03, экземпляр тинаму , существовавший в позднем плейстоцене в Аргентине , возможно, принадлежит к современному роду Eudromia и превосходит по размерам современные E. elegans и E. formosa на 2,2–8% и 6–14% соответственно. [537]
Слоновьи птицы (Aepyornithiformes)
[ редактировать ]Самой крупной птицей в летописи окаменелостей, возможно, являются вымершие птицы-слоны ( Vorombe , Aepyornis ) Мадагаскара, которые были родственниками страуса. Их рост превышал 3 м (9,8 фута) и вес 500 кг (1100 фунтов). [536]
Страусы (Struthioniformes)
[ редактировать ]с массой тела 450 кг (990 фунтов) Pachystruthio dmanisensis из нижнего плейстоцена Крыма . была самой крупной птицей, когда-либо зарегистрированной в Европе Несмотря на свои гигантские размеры, он был хорошим бегуном. [538] Возможный экземпляр Pachystruthio из нижнего плейстоцена провинции Хэбэй ( Китай ) имел вес около 300 кг (660 фунтов), что вдвое тяжелее обыкновенного страуса ( Struthio Camelus ). [539] Останки массивного азиатского страуса ( Struthio asiaticus ) из плиоцена. [540] указывают на размер на 20% больше взрослого самца сохранившегося Struthio Camelus . [541]
Голуби и голуби (Columbiformes)
[ редактировать ]
Самым крупным известным родственником голубя был дронт ( Raphus cucullatus ), возможно, превышающий 1 м (3,3 фута) в высоту и весивший целых 28 кг (62 фунта), хотя недавние оценки показали, что средний дикий дронт весил гораздо меньше, примерно 10,2 кг (22 фунта). [542] [543]
Фазаны, индейки, дичь и их союзники (Galliformes)
[ редактировать ]Самым крупным из известных курообразных , вероятно, была гигантская моллюска , вес которой мог достигать 7 кг (15 фунтов). [544]
Певчие птицы (Воробьиные)
[ редактировать ]Самая крупная известная певчая птица — вымерший гигантский гробоклюв ( Chloridops regiskongi ) длиной 280 мм (11 дюймов). [ нужна ссылка ]
Бакланы и союзники (Suliformes)
[ редактировать ]
- Самым крупным известным бакланом был очковый баклан северной части Тихого океана ( Phalacrocorax perspicillatus ), который вымер около 1850 года и весил в среднем около 6,4 кг (14 фунтов) и 1,15 м (3 фута 9 дюймов). [486]
- Самым крупным известным дартером был Гиганхинга с предполагаемым весом около 17,7 кг (39 фунтов). [481] В более раннем исследовании даже утверждается, что вес составляет 25,7 кг (57 фунтов). [545]
- Самой крупной известной плотоптеридной нелетающей птицей, похожей на пингвина, был Copepteryx titan , длина бедренной кости которого составляет 22 см (8,7 дюйма), что почти вдвое длиннее, чем у императорского пингвина . [546]
Поганки (Podicipediformes)
[ редактировать ]Самая крупная из известных поганок, Атитланская поганка ( Podylimbus gigas ), достигала в длину около 46–50 сантиметров (18–20 дюймов). [547]
Костистозубые птицы (Odontopterygiformes)
[ редактировать ]Самыми крупными известными из Odontopterygiformes — группы, которая по-разному была связана с Procellariiformes, Pelecaniformes и Anseriformes, а самыми крупными летающими птицами всех времен, кроме Argentavis, были огромные Pelagornis , Cyphornis , Dasornis , Gigantornis и Osteodontornis . [ нужна ссылка ] Они имели размах крыльев 5,5–6 м (18–20 футов) и высоту около 1,2 м (3 фута 11 дюймов). [ нужна ссылка ] Точные оценки размера и определение того, какая из них была самой крупной, пока невозможны для этих птиц, поскольку их кости были чрезвычайно тонкостенными, легкими и хрупкими, и поэтому большинство из них известны только по очень неполным останкам. [ нужна ссылка ]
Дятлы и союзники (Piciformes)
[ редактировать ]Самый крупный известный дятел — это, возможно, вымерший императорский дятел ( Campephilus Imperialis ) общей длиной около 56–60 см (22–24 дюйма). [548]
Попугаи (Psittaciformes)
[ редактировать ]Самый крупный известный попугай — вымерший Heracles inexpectatus длиной около 1 метра (3,3 фута). [549]
Пингвины (Sphenisciformes)
[ редактировать ]
Одним из самых тяжелых когда-либо известных пингвинов является Kumimanu fordycei , его масса тела оценивается от 148 до 159,7 кг (от 326 до 352 фунтов), что определяется на основе измерений плечевой кости. [550] Другой пример - Palaeeudyptes klekowski из Антарктиды , длина от клюва до хвоста оценивается в 2,02 м (6 футов 8 дюймов) и предполагаемая масса тела 84,2 кг (186 фунтов), что немного меньше, чем предыдущие оценки. [550] [551] Эоценовый Anthropornis nordenskjoeldi сопоставим по размеру: когда-то считалось, что он достигает длины 2,05 м (6 футов 9 дюймов) и веса 108 кг (238 фунтов). [552] Однако, по недавним оценкам, основанным на измерениях плечевой кости, вес A. nordenskjoeldi оказался в пределах 67 кг (148 фунтов). [550] Другие крупные пингвины включают новозеландского гигантского пингвина ( Pachydyptes pondeorsus ) весом от 65,4 до 94,6 кг (от 144 до 209 фунтов) и Icadyptes salasi от 52,8 до 73,0 кг (от 116,4 до 160,9 фунтов). [553] [550]
Совы (Strigiformes)
[ редактировать ]Самой крупной известной совой всех времен была кубинская орнимегалоникс высотой 1100 мм (43,3 дюйма), вероятно, превышающая 9 кг (20 фунтов). [554]
Земноводные (амфибии)
[ редактировать ]Самой крупной известной амфибией всех времен был темноспондил Prionosuchus длиной 9,1 м (30 футов) . [555]
Лиссамфибии (Lissamphibia)
[ редактировать ]Лягушки и жабы (Анура)
[ редактировать ]
Самая крупная из когда-либо известных лягушек была еще неназванным видом эоцена и имела длину около 58–59,1 сантиметра (22,8–23,3 дюйма). [556] Позднемеловой быка Beelzebufo вырастал как минимум до 23,2 см (9,1 дюйма) (длина морды и отверстия), что примерно соответствует размеру современной африканской лягушки- . [557]
Саламандры, тритоны и союзники (Уродела)
[ редактировать ]
- Андриас Маттеви был самым крупным из когда-либо известных лиссамфибий , его общая длина достигала 2,3 м (7 футов 7 дюймов). [558]
- Хаброзавр был самым крупным сиренидом . Его длина достигала 1,6 м (5 футов 3 дюйма). [559]
Диадектоморфы (Diadectomorpha)
[ редактировать ]
Самый крупный из известных диакектид, травоядный Diadectes , представлял собой тяжелое животное длиной до 3 м (9,8 футов) с толстыми позвонками и ребрами. [560] [561]
Антракозаврия
[ редактировать ]Самым крупным известным антракозавром был Anthracosaurus , с черепом около 40 см (16 дюймов) в длину. [562]
Эмболомеры
[ редактировать ]
Самым длинным членом этой группы был Eogyrinus attheyi , вид, который иногда относят к роду Pholiderpeton . [563] Его череп имел длину около 41 см (16 дюймов). [564]
Темноспондилы (Temnospondyli)
[ редактировать ]
Самая крупная известная темноспондильная амфибия — Prionosuchus , длина которой достигала 9 м (30 футов). [555] Еще одним огромным темноспондилом был Mastodonsaurus giganteus длиной 6 м (20 футов). [565] Безымянный вид темноспондилов из Лесото является частичным, но возможная оценка длины тела составляет 7 м (23 фута). [566]
Рыбы (Рыбы)
[ редактировать ]Рыбы — парафилетическая группа позвоночных, не относящихся к четвероногим .
Бесчелюстная рыба (Агната)
[ редактировать ]Конодонты (Conodonta)
[ редактировать ]По оценкам, Iowagnathus grandis имеет длину более 50 см (1,6 фута). [567]
Гетеростраки (Heterostraci)
[ редактировать ]По оценкам, некоторые представители Psammosteidae, такие как Obruchevia и Tartuosteus, достигают высоты до 2 м (6,6 футов). [568]
Телодонты (Thelodonti)
[ редактировать ]) известен по частичным материалам, Хотя Thelodus parvidens (= T. macintoshi по оценкам, он достигает 1 м (3,3 фута). [569]
Цефаласпидоморфы (Cephalaspidomorphi)
[ редактировать ]Разновидность Parameteoraspis достигала 1 м (3,3 фута). [570]
Spiny sharks (Acanthodii)
[ редактировать ]Самым крупным из ныне вымерших Acanthodii был Xylacanthus grandis , ишнакантиформный вид , основанный на челюстной кости длиной около 35 см (14 дюймов). Судя по пропорциям его родственника Ischnacanthus , общая длина X. grandis составляла 2,5 м (8 футов 2 дюйма). [571]
Плакодермы (Placodermi)
[ редактировать ]
Самой крупной известной плакодермой был гигантский хищный Дунклеостей . Самым крупным и наиболее известным видом был D. terrelli , по разным оценкам, его длина составляла около 4,1–10 м (13,5–32,8 футов) в длину и 1–4 т (1,1–4,4 коротких тонны) в весе. [572] Другая крупная плакодерма, Titanichthys , возможно, соперничала с ним по размеру. [573] По оценкам, титанихтис имеет длину около 4,1–7,5 м (13–25 футов). [574] [575] [576] [572]
Хрящевая рыба (Chondrichthyes)
[ редактировать ]Скумбрия-акулы (Lamniformes)
[ редактировать ]
- Виды вымершего рода Otodus были огромными. Гигантская акула . Otodus megalodon [577] [578] [579] на сегодняшний день является самой большой когда-либо известной акулой-макрелью . [580] Большинство оценок размера мегалодона экстраполируются на основе зубов, при этом максимальная длина оценивается до 10,6–20 м (35–66 футов). [578] [579] [581] и средняя оценка длины 10,5 м (34 фута). [582] [583] Из-за фрагментарных останков было сделано много противоречивых оценок размера мегалодона, поскольку их можно получить только по ископаемым зубам и позвонкам. [584] : 87 [585] Взрослый самец мегалодона мог иметь массу тела от 12,6 до 33,9 метрических тонн (от 13,9 до 37,4 коротких тонн), а взрослые самки могли иметь массу тела от 27,4 до 59,4 метрических тонн (от 30,2 до 65,5 коротких тонн), предполагая, что длина самцов могла варьироваться от От 10,5 до 14,3 м (от 34 до 47 футов) и у женщин от 13,3 до 17 м (от 44 до 56 футов). [586] Родственные мегалодону Otodus angustidens и O. chubutensis крупные размеры достигали . Высота каждого оценивалась в 9,3 м (31 фут). [587] и 12,2 м (40 футов), [588] соответственно.
- Другими гигантскими макрельевыми акулами были Pseudoscapanorhynchidae из мелового периода. Cretodus Размер составлял 9–11 м (30–36 футов) (для C. crassidens ), [589] Лептостиракс достигал длины 6,3–8,3 м (21–27 футов). [590]
- Кайнозойский . Parotodus достигал 7,6 м (25 футов) в длину [591]
- Самой тяжелой акулой-молотилкой, вероятно, была Alopias grandis . По размерам она была похожа или даже крупнее ныне живущей большой белой акулы и, вероятно, не имела удлиненного спинного хвоста, характерного для современных сородичей. [592]
Сухопутные акулы (Carcharhiniformes)
[ редактировать ]Кайнозойский . Hemipristis serra был значительно крупнее своих современных родственников и имел гораздо более крупные зубы Его общая длина оценивается в 6 метров (20 футов). [593]
Гибодонты (Hybodontiformes)
[ редактировать ]Одним из крупнейших гибодонтиформ был юрский астеракант с длиной тела до 3 м (9,8 футов). [594] Crassodus reifi известен из меньшего количества материалов, однако, по оценкам, он достигал более 3 м (9,8 футов). [595]
Ктенакантиформные
[ редактировать ]Самый крупный представитель гребневиков - Saivodus striatus , предполагаемая длина которого составляет около 6–9 м (20–30 футов). [596] [572]
Коньки и союзники (Rajiformes)
[ редактировать ]Гигантский склероринхид Onchopristis достигал около 4,25 м (13,9 футов) в длину. [597]
Евгенеодонт (Eugeneodontida)
[ редактировать ]
Самый крупный из известных эвгенеодонтов — пока еще неназванный вид Helicoprion , обнаруженный в Айдахо. Судя по образцам, это животное, длина которого, возможно, превышала 12 м (39 футов). [598] Другой довольно крупный эвгенеодонт — Парагеликоприон . Будучи более тонким, чем Геликоприон , он достиг почти такого же размера. [598] возможно, до 12 м (39 футов) в длину. [599] Оба имели самые большие размеры среди животных палеозойской эры. [600] [599]
Лопастеперая рыба (Sarcopterygii)
[ редактировать ]Целаканты (Actinistia)
[ редактировать ]
Самый крупный целакант — Mawsonia gigas мелового периода , общая длина которого оценивается до 5,3 м (17 футов). Юрский трахиметопон , возможно, достигал близкого к этому размера - около 5 м (16 футов). [601] Неопределенный мавсониид из маастрихтских отложений Марокко, вероятно, достигал 3,65–5,52 м (12,0–18,1 футов) в длину. [602] [601]
Двоякодышащая рыба (Дипной)
[ редактировать ]Меловой Ceratodus sp. По оценкам, из Западного интерьера его длина составляла около 4 м (13 футов). [603]
Стеблевые четвероногие (Tetrapodomorpha)
[ редактировать ]

- Не только самым крупным известным ризодонтом , но и самой крупной рыбой с лопастными плавниками был Rhizodus длиной 5,63–7 м (18,5–23,0 фута) . [604] [572] Другими крупными ризодонтами были Strepsodus с предполагаемой длиной около 3–5 м (9,8–16,4 футов) и Barameda с предполагаемой длиной 3–4 м (9,8–13,1 футов). [605] [606]
- Тристихоптерид Hyneria достигал длины до 3,5 м (11 футов). [572]
Лучеперая рыба (Actinopterygii)
[ редактировать ]Осетровые
[ редактировать ]- Gyrosteus , принадлежащий к вымершему остроконечных семейству Chondrosteidae , по оценкам, имеет стандартную длину около 6–7 м (20–23 футов). [607]
- Самый крупный известный ископаемый осетр - Acipenser gigantissimus , известный по фрагментарным останкам, длина которого, по оценкам, достигает 5,8 м (19 футов). [608]
- Самый крупный из известных ископаемых веслоносов - это безымянные останки из формации Джудит-Ривер , их размер может превышать 2 м (6 футов 7 дюймов), а известные останки превышают размер недавно вымершего китайского веслоноса , размер которого, по научным данным, превышает 3 м (9,8 футов). [609]
Пахикормиформные
[ редактировать ]
Самой крупной известной лучепёрой рыбой и самой крупной костной рыбой всех времён была пахикормида Leedsichthyspromaticus . длиной около 16,5 м (54 фута) [610] По более ранним оценкам, речь шла о более крупных особях длиной более 27 м (89 футов). [611] [612]
Ихтиодектиформы
[ редактировать ]
Самой крупной известной ихтиодектиформной рыбой был Xiphactinus , длина которого достигала 6,1 м (20 футов). [613] Ихтиодекты достигали 3 м (9,8 футов) в длину, что вдвое меньше Xiphactinus . [614]
Пикнодонтиформы
[ редактировать ]Самым крупным из известных пикнодонтиформ был Gyrodus roundis длиной до 2 м (6 футов 7 дюймов). [615]
Бичирсы (Polypteriformes)
[ редактировать ]Позднемеловой вероятно Бавитиус, , был самым крупным бичиром всех времен. Его длина достигала 3 м (9,8 футов). [616]
Опахи, ленточные рыбы, весла и их союзники (Lampriformes)
[ редактировать ]Мегаламприс, вероятно, был крупнейшей ископаемой опой . В живом состоянии эта рыба имела длину около 4 м (13 футов), что в два раза превышает длину самого крупного из ныне живущих видов опаха, Lampris Guttatus . [617]
Лосось и форель (Salmoniformes)
[ редактировать ]Самым крупным лососем был Oncorhynchus rastrosus , размер которого варьировался от 1,9 м (6 футов 3 дюйма) до 177 кг (390 фунтов). [618] до 2,4 м (7 футов 10 дюймов) и 200 кг (440 фунтов). [619] [618]
Иглобрюхи, самшиты, спинороги, океанические солнечные рыбы и их союзники (Tetraodontiformes)
[ редактировать ]- Austromola angerhoferi имела общую длину тела около 3,2 м (10 футов) и общую высоту 4 м (13 футов), что сравнимо с крупнейшей океанской солнечной рыбой . [620] [621]
- По оценкам, некоторые вымершие виды Balistes , такие как B. vegai и B. crassidens, имеют общую длину до 1,8 м (5 футов 11 дюймов). [622]
Рыбы-ящерицы (Aulopiformes)
[ редактировать ]Самой крупной рыбой-ящерицей был Stratodus , длина которого могла достигать 5 м (16 футов). [623]
Иглокожие (Echinodermata)
[ редактировать ]Кринозоа
[ редактировать ]
Морские лилии (Crinoidea)
[ редактировать ]Самый длинный стебель Seirocrinus subangularis достигал более 26 м (85 футов). [624]
Астероид
[ редактировать ]Морская звезда (астероид)
[ редактировать ]Гелиантастер из сланца Хунсрюк имел радиус около 25 см (9,8 дюйма). [625]
Граптолиты (Graptolithina)
[ редактировать ]Самый длинный известный граптолидный граптолит - Stimuloraptus Halli, его длина составляет 1,45 м (4,8 фута). Он найден в силурийских отложениях Соединенного Королевства . [626]
Киноринчи (Kinorhyncha)
[ редактировать ]Кембрийские киноринхи из биоты Цинцзян , также известные как «грязевые драконы», достигали 4 см (1,6 дюйма) в длину, что намного крупнее современных родственников, которые вырастают всего на несколько миллиметров в длину. [627] [628]
Членистоногие (Arthropoda)
[ редактировать ]динокаридид
[ редактировать ]Жаберные лобоподии
[ редактировать ]
Судя по исследованиям ротового аппарата, кембрийский жаберный лобопод Omnidens amplus оценивается в 1,5 метра (4,9 фута). [629] Он также известен как самое крупное из когда-либо существовавших кембрийских животных. [629]
Радиодонт (Radiodonta)
[ редактировать ]
Самый крупный из известных радиодонтов — Aegirocassis benmoulai , длина которого, по оценкам, составляла не менее 2 м (6 футов 7 дюймов). [630] [631]
Хелицерата
[ редактировать ]Морские пауки (Pycnogonida)
[ редактировать ]Самый крупный ископаемый морской паук — Palaeoisopuspromaticus с размахом ног около 32 см (13 дюймов). [632]
Мечехвосты и их союзники (Xiphosura)
[ редактировать ]- Willwerathia достигала 9 см (3,5 дюйма) в ширину панциря и была самым крупным видом базальных (« синзифосуриновых ») ксифосуран . [633] [634] Однако девонская Мальдыбулакия достигала почти 11,5 см (4,5 дюйма). [635] и был отнесен к ксифосуранам в 2013 году. [634]
- тропа мечехвоста Икноископаемая Kouphichnium Litographicum из Церина в Айне указывает на длину животного 77,4–85,1 см (30,5–33,5 дюйма). [636]
Часматаспидиды (Chasmataspidida)
[ редактировать ]
Самыми крупными хасматаспидидами были ордовикские Hoplitaspis длиной 29 см (11 дюймов) и аналогичные по размеру Chasmataspis . [637]
Эвриптериды (Eurypterida)
[ редактировать ]
- Самым крупным известным эвриптеридом был Jaekelopterus rhenaniae длиной 2,5 м (8 футов 2 дюйма), который также является самым крупным из известных членистоногих. [638] Erettopterus grandis , возможно, достиг такой же длины, но это основано только на неполном тельсоне . Близким соперником был Acutiramus bohemicus длиной 2,1 м (6 футов 11 дюймов). [639] Самым крупным мегалограптидом , а также крупнейшим ордовикским эвриптидом был Pentecopterus . Его длина достигала 1,7 м (5 футов 7 дюймов). [640] Все это были эвриптериновые эвриптериды. [639] [640]
- Самым крупным стилонуриновым эвриптеридом был Hibbertopterus , его длина составляла 1,8 м (5 футов 11 дюймов). [639]
Паукообразные (Arachnida)
[ редактировать ]- Есть три претендента на звание самого крупного из известных паукообразных, а также крупнейших скорпионов всех времен: Pulmonoscorpius kirktonensis , Brontoscorpio anglicus и Praearcturus gigas . По оценкам, каждый из них был 70 см (28 дюймов), [641] 90 см (35 дюймов) [642] и до 100 см (39 дюймов), [643] соответственно.
- Mongolarachne jurassica — самый крупный из описанных ископаемых пауков : общая длина тела самки составляет примерно 24,6 мм (0,97 дюйма), а длина передних ног — около 56,5 мм (2,22 дюйма). [644] Dinodiplura ambulacra имела большую длину тела, общая длина панциря и опистосомы достигает 26,15 мм (1,030 дюйма). [645]
- Самый крупный из доисторических скорпионов и, возможно, самый крупный из когда-либо обнаруженных скорпионов. [646] был Mesoproctus rayoli . У типового вида длина тела достигала 66,9 мм (2,63 дюйма) с панцирем 25,7 мм (1,01 дюйма) в длину, тогда как у другого экземпляра панцирь имел длину 32,5 мм (1,28 дюйма) и ширину 16 мм (0,63 дюйма). сопоставимы или даже больше, чем у современных мастигопроктусов . [647] [648]
- Самым крупным рицинулей из когда-либо существовавших был Curculioides bohemondi с длиной тела 21,77 мм (0,857 дюйма). [649]
- Самым крупным ископаемым акариформным клещом , а также самым крупным когда-либо зарегистрированным эритреоидным клещом был Immensmaris Chewbaccei с идиосомой более 8 мм (0,31 дюйма) в длину. [650]
- Самым крупным известным тригонотарбидом была Kreischeria с минимальной длиной 51 мм (2,0 дюйма). [651] Вторым по величине был Pleophrynus длиной 36 мм (1,4 дюйма). [651]
Парнокопытные (Artiopoda)
[ редактировать ]Ретифации, вероятно, достигали 55 см (22 дюйма). [652] Тегопельте - еще один пример крупных парнокопытных, не относящихся к трилобитам, достигавших 280 мм (11 дюймов) в длину. [653] и был самым крупным из билатерий Берджесс-Шейла , превосходя все другие донные организмы как минимум вдвое. [653]
Трилобиты (Trilobita)
[ редактировать ]Некоторые из трилобитов превышали 60 см (24 дюйма) в длину. Почти полный экземпляр Isotelus rex из Манитобы достигал длины более 70 см (28 дюймов), а Ogyginus forteyi из Португалии был почти такой же длины. Фрагменты трилобитов предполагают еще большие рекордные размеры. Изолированный пигидий Hungioides bohemicus предполагает, что длина всего животного составляла 90 см (35 дюймов). [654]
Многоножки (Myriapoda)
[ редактировать ]
Самым крупным из известных многоножек на сегодняшний день была Arthropleura . Длина 2,5 метра (8 футов 2 дюйма). [655] и шириной 50 сантиметров (20 дюймов). [656] Некоторые экземпляры могли быть еще больше: до 2,63 метра (8 футов 8 дюймов) в длину и 50 кг (110 фунтов) в весе. [657] [658]
Нешестиногие ракообразные (Crustacea)
[ редактировать ]Циклоиды (Cyclida)
[ редактировать ]Самой крупной циклидой была Opolanka Decorosa , позднетриасовая галициноподобная которой достигал более 6 см (2,4 дюйма) циклоида, размер панциря . [659]
Ремипеды (Ремипедия)
[ редактировать ]Tesnusocaris имел длину тела не менее 9,5 см (3,7 дюйма), [660] крупнее всех ныне живущих ремипедов , их длина может достигать 4,5 см (1,8 дюйма). [661]
Насекомые (Insecta)
[ редактировать ]Пилильщики, осы, пчелы, муравьи и их союзники (Hymenoptera)
[ редактировать ]
- Самым крупным из известных представителей этой группы был гигантский муравей Titanomyrma giganteum , чьи королевы вырастали до 6 см (2,4 дюйма). Размах его крыльев составлял 15 см (5,9 дюйма). [662]
- Apislithohermaea — одна из самых крупных когда-либо найденных медоносных пчел, по размеру сравнимая с современной Apis dorsata . [663]
- Гигантский рогохвост Ypresiosirex Orthosemos достигал 67,9 мм (2,67 дюйма) в длину, включая неполный яйцеклад . [664] Другой пример гигантского пилильщика — Hoplitolyda duolunica с размахом крыльев более 92 мм (3,6 дюйма). [665]
Блохи (Siphonaptera)
[ редактировать ]Самым крупным из известных представителей Siphonaptera был, вероятно, Pseudopulex magnus , выраставший до 0,90 дюйма (22,8 мм) в длину. [666]
Уховертки (Dermaptera)
[ редактировать ]
Вымерли совсем недавно, после 1967 года. [667] [668] а также представлены как голоцена окаменелости , [669] гигантская уховертка острова Святой Елены ( Labidura herculeana , синоним Labidura loveridgei ) достигала 84 мм (3,3 дюйма) в длину, включая щипцы длиной 34 мм (1,3 дюйма). [667]
Хресмодиды
[ редактировать ]У Chresmodidae были длинные специализированные ноги, как у современного семейства Gerridae . Один из Chresmodidae, Chresmoda obscura , мог достигать размера около 19 сантиметров (7,5 дюйма). [670]
Жуки (Coleoptera)
[ редактировать ]крупнейших известных ископаемых жуков надсемейства Одним из Scarabaeoidea является Protognathinus spielbergi . Его общая длина, включая челюсти, составляла около 5,5 см (2,2 дюйма). [671] Самым крупным ископаемым скарабеидом был Oryctoantiquus Borealis с предполагаемой длиной тела 5 сантиметров (2,0 дюйма). [672]
Титаноптераны (Titanoptera)
[ редактировать ]
Родственные современным периода были намного крупнее прямокрылым, титаноптераны триасового . Размах крыльев Gigatitan vulgaris достигал 40 сантиметров (16 дюймов). [673] Клатротитан андерсони также достиг огромных размеров: переднее крыло имело длину 13,8 сантиметра (5,4 дюйма). [674]
Муравьиные львы и родственные им сетчатокрылые насекомые (Neuroptera)
[ редактировать ]По оценкам, Makarkinia adamsi из формации Крато имеет самые длинные передние крылья среди всех видов нейроптерых , их длина оценивается в 160 мм (6,3 дюйма). [675]
Тараканы, термиты, богомолы и их союзники (Dictyoptera)
[ редактировать ]- Некоторые тараканоподобные насекомые каменноугольного периода группируются в Blattoptera, например Archoblattina beecheri. [676] и Necymylacris (Xenoblatta) scudderi. [677] [678] Общая длина могла достигать около 9 сантиметров, что сравнимо с современной Megaloblatta longipennis .
- Меловой таракан Ptiloteuthis foliatus имел крыло длиной 7,9 см (3,1 дюйма). [679]
- обнаруженный в миоцене в Австрии Гигантский термит Gyatermes styriensis, , достигал 25 мм (0,98 дюйма) в длину тела и имел размах крыльев 76 мм (3,0 дюйма). [680]
Стрекозы, стрекозы и грифоны (Odonatoptera)
[ редактировать ]
- Самым крупным известным насекомым -одонатоптерокрылом был Meganeuropsis permiana с одним крылом длиной 33 см (13 дюймов). Меганевра имела крыло длиной 32 см (13 дюймов). [681]
- Triadotypid odonatan Reisia gelasii (= Triadotypus guillaumei ) из триаса имел крыло длиной 136 миллиметров (5,4 дюйма), а размах крыльев мог составлять 280 миллиметров (11 дюймов). [682] [683]
Подёнки (Ephemeroptera)
[ редактировать ]- Самая крупная известная подёнка — пермская Ponalex maximus , с задним крылом длиной 55 мм (2 дюйма). [684] Меловой Epicharmeropsis Quadrivulosus имел переднее крыло длиной 37 мм (1 дюйм). [685]
- Хотя Bojophlebia prokopi из верхнего карбона Моравии , ( Чехия ) с размахом крыльев 45 см (18 дюймов) описывается как самая крупная подёнка [686] более позднее исследование показывает, что это насекомое не имеет отношения к подёнкам. [687]
Палеодиктиоптера
[ редактировать ]Самым крупным известным палеодиктиоптераном был Mazothairos с предполагаемым размахом крыльев до 560 мм (22 дюйма). [688] Если субкруглое крыло, известное из карьера Писберг, принадлежит палеодиктиоптерану, возможно, оно имело длину одного крыла не менее 30 см (12 дюймов). [689]
Archaeognatha (прыгающие щетинки) и другие бескрылые примитивные насекомые.
[ редактировать ]- Самый крупный известный махилид — Triassic Gigamachilis , его длина тела составляет 40 миллиметров (1,6 дюйма), не считая длины нити , а общая длина оценивается примерно в 80 миллиметров (3,1 дюйма). [690]
- Самые крупные экземпляры вымершего подотряда Monura достигали 30 миллиметров (1,2 дюйма) и более, не считая длины нити. [691]
- Хотя Рамсделепидион когда-то считался чешуйницей длиной 60 миллиметров (2,4 дюйма) , [692] Позже считалось, что классификация неопределенна и рассматривается как насекомое стеблевой группы . [693]
- Бескрылое раннее насекомое Carbotriplura имело длину тела около 103 миллиметров (4,1 дюйма) без хвостовых нитей. [694]
Четогната
[ редактировать ]Кембрийский , Timorebestia koprii жил 518 миллионов лет назад и был родственником современных стрелообразных червей обитающих в современных океанах. Вырастая до 30 см в длину, включая усики, они были намного крупнее и массивнее современных форм. До эволюции нектонных панартропод, челюстных позвоночных и головоногих эта группа крупных хищных гнатифер доминировала на вершине пищевой цепи. [695] [696]
Кольчатые черви (Annelida)
[ редактировать ]Вебстероприон — самый крупный из известных ископаемых евницидовых кольчатых червей , предполагаемая длина которого составляет 0,42–8,3 м (1 фут 5 дюймов — 27 футов 3 дюйма), однако сравнение с близкородственными современными таксонами показывает, что длина составляет около 1–2 м (3 фута 3 дюйма — 6 дюймов). футов 7 дюймов). [697] У него также были самые большие сколекодонты среди всех доисторических полихет , до 13,2 мм (0,52 дюйма) в длину и, возможно, больше. [697]
Моллюски (Mollusca)
[ редактировать ]Улитки и слизни (Gastropoda)
[ редактировать ]
- Самые крупные из известных брюхоногих моллюсков принадлежали к роду Campanile , при этом вымерший Campanile giganteum имел длину раковины до 90 см (35 дюймов). [698] или даже более 120 см (47 дюймов). [699]
- Самый крупный из известных каури — Vicetia bizzottoi , длина раковины которого составляет 33,5 см (13,2 дюйма). [700]
- Pebasiconcha immanis - самая крупная из когда-либо известных наземных улиток, высота раковины составляет 25,6 см (10,1 дюйма), а высота части экземпляра может превышать 30 см (12 дюймов). [701]
Двустворчатые моллюски (Двустворчатые моллюски)
[ редактировать ]- Самым крупным из когда-либо известных двустворчатых моллюсков , а также самой крупной иноцерамидой был Platyceramus platinus , гигант, осевая длина которого обычно составляла 1 м (3 фута 3 дюйма), но некоторые особи могли достигать осевой длины до 3 м (9,8 фута). . [702] Еще одним крупным доисторическим двустворчатым моллюском был Inoceramus . В 1952 году экземпляр Inoceramus steenstrupi длиной 187 см (6,14 фута) был найден в позднемеловых отложениях Гренландии . [703]
- Некоторые пермские алатоконхид рода , такие как Shikamaia, имели длину раковины около 1 м (3 фута 3 дюйма). [704] Предыдущая оценка реконструировала длину Шикамайи около 1,6 м (5 футов 3 дюйма). [705]
- Самая длинная остреида — Конбострея , высота панциря которой достигает 1,2 м (3 фута 11 дюймов). [706]
- Рудистские титаносарколиты имели общий размер около 2 м (6 футов 7 дюймов). [707]
Раковины бивней (Scaphopoda)
[ редактировать ]- Полная длина панциря бивня Prodentalium onoi оценивается более 30 см (12 дюймов). [708]
Головоногие моллюски (Cepphalopoda)
[ редактировать ]
Наутилоиды (Nautiloidea)
[ редактировать ]Самым крупным и длинным из известных наутилоидей был Endoceras giganteum с длиной раковины 5,73 м (18,8 футов). Есть запись об особи, длина панциря которой достигала 9,14 м (30,0 футов), но она сомнительна. [709]
Аммониты (Ammonoidea)
[ редактировать ]Самым крупным известным аммонитом была Parapuzosia seppenradensis . [710] Частичный ископаемый экземпляр, найденный в Германии, имел диаметр раковины 1,95 м (6 футов 5 дюймов), но жилая камера была неполной, поэтому предполагаемый диаметр раковины, вероятно, составлял около 3,5 м (11 футов), а вес - около 705 кг (1554 фунта). ), когда он был жив. [711] Однако, по оценкам более позднего исследования, диаметр раковины составляет примерно 2 м (6 футов 7 дюймов). [712]
Белемниты (Belemnoidea)
[ редактировать ]Самым крупным известным белемнитом был Megateuthis gigantea , максимальный диаметр и длина рострума которого достигали примерно 50 и 700 мм (2,0 и 27,6 дюйма) соответственно. [713]
Кальмары, осьминоги, каракатицы и их союзники (Neocoleoidea)
[ редактировать ]- Осьминог Enchoteuthis melanae (считающийся экземпляром Tusoteuthis longa ) имел длину мантии до 2 метров (6 футов 7 дюймов), что сравнимо с современным гигантским кальмаром . Ранее этот таксон считался звероподобным гигантским кальмаром, общая длина которого, включая руки, превышает 10 метров (33 фута). Однако, учитывая других ископаемых родственников, общая длина, включая руки, оценивается примерно в 3 метра (9,8 футов). [714]
- По оценкам, как неосьминогий Yezoteuthis , так и теутид Haboroteuthis по размеру аналогичны современному гигантскому кальмару . [715] [716]
Брахиоподы (Brachiopoda)
[ редактировать ]
Самым крупным брахиоподом , когда-либо существовавшим, была Striatifera striata из Аккермановского карьера , Россия , высотой до 0,5 метра (1 фут 8 дюймов). [717] Еще одним огромным брахиоподом был каменноугольный Gigantoproductus giganteus с шириной раковины от 30 см (12 дюймов). [718] до более 35 сантиметров (14 дюймов). [707] [719] Titanaria Costellata имела большую и длинную раковину шириной 35–36 см (14–14 дюймов), почти такую же большую, как Gigantoproductus . [720]
Гиолиты (Hyolitha)
[ редактировать ]Самый крупный гиолит — Macrotheca almgreeni , его длина около 50 сантиметров (20 дюймов). [707] [721]
Книдарийцы (Cnidaria)
[ редактировать ]Медузы и союзники (Medusozoa)
[ редактировать ]Самая крупная ископаемая медуза — кембрийская Cordubia gigantea , диаметром 88 сантиметров (35 дюймов). [722] Образцы из кембрия Висконсина достигали 70 см (28 дюймов) в длину. [723]
Вендобионты (Vendobionta)
[ редактировать ]Петалонамиды (Petalonamae)
[ редактировать ]
Самые длинные экземпляры Trepassia wardae (также известные как Charnia wardi ) достигали 185 см (73 дюйма) в длину. [724] Charnia masoni известна от экземпляров размером от 1 см (0,39 дюйма) до самых крупных экземпляров длиной 66 см (26 дюймов). [725]
Проговаривать
[ редактировать ]Dickinsonia tenuis достигала 1,4 м (4,6 фута) в длину, что делает его одним из крупнейших докембрийских организмов. [726] [727]
Губки (Porifera)
[ редактировать ]Самая крупная известная пермская губка Gigantospongia имела диаметр до 2,5 метров (8 футов 2 дюйма). [728]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Карбоне, Крис; Учитель, Эмбер; Роуклифф, Дж (2007). «Издержки плотоядства» . ПЛОС Биология . 5 (2): е22. doi : 10.1371/journal.pbio.0050022 . ПМК 1769424 . ПМИД 17227145 .
- ^ Хокканен, JEI (21 февраля 1986 г.). «Размер самого большого наземного животного». Журнал теоретической биологии . 118 (4): 491–499. Бибкод : 1986JThBi.118..491H . CiteSeerX 10.1.1.63.3969 . дои : 10.1016/S0022-5193(86)80167-9 . ПМИД 3713220 .
- ^ Романо, Марко; Циттон, Пол; Магануко, Симоне; Сакки, Ева; Карателли, Мартина; Рончи, Аусонио; Никосия, Умберто (2019). Сомервилл, Айдахо (ред.). «Новое открытие базальных синапсид на пермском обнажении Торре-дель-Портиччоло (Альгеро, Италия)» . Геологический журнал . 54 (3): 1554–1566. Бибкод : 2019GeolJ..54.1554R . дои : 10.1002/gj.3250 . ISSN 0072-1050 . S2CID 133755506 .
- ^ «Стратиграфия Перми - Международная комиссия по стратиграфии, Международный союз геологических наук» (PDF) . permian.stratigraphy.org . Архивировано (PDF) из оригинала 3 декабря 2018 года . Проверено 8 сентября 2022 г.
- ^ Рейс, Роберт Р.; Фрёбиш, Йорг (16 апреля 2014 г.). «Старейший казеидный синапсид из позднего пенсильванского периода Канзаса и эволюция травоядных животных у наземных позвоночных» . ПЛОС ОДИН . 9 (4): е94518. Бибкод : 2014PLoSO...994518R . дои : 10.1371/journal.pone.0094518 . ПМЦ 3989228 . ПМИД 24739998 .
- ^ Ромер, А.С.; Прайс, Л.В. (1940). «Обзор пеликозавров» . Специальные статьи Геологического общества Америки (28): 400, 403. ISBN. 978-0-8137-2028-9 .
- ^ «Эдафозавр» . Палеос . Архивировано из оригинала 20 февраля 2022 года . Проверено 9 сентября 2022 г.
- ^ Берман, Д.С.; Рейс, РР; Мартенс, Т.; Хенричи, AC (2001). «Новый вид Dimetrodon (Synapsida: Sphenacodontidae) из нижней перми Германии регистрирует первое появление этого рода за пределами Северной Америки» (PDF) . Канадский журнал наук о Земле . 38 (5): 803–812. Бибкод : 2001CaJES..38..803B . doi : 10.1139/cjes-38-5-803 .
- ^ Бринк, Кирстин С.; Рейс, Роберт Р. (16 октября 2014 г.). «Скрытое разнообразие зубов у старейшего наземного хищника Диметродона» . Природные коммуникации . 5 : 3269. Бибкод : 2014NatCo...5.3269B . дои : 10.1038/ncomms4269 . ПМИД 24509889 .
- ^ Олсон, ЕС (1955). «Параллелизм в эволюции пермских рептильных фаун Старого и Нового Света» . Филдиана . 37 (13): 395 . Проверено 8 сентября 2022 г.
- ^ Сен-Флер, Николас (4 января 2019 г.). «Родственник млекопитающих размером со слона, который пасся рядом с динозаврами» . Нью-Йорк Таймс . Проверено 9 сентября 2022 г.
- ^ Сулей, Томаш; Недзведски, Гжегож (4 января 2019 г.). «Синапсид позднего триаса размером с слона с прямыми конечностями» . Наука . 363 (6422): 78–80. Бибкод : 2019Sci...363...78S . дои : 10.1126/science.aal4853 . ПМИД 30467179 .
- ^ «Обнаружен «двоюродный брат» гигантского млекопитающего» . Наука Дейли . 23 ноября 2018 г. Архивировано из оригинала 22 августа 2022 г. . Проверено 10 сентября 2022 г.
- ^ ван Валкенбург, Блэр; Дженкинс, Ян (2002). «Эволюционные закономерности в истории пермо-триасовых и кайнозойских синапсидных хищников». Документы Палеонтологического общества 8: 267–288.
- ^ Перейти обратно: а б «Бритоподиды/Антеозавриды» . Хепер . М.Алан Казлев. Архивировано из оригинала 12 августа 2019 года . Проверено 9 сентября 2022 г.
- ^ Перейти обратно: а б «Эотитанозухиды» . Хепер . М.Алан Казлев. Архивировано из оригинала 6 августа 2019 года . Проверено 9 сентября 2022 г.
- ^ Протеро, Дональд Р. (18 апреля 2022 г.). «20. Синапсиды: происхождение млекопитающих». Эволюция позвоночных: от истоков до динозавров и далее . Бока-Ратон: CRC Press. дои : 10.1201/9781003128205-4 . ISBN 978-0-367-47316-7 . S2CID 246318785 .
- ^ Перейти обратно: а б с Каммерер, Кристиан Ф. (2016). «Систематика Rubidgeinae (Therapsida: Gorgonopsia)» . ПерДж . 4 : е1608. дои : 10.7717/peerj.1608 . ПМЦ 4730894 . ПМИД 26823998 .
- ^ Фрэнк Захос, Роберт Ашер (22 октября 2018 г.). Эволюция, разнообразие и систематика млекопитающих . Де Грютер. стр. 158–159. ISBN 978-3-11-034155-3 . Проверено 9 сентября 2022 г.
- ^ Дж. Ван Ден Хивер (1987), диссертация на степень доктора философии в Стелленбосском университете.
- ^ ЛИДЕККЕР, Р. (1908). «Годовая палеонтология позвоночных». Научный прогресс в двадцатом веке (1906–1916) . 2 (7): 501–524. JSTOR 43776634 .
- ^ Брум, Роберт (1903). «О некоторых новых примитивных териодонтах». Анналы Южноафриканского музея . 4 .
- ^ Толчард, Фредерик; Чемберлен, Кристиан Ф.; Батлер, Ричард Дж.; Хендрикс, Кристофер; Бенуа, Жюльен; Абдулла, Фердинанд; Шуаньер, Иона Н. (26 июля 2021 г.). «Новый крупный гомфодонт из триаса Южной Африки и его значение для биостратиграфии Гондваны» (PDF ) Журнал палеонтологии позвоночных . 41 (2): e1929265. Бибкод : 2021JVPal..41E9265T . дои : 10.1080/02724634.2021.1929265 . ISSN 0272-4634 . S2CID 237517965 .
- ^ Уильям А. Клеменс (2011). «Новые морганукодонтаны из заполнения раннеюрской трещины в Уэльсе (Великобритания)» . Палеонтология . 54 (5): 1139–1156. Бибкод : 2011Palgy..54.1139C . дои : 10.1111/j.1475-4983.2011.01094.x .
- ^ Роуз 2006 , с. 56
- ^ Пол Селден; Джон Наддс (19 сентября 2012 г.). Эволюция ископаемых экосистем. Второе издание . Академическая пресса. п. 178. ИСБН 978-0-12-404629-0 . Проверено 25 августа 2022 г.
- ^ Зофия Килан-Яворовска ; Ричард Л. Сифелли; Чжэ-Си Ло (2004). «Глава 12: Метатерийцы». Млекопитающие эпохи динозавров: происхождение, эволюция и строение . Нью-Йорк: Издательство Колумбийского университета. п. 243. ИСБН 978-0-231-11918-4 .
- ^ Перейти обратно: а б с Протеро, Дональд Р. (15 ноября 2016 г.). Принстонский полевой справочник по доисторическим млекопитающим . Издательство Принстонского университета. п. 33. ISBN 978-0-691-15682-8 . Проверено 22 сентября 2022 г.
- ^ Фрэнк Захос, Роберт Ашер (22 октября 2018 г.). Эволюция, разнообразие и систематика млекопитающих . Де Грютер. п. 224. ИСБН 978-3-11-034155-3 . Проверено 25 августа 2022 г.
- ^ Перейти обратно: а б Роуз 2006 , с. 62.
- ^ Перейти обратно: а б Килан-Яворовска, Зофия (2013). В поисках ранних млекопитающих . Издательство Университета Индианы. п. 115. ИСБН 978-0-253-00824-4 . Проверено 25 августа 2022 г.
- ^ Ху; и др. (2005). «Крупные млекопитающие мезозоя питались молодыми динозаврами» (PDF) . Природа . 433 (7022): 149–152. Бибкод : 2005Natur.433..149H . дои : 10.1038/nature03102 . ПМИД 15650737 . S2CID 2306428 .
- ^ Т. С. Кемп (2005). Происхождение и эволюция млекопитающих . Издательство Оксфордского университета, США. п. 183. ИСБН 978-0-19-850761-1 . Проверено 22 сентября 2022 г.
- ^ Роуз 2006 , с. 60
- ^ Томас Э. Уильямсон, Стивен Л. Брусатте, Росс Секорд, Сара Шелли, Новый многобугорчатый тэниолабидоид (Mammalia) из среднего Пуэркана формации Насимьенто, Нью-Мексико, и пересмотр систематики и филогении тэниолабидоидов, 5 октября 2015 г., doi : 10.1111/zoj.12336: «Taeniolabidoids претерпели умеренную таксономическую радиацию в раннем палеоцене Северной Америки и претерпели резкое увеличение размеров тела, при этом Taeniolabis taoensis, возможно, превысил 100 кг»
- ^ « Zaglossus hacketti – вымершая гигантская ехидна» . Туризм Западная Австралия . Архивировано из оригинала 26 января 2014 года . Проверено 28 апреля 2014 г.
- ^ Пиан, Ребекка; Арчер, Майкл; Хэнд, Сюзанна Дж. (1 ноября 2013 г.). «Новый гигантский утконос Obdurodon tharalkooschild, sp. nov. (Monotremata, Ornithorhynchidae), из зоны всемирного наследия Риверслей, Австралия». Журнал палеонтологии позвоночных . 33 (6): 1255–1259. Бибкод : 2013JVPal..33.1255P . дои : 10.1080/02724634.2013.782876 . ISSN 0272-4634 . S2CID 85776473 .
- ^ Вейль, Энн (2005). «Палеобиология млекопитающих: большая жизнь в меловом периоде». Природа . 433 (7022) (опубликовано 12 января 2005 г.): 116–117. Бибкод : 2005Natur.433..116W . дои : 10.1038/433116b . ПМИД 15650725 . S2CID 52869101 .
- ^ Превости, Франциско Дж.; Форазиепи, Аналия; Зимич, Наталья (5 ноября 2011 г.). «Эволюция кайнозойской гильдии хищников наземных млекопитающих в Южной Америке: конкуренция или замена?». Журнал эволюции млекопитающих . 20 (1): 3–21. дои : 10.1007/s10914-011-9175-9 . hdl : 11336/2663 . S2CID 15751319 .
- ^ Эрколи, Маркос Дарио; Превости, Франсиско Хуан (1 декабря 2011 г.). «Массовая оценка видов Святого Креста (ранний миоцен) Sparassodonta (Mammalia, Metatheria) по центроидному размеру аппендикулярных элементов: палеоэкологические выводы» [Массовая оценка видов Святого Креста (ранний миоцен) Sparassodonta (Mammalia, Metatheria) из Центроидный размер аппендикулярных элементов: палеоэкологические выводы. Амегиниана (на испанском языке). 48 (4): 462–479. дои : 10.5710/amgh.v48i4(347) . S2CID 129838311 .
- ^ Перейти обратно: а б с д и ж г час Соркин, Борис (декабрь 2008 г.). «Биомеханическое ограничение массы тела у наземных хищников-млекопитающих». Летайя . 41 (4): 333–347. Бибкод : 2008Лета..41..333С . дои : 10.1111/j.1502-3931.2007.00091.x .
- ^ Энгельман, Рассел К.; Флинн, Джон Дж. (Джон Джозеф); Висс, Эндрю Р.; Крофт, Дарин А. (17 июля 2020 г.). Eomakhaira molossus, новый саблезубый спарассодонт (Metatheria: Thylacosmilinae) из раннеолигоценового (?Tinguirirican) местонахождения Качапоал, Главный хребет Анд, Чили. (Новитаты Американского музея, № 3957) (Отчет). Американский музей естественной истории. hdl : 2246/7235 .
- ^ А. М. Форазиепи, М. Джудит Бабо и Н. Зимич. 2014. Australohyaena antiqua (Mammalia, Metatheria, Sparassodonta), крупный хищник из позднего олигоцена Патагонии. Журнал систематической палеонтологии 13(6):503–525 DOI: 10.1080/14772019.2014.926403
- ^ Роуз 2006 , с. 78
- ^ Зофия Килан-Яворовска ; Ричард Л. Сифелли; Чжэ-Си Ло (2004). «Глава 12: Метатерийцы». Млекопитающие эпохи динозавров: происхождение, эволюция и строение . Нью-Йорк: Издательство Колумбийского университета. п. 428. ИСБН 978-0-231-11918-4 .
- ^ Уилсон, врач общей практики; Экдейл, Э.Г.; Хогансон, Дж.В.; Каледе, Джей-Джей; Линден, А.В. (2016). «Крупное хищное млекопитающее позднего мела и североамериканского происхождения сумчатых» . Природные коммуникации . 7 : 13734. Бибкод : 2016NatCo...713734W . дои : 10.1038/ncomms13734 . ПМК 5155139 . ПМИД 27929063 .
- ^ «Сумчатые ледникового периода превысили три тонны, говорят ученые» . Архивировано из оригинала 13 апреля 2010 года.
- ^ Ричардс, Хейзел Л.; Уэллс, Род Т.; Эванс, Алистер Р.; Фицджеральд, Эрих М.Г.; Адамс, Джастин В. (13 сентября 2019 г.). «Необычайная остеология и функциональная морфология конечностей Palorchestidae, семейства странных вымерших сумчатых гигантов» . ПЛОС ОДИН . 14 (9): e0221824. Бибкод : 2019PLoSO..1421824R . дои : 10.1371/journal.pone.0221824 . ISSN 1932-6203 . ПМК 6744111 . ПМИД 31518353 .
- ^ Аллуан-Сегье, Леани; Санчес-Вильягра, Марсело Р.; Ли, Майкл С.Ю.; Лебрен, Рено (2013). «Костный лабиринт у дипротодонтовых сумчатых млекопитающих: разнообразие существующих и вымерших форм и взаимосвязь с размером и филогенией». Журнал эволюции млекопитающих . 20 (3): 191–198. дои : 10.1007/s10914-013-9228-3 . S2CID 16385939 .
- ^ Роу, С.; Майерс, Ти Джей; Уэллс, RT; Гиллеспи, А. (1999). «Оценка веса плейстоценового сумчатого льва Thylacoleo carnifex (Thylacoleonidae:Marsupialia): значение для экоморфологии сумчатого суперхищника и гипотезы обнищания австралийской фауны сумчатых хищников». Австралийский журнал зоологии . 47 (5): 489. дои : 10.1071/ZO99006 . ISSN 0004-959X .
- ^ Хокнулл, Скотт А.; Льюис, Ричард; Арнольд, Ли Дж.; Питч, Тим; Жоан-Бойо, Рено; Прайс, Гилберт Дж.; Мосс, Патрик; Вуд, Рэйчел; Доссето, Энтони; Луис, Жюльен; Олли, Джон; Лоуренс, Рошель А. (18 мая 2020 г.). «Вымирание мегафауны восточного Сахула совпадает с устойчивым ухудшением состояния окружающей среды» . Природные коммуникации . 11 (1): 2250. Бибкод : 2020NatCo..11.2250H . дои : 10.1038/s41467-020-15785-w . ISSN 2041-1723 . ПМК 7231803 . ПМИД 32418985 .
- ^ Хелген, КМ; Уэллс, RT; Кир, BP; Гердц, WR; Фланнери, ТФ (2006). «Экологическое и эволюционное значение размеров гигантских вымерших кенгуру». Австралийский журнал зоологии . 54 (4): 293–303. дои : 10.1071/ZO05077 .
- ^ Янис, СМ; Баттрилл, К; Фигейридо, Б (2014). «Передвижение вымерших гигантских кенгуру: были ли стенурины монстрами без хмеля?» . ПЛОС ОДИН . 9 (10): e109888. Бибкод : 2014PLoSO...9j9888J . дои : 10.1371/journal.pone.0109888 . ПМК 4198187 . ПМИД 25333823 .
- ^ Лонг, Джон А. (2002). Доисторические млекопитающие Австралии и Новой Гвинеи: сто миллионов лет эволюции . Издательство Университета Джонса Хопкинса. п. 174. ИСБН 978-0-8018-7223-5 .
- ^ Казимеж Ковальский (1976). Млекопитающие. Очерк териологии . Польские научные издательства. п. 442 . Проверено 17 сентября 2022 г.
- ^ Джон В. Хогансон (2007). Динозавры, акулы и шерстистые мамонты . Государственное историческое общество Северной Дакоты. п. 36. ISBN 978-1-891419-33-1 . Проверено 17 сентября 2022 г.
- ^ Уэн, Марк Д.; Джинджерич, Филип Д. (1995). «Эволюция корифодона (Mammalia, Pantodonta) в позднем палеоцене и раннем эоцене северо-западного Вайоминга» (PDF) . Материалы Музея палеонтологии Мичиганского университета . 29 (10): 264. OCLC 742731820 . Проверено 17 сентября 2022 г.
- ^ «Палеоценовые млекопитающие мира» . Архивировано из оригинала 11 сентября 2022 года.
- ^ Перейти обратно: а б «Палеоценовые млекопитающие мира» . www.paleocene-mammals.de . Архивировано из оригинала 3 августа 2022 года . Проверено 17 сентября 2022 г.
- ^ Перейти обратно: а б Ти Джей Михан, Ларри Д. Мартин (1 сентября 2012 г.). «Новое крупное насекомоядное лепптид из позднего палеогена Южной Дакоты, США» . Acta Palaeontologica Polonica . 57 (3): 509–518. дои : 10.4202/app.2011.0035 . S2CID 129358395 .
- ^ Макфи, Росс (январь 1994 г.). «Морфология, адаптации и взаимоотношения Plesiorycteropus, а также диагноз нового отряда эвтерийных млекопитающих» . Бюллетень Американского музея естественной истории (220): 148 – через ResearchGate.
- ^ «Гиппопотам — paleofiles.com» . Архивировано из оригинала 31 октября 2020 года.
- ^ Дональд Р. Протеро , Скотт Э. Фосс (2007). Эволюция парнокопытных . Издательство Университета Джонса Хопкинса. стр. 128–129. ISBN 978-0-8018-8735-2 . Проверено 26 сентября 2020 г.
- ^ Петерсон, О.А. (1909). «Пересмотр энтелодонтид» . Мемуары музея Карнеги . 4 (3): 41–158. дои : 10.5962/стр.234831 . hdl : 2027/mdp.39015017493571 . S2CID 247000277 .
- ^ Осборн, HF (11 ноября 1924 г.). «Андрусарх, гигантский мезонихид Монголии» (PDF) . Американский музей послушников (146). Американский музей естественной истории . Архивировано из оригинала 2 ноября 2020 года.
- ^ Тьювиссен, JGM ; Купер, Лиза Ноэль; Клементц, Марк Т.; Баджпай, Сунил; Тивари, Б.Н. (2009). «Тевиссен и др. Ответ» . Природа . 458 (7236): Е5. Бибкод : 2009Natur.458....5T . дои : 10.1038/nature07775 . S2CID 4431497 .
- ^ Р. Табусе, Дж. Клавель и М.Т. Антунес. 2011. Структурное промежуточное звено между триизодонтидами и мезонихиями (Mammalia, Acreodi) из раннего эоцена Португалии. Naturwissenschaften 98: 145–155.
- ^ Ист, Ширли Г. (29 декабря 2011 г.). Эпоха охотников за сновидениями: серия «Индейцы Палео» . Корпорация Xlibris. ISBN 978-1-4653-9694-5 .
- ^ Мука, Ричард А.; Вискайно, Серджио Ф.; Юлиис, Джерри Де (22 мая 2013 г.). Мегафауна: гигантские звери плейстоцена Южной Америки . Издательство Университета Индианы. ISBN 978-0-253-00719-3 .
- ^ «Бизон латифронс - характеристики, поведение и среда обитания бизона латифронса, гигантского зубра» . Архивировано из оригинала 10 ноября 2013 года.
- ^ Перейти обратно: а б «Вымершие длиннорогие бизоны и древние бизоны (Bison latifrons и B. antiquus). Информационный бюллетень: Краткое изложение. Глобальная библиотека зоопарка Сан-Диего» . Архивировано из оригинала 15 января 2021 года.
- ^ Куртен, Б; Андерсон, Э. (1980). «Отряд Парнокопытные » Плейстоценовые млекопитающие Северной Америки (1-е изд.). Нью-Йорк: Издательство Колумбийского университета. стр. 100-1 295–339. ISBN 0-231-03733-3 .
- ^ «Родео выходного дня Warkworth Western | Участники» . Архивировано из оригинала 21 августа 2011 года . Проверено 30 мая 2012 г.
- ^ Палмер, Дуглас, изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла . Лондон: Marshall Editions. п. 281. ИСБН 978-1-84028-152-1 .
- ^ Перейти обратно: а б Кэмпс, Габриэль (1992). «Античная месть» . В лагерях, Габриэль (ред.). Берберская энциклопедия (на французском языке). Экс-ан-Прованс: Эдисуд. стр. 1642–1647. doi : 10.4000/encyclepedieberbere.1875 . Архивировано из оригинала 18 июля 2020 года . Проверено 31 августа 2022 г.
- ^ Помель, Огюст (1893). Бубалус антиквус . Carte de Géologie de l’Algérie – Paléontologie Monographies de Vertébrés (на французском языке). Алжир: П. Фонтана импримери. стр. 1–94, табл. 1–10. дои : 10.5962/bhl.title.13867 .
- ^ Кисели, Рене. «Зуб и потенциальное скрещивание с домашним скотом в Центральной Европе в период энеолита. Метрический анализ костей из археологического памятника Кутна Гора-Дания (Чехия)». Антропозоология . 43 (2): 2008.
- ^ «Купрей (Бос саувели)» . Архивировано из оригинала 10 августа 2011 года.
- ^ Берни Д. и Уилсон Д.Э. (ред.), Животные: Полное визуальное руководство по дикой природе мира . ДК Взрослый (2005), ISBN 0-7894-7764-5
- ^ К. Сурпрасит; Ж.-Ж. Расположение; Ю. Чайман; О. Нож; К. Ями; П. Тянь; С. Панья (2016). «Фауна позвоночных среднего плейстоцена из Кхок Сунга (Накхонратчасима, Таиланд): биохронологические и палеобиогеографические значения» . ZooKeys (613): 1–157. Бибкод : 2016ZooK..613....1S . дои : 10.3897/zookeys.613.8309 . ПМК 5027644 . ПМИД 27667928 .
- ^ Дуглас М. Консидайн, Гленн Д. Консидайн (2013). Научная энциклопедия Ван Ностранда . Спрингер США. п. 446. ИСБН 978-1-4757-6918-0 . Проверено 28 августа 2022 г.
- ^ Херринг, Энди Д. (2014). Системы производства мясного скота . КАБИ. п. 22. ISBN 978-1-78064-507-0 . Проверено 28 августа 2022 г.
- ^ Терри Харрисон (27 января 2011 г.). Палеонтология и геология Лаэтоли: эволюция человека в контексте. Том 2: Ископаемые гоминины и связанная с ними фауна . Спрингер Нидерланды. п. 404. ИСБН 978-90-481-9962-4 . Проверено 17 сентября 2022 г.
- ^ Открытие. Журнал Музея естественной истории Пибоди Йельского университета. Тома 13–16 . Музей естественной истории Пибоди. 1978. с. 9 . Проверено 17 сентября 2022 г.
- ^ Отчет Южноафриканской ассоциации развития науки. Том 96 . Южноафриканская ассоциация развития науки. 2000. с. 163 . Проверено 17 сентября 2022 г.
- ^ Дэвид Петерсен (1989). «О лосях, мегалоцеросе и чудесах» . Motherearthnews.com . Архивировано из оригинала 4 мая 2017 года.
- ^ Моен, Рон А.; Пастор Джон; Коэн, Йосеф (1999). «Рост рогов и исчезновение ирландских лосей» (PDF) . Исследования в области эволюционной экологии : 235–249. Архивировано из оригинала (PDF) 12 августа 2022 года.
- ^ Харалампос Кеврекидис; Димитрис С. Костопулос (март 2017 г.). «Самое южное возникновение Cervalces latifrons (Johnson, 1874) (Artiodactyla: Cervidae) в Европе» . Исследовательские ворота . дои : 10.13140/RG.2.2.24751.53928 .
- ^ Гейст, Валериус (1998). Олени мира: их эволюция, поведение и экология . Книги Стэкпола. ISBN 978-0-8117-0496-0 . Книги Оксворта. стр. 111, 126, 247–250. (1998) ISBN 0-8117-0496-3
- ^ Бреда, Марсия (2010). « Сервальцес латифронс » . Музей естественной истории. Архивировано из оригинала 4 января 2014 года.
- ^ Штраус, Боб. «Лось-олень – факты и цифры» . Мыслько.com. Архивировано из оригинала 4 ноября 2021 года . Проверено 11 октября 2022 г.
- ^ «(на испанском)» . Laignoranciadelconocimiento.blogspot.com.es. 23 декабря 2011 г. Архивировано из оригинала 27 января 2022 г. . Проверено 11 октября 2022 г.
- ^ Басу, Кристофер; Фолкингем, Питер Л.; Хатчинсон, Джон Р. (январь 2016 г.). «Вымерший гигантский жираф Sivatherium giganteum : реконструкция скелета и оценка массы тела» . Письма по биологии . 12 (1): 20150940. doi : 10.1098/rsbl.2015.0940 . ПМЦ 4785933 . ПМИД 26763212 .
- ^ Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла . Лондон: Marshall Editions. п. 273. ИСБН 1-84028-152-9 .
- ^ Янис, К.М., Теодор, Дж.М., и Буасверт, Б. (2002). Еще раз о локомоторной эволюции верблюдов: количественный анализ анатомии педалей и освоение шагающей походки. Журнал палеонтологии позвоночных, 22 (1), 110–121.
- ^ Зубы: Kubanochoerus gigas lii (ГУАНЬ) . tesorosnaturales.es
- ^ Пикфорд, М. (2006). «Новые экземпляры суоидов из Гебель-Зельтена, Ливия» . Геологические исследования . 62 (1). дои : 10.3989/egeol.0662147 .
- ^ Палмер 1999 , стр. 269.
- ↑ Исчезнувшие виды гигантских верблюдов. Архивировано 10 ноября 2013 года в Wayback Machine . Intechinc.com (5 июля 2011 г.)
- ^ Мендоса, М.; Янис, СМ; Палмквист, П. (2006). «Оценка массы тела вымерших копытных: исследование использования множественной регрессии». Журнал зоологии . 270 (1): 90–101. CiteSeerX 10.1.1.541.9021 . дои : 10.1111/j.1469-7998.2006.00094.x .
- ^ Перейти обратно: а б «В Сирии найдена окаменелость гигантского верблюда» . Новости Би-би-си . 10 октября 2006 г. Архивировано из оригинала 27 сентября 2019 г. . Проверено 17 апреля 2013 г.
- ^ Ребекка Рэгг Сайкс (17 июня 2022 г.). Неандерталец, родственник: открытие нашего происхождения . Делашо и Нистле. ISBN 978-2-603-02968-8 . Проверено 28 августа 2022 г.
- ^ Энтони Дж. Стюарт, 2021, Исчезнувшие гиганты: Затерянный мир ледникового периода , «6.17 Вчерашний верблюд: Camelops Hesternus», стр.99, University of Chicago Press
- ^ Бьянуччи, Г.; Ламберт, О.; Урбина, М.; Мерелла, М.; Колларета, А.; Беннион, Р.; Салас-Жисмонди, Р.; Бенитес-Паломино, А.; Пост, К.; де Мюзон, К.; Босио, Г.; Ди Сельма, К.; Малинверно, Э.; Пьерантони, ПП; Вилла, ИМ; Амсон, Э. (2023). «Ранний кит-тяжеловес раздвигает границы морфологии позвоночных». Природа . 620 (7975): 824–829. Бибкод : 2023Natur.620..824B . дои : 10.1038/s41586-023-06381-1 . ПМИД 37532931 . S2CID 260433513 .
- ^ Джинджерич, PD; Ариф, М; Бхатти, М. Акрам; Анвар, М; Сандерс, Уильям Дж (1997). « Basilosaurus drazinda и Basiloterus hussaini , новые Archaeoceti (Mammalia, Cetacea) из среднеэоценовой формации Дразинда, с пересмотренной интерпретацией возраста китоносных слоев в группе Киртар Сулейманского хребта, Пенджаб (Пакистан)». Материалы Музея палеонтологии Мичиганского университета . 30 (2): 55–81. hdl : 2027.42/48652 . OCLC 742731913 .
- ^ Келлог Р. Обзор археоцитов. Публикации Института Карнеги в Вашингтоне. 1936 год; 482: 1–366.
- ^ Восс, Манья; Антар, Мохаммед Самех М.; Залмут, Ияд С.; Джинджерич, Филип Д. (2019). «Содержимое желудка археоцита Basilosaurus isis : высшего хищника океанов позднего эоцена» . ПЛОС ОДИН . 14 (1). е0209021. Бибкод : 2019PLoSO..1409021V . дои : 10.1371/journal.pone.0209021 . ПМК 6326415 . ПМИД 30625131 .
- ^ Джованни Бьянуччи; Джулия Босио; Элиза Малинверно; Кристиан Де Муизон; Игорь Вилла; Марио Урбина; Оливье Ламбер (2018). «Новый крупный сквалодельфинид (Cetacea, Odontoceti) из Перу проливает свет на неравенство и экологию платанистоидов раннего миоцена» . Королевское общество открытой науки . 5 (4): 172–302. Бибкод : 2018RSOS....572302B . дои : 10.1098/rsos.172302 . ПМЦ 5936943 . ПМИД 29765678 . S2CID 21680097 . Архивировано из оригинала 13 августа 2018 года.
- ^ Перейти обратно: а б Демере, штат Техас; Берта, А.; Макгоуэн, MR (2005). «Таксономическая и эволюционная история ископаемых и современных балаеноптероидных усатых китов». Журнал эволюции млекопитающих . 12 (1/2): 99–143. дои : 10.1007/s10914-005-6944-3 . S2CID 90231 .
- ^ Слейтер, Грэм Дж.; Голдбоген, Джереми А.; Пьенсон, Николас Д. (31 мая 2017 г.). «Независимая эволюция гигантизма усатых китов, связанная с динамикой океана плио-плейстоцена» . Труды Королевского общества B: Биологические науки . 284 (1855): 20170546. doi : 10.1098/rspb.2017.0546 . ISSN 0962-8452 . ПМЦ 5454272 . ПМИД 28539520 .
- ^ Перейти обратно: а б с д и ж г час Ларраменди, А. (2016). «Высота плеч, масса тела и форма хоботков» (PDF) . Acta Palaeontologica Polonica . 61 . дои : 10.4202/app.00136.2014 . S2CID 2092950 .
- ^ Фортелиус, М.; Каппельман, Дж. (1993). «Самое большое наземное млекопитающее, которое когда-либо можно было себе представить». Зоологический журнал Линнеевского общества . 108 : 85–101. дои : 10.1111/j.1096-3642.1993.tb02560.x .
- ^ Перейти обратно: а б Жегалло, В.; Каландадзе Н.; Шаповалов А.; Бессуднова З.; Носкова Н.; Тесакова, Е. (2005). «Об ископаемых носорогах Elasmotherium (в том числе из коллекций РАН)» (PDF) . Череп . 22 (1): 17–40.
- ^ Перейти обратно: а б Schvyreva, A.K. (2016). Эласмотерии плейстоцена Евразии (PDF) (in Russian). pp. 103–105.
- ^ Перейти обратно: а б Косинцев П.; Митчелл, К.Дж.; Девьезе, Т.; ван дер Плихт, Дж.; Куйтемс, М.; Петрова Е.; Тихонов А.; Хайэм, Т.; Комески, Д.; Терни, К.; Купер, А.; ван Кольфшотен, Т.; Стюарт, Эй Джей; Листер, AM (2019). «Эволюция и вымирание гигантского носорога Elasmotherium sibiricum проливает свет на позднечетвертичное вымирание мегафауны» . Экология и эволюция природы . 3 (1): 31–38. дои : 10.1038/s41559-018-0722-0 . hdl : 11370/78889dd1-9d08-40f1-99a4-0e93c72fccf3 . ПМИД 30478308 . S2CID 53726338 .
- ^ Краузе, Ганс (2011). «Ххпе 07 02» . hanskrause.de . Архивировано из оригинала 28 августа 2022 года . Проверено 10 сентября 2022 г.
- ^ Ханс-Дитер Сьюс, Росс Д.Е. Макфи (30 июня 1999 г.). Вымирания в ближайшем времени. Причины, контексты и последствия . Спрингер США. п. 262. ИСБН 978-0-306-46092-0 . Проверено 17 сентября 2022 г.
- ^ Боескоров, Г.Г. (2012). «Некоторые специфические морфологические и экологические особенности ископаемого шерстистого носорога ( Coelodonta antiquitatis Blumenbach 1799)». Биологический вестник . 39 (8): 692–707. Бибкод : 2012BioBu..39..692B . дои : 10.1134/S106235901208002X . S2CID 24868968 .
- ^ «Coelodonta antiquitatis (Млекопитающее)» . Triebold Paleontology, Inc. Архивировано из оригинала 2 июня 2012 года . Проверено 28 августа 2022 г.
- ^ «Аминодонты» . Архивировано из оригинала 8 сентября 2006 года.
- ^ Макларен, Джейми А; Халберт, Ричард С; Уоллес, Стивен С; Наувелартс, Сандра (5 октября 2018 г.). «Морфометрический анализ передних конечностей рода Tapirus (Perissodactyla: Tapiridae) выявляет влияние среды обитания, филогении и размера во времени и в географическом пространстве» . Зоологический журнал Линнеевского общества . 184 (2): 499–515. doi : 10.1093/zoolinnean/zly019 . ISSN 0024-4082 .
- ^ Перейти обратно: а б Stanislav Drobyshevsky (2021). Палеонтология антрополога. Том 3. Кайнозой (Paleontology of anthropologist. Volume 3. Cenozoic) . LitRes. ISBN 978-5-04-380567-6 . Проверено 19 сентября 2022 г.
- ^ Патрисия Викерс Рич ; Томас Хьюитт Рич; Милдред Адамс Фентон ; Кэрролл Лейн (15 января 2020 г.). Книга окаменелостей: летопись доисторической жизни . Дуврские публикации. п. 573. ИСБН 978-0-486-83855-7 . Проверено 25 августа 2022 г.
- ^ Национальный памятник окаменелостей агата, отчет NPS о природных ресурсах NPS/NRPC/GRD/NRR-2009/080 (Дж. Грэм, март 2009 г.)
- ^ Перейти обратно: а б Боб Штраусс. «Обзор бронтотерия (мегацеропса)» . О . Архивировано из оригинала 21 ноября 2021 года.
- ^ «Бронтотерий – Титанотер – эпоха олигоцена» . Архивировано из оригинала 26 мая 2013 года.
- ^ Перейти обратно: а б Грегори С. Пол; Гай Д. Лихи (1994). «Террамегатермия во времена титанов: восстановление метаболизма колоссальных динозавров» (PDF) . Специальные публикации Палеонтологического общества . 7: Дино Фест: 177–198. дои : 10.1017/S2475262200009515 . Архивировано из оригинала (PDF) 20 сентября 2022 года.
- ^ «Brontotheriidae. Американский музей естественной истории» . Архивировано из оригинала 20 ноября 2021 года.
- ^ Эйзенманн, Вера (2003). «Гигантские лошади» (PDF) . Достижения в палеонтологии позвоночных .
- ^ Перейти обратно: а б Брюс Дж. Макфадден (1992). Ископаемые лошади: систематика, палеобиология и эволюция семейства лошадиных . Издательство Кембриджского университета. п. 284. ИСБН 0-521-47708-5 . Проверено 21 сентября 2022 г.
- ^ Кэтлин М. Дженис (1998). Эволюция третичных млекопитающих Северной Америки: Том 1, Наземные, хищные, копытные и копытные млекопитающие . Издательство Кембриджского университета. п. 545. ИСБН 978-0-521-35519-3 . Проверено 21 сентября 2022 г.
- ^ Т. С. Кемп (4 ноября 2004 г.). Происхождение и эволюция млекопитающих . Издательство Оксфордского университета, США. п. 237. ИСБН 978-0-19-154517-7 . Проверено 21 сентября 2022 г.
- ^ Джерисон, Гарри Дж (декабрь 2007 г.). «Что окаменелости говорят нам об эволюции неокортекса» (PDF) . Эволюция нервных систем : 1–12. дои : 10.1016/B0-12-370878-8/00065-3 . ISBN 978-0-12-370878-6 .
- ^ Перейти обратно: а б с Патрисия Викерс Рич; Томас Хьюитт Рич; Милдред Адамс Фентон; Кэрролл Лейн Фентон (15 января 2020 г.). Книга окаменелостей: летопись доисторической жизни . Дуврские публикации. п. 555. ИСБН 978-0-486-83855-7 . Проверено 4 сентября 2022 г.
- ^ Перейти обратно: а б с д «Млекопитающие ледникового периода — EnchantedLearning.com» .
- ^ Сойбельзон, Л.Х.; Шуберт, BW (январь 2011 г.). «Самый крупный известный медведь, Arctotherium angustidens , из пампейского региона раннего плейстоцена Аргентины: с обсуждением тенденций размера и питания медведей» . Журнал палеонтологии . 85 (1): 69–75. дои : 10.1666/10-037.1 . hdl : 11336/104215 . S2CID 129585554 . Проверено 1 июня 2011 г.
- ^ Делл'Амор, К. (2011): Самый большой медведь, когда-либо найденный , National Geographic News, опубликовано 3 февраля 2011 г.
- ^ Фигейридо; и др. (2010). «Демифологизация Arctodus simus, длинноногого и хищного медведя с коротким лицом, которого никогда не было». Журнал палеонтологии позвоночных . 30 (1): 262–275. Бибкод : 2010JVPal..30..262F . дои : 10.1080/02724630903416027 . S2CID 85649497 .
- ^ СОЙБЕЛЬЗОН, ЛЕОПОЛЬДО Х.; ШУБЕРТ, БЛЕЙН В. (2011). «Самый крупный из известных медведей, Arctotherium Angustidens, из раннего плейстоцена Пампейского региона Аргентины: с обсуждением размеров и тенденций в питании медведей». Журнал палеонтологии . 85 (1): 69–75. дои : 10.1666/10-037.1 . hdl : 11336/104215 . JSTOR 23019499 . S2CID 129585554 .
- ^ Live Science Staff (25 ноября 2008 г.). «Огромные пещерные медведи: когда и почему они исчезли» . Живая наука .
- ^ К. Джин, Р. Л. Чочон, В. Донг, Р. М. Хант-младший, Дж. Лю, М. Джегер и К. Чжу. 2007. « Первый череп древнейшей гигантской панды ». Труды Национальной академии наук 104: 10932-10937.
- ^ Перейти обратно: а б Берта, Анналиса (2017). Возникновение морских млекопитающих: 50 миллионов лет эволюции . Издательство Университета Джонса Хопкинса. п. 110. ИСБН 978-1-4214-2325-8 . Проверено 21 августа 2022 г.
- ^ Перейти обратно: а б Ксения Кейли; Мортен Танге Олсен; Питер Джордан; Шон П.А. Дежарден (2021). Атлантический морж: междисциплинарный взгляд на взаимодействие человека и животных . Шарлотта Кокл. п. 17. ISBN 978-0-12-817431-9 . Проверено 21 августа 2022 г.
- ^ Морган Черчилль, Марк Т. Клементц, Наоки Коно. «Правило Коупа и эволюция размеров тела»в Pinnipedimorpha (Mammalia: Carnivora)». Эволюция. Январь 2015 г.; 69 (1): 201-15. doi: 10.1111 / evo.12560
- ^ Иштван Фози; Иштван Сенте; Гарет Дайк (1977). Отариоидные печати неогена. Профессиональный доклад геологической службы, том 992 . Геологическая служба (США). стр. 61–62 . Проверено 21 августа 2022 г.
- ^ Перейти обратно: а б Гроэ, Камилла; Один, Кевин; Буассери, Жан-Рено (2022). «Lutrinae Bonaparte, 1838 (Carnivora, Mustelidae) из плио-плейстоцена долины Нижнего Омо, юго-западная Эфиопия: систематика и новые взгляды на палеоэкологию и палеобиогеографию выдр Туркана» . Comptes Rendus Palevol (на французском языке). 30 (30): 684–693. дои : 10.5852/cr-palevol2022v21a30 . S2CID 252106648 .
- ^ Гераадс, Денис; Алемсегед, Зересенай; Бобе, Рене; Рид, Денне (2011). «Enhydriodon dikikae, sp. nov. (Carnivora: Mammalia), гигантская выдра из плиоцена Дикики, Нижний Аваш, Эфиопия». Журнал палеонтологии позвоночных . 31 (2): 447–453. Бибкод : 2011JVPal..31..447G . дои : 10.1080/02724634.2011.550356 . S2CID 84797296 .
- ^ «Медвежья выдра» . Проводной . 26 марта 2011 г.
- ^ «Siamogale melilutra: окаменелости гигантской выдры открывают новые виды - Палеонтология - Sci-News.com» . sci-news.com .
- ^ «Острова выдр и странных лисиц» . scienceblogs.com .
- ^ Перейти обратно: а б с Валенсиано, Альберто; Баскин, Джон А.; Абелла, Джон; Перес-Рамос, Александр; Альварес-Сьерра, М. Анхелес; Моралес, Джордж; Хартстон-Роуз, Адам (7 апреля 2016 г.). «Megalictis, гигантский куньих-костолом (Carnivora, Mustelidae, Oligobuninae) из раннего миоцена Северной Америки» . ПЛОС ОДИН 11 (4): e0152430. Бибкод : 2016PLoSO..1152430V . дои : 10.1371/journal.pone.0152430 . ISSN 1932-6203 . ПМЦ 4824437 . ПМИД 27054570 .
- ^ Протеро, Дональд Р. (2016). Принстонский полевой справочник по доисторическим млекопитающим . Издательство Принстонского университета. п. 144. ИСБН 978-1-4008-8445-2 . Проверено 28 августа 2022 г.
- ^ Тернер, Алан (2004). Развивающийся Эдем: иллюстрированное руководство по эволюции фауны крупных млекопитающих Африки . Морис Энтони. Нью-Йорк: Издательство Колумбийского университета. стр. 100-1 106–107. ISBN 0-231-11944-5 . OCLC 53900492 .
- ^ Орлов Ю. Ю. 1948. Perunium ursogulo Orlov, новый гигантский вымерший куньих (вклад в морфологию черепа и мозга, а также в филогению куньих). Acta Zoologica 29:63–105.
- ^ Валенсиано, Альберто; Абелла, Джон; Сансидро, Оскар; Хартстон-Роуз, Адам; Альварес-Сьерра, Мэри Энджелс; Моралес, Хорхе (27 мая 2015 г.). «Полное описание черепа и нижней челюсти гигантского куньего Eomellivora piveeaui Ozansoy, 1965 (Mammalia, Carnivora, Mustelidae) из Batallones (MN10), поздний миоцен (Мадрид, Испания) » Журнал палеонтологии позвоночных . 35 (4): e934570. Бибкод : 2015JVPal..35E4570V . дои : 10.1080/02724634.2014.934570 . S2CID 86216613 .
- ^ Форасиепи, Аналия М.; Превости, Франциско Дж. (2018). Эволюция южноамериканских млекопитающих-хищников в кайнозое: палеобиогеографические и палеоэкологические факторы . Чам: Спрингер. стр. 124–125. ISBN 978-3-319-03701-1 .
- ^ Тарквини, Юлиана; Толедо, Нестор; Сойбельзон, Леопольдо Х.; Морган, Сесилия К. (март 2017 г.). «Оценка массы тела † Cyonasua (Procyonidae, Carnivora) и родственных таксонов на основе посткраниального скелета» . Историческая биология . 30 (4): 496–506. дои : 10.1080/08912963.2017.1295042 . hdl : 11336/49670 . S2CID 90408657 .
- ^ Диас-Сибаха, Р. (2010). «Титаны. Том 1. Млекопитающие». Ископаемый журнал палеонтологии . ISSN 0717-9235
- ^ Андерссон, Ки (2005). «Существовали ли в третичном периоде псовые, охотящиеся стаями, и как мы можем это знать?». Палеобиология . 41 (4): 333–347. Бибкод : 2005Pbio...31...56A . doi : 10.1666/0094-8373(2005)031<0056:WTPCIT>2.0.CO;2 . S2CID 85306826 .
- ^ Ван, Сяомин ; Тедфорд, Ричард Х. (2008). Собаки: их ископаемые родственники и эволюционная история .
- ^ «Волки, койоты и собаки (род Canis )» . Museum.state.il.us . Проверено 23 октября 2011 г.
- ^ Будади-Малин, Мириам (2012). «Новый подвид волка (Canis lupus maximus nov. subsp.) из верхнего плейстоцена Западной Европы» . Об этом сообщает Палевол . 11 (7): 475. doi : 10.1016/j.crpv.2012.04.003 .
- ^ Берте, Э.; Пандольфи, Л. (2014). « Canis lupus (Mammalia, Canidae) из позднеплейстоценового отложения Аветрана (Таранто, Южная Италия)» . Итальянский журнал палеонтологии и стратиграфии . 120 (3): 367–379.
- ^ Фигейридо, Борха; Перес-Кларос, Хуан А.; Хант, Роберт М. младший; Пальмквист, Пол. «Оценка массы тела амфиционидных хищных млекопитающих: подход множественной регрессии по черепу и скелету - Acta Palaeontologica Polonica» (PDF) . www.app.pan.pl. дои : 10.4202/app.2010.0005 . S2CID 56051166 .
- ^ Хорди Агусти и Маурисио Антон: мамонты, саблезубые и гоминиды 65 миллионов лет эволюции млекопитающих в Европе , Columbia University Press, 2002, стр. 81–83
- ^ Барретт, Пол Закари (26 октября 2021 г.). «Самый большой гоплофонин и сложная новая гипотеза эволюции нимравидов» . Научные отчеты . 11 (1): 21078. Бибкод : 2021NatSR..1121078B . дои : 10.1038/s41598-021-00521-1 . ПМЦ 8548586 . ПМИД 34702935 . S2CID 240000358 .
- ^ Пенье, С.; де Бонис, Л.; Ликиус, А.; Маккей, Х.Т.; Виньо, П.; Брюне, М. (2005). «Новый махайродонт (Carnivora, Felidae) из позднемиоценового местонахождения гоминид TM 266, Торос-Меналла, Чад». Comptes Рендус Палевол . 4 (3): 243–253. Бибкод : 2005CRPal...4..243P . дои : 10.1016/j.crpv.2004.10.002 .
- ^ Перейти обратно: а б с Шерани, Шахир (2016). «Новый метод оценки массы тела кошачьих, зависящий от образца» (PDF) . Препринты PeerJ : 16. doi : 10.7287/peerj.preprints.2327v2 . Архивировано из оригинала (PDF) 20 сентября 2022 года . Проверено 25 августа 2022 г.
- ^ Перейти обратно: а б Джованни Дж. Беллани (2019). Кошачьи мира. Открытия в таксономической классификации и истории . Эльзевир Наука. стр. 30–31. ISBN 978-0-12-817277-3 . Проверено 19 сентября 2022 г.
- ^ «Недавно идентифицированная саблезубая кошка — одна из крупнейших в истории» .
- ^ Оркатт, Джон Д.; Каледе, Джонатан Дж. М. (2021). «Количественный анализ Feliform Humeri показывает существование очень крупной кошки в Северной Америке во время миоцена» . Журнал эволюции млекопитающих . 28 (3): 729–751. дои : 10.1007/s10914-021-09540-1 . S2CID 235541255 .
- ^ «Гигантский саблезубый кот бродил по Северной Америке во время миоцена | Палеонтология» . Sci-News.com. 4 мая 2021 г. Проверено 19 сентября 2022 г.
- ^ «Ксеносмилюс» . ucmp.berkeley.edu . Проверено 4 сентября 2022 г.
- ^ Мерриам, JC и Сток, C. 1932: Кошачьи с ранчо Ла Бреа. Публикации Вашингтонского института Карнеги 442, 1–231.
- ^ ДеСантис, ЛР; Шуберт, Б.В.; Скотт, младший; Унгар, П.С. (2012). «Последствия диеты для вымирания саблезубых кошек и американских львов» . ПЛОС ОДИН . 7 (12): е52453. Бибкод : 2012PLoSO...752453D . дои : 10.1371/journal.pone.0052453 . ПМЦ 3530457 . ПМИД 23300674 .
- ^ Манти, Фредрик К.; Браун, Фрэнсис Х.; Плавкан, Майкл Дж.; Верделин, Ларс (март 2018 г.). «Гигантский лев Panthera leo из плейстоцена Натодомери, Восточная Африка » Журнал палеонтологии . 92 (2): 305–312. Бибкод : 2018JPal...92..305M . дои : 10.1017/jpa.2017.68 . ISSN 0022-3360 .
- ^ Перейти обратно: а б Арган, Ален; Аргант, Жаклин (2011). « История Panthera gombaszogensis : вклад Шато Брекча (Сона и Луара, Бургундия, Франция)» . Четвертичный период (Спецвыпуск 4): 247–269.
- ^ Энн Шмидт-Кюнцель; Лори Маркер; Лоррейн К. Боаст (2017). Гепарды: биология и охрана . Эльзевир Наука. п. 30. ISBN 978-0-12-804120-8 . Проверено 4 сентября 2022 г.
- ^ Лейн, Х.Х. (1947). «Обзор ископаемых позвоночных Канзаса: Часть V: Млекопитающие (продолжение)» (PDF) . Труды Канзасской академии наук (1903–) . 50 (3/4). Канзасская академия наук (опубликовано в декабре 1947 г.): 273–314. дои : 10.2307/3625600 . JSTOR 3625600 .
- ^ Энтони, Морис (22 ноября 2013 г.). Саблезубый . Издательство Университета Индианы. стр. 100-1 104–107. ISBN 978-0-253-01049-0 . Проверено 26 августа 2022 г.
- ^ Алан Тернер, National Geographic Доисторические млекопитающие National Geographic, 2004 г., ISBN 0-7922-7134-3
- ^ Тернер, Алан; Антон, Маурисио (1996). «Гигантская гиена Pachycrocuta brevirostris (Mammalia, Carnivora, Hyaenidae)». Геобиос . 29 (№ 4): 455–468. Бибкод : 1996Geobi..29..455T . дои : 10.1016/S0016-6995(96)80005-2 .
- ^ Фланнери, Тим (2018). Европа. Первые 100 миллионов лет . Пингвин Букс Лимитед. п. 29: Другие гиганты умеренного пояса. ISBN 978-0-14-198903-7 . Проверено 21 августа 2022 г.
- ^ «Проклятие Саблезубого Зуба: Знакомство с Динокрокутой» . WordPress.com . 12 января 2016 г. Архивировано из оригинала 12 ноября 2020 г.
- ^ Личное сообщение Р. Дьюера в Бернессе и др., 2001, таблица 1.
- ^ Роу и др., 2004, с. 297
- ^ Гудман, С. (2009). «Семейство Eupleridae (Мадагаскарские хищники)». В Уилсоне, Д.; Миттермайер, Р. (ред.). Справочник млекопитающих мира. Том 1: Хищники . Рысь Эдиционс . ISBN 978-84-96553-49-1 . Архивировано из оригинала 25 июля 2011 года . Проверено 31 мая 2010 г.
- ^ Бортс, MR; Стивенс, Нью-Джерси (2019). « Simbakubwa kutokaafrika , gen. et sp. nov. (Hyainailourinae, Hyaenodonta, «Creodonta», Mammalia), гигантское хищное животное из раннего миоцена Кении» . Журнал палеонтологии позвоночных . 39 (1): e1570222. Бибкод : 2019JVPal..39E0222B . дои : 10.1080/02724634.2019.1570222 . S2CID 145972918 .
- ^ Сэвидж, RJG (1973). « Мегистотерий , гигантский гиенодонт из миоцена Гебель-Зельтена, Ливия» . Бюллетень Британского музея (Естественная история), Геология . 22 (7): 483–511. дои : 10.5962/стр.150151 .
- ^ Перейти обратно: а б N. N. Kramarenko (1974). Зоогеография палеогена Азии (Zoogeography of Paleogene of Asia) . Publishing office "Nauka". pp. 113–114 . Retrieved 18 September 2022 .
- ^ О'Лири, Морин А.; Лукас, Спенсер Г.; Уильямсон, Томас Э. (2000). «Новый экземпляр Анкалагона (Mammalia, Mesonychia) и свидетельства полового диморфизма у мезонихий». Журнал палеонтологии позвоночных . 20 (2): 387–93. doi : 10.1671/0272-4634(2000)020[0387:ANSOAM]2.0.CO;2 . JSTOR 4524103 . S2CID 86542114 .
- ^ Палеоценовые млекопитающие мира «Плотоядные, креодонты и хищные копытные: млекопитающие становятся хищниками»
- ^ «В Аргентине найдена окаменелость гигантской летучей мыши-вампира возрастом 100 000 лет - Палеонтология - Sci-News.com» . sci-news.com . 26 июля 2021 г.
- ^ Вилье, Борис (2010). «Дейногалерикс: гигантский еж из миоцена». Анналы Феррарского университета . 6 :93–102. ISSN 1824-2707 .
- ^ Фрейденталь, М. (1972). « Deinogalerix koenigswaldi nov. gen., nov. spec., гигантское насекомоядное животное из неогена Италии». Скрипта Геологика . 14 : 1–19.
- ^ Перейти обратно: а б Энгельман, Рассел К. (июнь 2022 г.). «Изменение размера крупнейших известных вымерших грызунов (Caviomorpha: Dinomyidae, Neoepiblemidae) с использованием ширины затылочного мыщелка» . Королевское общество открытой науки . 9 (6): 220370. Цифровой код : 2022RSOS....920370E . дои : 10.1098/rsos.220370 . ПМЦ 9198521 . ПМИД 35719882 .
- ^ Перейти обратно: а б с Дефлер, Томас (2018). История наземных млекопитающих Южной Америки . Международное издательство Спрингер. стр. 151–154. ISBN 978-3-319-98449-0 . Проверено 27 августа 2022 г.
- ^ Бикневичус, Арканзас; Макфарлейн, округ Колумбия; Макфи, RDE (1993). «Размер тела Amblyrhiza inundata (Rodentia: Caviomorpha), вымершего грызуна-мегафауны с берега Ангильи, Вест-Индия: оценки и последствия». Американский музей послушников (3079 г.). Нью-Йорк: Американский музей естественной истории : 1–25. hdl : 2246/4976 .
- ^ Суайнхарт, Энтони Л.; Ричардс, Рональд Л. (2001). «Палеоэкология водно-болотных угодий северо-восточной Индианы, где обитают останки плейстоценового гигантского бобра (Castoroides Ohioensis)» . Труды Академии наук Индианы . 110 :151 . Проверено 18 августа 2022 г.
- ^ Милиус, Сьюзен (2011). « Кролик, который правил Меноркой ». Новости науки . 179 (9): 18. doi : 10.1002/scin.5591790921 .
- ^ Годен, Тимоти Дж.; Эмри, Роберт Дж.; Уибл, Джон Р. (декабрь 2009 г.). «Филогения ныне живущих и вымерших панголинов (Mammalia, Pholidota) и связанных с ними таксонов: морфологический анализ» . Журнал эволюции млекопитающих . 16 (4): 235–305. дои : 10.1007/s10914-009-9119-9 . ISSN 1064-7554 . S2CID 1773698 .
- ^ Хойер, Д.А., 1947 - Бедренная кость Manis palaeojavanica Dubois с Западной Явы - Труды Королевской Нидерландской академии искусств и наук 50 (4): 423–418
- ^ Чочон, Рассел Л. «Обезьяна, которая была - азиатские окаменелости раскрывают гигантского кузена человечества» . Университет Айовы. Архивировано из оригинала 25 мая 2015 года . Проверено 7 сентября 2022 г.
- ^ Петтифор, Эрик (2000) [1995]. «Из зубов дракона: Гигантопитек Блэки » . Избранные материалы по физической антропологии . Кендалл/Хант Издательская компания. стр. 143–149. ISBN 978-0-7872-7155-8 . Архивировано из оригинала 5 марта 2016 года . Проверено 7 сентября 2022 г.
- ^ Чжан, Инци; Харрисон, Терри (январь 2017 г.). «Gigantopithecus blacki: повторное посещение гигантской обезьяны из плейстоцена Азии» . Американский журнал физической антропологии . 162 (С63): 153–177. дои : 10.1002/ajpa.23150 . ISSN 0002-9483 . ПМИД 28105715 . S2CID 46838584 .
- ^ Вибово, А. « Шаги азиатских гигантов: моделирование экологии кормления Meganthropus palaeojavanicus, связанной с размером тела, 8-футового гоминида в Центральной Яве ». Препринты 2020, 2020110504 (doi: 10.20944/preprints202011.0504.v1).
- ^ Занолли, Клеман; Куллмер, Оттмар; Келли, Джей; Бэкон, Анн-Мари; Деметра, Фабрис; Дюмонсель, Жан; Фиоренца, Лука; Грин, Фредерик Э.; Юблин, Жан-Жак; Нгуен, Ань Туан; Нгуен, Ти Май Хуонг (май 2019 г.). «Доказательства увеличения разнообразия гоминид в раннем и среднем плейстоцене Индонезии» . Экология и эволюция природы . 3 (5): 755–764. Бибкод : 2019NatEE...3..755Z . дои : 10.1038/s41559-019-0860-z . ISSN 2397-334X . ПМИД 30962558 . S2CID 102353734 .
- ^ Фрол А.В., Черчилль С.Е. (2009). «Энергетическая конкуренция между неандертальцами и анатомически современными людьми» (PDF) . Палеоантропология : 96–116 . Проверено 12 сентября 2022 г.
- ^ Перейти обратно: а б Роудман, Джозеф-Майкл; Родригес, Лаура; Гарсиа-Гонсалес, Ребекка; Арсуага, Джон-Луи; Гомес-Оливенс, Азиер; Лоуренс, Чарльз; Бонмати, Александр; Грейс, Ханна; Мартинес, Игнатиус (2012). «Оценка роста по целым длинным костям людей среднего плейстоцена из Сима-де-лос-Бонес, Сьерра-де-Атапуэрка (Испания)» (PDF) . Журнал эволюции человека . 62 (2): 242–255. Бибкод : 2012JHumE..62..242C . дои : 10.1016/j.jhevol.2011.11.004 . ПМИД 22196156 .
- ^ Алан Уокер, Ричард Лики (1993). Скелет Нариокотомэ человека прямоходящего . Издательство Гарвардского университета. п. 412. ИСБН 978-0-674-60075-1 . Проверено 2 октября 2022 г.
- ^ Мильяно AB, Гийон М (2012). «Влияние смертности, существования и экологии на рост взрослого человека и последствия для эволюции человека » . Современная антропология . 53 (С6): 359–368. дои : 10.1086/667694 . S2CID 84442763 .
- ^ Делсон, Эрик; Терранова, Карл Дж.; Юнгерс, Уильям Дж; Саргис, Саргис; Яблонски, Нина Г.; Дешоу, Пол К. (2000). «Масса тела Cercopithecidae (приматы, млекопитающие): оценка и масштабирование вымерших и современных таксонов». Антропологические документы Американского музея естественной истории . 83 : 1–159.
- ^ Яблонски, Нина; Лики, Мив; Антон, Маурисио (1 января 2008 г.), Яблонски, Н.Г. Лики, М.Г. и Антон, М. (2008) Систематическая палеонтология церкопитеков. В: Яблонски Н.Г. и Лики М.Г. (ред.) Исследовательский проект Koobi Fora. Том 6. Ископаемые обезьяны. Калифорнийская академия наук, Сан-Франциско, стр. 103–300. , стр. 103–300 , получено 10 сентября 2022 г.
- ^ Перри, Джонатан М.Г.; Кук, Шивон Б.; Рунестад Коннур, Жаклин А.; Берджесс, М. Лоринг; Рафф, Кристофер Б. (2018). «Суставное масштабирование и оценка массы тела у платиррин и катаров: современные вариации и применение к ископаемым антропоидам» . Журнал эволюции человека . 115 : 20–35. Бибкод : 2018JHumE.115...20P . дои : 10.1016/j.jhevol.2017.10.008 . ISSN 1095-8606 . ПМИД 29150186 .
- ^ Рэйчел Х. Данн. « Дополнительные посткраниальные останки омомидных приматов из формации Уинта, штат Юта, и последствия для двигательного поведения крупнотелых омомид ». Журнал эволюции человекаТом 58, выпуск 5, май 2010 г., стр. 406–417.
- ^ Юнгерс, В.Л.; Демес, Б.; Годфри, ЛР (2008). «Насколько большими были «гигантские» вымершие лемуры Мадагаскара?». Во Флигле, JG; Гилберт, CC (ред.). Элвин Саймонс: В поисках истоков . Развитие приматологии: прогресс и перспективы. п. 350. дои : 10.1007/978-0-387-73896-3_23 . ISBN 978-0-387-73895-6 .
- ^ Годфри, ЛР; Юнгерс, WL (2002). «Четвертичные ископаемые лемуры». В Хартвиге, У.К. (ред.). Летопись окаменелостей приматов . Издательство Кембриджского университета . п. 101. ИСБН 978-0-521-66315-1 .
- ^ Перейти обратно: а б Кроули, Б.Э., и Годфри, Л.Р. (2019). « Изотопы стронция поддерживают небольшие места обитания вымерших лемуров ». Frontiers in Ecology and Evolution, 7, 490. doi: 10.1002/ajp.20817.
- ^ Megaladapis edwardsi : Научная информация . Архивная копия от 20 января 2021 года.
- ^ Осборн, HF (1942). Хоботки, Vol. II . Нью-Йорк: Издательство Американского музея.
- ^ Листер, А.; Бан, П. (2007). Мамонты – гиганты ледникового периода (3-е изд.). Лондон: Фрэнсис Линкольн. ISBN 978-0-520-26160-0 .
- ^ Мол, Д. и ван Логчем, В. 2009. Мастодонт Милии: самые длинные бивни в мире. Депозиты 19: 28–32.
- ^ Марш, Хелен ; О'Ши, Томас Дж.; Рейнольдс III, Джон Э. (2011). «Стеллерова морская корова: открытие, биология и эксплуатация реликтовой гигантской сирены» . Экология и охрана сирен: дюгони и ламантины . Нью-Йорк, Нью-Йорк: Издательство Кембриджского университета . стр. 18–35. ISBN 978-0-521-88828-8 . OCLC 778803577 .
- ^ Шеффер, Виктор Б. (ноябрь 1972 г.). «Вес стеллеровой коровы». Журнал маммологии . 53 (4): 912–914. дои : 10.2307/1379236 . JSTOR 1379236 .
- ^ Эндрюс, CW (1906). Описательный каталог третичных позвоночных Фаюма. Британский музей, Лондон . Тейлор и Фрэнсис. п. 324.
- ^ Мондехар-Фернандес; и др. (2008). «Род Arsinoitherium : каталог неопубликованной коллекции Музея естественной истории в Париже и проблема количества видов» . Palaeontologica Nova (на испанском языке). СЕПАЗ (8): 292–304.
- ^ Сандерс, В.Дж., Каппельман, Дж. и Расмуссен, Д.Т., 2004. Польский палеонтологический акт 49 (3): 365–392. [1] .
- ^ Перейти обратно: а б Роуз 2006 , с. 260
- ^ Родольф Табусе (2016). «Нижняя челюсть гиракоидного млекопитающего Titanohyrax andrewsi в коллекции Национального музея естественной истории, Париж (Франция) с переоценкой вида» . Палеовертебрата . 40 (1): e4. дои : 10.18563/pv.40.1.e4 .
- ^ JGM Тьювиссен; Э. Л. Саймонс (2001). «Череп Megalohyrax eocaenus (Hyracoidea, Mammalia) из олигоцена Египта». Журнал палеонтологии позвоночных . 21 (1): 98–106. doi : 10.1671/0272-4634(2001)021[0098:SOMEHM]2.0.CO;2 . JSTOR 4524175 . S2CID 86063305 .
- ^ Дональд Р. Протеро; Роберт М. Шох (1989). Эволюция непарнокопытных . Издательство Оксфордского университета. п. 65. ИСБН 978-0-19-506039-3 . Проверено 20 сентября 2022 г.
- ^ Скиннер, доктор медицинских наук; Чимимба, Кристиан Т. (2005). Млекопитающие южноафриканского субрегиона . Издательство Кембриджского университета. п. 41. ИСБН 978-1-107-39405-6 . Проверено 20 сентября 2022 г.
- ^ Верделин, Ларс; Сандерс, Уильям Джозеф (2010). Кайнозойские млекопитающие Африки . Издательство Калифорнийского университета. п. 143. ИСБН 978-0-520-25721-4 . Проверено 20 сентября 2022 г.
- ^ Пьенсон, Николас Д.; Вермей, Гират Дж. (2016). «Восстание океанских гигантов: максимальный размер тела кайнозойских морских млекопитающих как индикатор продуктивности Тихого и Атлантического океанов» . Письма по биологии . 12 (7). дои : 10.1098/rsbl.2016.0186 . ПМЦ 4971165 . ПМИД 27381883 . S2CID 1652462 .
- ^ Хаяси, Сёдзи; Уссей, Александра; Накадзима, Ясухиса; Тиба, Кентаро; Андо, Три; Савамура, Хироши; Инузука, Разрушитель; Канеко, Наотомо; Осаки, Томохиро (2 апреля 2013 г.). «Внутренняя структура костей предполагает усиление водной адаптации у Desmostylia (Mammalia, Afrotheria) » ПЛОС ОДИН 8 (4): e59146. Бибкод : 2013PLoSO... 859146H дои : 10.1371/journal.pone.0059146 . ISSN 1932-6203 . ПМК 3615000 . ПМИД 23565143 .
- ^ Листер, Адриан (24 апреля 2018 г.). Окаменелости Дарвина. Коллекция, сформировавшая теорию эволюции . Смитсоновский институт. п. 57. ИСБН 978-1-58834-617-9 . Проверено 4 сентября 2022 г.
- ^ Лармон, Джин Т.; Макдональд, Х. Грегори; Эмброуз, Стэнли; ДеСантис, Лариса Р.Г.; Лусеро, Лиза Дж. (27 февраля 2019 г.). «Год из жизни гигантского наземного ленивца во время последнего ледникового максимума в Белизе» . Достижения науки . 5 (2): eaau1200. Бибкод : 2019SciA....5.1200L . дои : 10.1126/sciadv.aau1200 . ПМК 6392778 . ПМИД 30820449 .
- ^ «BBC – Наука и природа – Дикие факты – Мегатерий» . 1 февраля 2014 года. Архивировано из оригинала 1 февраля 2014 года . Проверено 29 июня 2017 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Джонсон, Стивен К. и Мэдден, Ричард Х.. Uruguaytheriinae Astrapotheres тропической Южной Америки. Глава 22 в «Палеонтологии позвоночных в неотропах. Миоценовая фауна Ла-Венты, Колумбия». Под редакцией Ричарда Ф. Кея, Ричарда Х. Мэддена, Ричарда Л. Чифелли и Джона Дж. Флинна. Издательство Смитсоновского института. Вашингтон и Лондон.
- ^ Крамарц, Александр Георгиевич; Бонд, Мариано (2008). «Ревизия парастрапотериев (Mammalia, Astrapotheria) и других желательных астрапотериев Патагонии » Амегиниана . 45 (3 ) Получено 1 марта.
- ^ Каррильо, Джон Д.; Амсон, Эли; Харамильо, Чарльз; Санчес, Родольфо; Кирос, Луи; В-четвертых, Чарльз; Корнер, Альдо Ф.; Санчес-Вильягра, Марсело Р. (2018). «Неогеновые находки местных копытных северной части Южной Америки» . Смитсоновский вклад в палеобиологию . 101 (101): iv-6 дои : 10.5479/si.1943-6688.101 . S2CID 135113342 .
- ^ Джинджерич, Филип Д. (1998). «Палеобиологические перспективы мезонихии, археоцитов и происхождения китов». В Тьювиссене, JGM (ред.). Появление китов: эволюционные закономерности происхождения китообразных . Нью-Йорк: Пленум Пресс. стр. 423–450. ISBN 978-0-306-45853-8 .
- ^ Элиссамбуру, А (2012). «Оценка массы тела у родов отряда Notoungulata» . Геологические исследования . 68 (1): 91–111. дои : 10.3989/egeol.40336.133 . hdl : 11336/197170 .
- ^ Крофт, округ Колумбия; Гелфо, Дж. Н.; Лопес, генеральный менеджер (2020). «Великолепные инновации: вымершие коренные копытные Южной Америки» . Ежегодный обзор наук о Земле и планетах . 48 : 259–290. Бибкод : 2020AREPS..48..259C . doi : 10.1146/annurev-earth-072619-060126 . S2CID 213737574 .
- ^ Дортангс, Руди В.; Шулп, Энн С.; Эрик В.А. Малдер; Джон В.М. Джагт (2002). «Большой новый мозазавр из верхнего мела Нидерландов» . Нидерландский журнал геонаук . 81 (1): 1–8. Бибкод : 2002NJGeo..81....1D . дои : 10.1017/S0016774600020515 .
- ^ Григорьев, Дмитрий В. (2014). «Гигантский Mosasaurus hoffmanni (Squamata, Mosasauridae) из позднего мела (маастрихта) Пензы, Россия» (PDF ) Труды Зоологического института РАН . 318 (2): 148–167. дои : 10.31610/трудызин/2014.318.2.148 . S2CID 53574339 .
- ^ Фанти, Федерико; Кау, Андреа; Негри, Алессандра (1 мая 2014 г.). «Гигантский мозазавр (Reptilia, Squamata) с необычно искривленным зубным рядом из комплекса Аржилле Скальозе (поздний кампан) Северной Италии» . Меловые исследования . 49 : 91–104. Бибкод : 2014CrRes..49...91F . дои : 10.1016/j.cretres.2014.01.003 . ISSN 0195-6671 .
- ^ Эверхарт, Майк. «Исследование: Tylosaurus proriger – новая находка большого мозазавра из мела Смоки-Хилл» . Океаны Канзаса . Проверено 12 мая 2010 г.
- ^ «Факты: Тилозавр Проригер из National Geographic» . Архивировано из оригинала 7 февраля 2010 года . Проверено 12 мая 2010 г.
- ^ Руководитель Джейсон; Блох, Джонатан; Морено Берналь, Хорхе; Ринкон Бурбано, Альдо Фернандо; Бурк, Джейсон (2013), Краниальная остеология, размер тела, систематика и экология гигантской палеоценовой змеи Titanoboa cerrejonensis , Общество палеонтологии позвоночных , стр. 140–141 , получено 22 мая 2017 г.
- ^ Перейти обратно: а б Руководитель Джейсон Дж.; и др. (5 февраля 2009 г.). «Гигантская змея-боид из неотропов палеоцена демонстрирует более высокие температуры за экваториальными температурами» (PDF) . Природа . 457 (7230): 715–717. Бибкод : 2009Natur.457..715H . дои : 10.1038/nature07671 . ПМИД 19194448 . S2CID 4381423 . Проверено 12 мая 2010 г.
- ^ Датта, Дебаджит; Баджпай, Сунил (18 апреля 2024 г.). «Самая крупная известная змея-мадцоид из теплого эоценового периода Индии предполагает межконтинентальное распространение в Гондване» . Научные отчеты . 14 (1): 8054. Бибкод : 2024NatSR..14.8054D . дои : 10.1038/s41598-024-58377-0 . ISSN 2045-2322 . ПМИД 38637509 .
- ^ Ярость, Жан-Клод; Баджпай, Сунил; Йоханнес Г.М. Тьювиссен; Тивари, Брахма Н. (2003). «Змеи раннего эоцена из Катча, Западная Индия, с обзором Palaeophiidae» (PDF) . Георазнообразие . 25 (4): 695–716. ISSN 1280-9659 . Проверено 12 мая 2010 г.
- ^ Ярость, Ж.-К. (1983). « Palaeophis colossaeus nov. sp. (крупнейший известный сепрент?) из эоцена Мали и проблема рода среди Palaeopheinae». Отчеты сессий Академии наук . 3 (296): 1741–1744.
- ^ «Большие палеофиидные и нигерофиидные змеи из палеогеновых отложений Транссахарского морского пути в Мали - Acta Palaeontologica Polonica» . www.app.pan.pl. Проверено 8 января 2021 г.
- ^ Хэд, Дж. и Полли, Д. 2004. Они могут быть гигантами: морфометрические методы реконструкции размера тела крупнейших змей в мире. Журнал палеонтологии позвоночных 24 (Приложение 3), 68A-69A.
- ^ «Гигант среди змей» . newscientist.com .
- ^ Рио, Джонатан П.; Маннион, Филип Д. (4 июля 2017 г.). «Остеология гигантской змеи Gigantophis garstini из верхнего эоцена Северной Африки и ее влияние на филогенетические отношения и биогеографию Madtsoiidae». Журнал палеонтологии позвоночных . 37 (4): e1347179. Бибкод : 2017JVPal..37E7179R . дои : 10.1080/02724634.2017.1347179 . hdl : 10044/1/48946 . ISSN 0272-4634 . S2CID 90335531 .
- ^ Скэнлон, Джон Д.; Макнесс, Брайан С. (1 января 2001 г.). «Новый гигантский питон из плиоценовой местной фауны Блафф-Даунс на северо-востоке Квинсленда». Алчеринга: Австралазийский журнал палеонтологии . 25 (4): 425–437. Бибкод : 2001Alch...25..425S . дои : 10.1080/03115510108619232 . ISSN 0311-5518 . S2CID 85185368 .
- ^ Оуэн, Р. (1857). «Об ископаемых позвонках змеи (Laophis crotaloides, Ow.), обнаруженных капитаном Спраттом, штат Р.Н., в третичной формации в Салониках» . Ежеквартальный журнал Геологического общества . 13 (1–2): 196–199. дои : 10.1144/GSL.JGS.1857.013.01-02.28 . S2CID 131142130 .
- ^ Бенджамин П. Кир (2014). «Повторное открытие Laophis crotaloides – крупнейшей гадюки в мире?» . www.researchgate.net . Проверено 14 сентября 2022 г.
- ^ Бэйлон С., Бовер П., Кинтана Дж. и Альковер Дж. А. (2010). Первая находка окаменелостей гадюки Лауренти 1768 г. «Комплекс восточных гадюк» (Serpentes: Viperidae) из раннего плиоцена островов западного Средиземноморья . Комптес Рендус Палевол, 9, 147–154.
- ^ Фачини, Тьяго Шнайдер; Онари, Сильвио; Пальчи, Алессандро; Ли, Майкл С.Ю.; Бронзати, Марио; Сю, Энни Шмальц (18 декабря 2020 г.). «Меловая слепая змея из Бразилии заполняет серьезный пробел в эволюции змей» . iScience . 23 (12): 101834. Бибкод : 2020iSci...23j1834F . дои : 10.1016/j.isci.2020.101834 . ISSN 2589-0042 . ПМЦ 7718481 . ПМИД 33305189 .
- ^ Перейти обратно: а б с Молнар, Р.Э. (2004). Палеобиология гигантского варана Мегалании . Издательство Университета Индианы . стр. 100-1 174–175. ISBN 0-253-34374-7 . OCLC 52775128 .
- ^ Фрай, Б.; Роу, С.; Теувисс, В.; Ван Ош, MJP; Морено, К.; Ингл, Дж.; МакГенри, К.; Феррара, Т.; Клаузен, П.; Шейб, Х.; Зима, КЛ; Грейсман, Л.; Ролантс, К.; Ван Дер Верд, Л.; Клементе, CJ; Яннакис, Э.; Ходжсон, WC; Луз, С.; Мартелли, П.; Кришнасами, К.; Кочва, Е.; Квок, ХФ; Скэнлон, Д.; Карас, Дж.; Цитрон, ДМ; Гольдштейн, ЕЕК; Макнотан, JE; Норман, Дж.А.; и др. (2009). «Центральная роль яда в хищниче Varanus komodoensis (Комодский варан) и вымершего гиганта Varanus ( Megalania ) priscus » . ПНАС . 106 (22): 8969–74. Бибкод : 2009PNAS..106.8969F . дои : 10.1073/pnas.0810883106 . ПМК 2690028 . ПМИД 19451641 .
- ^ «Обзор драконов в пыли: палеобиология гигантского варана Мегалании» , Ральф Э. Молнар, 2004 г., в Журнале палеонтологии позвоночных 25 (2): 479, июнь 2005 г. (PDF) . Архивировано из оригинала (PDF) 27 января 2016 года . Проверено 16 сентября 2022 г.
- ^ Колстон, Ти Джей; Кулкарни, П.; Джетц, В.; Пайрон, РА (2020). «Филогенетическое и пространственное распределение эволюционного разнообразия, изоляции и угроз у черепах и крокодилов (нептичьих архозавроморфов)» . Эволюционная биология BMC . 20 (1): 81. Бибкод : 2020BMCEE..20...81C . дои : 10.1186/s12862-020-01642-3 . ПМЦ 7350713 . ПМИД 32650718 .
- ^ Перейти обратно: а б Майк Эверхарт. «Морские черепахи Западного Внутреннего моря» . Океаны Канзаса . Архивировано из оригинала 7 апреля 2022 года.
- ^ Игорь Геннадьевич Данилов; Образцова Екатерина Михайловна; Максим Саввич Архангельский; Алексей Владимирович Иванов; Александр Аверьянов (2022). «Protostega gigas и другие морские черепахи кампана Восточной Европы, Россия» . Меловые исследования . 135 : 105196. Бибкод : 2022CrRes.13505196D . дои : 10.1016/j.cretres.2022.105196 . S2CID 247431641 .
- ^ Дерстлер, К.; Лейтч, AD; Ларсон, Польша; Финсли, К.; Хилл, Л. (1993). «Самые большие черепахи в мире - Венский архелон (4,6 м) и Далласская протостега (4,2 м), верхний мел Южной Дакоты и Техаса» . Журнал палеонтологии позвоночных . 13 (дополнение к № 3) (33А).
- ^ Кир, Бенджамин П. (11 сентября 2006 г.). «Переоценка Cratochelone Berneyi Longman, 1915, гигантской морской черепахи из раннего мела Австралии» . Журнал палеонтологии позвоночных . 26 (3): 779–783. doi : 10.1671/0272-4634(2006)26[779:ROCBLA]2.0.CO;2 . ISSN 0272-4634 . S2CID 140149175 .
- ^ HF Кадмий (2006). «Новый род морских черепах (Chelonioidea: Cheloniidae) из маастрихта фауны Харраны-Иордания» (PDF ) Журнал палеонтологии позвоночных PalArch . 3 (1): 1–14. Архивировано из оригинала (PDF) 24 февраля . Получено 4 февраля.
- ^ Р. Колер (1995). «Новый вид ископаемой черепахи Psephophorus (отряд Testudines) из эоцена Южного острова, Новая Зеландия» . Журнал Королевского общества Новой Зеландии . 25 (3): 371–384. Бибкод : 1995JRSNZ..25..371K . дои : 10.1080/03014223.1995.9517495 . Архивировано из оригинала 4 октября 2021 года.
- ^ Нэйш, Даррен (31 января 2024 г.). «Мегалохелис, настоящая гигантская черепаха» . Зоология четвероногих . Проверено 30 мая 2024 г.
- ^ Перейти обратно: а б с д и Родин, Андерс Г.Дж.; Томсон, Скотт; Георгалис, Георгиос Л.; Карл, Ганс Фолькер; Данилов Игорь Георгиевич; Такахаши, Акио; де ла Фуэнте, Марсело СаулИкон; Бурк, Джейсон; Дельфино, Массимо; Бур, Роджер; Айверсон, Джон Б.; Шаффер, Брэдли Х.; ван Дейк, Питер Пол (2015). «Черепахи и черепахи мира во время подъема и глобального распространения человечества: первый контрольный список и обзор вымерших плейстоценовых и голоценовых хелоний» (PDF) . Челонианские исследовательские монографии . 5 : 1–66. doi : 10.3854/crm.5.000e.fossil.checklist.v1.2015 . ISBN 978-0-9653540-9-7 . ISSN 1088-7105 . Архивировано (PDF) из оригинала 21 сентября 2022 года.
- ^ Перес-Гарсия, Адан; Влахос, Евангелос (1 ноября 2014 г.). «Новое родовое предложение для крупных тестудинид европейского неогена (Cryptodira) и первая филогенетическая гипотеза для средних и крупных представителей европейской кайнозойской летописи» . Зоологический журнал Линнеевского общества . 172 (3): 653–719. дои : 10.1111/zoj12183 . ISSN 0024-4082 .
- ^ Перес-Гарсия, Адам; Влахос, Евангелия; Аррибас, Альфонсо (март 2017 г.). «Последняя гигантская континентальная черепаха Европы: выживший на стоянке Фонелас P-1 в испанском плейстоцене» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 470 : 30–39. Бибкод : 2017PPP...470...30P . дои : 10.1016/j.palaeo.2017.01.011 . hdl : 10261/277114 . Архивировано из оригинала (PDF) 26 января 2022 года.
- ^ Перейти обратно: а б Джейсон Дж. Хед; Филип Д. Джинджерич; С. Махмуд Раза (1999). «Drazinderetes tethyensis, новый крупный трионихид (Reptilia: Testudines) из морской эоценовой формации Дразинда Сулейманского хребта, Пенджаб (Пакистан)» (PDF) . Музей палеонтологии Мичиганского университета . стр. 100-1 199–214. Архивировано (PDF) из оригинала 13 октября.
- ^ Гранде, Лэнс (14 июня 2013 г.). Затерянный мир Ископаемого озера: снимки из глубокого времени . Издательство Чикагского университета. п. 200. ИСБН 978-0-226-92296-6 .
- ^ Цепь, Э.-А.; Шайер, ТМ; Лейн-Брицено, доктор юридических наук; Санчес, Р.; Игл-Рельеф, ОА; Ванегас, А.; Браун, М.; Хансен, DM; Санчес-Вильягра, MR (12 февраля 2020 г.). «Анатомия, палеобиология и эволюционные взаимоотношения крупнейшей вымершей боковой черепахи» . Достижения науки . 6 (7): иаай4 Бибкод : 2020SciA.... 6.4593C дои : 10.1126/sciadv.aay4593 . ПМК 7015691 . ПМИД 32095528 .
- ^ «Исследователи обнаружили окаменелость древней гигантской черепахи» . Физика.орг . 17 мая 2012 г. Архивировано из оригинала 22 сентября 2022 г. . Проверено 13 октября 2022 г.
- ^ Мо II, Томас Х. (18 мая 2012 г.). «Исследователи нашли окаменелость черепахи размером с автомобиль Smart» . Лос-Анджелес Таймс . Архивировано из оригинала 23 сентября 2022 года . Проверено 13 октября 2022 г.
- ^ Фриман, Дэвид (17 мая 2012 г.). «Рептилия размером с автомобиль жила рядом с Титанобоа, говорят ученые» . Хаффингтон Пост . Архивировано из оригинала 23 сентября 2022 года . Проверено 13 октября 2022 г.
- ^ Эдвин А. Кадена; Дэниел Т. Ксепка; Карлос А. Харамильо; Джонатан И. Блох (2012). «Новые пеломедузоидные черепахи из позднепалеоценовой формации Серрехон в Колумбии и их значение для филогении и эволюции размеров тела» . Журнал систематической палеонтологии . 10 (2): 313–331. Бибкод : 2012JSPal..10..313C . дои : 10.1080/14772019.2011.569031 . S2CID 59406495 .
- ^ Тонг, Хайян; Чантасит, Форнфен; Наксри, Вилайлак; Дитбанджонг, Питаксит; Сутитхорн, Суравек; Баффето, Эрик; Сутитхорн, Варавуд; Вонгко, Камонлак; Дисри, Утумпорн; Клод, Жюльен (ноябрь 2021 г.). «Yakemys multiporcata ngn sp., крупная черепаха-макробенид из базального мела Таиланда, с обзором черепаховой фауны формации Фу-Крадунг и ее стратиграфическими последствиями» . Разнообразие . 13 (12): 630. дои : 10.3390/d13120630 .
- ^ Лорен Э. Браун, Дон Молл (октябрь 2019 г.). «Загадочная палеоэкология и палеобиогеография гигантских рогатых ископаемых черепах Австралазии: обзор и повторный анализ данных» . Герпетологический журнал . 29 (4): 252–263. дои : 10.33256/hj29.4.252263 . ISSN 0268-0130 . Архивировано из оригинала 18 июня 2022 года.
- ^ Росс Д.Е. Макфи, Ханс-Дитер Сьюс (1999). Вымирания в ближайшем будущем: причины, контексты и последствия . Издательство Kluwer Academic/Plenum. п. 251. ИСБН 978-0-306-46092-0 .
- ^ «ПЛАКОДОНТЫ: Причудливые «моржи-черепахи» триаса» . Океаны Канзаса . Архивировано из оригинала 30 января 2022 года.
- ^ Лю, Цзюнь (27 ноября 2014 г.). «Гигантский нотозавр (Reptilia: Sauropterygia) из среднего триаса на юго-западе Китая и его значение для триасового восстановления биотики» . Научные отчеты . 4 : 7142. Бибкод : 2014NatSR...4E7142L . дои : 10.1038/srep07142 . ПМЦ 4245812 . ПМИД 25429609 .
- ^ О'Горман, JP; Сантильяна, С.; Отеро, Р.; Регеро, М. (1 октября 2019 г.). «Гигантский эласмозаврид (Sauropterygia; Plesiosauria) из Антарктиды: новая информация о разнообразии размеров тела эласмозаврид и сценариях эволюции аристонектина» . Меловые исследования . 102 : 37–58. Бибкод : 2019CrRes.102...37O . дои : 10.1016/j.cretres.2019.05.004 . ISSN 0195-6671 . S2CID 181725020 .
- ^ Кубо, Т.; Митчелл, Монтана; Хендерсон, DM (2012). « Albertonectes vanderveldei , новый эласмозавр (Reptilia, Sauropterygia) из верхнего мела Альберты». Журнал палеонтологии позвоночных . 32 (3): 557–572. Бибкод : 2012JVPal..32..557K . дои : 10.1080/02724634.2012.658124 . S2CID 129500470 .
- ^ Смит, Эллиотт (1 января 2020 г.). «Ревизия рода Styxosaurus и взаимоотношений позднемеловых эласмозаврид (Sauropterygia: Plesiosauria) Западного внутреннего морского пути» . Диссертации, диссертации и основные положения .
- ^ Майк Эверхарт (2005). «Следы укусов на весле эласмозавра (Sauropterygia; Plesiosauria) из мела Ниобрара (верхний мел) как вероятное свидетельство питания пластинчатой акулой Cretoxyrhina mantelli » (PDF) . Журнал палеонтологии позвоночных PalArch . 2 (2). Архивировано (PDF) из оригинала 16 июля 2011 года . Проверено 2 октября 2009 г.
- ^ Перейти обратно: а б с О'Горман, JP (2016). «Неаристонектиновый эласмозаврид небольшого размера (Sauropterygia, Plesiosauria) из позднего мела Патагонии с комментариями о взаимоотношениях патагонских и антарктических эласмозавридов» . Амегиниана . 53 (3): 245–268. дои : 10.5710/AMGH.29.11.2015.2928 . S2CID 133139689 .
- ^ Хиллер, Нортон; Мэннеринг, А.А.; Джонс, CM; Круикшанк, ARI (2005). «Природа Mauisaurus haasti Hector, 1874 (Reptilia: Plesiosauria)» . Журнал палеонтологии позвоночных . 25 (3): 588–601. doi : 10.1671/0272-4634(2005)025[0588:TNOMHH]2.0.CO;2 . S2CID 130607702 .
- ^ Хиллер, Нортон; О'Горман, Хосе П.; Отеро, Родриго А.; Мэннеринг, Эл А. (2017). «Переоценка позднемелового рода плезиозавров Уэдделла Mauisaurus Hector, 1874» . Новозеландский журнал геологии и геофизики . 60 (2): 112–128. Бибкод : 2017NZJGG..60..112H . дои : 10.1080/00288306.2017.1281317 . S2CID 132037930 .
- ^ Кнутсен, Эспен М.; Дракенмиллер, Патрик С.; Хурум, Йорн Х. (2012). «Новый вид плиозавра (Sauropterygia: Plesiosauria) из средней волги центрального Шпицбергена, Норвегия» (PDF) . Норвежский геологический журнал . 92 : 235–258. ISSN 1502-5322 . Архивировано из оригинала (PDF) 1 марта 2020 года.
- ^ Перейти обратно: а б Буши, М.-К.; Фрей, Э.; Стиннесбек, В.; Лопес-Олива, JG (2003). «Первое появление гигантского плиозавридного плезиозавра в поздней юре (кимеридже) Мексики» (PDF) . Бюллетень геологического общества Франции . 174 (3): 271–278. дои : 10.2113/174.3.271 . HDL : 2042/260 .
- ^ МакГенри, Колин Ричард (2009). Пожиратель богов: палеоэкология мелового плиозавра Kronosaurus queenslandicus (PDF) (доктор философии). стр. 1–460.
- ^ «Монстр фон Аррамберри» . Архивировано из оригинала 3 марта 2012 года . Проверено 10 сентября 2022 г.
- ^ «Чудовищная нижняя челюсть Камнора» . Архивировано из оригинала 11 октября 2011 года . Проверено 10 сентября 2022 г.
- ^ Бенджамин П. Кир (8 апреля 2003 г.). «Меловые морские рептилии Австралии: обзор систематики и распространения» . Меловые исследования . 24 (3): 277–303. Бибкод : 2003CrRes..24..277K . дои : 10.1016/S0195-6671(03)00046-6 .
- ^ МакГенри, Колин Р. 2009. Пожиратель богов: палеоэкология мелового плиозавра Kronosaurus queenslandicus . Университет Ньюкасла, Новый Южный Уэльс, Австралия, Интернет. [2]
- ^ «Самый крупный плиозаврид Северной Америки» . Архивировано из оригинала 1 августа 2016 года.
- ^ Джуди Массаре, Уильям Р. Уол, Мелисса Коннели, Майк Росс (2014). Палеоэкология морских рептилий сланцевой пачки Редуотер формации Сандэнс (юрский период) центрального Вайоминга, США. Геологический журнал 151(01):167–182 DOI:10.1017/S0016756813000472
- ^ Шумахер, бакалавр; Карпентер, К.; Эверхарт, MJ (2013). «Новый меловой плиозаврид (рептилия, плезиозаврия) из сланцев Карлайл (средний турон) округа Рассел, штат Канзас» . Журнал палеонтологии позвоночных . 33 (3): 613–628. Бибкод : 2013JVPal..33..613S . дои : 10.1080/02724634.2013.722576 . S2CID 130165209 .
- ^ Desojo 2013 , с. 20
- ^ Desojo 2013 , с. 22
- ^ «Редондазавр» . Музей естественной истории и науки Нью-Мексико . Архивировано из оригинала 20 декабря 2015 года . Проверено 15 октября 2022 г.
{{cite web}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Ханс-Дитер Сьюс (2019). Возникновение рептилий: 320 миллионов лет эволюции . Издательство Университета Джонса Хопкинса. п. 176. ИСБН 978-1-4214-2867-3 . Проверено 28 августа 2022 г.
- ^ Перейти обратно: а б Стерлинг Дж. Несбитт (2011). «Ранняя эволюция архозавров: взаимоотношения и происхождение основных клад» . Бюллетень Американского музея естественной истории . 352 : 1–292. дои : 10.1206/352.1 . hdl : 2246/6112 . S2CID 83493714 .
- ^ Перейти обратно: а б Дезохо 2013 , с. 260
- ^ Несбитт, С.Дж., Брусатте, С.Л., Дезохо, Дж.Б., Липарини, А., Франса, MAGD, Вайнбаум, Дж.К., и Гауэр, DJ (2013). Рауизуха. Геологическое общество, Лондон, Специальные публикации, 379 (1), 241–274. https://doi.org/10.1144/SP379.1
- ^ Desojo 2013 , с. 527
- ^ Майкл Дж. Бентон (2015). Палеонтология позвоночных . Уайли Блэквелл. п. 158. ИСБН 978-1-118-40755-4 . Проверено 15 сентября 2022 г.
- ^ Липарини, А.; Шульц, CL (2013). «Реконструкция мускулатуры бедра вымершего псевдозухия Prestosuchus chiniquensis из зоны объединения динодонтозавров (эпоха среднего триаса), последовательность Санта-Мария 1, южная Бразилия». Геологическое общество, Лондон, специальные публикации . 379 (1): 441–468. Бибкод : 2013GSLSP.379..441L . CiteSeerX 10.1.1.985.9550 . дои : 10.1144/SP379.20 . S2CID 130984792 .
- ^ Паркер, WG (2005). «Новый вид позднетриасового этозавра Desmatosuchus (Archosauria: Pseudosuria)». Comptes Рендус Палевол . 4 (4): 327–340. Бибкод : 2005CRPal...4..327P . дои : 10.1016/j.crpv.2005.03.002 .
- ^ Дезохо, Дж.Б.; Хекерт, AB; Марц, Дж.В.; Паркер, В.Г.; Шох, Р.Р.; Смолл, Би Джей; Сулей, Т. (2013). «Этозаврия: клада панцирных псевдозухий из континентальных отложений верхнего триаса» . Геологическое общество, Лондон, специальные публикации . 379 (1): 203–239. Бибкод : 2013GSLSP.379..203D . дои : 10.1144/SP379.17 . S2CID 129267515 .
- ^ фон Бачко, МБ, Десохо, Дж. Б., Гауэр, DJ, Риджли, Р., Бона, П. иВитмер, LM (2021). Новые цифровые эндокасты мозговой оболочки двух видов Desmatosuchus и нейрокраниальное разнообразие внутри Aetosauria (Archosauria: Pseudosuchia) . Анатомические записи, 1–20. https://doi.org/10.1002/ar.24798
- ^ Перейти обратно: а б Кейси М. Холлидей и Николас М. Гарднер (2012). «Новая евзуховая крокодилиформная форма с новым черепным покровом и ее значение для происхождения и эволюции Crocodylia» . ПЛОС ОДИН . 7 (1): e30471. Бибкод : 2012PLoSO...730471H . дои : 10.1371/journal.pone.0030471 . ПМЦ 3269432 . ПМИД 22303441 .
- ^ Бокантен, Жан; Мело, Джанира (2006). « Stupendemys souzai sp. nov (Pleurodira, Podocnemididae) из миоцена-плиоцена формации Солимойнс, Бразилия» . Бразильский журнал палеонтологии . 9 (2): 187–192. дои : 10.4072/rbp.2006.2.02 .
- ^ Перейти обратно: а б с д "Крупнейшие крокодиломорфы" . vk.com (in Russian) . Retrieved 6 September 2022 .
- ^ Сидаде, Джованни М.; Ринкон, Асканио Д.; Солорсано, Андрес (2020). «Новые краниальные и посткраниальные элементы Mourasuchus (Alligatoroidea: Caimaninae) из позднего миоцена Венесуэлы и их палеобиологические последствия» . Историческая биология . 33 (10): 2387–2399. дои : 10.1080/08912963.2020.1795844 . S2CID 225395230 .
- ^ Пайва, Ана Лаура С.; Годой, Педро Л.; Соуза, Рэй Б.Б.; Кляйн, Вильфрид; Сю, Энни С. (13 августа 2022 г.). «Оценка размеров тела особей Caimaninae из миоцена Южной Америки» . Журнал южноамериканских наук о Земле . 118 : 103970. Бибкод : 2022JSAES.11803970P . дои : 10.1016/j.jsames.2022.103970 . ISSN 0895-9811 . S2CID 251560425 .
- ^ Швиммер, Дэвид Р. (2002). «Размер дейнозуха ». Король крокодилов: Палеобиология дейнозуха . Издательство Университета Индианы . стр. 42–63. ISBN 978-0-253-34087-0 .
- ^ Лукас, Спенсер Г.; Салливан, Роберт М.; Шпильманн, Джастин А. (2006). «Гигантский крокодиловый дейнозух из верхнего мела бассейна Сан-Хуан, Нью-Мексико» (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико . 35 . Архивировано из оригинала (PDF) 17 июня 2009 года . Проверено 11 мая 2010 г.
- ^ Стаут, Джей Би (2020). «Новый аллигатор раннего плейстоцена (Eusuchia: Crocodylia) из Флориды устраняет пробел в эволюции аллигаторов » . Зоотакса . 4868 (1): 41–60. дои : 10.11646/zootaxa.4868.1.3 . ПМИД 33311408 . S2CID 226337860 .
- ^ Хед, Джей-Джей (2001). «Систематика и размер тела гигантского загадочного крокодилоида Rhamphosuchus crassidens и история фауны крокодилов группы Сивалик (миоцен)». Журнал палеонтологии позвоночных . 21 (Дополнение к № 3): 1–117. дои : 10.1080/02724634.2001.10010852 . S2CID 220414868 .
- ^ Перейти обратно: а б с д и Мартин, Джереми Э.; Антуан, Пьер-Оливье; Перье, Винсент; Добро пожаловать, Жан-Лу; Метаис, Грегуар; Мариво, Лоран (4 июля 2019 г.). «Большой крокодилоид из олигоцена холмов Бугти, Пакистан» (PDF) . Журнал палеонтологии позвоночных . 39 (4): e1671427. Бибкод : 2019JVPal..39E1427M . дои : 10.1080/02724634.2019.1671427 . ISSN 0272-4634 . S2CID 209439989 . Архивировано (PDF) из оригинала 9 сентября 2022 года.
- ^ Рифф, Д.; Агилера, ОА (2008). «Крупнейшие в мире гавиалы Gryposuchus: описание G. croizati n. sp. (Crocodylia, Gavialidae) из верхнемиоценовой формации Урумако, Венесуэла» . Палеонтологический журнал . 82 (2): 178–195. Бибкод : 2008PalZ...82..178R . дои : 10.1007/bf02988408 . S2CID 85172486 .
- ^ Сторрс, GW (2003). Позднемиоцен-раннеплиоценовая крокодиловая фауна Лотагама, юго-западная часть бассейна Туркана, Кения. В: Лотагам: Рассвет человечества в Восточной Африке, стр. 101-1. 137–159. Нью-Йорк. Издательство Колумбийского университета. ISBN 0-231-11870-8 .
- ^ Брочу, Калифорния; Сторрс, GW (2012). «Гигантский крокодил из плио-плейстоцена Кении, филогенетические связи неогеновых африканских крокодилов и древность Crocodylus в Африке» . Журнал палеонтологии позвоночных . 32 (3): 587. Бибкод : 2012JVPal..32..587B . дои : 10.1080/02724634.2012.652324 . S2CID 85103427 .
- ^ « Крокодил антропофаг » . Палеокритти.com . Архивировано из оригинала 14 июня 2016 года.
- ^ Юэн Каллауэй (24 февраля 2010 г.). «Чудовищный крокодил был кошмаром древнего человека» . Новый учёный . Архивировано из оригинала 7 декабря 2021 года.
- ^ Дельфино, Массимо; Де Вос, Джон (март 2014 г.). «Гигантский крокодил в коллекции Дюбуа из плейстоцена Кали-Геде (Ява)» . Интегративная зоология . 9 (2): 141–147. дои : 10.1111/1749-4877.12065 . hdl : 2318/141647 . ПМИД 24673759 .
- ^ «Блин! Огромный доисторический крокодил появился на юго-востоке Квинсленда» . ScienceDaily . Проверено 9 января 2021 г.
- ^ Роу, С.; Филд, Дж. Х.; Арчер, М.; Грейсон, Дания; Цена, ГДж; Луис, Дж.; Вера, Т.; Уэбб, GE; Дэвидсон, И.; Муни, SD (2013). «Изменение климата лежит в основе дебатов по поводу исчезновения мегафауны в Сахуле (плейстоценовая Австралия-Новая Гвинея)» . Труды Национальной академии наук . 110 (22): 8777–8781. Бибкод : 2013PNAS..110.8777W . дои : 10.1073/pnas.1302698110 . ПМК 3670326 . ПМИД 23650401 .
- ^ Роу, Стивен (2002). «Обзор экологии наземных млекопитающих и рептилий-хищников в ископаемых фаунах Австралии, а также факторов, влияющих на их разнообразие: миф о доминировании рептилий и его более широкие последствия». Австралийский журнал зоологии . 50 (1): 1. дои : 10.1071/zo01053 . ISSN 0004-959X .
- ^ Фланнери, ТФ (1990). «Утрата фауны плейстоцена: последствия афтершока для прошлого и будущего Австралии». Археология в Океании . 25 (2): 45–55. дои : 10.1002/j.1834-4453.1990.tb00232.x .
- ^ Соббе, Ян Х.; Прайс, Гилберт Дж.; Кнезур, Роберт А. (2013). «Зифодонтный крокодил из водосбора Кинг-Крик позднего плейстоцена, Дарлинг-Даунс, Квинсленд» . Мемуары Квинслендского музея . 52 (2): 601–606.
- ^ Сторрс, Г.В.; Ефимов, М.Б. (2000). «Мезозойские крокодилоформы северо-центральной Евразии». У Майкла Дж. Бентона; Михаил А. Шишкин; Дэвид М. Анвин; Евгений Н. Курочкин (ред.). Эпоха динозавров в России и Монголии . Издательство Кембриджского университета. стр. 402–419.
- ^ Лион, Габриэль (9 декабря 2001 г.). «Информационный бюллетень» . СуперКрок . Разведка проекта . Проверено 22 сентября 2007 г.
- ^ Хейли Д. О'Брайен, Лейга М. Линч; Кент А. Влит; Брюгген, Джон; Грегори М. Эриксон; Поль М. Жиньяк (2019). «Аллометрия ширины головы крокодилов и филогенетическое предсказание размера тела у вымерших крокодилообразных» . Интегративная организменная биология . 1 (1): obz006. дои : 10.1093/iob/obz006 . ПМЦ 7671145 . ПМИД 33791523 .
- ^ Мартин, Дж. Э.; Лаупрасерт, К.; Бюффето, Э.; Лиард Р. и Сютитхорн В. (2013). «Крупный фолидозавр в формации Фу Крадунг на северо-востоке Таиланда» . Палеонтология . 57 (4): 757–769. дои : 10.1111/пала.12086 . S2CID 128482290 .
- ^ Баффето, Э. (1978). «Дирозавриды (Crocodylia, Mesosuchia) из нижнеэоценовых фосфатов Туниса: Dyrosaurus , Rhabdognathus , Phosphatosaurus ». Средиземноморская геология . 5 (2): 237–256. дои : 10.3406/geolm.1978.1046 .
- ^ Нэйш, Д. 2002. Объяснение окаменелостей 34: Крокодилы . Геология сегодня 2 : 71–77. Архивная копия от 24 января 2019 г.
- ^ Перейти обратно: а б Мольнар Р.Э., де Васконселлос FM (2016). «Кайнозойские динозавры в Южной Америке – еще раз» . Мемуары музея Виктории . 74 : 363–377. дои : 10.24199/j.mmv.2016.74.25 .
- ^ Перейти обратно: а б Мартин, Джереми Э.; Поша-Котийу, Йохан; Лоран, Ив; Перье, Винсент; Роберт, Эммануэль; Антуан, Пьер-Оливье (28 октября 2022 г.). «Анатомия и филогения исключительно крупного себецида (Crocodylomorpha) из среднего эоцена южной Франции» . Журнал палеонтологии позвоночных . 42 (4). Бибкод : 2022JVPal..42E3828M . дои : 10.1080/02724634.2023.2193828 . ISSN 0272-4634 . S2CID 258361595 .
- ^ Рифф, Д.; Келлнер, AWA (2011). «Крокодилформы Baurusuchid как имитаторы теропод: подсказки по черепу и аппендикулярной морфологии Stratiotosuchus maxhechti (верхний мел Бразилии)» . Зоологический журнал Линнеевского общества . 163 (с1): с37–с56. дои : 10.1111/j.1096-3642.2011.00713.x .
- ^ От Сассо, К.; Пасини, Г.; Флери, Г.; Магануко, С. (2017). « Razanandrongobe sakalavae , гигантский мезоэвкрокодил из средней юры Мадагаскара, является старейшим из известных нотозухов » ПерДж . 5 : е3 дои : 10.7717/peerj.3481 . ПМК 5499610 . ПМИД 28690926 .
- ^ «У гигантского крокодила были зубы, как у тиранозавра» . Новости Би-би-си . Би-би-си . 4 июля 2017 года . Проверено 22 апреля 2020 г.
- ^ С любезного разрешения, Дирли; Ларссон, Ханс CE; Максвелл, Эрин Э.; Парра Руге, Мэри Луз; Патарройо, Педро; Уилсон, Джеффри А. (6 октября 2019 г.). «Раннемеловой телеозавроид (Crocodylomorpha: Thalattosuria) из Колумбии». Амегиниана . 56 (5): 365. doi : 10.5710/amgh.26.09.2019.3269 . ISSN 0002-7014 . S2CID 210110716 .
- ^ Перейти обратно: а б Янг, Монтана; Раби, М.; Белл, Массачусетс; Фоффа, Д.; Сталь, Л.; Сакс, С.; Пейер, К. (2016). «Большеголовые морские крокодилоформы и почему мы должны быть осторожны при использовании существующих видов в качестве показателя длины тела для давно вымерших родственников» . Электронная палеонтология . 19 (3): 1–14. дои : 10.26879/648 .
- ^ Джонсон, Микела М.; Янг, Марк Т.; Брусатте, Стивен Л. (2020). «Филогенетика Teleosauroidea (Crocodylomorpha, Thalattosuria) и значение для их экологии и эволюции» . ПерДж . 8 : е9808. дои : 10.7717/peerj.9808 . ПМЦ 7548081 . ПМИД 33083104 .
- ^ Янг, Монтана; Брусатте, СЛ; Де Андраде, МБ; Дезохо, Дж.Б.; Битти, БЛ; Сталь, Л.; Фернандес, MS; Сакамото, М.; Руис-Оменьака, Дж.И.; Шох, Р.Р. (2012). Батлер, Ричард Дж (ред.). «Краниальная остеология и экология питания Metriorhynchid Crocodylomorph родов Dakosaurus и Plesiosuchus из поздней юры Европы» . ПЛОС ОДИН . 7 (9): e44985. Бибкод : 2012PLoSO...744985Y . дои : 10.1371/journal.pone.0044985 . ПМЦ 3445579 . ПМИД 23028723 .
- ^ Янг, Монтана; Де Андраде, МБ; Брусатте, СЛ; Сакамото, М.; Листон, Дж. (2013). «Самый старый известный суперхищник метриоринхид: новый род и вид из средней юры Англии, имеющий значение для зубчатости и эволюции нижней челюсти в хищных кладах» . Журнал систематической палеонтологии . 11 (4): 475–513. Бибкод : 2013JSPal..11..475Y . дои : 10.1080/14772019.2012.704948 . S2CID 85276836 .
- ^ Янг, Марк Т.; Брусатте, Стивен Л.; Де Андраде, Марко Брандалис; Дезохо, Джулия Б.; Битти, Брайан Л.; Сталь, Лорна; Фернандес, Марта С.; Сакамото, Манабу; Руис-Оменьака, Хосе Игнасио; Шох, Райнер Р.; (2012) « Краниальная остеология и экология питания Metriorhynchid Crocodylomorph родов Dakosaurus и Plesiosuchus из поздней юры Европы », в Батлере, Ричарде Дж. (ред.), PLoS ONE, vol. 7, нет. 9, с. e44985, pmid 23028723, pmc 3445579, doi:10.1371/journal.pone.0044985
- ^ Перейти обратно: а б Линдси Э. Занно, Сьюзан Дримала, Стерлинг Дж. Несбитт, Винсент П. Шнайдер (2015) Ранние крокодиломорфы увеличивают разнообразие хищников высшего уровня во время появления динозавров . Научные отчеты, том 5, номер статьи: 9276
- ^ Несбитт, Стерлинг Дж.; Ирмис, Рэндалл Б.; Лукас, Спенсер Г.; Хант, Адриан П. (2005). «Гигантский крокодиломорф из верхнего триаса Нью-Мексико». Палеонтологическая газета . 79 (4): 471–478. Бибкод : 2005PalZ...79..471N . дои : 10.1007/bf02988373 . S2CID 128541365 .
- ^ Крокодиломорфа. Крокодилы и их сородичи , архивная копия от 20 июня 2022 года.
- ^ Занно, Линдси Э.; Дримала, Сьюзен; Несбитт, Стерлинг Дж.; Шнайдер, Винсент П. (19 марта 2015 г.). «Ранние крокодиломорфы увеличивают разнообразие хищников высшего уровня во время появления динозавров» . Научные отчеты . 5 : 9276. Бибкод : 2015NatSR...5E9276Z . дои : 10.1038/srep09276 . ISSN 2045-2322 . ПМЦ 4365386 . ПМИД 25787306 .
- ^ Перейти обратно: а б Уиттон, член парламента; Нэйш, Д. (2008). Макклейн, Крейг Р. (ред.). «Переоценка функциональной морфологии и палеоэкологии аждархид-птерозавров» . ПЛОС ОДИН . 3 (5): е2271. Бибкод : 2008PLoSO...3.2271W . дои : 10.1371/journal.pone.0002271 . ПМК 2386974 . ПМИД 18509539 .
- ^ Бюффето, Э.; Григореску, Д.; Чики, З. (2002). «Новый гигантский птерозавр с крепким черепом из позднего мелового периода Румынии» (PDF) . Naturwissenschaften . 89 (4): 180–184. Бибкод : 2002NW.....89..180B . дои : 10.1007/s00114-002-0307-1 . ПМИД 12061403 . S2CID 15423666 .
- ^ Перейти обратно: а б Пол, Грегори С. (2022). Принстонский полевой справочник по птерозаврам . Издательство Принстонского университета. стр. 155–172. дои : 10.1515/9780691232218 . ISBN 978-0-691-23221-8 . S2CID 249332375 .
- ^ Перейти обратно: а б Таканобу Цуихидзи; Брайан Андрес; Патрик М. О'Коннор; Махито Ватабе; Хишигджав Цогтбаатар; Майнбаяр Буувей (2017). «Гигантские останки птерозавров из верхнего мела Монголии» . Журнал палеонтологии позвоночных . 37 (5): e1361431. Бибкод : 2017JVPal..37E1431T . дои : 10.1080/02724634.2017.1361431 . S2CID 134424023 .
- ^ «Древний крылатый ужас был одним из крупнейших летающих животных» . 31 октября 2017 г. Архивировано из оригинала 30 марта 2019 г.
- ^ Келлнер, AWA; КАМПОС, ДА; Саяо, Дж. М.; САРАИВА, АНАФ; РОДРИГЕС, Т.; ОЛИВЕЙРА, Г.; КРУЗ, Луизиана; КОСТА, Франция; СИЛЬВА, HP; Феррейра, Дж.С. (2013). «Самый крупный летающий повтор из Гондваны: новый экземпляр Tropeognathus cf. T. Mesembrinus Wellnhofer, 1987 (Pterodactyloidea, Anhangueridae) и других крупных птерозов из формации Ромуальдо, ранний мел, Бразилия» . Анналы Бразильской академии наук . 85 (1): 113–135. дои : 10.1590/S0001-37652013000100009 . ПМИД 23538956 .
- ^ Родригес, Т.; Келлнер, А. (2013). «Таксономический обзор комплекса Ornithocheirus (Pterosauria) мела Англии» . ZooKeys (308): 1–112. Бибкод : 2013ZooK..308....1R . дои : 10.3897/zookeys.308.5559 . ПМЦ 3689139 . ПМИД 23794925 .
- ^ «Ископаемое крупнейшего юрского птерозавра, найденное на острове Скай» . Новости Би-би-си . 22 февраля 2022 г. Проверено 22 февраля 2022 г.
- ^ Наталья Ягельская; Майкл О'Салливан; Грегори Ф. Фанстон; и др. (февраль 2022 г.). «Скелет из средней юры Шотландии проливает свет на более раннее происхождение крупных птерозавров». Современная биология . 32 : 1–8. дои : 10.1016/J.CUB.2022.01.073 . ISSN 0960-9822 . Викиданные Q110984418 .
- ^ Перейти обратно: а б Шпиндлер, Фредерик; Ифрим, Кристина (2021). «След следа – возможного первого птерозавра из Эттлинга. След следа – предполагаемого первого птерозавра из местонахождения Эттлинга» . Археоптерикс . 37 :75–83.
- ^ Перейти обратно: а б Андрес, Б.; Кларк, Дж. М.; Син, X. (2010). «Новый рамфоринхидный птерозавр из верхней юры Синьцзяна, Китай, и филогенетические отношения базальных птерозавров» (PDF) . Журнал палеонтологии позвоночных . 30 (1): 163–187. Бибкод : 2010JVPal..30..163A . дои : 10.1080/02724630903409220 . S2CID 53688256 . Архивировано из оригинала (PDF) 31 июля 2021 года.
- ^ Перейти обратно: а б Браунштейн, CD (2022). «Высокое морфологическое разнообразие в причудливой палеоценовой фауне хищных пресноводных рептилий» . БМК Эколь Эвол . 22 (34): 34. дои : 10.1186/s12862-022-01985-z . ПМЦ 8935759 . ПМИД 35313822 .
- ^ Мацумото, Р.; Эванс, SE (10 декабря 2010 г.). «Хористодеры и пресноводные комплексы Лавразии» . Журнал иберийской геологии . 36 (2): 253–274. Бибкод : 2010JIbG...36..253M . дои : 10.5209/rev_JIGE.2010.v36.n2.11 . ISSN 1886-7995 . Архивировано из оригинала 18 августа 2020 года.
- ^ Тиана М. Де-Оливейра, Фелипе Л. Пиньейру, Атила Аугусто Сток Да-Роза, Сержио Диас-Да-Сильва, Леонардо Кербер. 2018. « Новый архозавроморф из Южной Америки дает представление о ранней диверсификации танистрофеидов ». PLOS ONE 15(5): e0233216. doi.org/10.1371/journal.pone.0230890
- ^ Чжао, Ли-Цзюнь; Сато, Тамаки; Лю, Цзюнь; Ли, Чун; Ву, Сяо-Чун (2010). «Новый скелет Miodentosaurus brevis (Diapsida: Thallatosauria)» (PDF ) позвоночные Палазиатские 48 (1): 1–10.
- ^ Лю, Дж.; Чжао, LJ; Ли, К.; Он, Т. (2013). «Остеология Concavispina biseridens (Reptilia, Thalattosauria) из формации Сяова (Карний), Гуанлин, Гуйчжоу, Китай» . Журнал палеонтологии . 87 (2): 341–350. Бибкод : 2013JPal...87..341L . дои : 10.1666/12-059R1.1 . S2CID 83684967 .
- ^ Де ла Саль П., Р. Ломакс Д., А. Массаре Дж., Галлуа Р. (2018). «Гигантский ихтиозавр позднего триаса из Великобритании и новая интерпретация «динозавровых» костей из Ост-Клифф» . ПЛОС ОДИН . 13 (4). doi : 10.6084/m9.figshare.5975440 . Архивировано из оригинала 27 февраля 2022 года.
- ^ Пикрелл, Джон (2018). «Доисторическое «морское чудовище», возможно, самое большое из когда-либо живших» . Архивировано из оригинала 31 мая 2022 года.
- ^ Ломакс, Дин Р.; Салле, Поль де ла; Перилло, Марчелло; Рейнольдс, Джастин; Рейнольдс, Руби; Уолдрон, Джеймс Ф. (17 апреля 2024 г.). «Последние гиганты: новые доказательства существования гигантских ихтиозавров позднего триаса (ретийского периода) из Великобритании» . ПЛОС ОДИН . 19 (4): e0300289. Бибкод : 2024PLoSO..1900289L . дои : 10.1371/journal.pone.0300289 . ISSN 1932-6203 . ПМЦ 11023487 . ПМИД 38630678 .
- ^ «Триасовый гигант» . Архивировано из оригинала 16 июля 2011 года . Проверено 13 октября 2022 г.
- ^ Перейти обратно: а б Сандер, премьер-министр; Ромеро Перес де Вильяр, П.; Фуррер, Х.; Винтрич, Т. (2022). «Гигантские ихтиозавры позднего триаса из формации Кёссен в Швейцарских Альпах и их палеобиологические последствия» (PDF) . Журнал палеонтологии позвоночных . 41 (6): e2046017. дои : 10.1080/02724634.2021.2046017 . S2CID 248444094 . Архивировано (PDF) из оригинала 20 сентября 2022 года.
- ^ Перейти обратно: а б с Сандер, премьер-министр; Грибелер, Э.М.; Кляйн, Н.; Хуарбе, СП; Винтрич, Т.; Ревелл, LJ; Шмитц, Л. (2021). «Ранние гиганты демонстрируют более быструю эволюцию крупных размеров тела у ихтиозавров, чем у китообразных» (PDF) . Наука . 374 (6575): eabf5787. дои : 10.1126/science.abf5787 . ПМИД 34941418 . S2CID 245444783 . Архивировано (PDF) из оригинала 10 октября 2022 года.
- ^ Перейти обратно: а б «Исследователи нашли челюстную кость возрастом 205 миллионов лет одного из крупнейших животных, когда-либо живших на Земле» . Newsweek . 9 апреля 2018 г. Архивировано из оригинала 21 сентября 2022 г.
- ^ Пи Джей Карри. « Hovasaurus boulei, водный эозух из верхней перми Мадагаскара ». Палеонт. афр., 24 (1981), с. 112.
- ^ Перейти обратно: а б с Марко Романо; Фабио Мануччи; Брюс Рубидж; Марк Дж. Ван ден Брандт (17 июня 2021 г.). «Объемная оценка массы тела и реконструкция in vivo российского парейазавра Scutosaurus karpinski » . Границы экологии и эволюции . 9 . дои : 10.3389/fevo.2021.692035 . hdl : 11573/1634310 .
- ^ МакГи, Джордж Р. младший (7 августа 2018 г.). Каменноугольные гиганты и массовое вымирание: мир позднепалеозойского ледникового периода . Издательство Колумбийского университета. ISBN 978-0-231-54338-5 . Проверено 16 октября 2019 г. - через Google Книги.
- ^ Пинейро, Грасиела; Нуньес Демарко, Пол; Менегель, Мелитта Д. (17 мая 2016 г.). «Онтогенетическая трансформация лапки мезозаврид: вклад в происхождение примитивного амниотического астрагала» . ПерДж . 4 : е2036. дои : 10.7717/peerj.2036 . ПМЦ 4878385 . ПМИД 27231658 .
- ^ Додик, Дж. Т. и Модесто, С. П., 1995. Черепная анатомия рептилии-капторинида Labidosaurikos meachami из нижней перми Оклахомы. Палеонтология , 38(3), с.687.
- ^ Патрисия Викерс Рич ; Томас Хьюитт Рич; Милдред Адамс Фентон ; Кэрролл Лейн (15 января 2020 г.). Книга окаменелостей: летопись доисторической жизни . Дуврские публикации. п. 444. ИСБН 978-0-486-83855-7 . Проверено 13 октября 2022 г.
- ^ Лу Дж., Ли Т., Чжун С., Адзума Ю., Фудзита М., Донг З. и Джи К. (2007). « Новый динозавр-юннанозаврид (Dinosauria, Prosauropoda) из среднеюрской формации Чжанхэ в Юаньмоу, провинция Юньнань, Китай ». Мемуары Музея динозавров префектуры Фукуи, 6: 1–15.
- ^ Перейти обратно: а б с Кеннет Карпентер (2006). «Самый большой из больших: критическая переоценка мегазауропода Amphicoelias fragillimus » . В книге «Палеонтология и геология верхнеюрской формации Моррисон». Бюллетень Музея естественной истории и науки Нью-Мексико . 36 : 131–138.
- ^ Перейти обратно: а б Пол, Грегори С. (2019). «Определение самого крупного известного наземного животного: критическое сравнение различных методов восстановления объема и массы вымерших животных» (PDF) . Анналы музея Карнеги . 85 (4): 335–358. дои : 10.2992/007.085.0403 . S2CID 210840060 . Архивировано (PDF) из оригинала 29 сентября 2022 года.
- ^ Гальтон, Питер М.; Айясами, Кришнан (1 июля 2017 г.). «Предполагаемая новейшая кость пластинчатого динозавра (Ornithischia: Stegosauria), «кожная пластина» из маастрихта (верхний мел) на юге Индии» . Новогодний ежегодник по геологии и палеонтологии-трактаты . 285 (1): 91–96. дои : 10.1127/njgpa/2017/0671 . ISSN 0077-7749 .
- ^ Перейти обратно: а б Молина-Перес и Ларраменди (2020). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы . Нью-Джерси: Издательство Принстонского университета. п. 46. ИСБН 978-0-691-19069-3 .
- ^ Пол, Грегори С.; Ларраменди, Азиер (11 апреля 2023 г.). «Оценка массы тела брухаткайозавра и других фрагментарных останков зауроподов позволяет предположить, что крупнейшие наземные животные были примерно такими же большими, как и величайшие киты». Летайя . 56 (2): 1–11. Бибкод : 2023Лета..56..2.5П . дои : 10.18261/let.56.2.5 . ISSN 0024-1164 . S2CID 259782734 .
- ^ Майк Тейлор (2019). «Суперзавр, Ультразавр и Дистилозавр в 2019 году, часть 2: что мы нашли в Юте» . svpow.com . Архивировано из оригинала 21 сентября 2022 года.
- ^ Майк Тейлор (16 сентября 2016 г.). «Насколько ужасающей была шея Барозавра?» . svpow.com . Архивировано из оригинала 22 сентября 2022 года.
- ^ Перейти обратно: а б Молина-Перес и Ларраменди (2020). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы . Нью-Джерси: Издательство Принстонского университета. п. 36. ISBN 9780691190693 .
- ^ Перейти обратно: а б Кертис, Брайан (2021). «Новые осевые элементы карьера динозавров Сухой Мезы Supersaurus vivianae (Jensen 1985) дают дополнительное представление о его филогенетических взаимоотношениях и размерах, что позволяет предположить, что длина животного превышала 39 метров» (PDF) . п. 92. Архивировано (PDF) из оригинала 8 октября 2022 года.
- ^ Майк Тейлор (16 июня 2019 г.). «Размеры животного BYU 9024» . svpow.com . Архивировано из оригинала 16 апреля 2022 года.
- ^ Молина-Перес и Ларраменди (2020). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы . Нью-Джерси: Издательство Принстонского университета. п. 156. ИСБН 978-0-691-19069-3 .
- ^ Питер Мэтьюз (1992). Книга рекордов Гиннесса . Издательство Гиннесс. п. 42. ИСБН 9780851129785 .
- ^ Тим Футман, Марк К. Янг (май 2001 г.). Книга рекордов Гиннесса 2001 года . Бантамские книги. п. 276. ИСБН 978-0-553-58375-5 .
- ^ Молина-Перес и Ларраменди (2020). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы . Нью-Джерси: Издательство Принстонского университета. п. 32. ISBN 9780691202976 .
- ^ Ведель, Мэтью Дж.; Чифелли, РЛ; Сандерс, Р.К. (2000). «Остеология, палеобиология и взаимоотношения динозавра-зауропода Sauroposeidon » (PDF) . Acta Palaeontologica Polonica . 45 : 343–388. S2CID 59141243 . Архивировано из оригинала (PDF) 26 июня 2020 года.
- ^ Книги рекордов Гиннесса . Бантамские книги. 2004. с. 110. ИСБН 9780553587128 .
- ^ Перейти обратно: а б Молина-Перес, Рубен; Ларраменди, Азиер (29 сентября 2020 г.). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы . Издательство Принстонского университета. п. 55. ИСБН 978-0-691-19069-3 .
- ^ Пол, GS (1988). «Брахиозавры-гиганты Моррисона и Тендагуру с описанием нового подрода Giraffatitan и сравнением крупнейших в мире динозавров» (PDF) . Хантерия . 2 (3): 1–14. Архивировано (PDF) из оригинала 27 июня 2022 года.
- ^ Перейти обратно: а б Пол, GS, 2010, Принстонский полевой справочник по динозаврам, Princeton University Press.
- ^ «Гигантский динозавр похудел... немного» . Новости Би-би-си . 10 августа 2017 года. Архивировано из оригинала 21 сентября 2022 года . Проверено 17 октября 2022 г.
- ^ Перейти обратно: а б Отеро, Алехандро; Карбаллидо, Хосе Л.; Перес Морено, Агустин (2020). «Аппендикулярная остеология Patagotitan majorum (Dinosauria, Sauropoda)» . Журнал палеонтологии позвоночных . 40 (4): e1793158. Бибкод : 2020JVPal..40E3158O . дои : 10.1080/02724634.2020.1793158 . S2CID 225012747 .
- ^ «Не верьте шумихе: Патаготитан был не больше аргентинозавра» . Изображение недели зауропода Vertebra . 9 августа 2017 года . Проверено 18 ноября 2018 г.
- ^ Лаковара, Кеннет Дж.; Ламанна, MC; Ибирику, LM; Пул, Джей Си; Шретер, ER; Ульманн, П.В.; Фогеле, КК; Болес, З.М.; Картер, AM; Фаулер, ЕК; Эгертон, ВМ; Мойер, А.Е.; Куэнур, CL; Шейн, JP; Харрис, доктор юридических наук; Мартинес, РД; Новас, FE (4 сентября 2014 г.). «Гигантский, исключительно полный динозавр-титанозавр-зауропод из Южной Патагонии, Аргентина» . Научные отчеты . 4 (1): 6196. Бибкод : 2014NatSR...4E6196L . дои : 10.1038/srep06196 . ПМЦ 5385829 . ПМИД 25186586 .
- ^ Рафаэль Ройо-Торрес; Альберто Кобос; Луис Алькала (2006). «Гигантский европейский динозавр и новая группа зауроподов» (PDF) . Наука . 314 (5807): 1925–1927. Бибкод : 2006Sci...314.1925R . дои : 10.1126/science.1132885 . ПМИД 17185599 . S2CID 9343711 .
- ^ Перейти обратно: а б с д и ж г Хольц, Томас Р. младший (2012). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов (PDF) . Архивировано (PDF) из оригинала 2 октября 2022 года.
Зима 2011 г. Приложение.
- ^ Молина-Перес, Рубен; Ларраменди, Азиер (29 сентября 2020 г.). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы . Издательство Принстонского университета. п. 28. ISBN 978-0-691-19069-3 .
- ^ «Документ, описывающий NHMUK PV R5937, брахиозаврид Тендагуру, известный как «Архиепископ» » . Гитхаб . 25 августа 2022 г.
- ^ де Лаппарент, AF (1960). интеркалара» центральной Сахары » континентального (PDF) . Mémoires de la Société Géologique de France . «Динозавры « (на французском языке )
- ^ Мэннион, Полиция; Апчерч, Пол; Барнс, Рози Н.; Матеус, Октавио (2013). «Остеология позднеюрского португальского динозавра-зауропода Lusotitan atalaiensis (Macronaria) и история эволюции базальных титанозавроформ» (PDF) . Зоологический журнал Линнеевского общества . 168 : 98–206. дои : 10.1111/zoj.12029 .
- ^ Нима. «Является ли французское чудовище эвгелоподидом?» . paleoking.blogspot.com .
- ^ Томпсон, Хелен. «Знакомьтесь: могучий спинозавр, первый динозавр, приспособленный к плаванию» . Смитсоновский институт . Проверено 22 ноября 2017 г. .
- ^ Перейти обратно: а б Терриен, Ф.; Хендерсон, DM (2007). «Мой теропод больше вашего… или нет: оцениваем размер тела теропод по длине черепа». Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . ISSN 0272-4634 . S2CID 86025320 .
- ^ даль Сассо, К.; Магануко, С.; Бюффето, Э.; Мендес, Массачусетс (2005). «Новая информация о черепе загадочного теропода спинозавра с замечаниями о его размерах и сходстве». Журнал палеонтологии позвоночных . 25 (4): 888–896. doi : 10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2 . S2CID 85702490 .
- ^ Ибрагим, Низар ; Серено, Пол С .; Даль Сассо, Криштиану; Магануко, Симоне; Фабри, Маттео; Мартилл, Дэвид М.; Зухри, Самир; Мирволд, Натан; Лурино, Дэвид А. (2014). «Полуводные адаптации гигантского хищного динозавра» . Наука . 345 (6204): 1613–6. Бибкод : 2014Sci...345.1613I . дои : 10.1126/science.1258750 . ПМИД 25213375 . S2CID 34421257 . Дополнительная информация
- ^ Перейти обратно: а б Хендерсон, DM (2018). «Вызов плавучести, баланса и устойчивости гипотезе полуводного спинозавра Стромера, 1915 (Dinosauria: Theropoda)» . ПерДж . 6 : е5409. дои : 10.7717/peerj.5409 . ПМК 6098948 . ПМИД 30128195 .
- ^ Перейти обратно: а б Молина-Перес и Ларраменди 2016. Записи и курьезы динозавров, тероподов и других динозавроморфов , Ларусс. Барселона, Испания с. 259
- ^ Нэйш, Даррен (9 июня 2022 г.). «Гигантский динозавр-спинозавр из мелового периода острова Уайт» . Зоология четвероногих . Проверено 29 августа 2022 г.
- ^ Кальво, Хорхе О.; Кориа, Родольфо (1998). «Новый экземпляр Giganotosaurus Carolinii» (PDF) . ГЕЯ : 117–122. Архивировано из оригинала (PDF) 16 февраля 2008 года.
- ^ Перейти обратно: а б Хендерсон Д.М. (1 января 1999 г.). «Оценка массы и центров масс вымерших животных методом трехмерного математического разрезания» . Палеобиология . 25 (1): 88–106.
- ^ Кориа, РА; Карри, Пи Джей (2002). «Мозговая черепная коробка Giganotosaurus carolinii (Dinosauria: Theropoda) из верхнего мела Аргентины». Журнал палеонтологии позвоночных . 22 (4): 802–811. doi : 10.1671/0272-4634(2002)022[0802:TBOGCD]2.0.CO;2 . S2CID 85678725 .
- ^ Кориа, РА; Сальгадо, Л. (1995). «Новый гигантский плотоядный динозавр из мелового периода Патагонии». Природа . 377 (6546): 225–226. Бибкод : 1995Natur.377..224C . дои : 10.1038/377224a0 . S2CID 30701725 .
- ^ Кальво, Джо; Кориа, Р.А. (1998). «Новый экземпляр Giganotosaurus carolinii (CORIA & SALGADO, 1995) подтверждает, что это самый крупный теропод из когда-либо найденных» (PDF) . Гайя . 15 : 117–122. Архивировано из оригинала (PDF) 16 февраля 2008 года.
- ^ Андерсон, Дж. Ф.; Холл-Мартин, AJ; Рассел, Д. (1985). «Окружность и вес длинных костей у млекопитающих, птиц и динозавров». Журнал зоологии . 207 (1): 53–61. дои : 10.1111/j.1469-7998.1985.tb04915.x .
- ^ Баккер, RT (1986). Ересь динозавров . Нью-Йорк: Издательство Кенсингтон. п. 241 . ISBN 978-0-688-04287-5 . OCLC 13699558 .
- ^ Саймон, диджей (2014). «Яйца гигантского динозавра (теропода) Oogenus Macroelongatoolithus (Elongatoolithidae) из юго-восточного Айдахо: таксономические, палеобиогеографические и репродуктивные последствия» (PDF) . Докторская диссертация, Государственный университет Монтаны . Бозман. Архивировано (PDF) из оригинала 11 августа 2021 года.
- ^ Перейти обратно: а б г-н, Х.; Зеленицкий, ДК; Лю, Дж.; Карри, Пи Джей; Карпентер, К.; Сюй, Л.; Коппельхус, Э.Б.; Цзя, С.; Сяо, Л.; Чуанг, Х.; Ли, Т.; Кундрат, М.; Шен, К. (2017). «Перинат и яйца гигантского ценагнатидного динозавра из позднего мела центрального Китая» . Природные коммуникации . 8 : 14952. Бибкод : 2017NatCo...814952P . дои : 10.1038/ncomms14952 . ПМЦ 5477524 . ПМИД 28486442 .
- ^ Викариус, М.К., Марьянска, Т. и Вейшампель, Д.Б. 2004. Анкилозаврия. В: Вейшампель Д.Б., Додсон П. и Осмольска Х. (ред.). Динозаврия (2-е издание). Беркли: Издательство Калифорнийского университета. Стр. 363–392.
- ^ Кумбс, Уолтер П. (декабрь 1978 г.). «Теоретические аспекты беговой адаптации динозавров». Ежеквартальный обзор биологии . 53 (4): 393–418. дои : 10.1086/410790 . S2CID 84505681 .
- ^ Пол, Грегори С. (2010). Принстонский полевой справочник по динозаврам . Принстон, Нью-Джерси: Издательство Принстонского университета. п. 244 . ISBN 978-0-691-13720-9 .
- ^ Хольц, Томас Р. младший (2008). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов (PDF) . Нью-Йорк: Рэндом Хаус. стр. обновленное приложение . ISBN 978-0-375-82419-7 .
- ^ Лонгрич, Северная Каролина (2011). « Титаноцератопс уранос , гигантский рогатый динозавр из позднего кампана Нью-Мексико». Меловые исследования . 32 (3): 264–276. Бибкод : 2011CrRes..32..264L . дои : 10.1016/j.cretres.2010.12.007 .
- ^ Перейти обратно: а б Чжао, X.; Ли, Д.; Вешать.; Хао, Х.; Лю, Ф.; Ли, Л.; Фанг, X. (2007). « Zzhuchengosaurus maximus из провинции Шаньдун». Acta Geoscientia Sinica . 28 (2): 111–122. дои : 10.1007/s10114-005-0808-x . S2CID 119700784 .
- ^ Моррис, Уильям Дж. (1981). «Новый вид гадрозавровых динозавров из верхнего мела Нижней Калифорнии: ? Lambeosaurus laticaudus ». Журнал палеонтологии . 55 (2): 453–462.
- ^ Прието-Маркес, А.; Кьяппе, LM; Джоши, SH (2012). Додсон, Питер (ред.). «Ламбеозавровый динозавр Magnapaulia laticaudus из позднего мела Нижней Калифорнии, северо-запад Мексики» . ПЛОС ОДИН . 7 (6): e38207. Бибкод : 2012PLoSO...738207P . дои : 10.1371/journal.pone.0038207 . ПМЦ 3373519 . ПМИД 22719869 .
- ^ Моррис, Уильям Дж. (1981). «Новый вид гадрозавровых динозавров из верхнего мела Нижней Калифорнии: ? Lambeosaurus laticaudus ». Журнал палеонтологии . 55 (2): 453–462.
- ^ Перейти обратно: а б Пол, Грег (2010). «Принстонский полевой справочник по динозаврам». Нью-Джерси: Издательство Принстонского университета. п. 335.
- ^ Восик, М.; Эванс, округ Колумбия (2022 г.). «Остеогистологическая и тафономическая оценка истории жизни Edmontosaurus annectens (Ornithischia: Hadrosauridae) из позднемелового (маастрихтского) карьера динозавров Рут Мейсон, Южная Дакота, США, с учетом онтогенетической сегрегации между молодыми и взрослыми гадрозавридами» . Журнал анатомии . 241 (2): 272–296. дои : 10.1111/joa.13679 . ПМЦ 9296034 . ПМИД 35801524 .
{{cite journal}}: CS1 maint: Срок эмбарго PMC истек ( ссылка ) - ^ Пол, GS, 2010, Принстонский полевой справочник по динозаврам, Princeton University Press, стр. 304
- ^ Конти, С.; Вила, Б.; Селлес, АГ; Галобарт, А.; Бентон, MJ; Прието-Маркес, А. (2020). «Самый старый динозавр-ламбеозавр из Европы: информация о прибытии цинтаозаурини» (PDF) . Меловые исследования . 107 : 40. Бибкод : 2020CrRes.10704286C . дои : 10.1016/j.cretres.2019.104286 . hdl : 1983/be876efb-979c-4237-94f9-5f8d80121f7e . S2CID 208195457 . Архивировано (PDF) из оригинала 27 апреля 2021 года.
- ^ Киркланд, Дж.И.; Эрнандес-Ривера, Р.; Гейтс, Т.; Пол, GS; Несбитт, С.; Серрано-Браньяс, CI; Гарсиа-де ла Гарса, JP (2006). «Большие гадрозавровые динозавры из позднего кампана Коауила, Мексика». В Лукасе, СГ; Салливан, Р.М. (ред.). Позднемеловые позвоночные западного региона (PDF) . Бюллетень Музея естественной истории и науки Нью-Мексико. Том. 35. Альбукерке: Музей естественной истории и науки Нью-Мексико. стр. 299–315. Архивировано (PDF) из оригинала 14 июля 2019 года.
- ^ Пауэрс, Марк (11 июля 2004 г.). «Холден подписывает государственный законопроект о динозаврах» . Дейли Данкин Демократ . Дейли Данклин Демократ. Архивировано из оригинала 4 февраля 2011 года . Проверено 29 августа 2022 г.
- ^ Пауэрс, Марк (19 февраля 2004 г.). «Кость для Миссури» . Юго-восточный штат Миссури . Архивировано из оригинала 18 ноября 2015 года . Проверено 29 августа 2022 г.
- ^ Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 342. ИСБН 978-1-78684-190-2 . OCLC 985402380 .
- ^ Верду, Ф.Дж.; Годфруа, П.; Ройо-Торрес, Р.; Кобос, А.; Алькала, Л. (2017). «Индивидуальная вариация посткраниального скелета раннемелового Iguanodon bernissartensis (Dinosauria: Ornithopoda)». Меловые исследования . 74 : 65–86. Бибкод : 2017CrRes..74...65В . дои : 10.1016/j.cretres.2017.02.006 .
- ^ Гасулла, Дж. М.; Эскасо, Ф.; Нарваес, И.; Санс, Дж.Л.; Ортега, Ф. (2022). «Новые осевые кости Iguanodon bernissartensis (Dinosauria, Ornithopoda) из раннего мела Мореллы, Испания» . Разнообразие . 14 (2): 63. дои : 10.3390/d14020063 .
- ^ Най, Э.; Файст-Буркхардт, С.; Хорн, диджей; Росс, Эй Джей; Уиттакер, Дж. Э. (2008). «Палеосреда, связанная с частичным скелетом игуанодона из глины Верхнего Уилда (баррем, ранний мел) на кирпичном заводе Smokejacks (Окли, Суррей, Великобритания), на основе палиноморфов и остракод». Меловые исследования . 29 (3): 417–444. Бибкод : 2008CrRes..29..417N . дои : 10.1016/j.cretres.2008.01.004 .
- ^ Макдональд, AT; Киркланд, Дж.И.; ДеБлё, Д.Д.; Мадсен, СК; Кэвин, Дж.; Милнер, Арканзас; Панзарин, Л. (2010). «Новые базальные игуанодонты из формации Кедровой горы в штате Юта и эволюция динозавров с шипами на больших пальцах» . ПЛОС ОДИН . 5 (11): e14075. Бибкод : 2010PLoSO...514075M . дои : 10.1371/journal.pone.0014075 . ПМЦ 2989904 . ПМИД 21124919 .
- ^ Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам, 2-е издание . Нью-Джерси: Издательство Принстонского университета. п. 319.
- ^ Билл, А. (2017). «У этого «уродливого» динозавра были гигантские зубы, похожие на ножницы» . Альфр . Деннис Паблишинг.
Думаю, он был метров пять в длину.
- ^ Пол, Грегори С. (2016). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. п. 312. ИСБН 978-1-78684-190-2 . OCLC 985402380 .
- ^ Перейти обратно: а б с д и ж г час я Мюррей, П.Ф. и Викерс-Рич, П. (2004)
- ^ Перейти обратно: а б с Дегранж, Ф.Дж., Норьега, Дж.И., и Арета, Дж.И. (2012). Разнообразие и палеобиология сантакруцианских птиц. Палеобиология раннего миоцена в Патагонии: высокоширотные палеосообщества формации Санта-Крус , 138–155.
- ^ Перейти обратно: а б Лазаро, Энрико де (28 июня 2019 г.). «Гигантские птицы бродили по Европе два миллиона лет назад | Палеонтология | Sci-News.com» . Sci.News: Последние новости науки . Проверено 3 марта 2023 г.
- ^ Перейти обратно: а б Молина-Перес, Рубен; Ларраменди, Азиер (2019). Факты и цифры о динозаврах: Тероподы Издательство Принстонского университета. п. 179. ИСБН 978-0-691-18031-1 .
- ^ Баффо1, Эрик; Ага, Дельфин (2017). «Насколько большим был гигантский китайский страус?». . ЭВОЛЮЦИЯ - Журнал Geistória и предыстории . 2 (1): 6–8 . Проверено 9 января 2023 г.
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Бреннан, Имоджен (19 февраля 2016 г.). «Кости предка «гигантской утки» обнаружены в Австралии» . ПМ . www.abc.net.au.
- ^ Тамбусси, CP, и Дегранж, FJ (2013). Доминирование птиц-зоофагов: просто клише?. У южноамериканских и антарктических континентальных кайнозойских птиц (стр. 87–102). Спрингер, Дордрехт.
- ^ Гулд, GC; Куитмайер, ИК (2005). «Титанис Валлери: Кости раздора» (PDF) . Бюллетень Музея естественной истории Флориды . 45 (4): 201–229. дои : 10.58782/flmnh.xumx1681 .
- ^ Перейти обратно: а б Эрик Бюффето и Дельфина Ангст: Стратиграфическое распространение крупных нелетающих птиц в палеогене Европы и его палеобиологические и палеогеографические последствия. Earth-Science Reviews 32 (2), 2014, С. 394–408.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Вес мяса и пищевая ценность археофауны Новой Зеландии
- ^ Перейти обратно: а б Рубен Молина-Перес; Асьер Ларраменди; Дэвид Коннолли; Гонсало Анхель Рамирес Крус; Андрей Атучин (25 июня 2019 г.). Факты и цифры о динозаврах: тероподы и другие динозаврообразные Издательство Принстонского университета. п. 282. ИСБН 978-0-691-19059-4 . Проверено 30 августа 2022 г.
- ^ Госпиталече, Каролина А. (2014). «Новые кости гигантских пингвинов из Антарктиды: систематическое и палеобиологическое значение» . Comptes Рендус Палевол . 13 (7): 555–560. Бибкод : 2014CRPal..13..555A . дои : 10.1016/j.crpv.2014.03.008 . hdl : 11336/32571 .
- ^ Перейти обратно: а б с Александр, Дэвид Э. (24 июля 2007 г.). «Древний Аргентавис снова парит» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (30): 12233–12234. Бибкод : 2007PNAS..10412233A . дои : 10.1073/pnas.0705515104 . ЧВК 1941455 . ПМИД 17640902 .
- ^ Перейти обратно: а б с Палеобиология раннего миоцена в Патагонии: высокоширотные палеосообщества Патагонии»
- ^ Амадон (1947), Расмуссен и др. (1987, 2001)
- ^ Фишер, Харви И. (1945). «Передвижение ископаемых грифов Teratornis». Американский натуралист из Мидленда . 33 (3): 725–742. дои : 10.2307/2421186 . ISSN 0003-0031 . JSTOR 2421186 .
- ^ Ксепка, Дэниел Т. (7 июля 2014 г.). «Летные качества самой крупной летучей птицы» . ПНАС . 111 (29): 10624–10629. Бибкод : 2014PNAS..11110624K . дои : 10.1073/pnas.1320297111 . ПМЦ 4115518 . ПМИД 25002475 .
- ^ Перкинс, Сид (9 апреля 2015 г.). «Обнаружен новый вид «птицы ужаса»» . Наука . дои : 10.1126/science.aab2465 . Проверено 14 апреля 2015 г.
- ^ Перейти обратно: а б Арета, Дж.И.; Норьега, Дж.И.; Агнолин, Ф. (2007). «Гигантский дротик (Pelecaniformes: Anhingidae) из верхнего миоцена Аргентины и расчет веса ископаемых Anhingidae». Новогодний альбом по геологии и палеонтологии – Abhandlungen . 243 (3): 343–350. дои : 10.1127/0077-7749/2007/0243-0343 . hdl : 11336/80812 .
- ^ НОРТКОТ, EM (2008). РАЗМЕР, ФОРМА И ПРИВИТИКА ВЫМЕРШЕГО МАЛЬТИЙСКОГО ЛЕБЕДЯ CYGNUS FALCONERI. Ибис, 124 (2), 148–158. https://doi.org/10.1111/j.1474-919x.1982.tb03753.x
- ^ Перейти обратно: а б Мейер Х.Дж., Р.А.Дью (2010). «Новый вид гигантского аиста марабу (Aves: Ciconiiformes) из плейстоцена Лян Буа, Флорес (Индонезия)» . Зоологический журнал Линнеевского общества . 160 (4): 707–724. дои : 10.1111/j.1096-3642.2010.00616.x .
- ^ Ангст, Д.; Бюффето, Э.; Абурашид, А. (март 2011 г.). «Конец толстого дронта? Новая оценка массы Raphus cucullatus ». Naturwissenschaften . 98 (3): 233–236. Бибкод : 2011NW.....98..233A . дои : 10.1007/s00114-010-0759-7 . ПМИД 21240603 . S2CID 29215473 .
- ^ Достойно, Тревор Х.; Хэнд, Сюзанна Дж.; Арчер, Майкл; Скофилд, Р. Пол; Де Пьетри, Ванеса Л. (август 2019 г.). «Свидетельства существования гигантского попугая из раннего миоцена Новой Зеландии» . Письма по биологии . 15 (8): 20190467. doi : 10.1098/rsbl.2019.0467 . ISSN 1744-9561 . ПМК 6731479 . ПМИД 31387471 .
- ^ Перейти обратно: а б Калифорнийская академия наук - Наука под парусом. Архивировано 7 июня 2012 года в Wayback Machine . Calacademy.org
- ^ Перейти обратно: а б с Колин Тадж (20 октября 2009 г.). Птица: естественная история того, кто такие птицы, откуда они пришли и как они живут . Корона. п. 64. ИСБН 978-0-307-45976-3 . Проверено 29 августа 2022 г.
- ^ Перейти обратно: а б с д и Рубен Молина-Перес; Асьер Ларраменди; Дэвид Коннолли; Гонсало Анхель Рамирес Крус; Андрей Атучин (25 июня 2019 г.). Факты и цифры о динозаврах: тероподы и другие динозаврообразные Издательство Принстонского университета. п. 281. ИСБН 978-0-691-19059-4 . Проверено 29 августа 2022 г.
- ^ Уокер, Сирил А.; Дайк, Гарет Дж. (2009). «Эуэнантиорнитиновые птицы из позднего мела Эль-Брете (Аргентина)» . Ирландский журнал наук о Земле . 27 :15–62. дои : 10.1353/ijes.2009.a810006 . ISSN 0790-1763 . JSTOR 25780698 .
- ^ Чьяппе, Луис М. «Энантиорнитин (Aves) tarsometatarsi из меловой формации Лечо на северо-западе Аргентины. Американский музей новитирует; № 3083» . Библиотека наследия биоразнообразия . Проверено 20 декабря 2023 г.
- ^ Аттерхольт, Джесси; Хатчисон, Дж. Ховард; О'Коннор, Цзинмай К. (13 ноября 2018 г.). «Наиболее полный энантиорнитин из Северной Америки и филогенетический анализ Avisauridae» . ПерДж . 6 : е5910. дои : 10.7717/peerj.5910 . ПМК 6238772 . ПМИД 30479894 .
- ^ Чжоу, Чжунхэ; Кларк, Джулия; Чжан, Фучэн (май 2008 г.). «Понимание разнообразия, размера тела и морфологической эволюции крупнейшей энантиорнитиновой птицы раннего мела» . Журнал анатомии . 212 (5): 565–77. дои : 10.1111/j.1469-7580.2008.00880.x . ПМК 2409080 . ПМИД 18397240 .
- ^ Бюффето, Э.; Ле Лефф, Ж. (1998). «Новая гигантская наземная птица из верхнего мела на юге Франции». Журнал Геологического общества, Лондон . 155 (155): 1–4. Бибкод : 1998JGSoc.155....1B . дои : 10.1144/gsjgs.155.1.0001 . S2CID 128496095 .
- ^ Мюссер, Энн. «Громовая птица Стиртона» . Австралийский музей . Архивировано из оригинала 31 августа 2022 года . Проверено 10 сентября 2022 г.
- ^ Чинсами, А., Хэндли, В.Д., Йейтс. AM, Уорти, TH (2016). Половой диморфизм позднемиоценового mihirung Dromornis stirtoni (Aves: Dromornithidae) из местной фауны Алкуты центральной Австралии. Журнал палеонтологии позвоночных, 36:5, DOI: 10.1080/02724634.2016.1180298.
- ^ Уитмер, Лоуренс; Роуз, Кеннет (1991). «Биомеханика челюстного аппарата гигантской эоценовой птицы Diatryma : Значение для диеты и образа жизни» (PDF) . Палеобиология . 17 (2): 95–120. Бибкод : 1991Pbio...17...95W . дои : 10.1017/S0094837300010435 . S2CID 18212799 . Архивировано из оригинала (PDF) 28 июля 2010 года.
- ^ Джеральд Майр: Gastornithidae. В: Джеральд Майр: Ископаемые птицы палеогена. Springer-Verlag, Берлин и Гейдельберг, 2009 г., стр. 44–47.
- ^ Дельфина Ангст и Эрик Баффето: Палеобиология гигантских нелетающих птиц. Оксфорд, 2017, С. 1–282 (С. 173–214).
- ^ Мейер, HJM (2014). «Своеобразный гусеобразный (Aves: Anseriformes) из миоцена Гаргано (Италия)» . Comptes Рендус Палевол . 13 (1): 19–26. Бибкод : 2014CRPal..13...19M . дои : 10.1016/j.crpv.2013.08.001 .
- ^ TH Worthy , Ричард Н. Холдэуэй (2002). Затерянный мир Моа. Доисторическая жизнь Новой Зеландии . Издательство Университета Индианы. п. 232. ИСБН 978-0-253-34034-4 . Проверено 1 октября 2022 г.
- ^ Сноу, Д.В.; Перринс, CM (1998). Птицы Западной Палеарктики (Краткое изд.). ОУП. ISBN 0-19-854099-Х .
- ^ НОРТКОТ, EM (2008). Размер, форма и облик вымершего мальтийского лебедя Cygnus falconeri. Ибис, 124 (2), 148–158. https://doi.org/10.1111/j.1474-919x.1982.tb03753.x
- ^ Ватанабэ Дж. и Мацуока Х. (2015). Нелетающая ныряющая утка (Aves, Anatidae) из плейстоцена Ширии, северо-восток Японии. Журнал палеонтологии позвоночных, 35 (6), e994745. https://doi.org/10.1080/02724634.2014.994745
- ^ Ватанабэ, Дж. (2017). Количественное распознавание нелетания у ископаемых Anatidae по пропорциям скелета. Аук, 134 (3), 672–695. https://doi.org/10.1642/auk-17-23.1
- ^ Гибб, Джон (27 июля 2017 г.). «Наконец-то собственный лебедь Новой Зеландии» . Отаго Дейли Таймс . Архивировано из оригинала 17 августа 2017 года . Проверено 1 октября 2022 г.
- ^ Мацуока Х, Хасегава Ю (2022). « Annakacygna , новый род двух замечательных нелетающих лебедей (Aves, Anatidae, Cygnini) из миоцена Гуммы, центральная Япония: с примечанием о сдвиге пищевой ниши птиц и специализации крыльев для родительской заботы» (PDF) . Вестник Музея естественной истории Гумма . 26 : 1–30.
- ^ Достойно, Тревор Х.; Олсон, Сторрс Л. (2002). «Отношения, адаптации и повадки вымершей утки «Euryanas» finschi» (PDF) . Ноторнис . 49 (1): 1–17. Архивировано из оригинала (PDF) 19 октября 2007 года.
- ^ Олсон, Сторрс Л. (1999). «Новый вид пеликана (Aves: Pelecanidae) из нижнего плиоцена Северной Каролины и Флориды» (PDF) . Труды Биологического общества Вашингтона . 112 (3): 503–509. Архивировано из оригинала (PDF) 10 октября 2022 года.
- ^ Каллахан, Д. (2014). История наблюдения за птицами в 100 объектах . Блумсбери Паблишинг Plc. п. 15. ISBN 978-1408-1-8618-3 .
- ^ «На острове хоббитов найдена гигантская птица» . 7 декабря 2010 г. - через BBC News.
- ^ Халберт-младший, Ричард К. и Вальдес, Натали (2013). «Окаменелости позвоночных Флориды: Ciconia Maltha » . Флоридский музей естественной истории . Проверено 30 сентября 2015 г.
- ^ Г. Майр; Томас Лехнер; М. Бёме (14 июля 2020 г.). «Череп очень крупного журавля из позднего миоцена Южной Германии с заметками о филогенетических взаимоотношениях современных Gruinae» . Журнал орнитологии . 161 (4): 923–933. дои : 10.1007/s10336-020-01799-0 . S2CID 220505689 .
- ^ Смит, Н. 2015. Эволюция массы тела у Pan-Alcidae (Aves, Charadriiformes): эффекты объединения неонтологических и палеонтологических данных. Палеобиология. дои: 10.1017/pab.2015.24
- ^ Уилсон, Лаура; Чин, Карен; Камба, Стивен; Дайк, Гарет (1 марта 2011 г.). «Высокоширотная гесперорнитиформа (Aves) с острова Девон: палеобиогеография и размерное распределение гесперорнитиформ Северной Америки» . Журнал систематической палеонтологии . 9 (1): 9–23. Бибкод : 2011JSPal...9....9W . дои : 10.1080/14772019.2010.502910 . S2CID 83749554 .
- ^ Ксепка, Дэниел Т. (22 июля 2014 г.). «Летные качества самой крупной летучей птицы» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (29): 10624–10629. Бибкод : 2014PNAS..11110624K . дои : 10.1073/pnas.1320297111 . ПМЦ 4115518 . ПМИД 25002475 .
- ^ Чаттерджи, С.; Темплин, Р.Дж.; Кэмпбелл, Кентукки (24 июля 2007 г.). «Аэродинамика Аргентависа , крупнейшей в мире летающей птицы из миоцена Аргентины» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (30): 12398–12403. Бибкод : 2007PNAS..10412398C . дои : 10.1073/pnas.0702040104 . ЧВК 1906724 . ПМИД 17609382 .
- ^ Кэмпбелл, Кеннет Э. младший; Тони, EP (1983). «Размер и передвижение тераторнов» (PDF ) Аук . 100 (2): 390–403. дои : 10.1093/au/100.2.390 .
- ^ Кэмпбелл, Кеннет; Скотт, Эрик; Спрингер, Кэтлин (1999). «Новый род невероятного тераторна» . Смитсоновский вклад в палеобиологию . 89 : 169–175 – через Researchgate.
- ^ Кеннет Э. Кэмпбелл-младший, Эдвард П. Тонни. « Размер и передвижение тераторов (Teratornithidae) ». The Auk, том 100, выпуск 2, апрель 1983 г., с. 390–403, https://doi.org/10.1093/au/100.2.390
- ^ Альваренга, Эркулано МФ; Хёфлинг, Элизабет (2003). «Синтетический обзор Phorusrhacidae (Птицы: Ralliformes)» . Отдельные статьи по зоологии . 43 (4). дои : 10.1590/S0031-10492003000400001 .
- ^ Перейти обратно: а б Ксепка, Дэниел Т. (2014). «Полёты фантазии в эволюции птиц». Американский учёный . 102 (1): 39. дои : 10.1511/2014.106.36 . ISSN 0003-0996 . JSTOR 43707746 .
- ^ Тамбусси, Клаудия П.; Дегранж, Федерико Дж. (2013). «Неогеновые птицы Южной Америки». Южноамериканские и антарктические континентальные кайнозойские птицы . Springer Briefs по наукам о системе Земли. Спрингер. стр. 59–86. дои : 10.1007/978-94-007-5467-6_7 . ISBN 978-94-007-5466-9 .
- ^ Крофт, Дарин А. (2016). Рогатые броненосцы и рафтинговые обезьяны: удивительные ископаемые млекопитающие Южной Америки . Издательство Университета Индианы. п. 120. ИСБН 978-0-253-02084-0 . Проверено 30 марта 2020 г.
- ^ Альваренга, Эркулано МФ; Хёфлинг, Элизабет (2003). «Синтетический обзор Phorusrhacidae (Птицы: Ralliformes)» . Отдельные статьи по зоологии . 43 (4): 55–91. дои : 10.1590/S0031-10492003000400001 .
- ^ Рид Викандер, Джеймс С. Монро (2015). Историческая геология . Cengage Обучение. п. 368. ИСБН 9781305480766 . Проверено 27 августа 2022 г.
- ^ Маас, П. «Недавно вымершие животные – Информация о видах – Орел Хааста» . Шестое вымирание. Архивировано из оригинала 20 января 2013 года . Проверено 19 апреля 2013 г.
- ^ «Орел Хааста» . Группа исследований палеобиологии и биоразнообразия. Архивировано из оригинала 5 мая 2014 года . Проверено 19 апреля 2013 г.
- ^ Уорти, Т. и Холдэуэй, Р., Затерянный мир Моа: доисторическая жизнь Новой Зеландии . Издательство Университета Индианы (2003), ISBN 978-0-253-34034-4
- ^ Брэтуэйт, Д.Х. (декабрь 1992 г.). «Заметки о весе, летных способностях, среде обитания и добыче орла Хааста ( Harpagornis moorei )» (PDF) . Ноторнис . 39 (4). Орнитологическое общество Новой Зеландии : 239–247. Архивировано из оригинала (PDF) 19 января 2012 года . Проверено 26 января 2014 г.
- ^ Даррен Нэйш (27 января 2008 г.). «Титаны-ястребы и другие суперхищники» . Научные блоги. Архивировано из оригинала 25 сентября 2022 года . Проверено 30 сентября 2022 г.
- ^ Дэвид В. Стедман, Хуан Н. Альмонте Милан и Алексис М. Мичайлив « Вымерший орел (Aves: Accipitridae) из четвертичного периода Эспаньолы ». Журнал исследований хищников 53 (3), 319–333, (13 августа 2019 г.). https://doi.org/10.3356/JRR-18-769
- ^ Олсон, Сторрс Л.; Суарес, Уильям (20 апреля 2007 г.). «Кубинский ископаемый орел Aquila borrasi Arredondo: увеличенная версия большого черного ястреба Buteogallus urubitinga (Gmelin)» (PDF) . Журнал исследований хищников . 41 (4). Фонд исследования хищников : 288. doi : 10.3356/0892-1016(2007)41[288:TCFEAB]2.0.CO;2 . S2CID 55380044 .
- ^ Олсон, Сторрс Л. (2007). «Ходячий орел» Wetmoregyps daggetti Miller: увеличенная версия саванного ястреба ( Buteogallus meridionalis )» (PDF) . Орнитологические монографии (63): 110–114. дои : 10.2307/40166902 . JSTOR 40166902 . Проверено 30 сентября 2022 г.
- ^ Холдэуэй, Ричард Н.; Достойно, Тревор Х. (март 1997 г.). «Переоценка ископаемых позвоночных позднечетвертичного периода болота Пирамидальной долины, Северный Кентербери, Новая Зеландия» . Новозеландский журнал зоологии . 24 (1): 69–121. дои : 10.1080/03014223.1997.9518107 .
- ^ Рахаман, Садекур (8 ноября 2014 г.). «10 крупнейших птиц в мире (живые)» AllTopTens.com» .
- ^ Перейти обратно: а б Вуд, Джеральд Книга рекордов Гиннеса по фактам и подвигам животных (1983) ISBN 978-0-85112-235-9
- ^ Марк Эш; Хорхе Норьега; Джон Дидерле; Стивен Сойбельзон; Леопольд Сойбельзон; Серхио Родригес; Элиша Бейлинсон (2018). «Неожиданный крупный хохлатый тинаму (Eudromia, Tinamidae, Birds) вблизи последнего ледникового максимума (MIS 2, поздний плейстоцен) в аргентинских пампасах » Историческая биология 32 (3): 330–338. дои : 10.1080/08912963.2018.1491568 . hdl : 11336/84901 . S2CID 91851921 .
- ^ Зеленков, Н.В.; Лавров А.В.; Старцев Д.Б.; Вислобокова И.А.; Лопатин, А.В. (2019). «Гигантская птица раннего плейстоцена из Восточной Европы: неожиданный компонент наземной фауны во время прибытия раннего человека». Журнал палеонтологии позвоночных . 39 (2): e1605521. Бибкод : 2019JVPal..39E5521Z . дои : 10.1080/02724634.2019.1605521 . S2CID 198384367 .
- ^ Эрик Баффето, Дельфин Ангст. «Гигантский страус из формации Нихэван нижнего плейстоцена в Северном Китае, с обзором ископаемых страусов Китая». Разнообразие 2021, 13, 47. https://doi.org/10.3390/d13020047 .
- ^ Джеральд Майр (31 октября 2016 г.). Птичья эволюция. Ископаемая летопись птиц и ее палеобиологическое значение . Уайли. п. 98. ИСБН 978-1-119-02076-9 . Проверено 30 сентября 2022 г.
- ^ Сесиль Мурер-Шовир, Денис Гераадс (январь 2008 г.). «Struthionidae и Pelagornithidae позднего плиоцена Ахль-эль-Углама, Марокко» . Ориктос 7 : 169–194.
- ^ Вермей, Гират Дж. (2004). Природа: экономическая история . Издательство Принстонского университета, ISBN 0-691-11527-3
- ^ Ангст, Д.; Бюффето, Э.; Абурашид, А. (2011). «Конец толстого дронта? Новая оценка массы Raphus cucullatus». Naturwissenschaften . 98 (3): 233–236. Бибкод : 2011NW.....98..233A . дои : 10.1007/s00114-010-0759-7 . ПМИД 21240603 . S2CID 29215473 .
- ^ Викерс-Рич, Патрисия; Найт, Фрэнк (1985). Кадимакара – вымершие позвоночные Австралии . Издательство Принстонского университета. ISBN 978-0-909674-26-7 .
- ^ Риндеркнехт, А.; Норьега, Дж.И. (2002). «Новый род Anhingidae (Птицы: Pelecaniformes) из плиоцена-плейстоцена Уругвая (формация Сан-Хосе)» . Амегиниана . 39 (2): 183–191. Архивировано (PDF) из оригинала 9 мая. Получено 5 мая.
- ^ Мори, Хироцугу; Мията, Казунори (2021). «Особи ранних Plotopteridae (птиц) из формаций Итанура и Какиноура (от позднего эоцена до раннего олигоцена), Сайкай, префектура Нагасаки, Западная Япония» . Палеонтологические исследования . 25 (2): 145–159. дои : 10.2517/2020PR018 . ISSN 1342-8144 . S2CID 233029559 .
- ^ Гриском, Ладлоу (1929). «Исследования птиц Гватемалы из коллекции Дуайта. 1». Американский музей послушников (379). hdl : 2246/5418 .
- ^ «Императорский дятел Campephilus Imperialis » . БердЛайф Интернэшнл. Архивировано из оригинала 4 марта 2016 года . Проверено 28 февраля 2011 г.
- ^ Достойно, Тревор Х.; Хэнд, Сюзанна Дж.; Арчер, Майкл; Скофилд, Р. Пол; Де Пьетри, Ванеса Л. (2019). «Свидетельства существования гигантского попугая из раннего миоцена Новой Зеландии» . Письма по биологии . 15 (8). дои : 10.1098/rsbl.2019.0467 . ПМК 6731479 . ПМИД 31387471 .
- ^ Перейти обратно: а б с д Ксепка, Дэниел Т.; Филд, Дэниел Дж.; Хит, Трейси А.; Петт, Уокер; Томас, Дэниел Б.; Джованарди, Симоне; Теннисон, Алан Джей Ди (8 февраля 2023 г.). «Самый крупный из известных ископаемых пингвинов дает представление о ранней эволюции размеров клиновидного тела и анатомии ласт» . Журнал палеонтологии . 97 (2): 434–453. Бибкод : 2023JPal...97..434K . дои : 10.1017/jpa.2022.88 . ISSN 0022-3360 . S2CID 256709376 .
- ^ Акоста Госпиталече, Каролина (1 октября 2014 г.). «Новые кости гигантских пингвинов из Антарктиды: систематическое и палеобиологическое значение» . Comptes Рендус Палевол . 13 (7): 555–560. Бибкод : 2014CRPal..13..555A . дои : 10.1016/j.crpv.2014.03.008 . hdl : 11336/32571 . ISSN 1631-0683 .
- ^ «Размеры тела эоценовых антарктических пингвинов» . Польские полярные исследования .
- ^ Шмид, Рэндольф Э. (25 июня 2007 г.). «Гигантские пингвины, возможно, бродили по Перу» . Associated Press через Washington Post . Проверено 29 июня 2007 г.
- ^ Арредондо, Оскар (1976) перевел Олсона, Сторрса Л. Великие хищные птицы плейстоцена Кубы, стр. 169–187 в «Смитсоновский вклад в палеобиологию, номер 27; Сборник статей по палеонтологии птиц в честь 90-летия со дня рождения Александра Ветмора»
- ^ Перейти обратно: а б Дэниел Л. Леви, Ребекка Хилд (август 2015 г.). «Проблемы биологического масштабирования и решения у амфибий» . Перспективы Колд-Спринг-Харбор в биологии . 8 (1): а019166. doi : 10.1101/cshperspect.a019166 . ПМЦ 4691792 . ПМИД 26261280 .
- ^ Родриго А. Отеро; Паулина Хименес-Уидоброб; Серджио Сото-Акуна; Роберто Э. Юрий-Янес (2014). «Свидетельства существования гигантской шлемованной лягушки (Australobatrachia, Calyptocephalellidae) с эоценовых уровней бассейна Магальянес, на самом юге Чили» . Журнал южноамериканских наук о Земле . 55 : 133–140. Бибкод : 2014JSAES..55..133O . дои : 10.1016/j.jsames.2014.06.010 .
- ^ Эванс, Сьюзен Э.; Гроенке, Джозеф Р.; Джонс, Марк Э.Х.; Тернер, Алан Х.; Краузе, Дэвид В. (2014). «Новый материал о Beelzebufo , гипероссифицированной лягушке (амфибия: Anura) из позднего мела Мадагаскара» . ПЛОС ОДИН . 9 (1): e87236. Бибкод : 2014PLoSO...987236E . дои : 10.1371/journal.pone.0087236 . ПМК 3905036 . ПМИД 24489877 .
- ^ Нейлор, Брюс Г. (1981). «Криптожаберные саламандры из палеоцена и миоцена Саскачевана». Копейя . 1981 (1): 76–86. дои : 10.2307/1444042 . ISSN 0045-8511 . JSTOR 1444042 .
- ^ Уилсон, Грегори П.; Клеменс, Уильям А.; Хорнер, Джон Р.; Хартман, Джозеф Х. (21 января 2014 г.). В конце мелового периода в типовом местонахождении формации Хелл-Крик в Монтане и прилегающих районах . Геологическое общество Америки. ISBN 978-0-8137-2503-1 .
- ^ Лори Дж. Витт, Джанали П. Колдуэлл (25 марта 2013 г.). Герпетология. Вводная биология амфибий и рептилий . Эльзевир Наука. п. 96. ИСБН 978-0-12-386920-3 . Проверено 25 сентября 2022 г.
- ^ Георг Ф. Стридтер , Р. Гленн Норткатт (2020). Мозг сквозь время. Естественная история позвоночных . Издательство Оксфордского университета. п. 276. ИСБН 978-0-19-512568-9 . Проверено 25 сентября 2022 г.
- ^ Панчен, Ал. (1977). «Об Anthracosaurus russelli Huxley (амфибия: Labyrinthodontia) и семействе Anthracosauridae». Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 279 (968): 447–512. Бибкод : 1977РСТБ.279..447П . дои : 10.1098/rstb.1977.0096 . ISSN 0080-4622 . JSTOR 2417840 .
- ^ Марьянович, Давид; Лорен, Мишель (4 января 2019 г.). «Филогения палеозойских конечностей позвоночных, заново оцененная посредством пересмотра и расширения крупнейшей опубликованной матрицы соответствующих данных» . ПерДж . 6 : е5565. дои : 10.7717/peerj.5565 . ISSN 2167-8359 . ПМК 6322490 . ПМИД 30631641 .
- ^ Панчен, Ал. (1972). «Череп и скелет Eogyrinus attheyi Watson (амфибия: Labyrinthodontia)». Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 263 (851): 279–326. Бибкод : 1972РСТБ.263..279П . дои : 10.1098/rstb.1972.0002 . ISSN 0080-4622 . JSTOR 2417176 .
- ^ Шох, Р.Р. (1999). «Сравнительная остеология Mastodonsaurus giganteus (Jaeger, 1828) из среднего триаса (Lettenkeuper: Longobardian) Германии (Баден-Вюртемберг, Бавария, Тюрингия)» (PDF) . Вклад Штутгарта в серию по естествознанию B. 278 : 1–175.
- ^ Штайер, Ж. Себастьян; Дамиани, Росс (1 мая 2005 г.). «Гигантский брахиопоидный темноспондил из верхнего триаса или нижней юры Лесото». Бюллетень Геологического общества Франции . 176 (3): 243–248. дои : 10.2113/176.3.243 . ISSN 0037-9409 .
- ^ Лю, Хуайбао П.; Бергстрем, Стиг М.; Витцке, Брайан Дж.; Бриггс, Дерек Э.Г.; Маккей, Роберт М.; Ферретти, Анналиса (2017). «Исключительно сохранившиеся аппараты конодонтов с гигантскими элементами из среднего ордовика Виннешик Консерват-Лагерштетте, Айова, США» . Журнал палеонтологии . 91 (3): 493–511. Бибкод : 2017JPal...91..493L . дои : 10.1017/jpa.2016.155 . ISSN 0022-3360 . S2CID 132698401 .
- ^ «Систематика, филогенетика и макроэволюция гетеростраков: исследование закономерностей эволюции вымерших бесчелюстных позвоночных | Research Explorer | Манчестерский университет» . www.research.manchester.ac.uk . Проверено 24 августа 2022 г.
- ^ Тернер, Сьюзен (1986). «Thelodus macintoshi Stetson 1928, самый крупный из известных телодонтов (Agnatha: Thelodonti)» . Бревиора . 486 : 1–18. ISSN 0006-9698 .
- ^ Альберт, Джеймс С.; Джонсон, Дерек М.; Кнауфт, Джейсон Х. (2009). «Окаменелости позволяют лучше оценить размеры тела предков, чем современные таксоны рыб» . Акта Зоология . 90 : 357–384. дои : 10.1111/j.1463-6395.2008.00364.x .
- ^ Орвиг, Тор (1967). «Некоторые новые акантодические материалы из нижнего девона Европы» . Журнал Лондонского Линнеевского общества, Зоология . 47 (311): 131–153. дои : 10.1111/j.1096-3642.1967.tb01400.x .
- ^ Перейти обратно: а б с д и Энгельман, Рассел К. (2023). «Сказка о девонских рыбах: новый метод оценки длины тела предполагает гораздо меньшие размеры Dunkleosteus terrelli (Placodermi: Arthrodira)» . Разнообразие . 15 (3): 318. дои : 10.3390/d15030318 . ISSN 1424-2818 .
- ^ Гилберт Пауэлл Ларвуд, Пол Д. Тейлор (1990). Основные эволюционные излучения . Ассоциация систематиков. п. 310. ИСБН 978-0-19-857718-8 . Проверено 28 августа 2022 г.
- ^ Бюллетень 70 . Огайо. Отдел геологической разведки. 1996. с. 290. ИСБН 9780931079054 . Проверено 28 августа 2022 г.
- ^ Чарли Андервуд; Марта Рихтер; Зерина Йохансон (2019). Эволюция и развитие рыб . Издательство Кембриджского университета. п. 13. ISBN 978-1-107-17944-8 . Проверено 31 августа 2022 г.
- ^ Башфорд Дин (1895). Рыбы, живые и ископаемые: очерк их форм и вероятных взаимоотношений . Макмиллан и компания. п. 130 . Проверено 28 августа 2022 г.
- ^ Шимада, К.; Чендлер, RE; Лам, ОЛТ; Танака, Т.; Уорд, диджей (2016). «Новая неуловимая отодонтидная акула (Lamniformes: Otodontidae) из нижнего миоцена и комментарии к таксономии родов отодонтид, включая кладу «мегатубатых». Историческая биология . 29 (5): 1–11. дои : 10.1080/08912963.2016.1236795 . S2CID 89080495 .
- ^ Перейти обратно: а б Симада, Кенсю (2019). «Возвращение к размеру большезубой акулы Otodus megalodon (Lamniformes: Otodontidae)». Историческая биология . 33 (7): 1–8. дои : 10.1080/08912963.2019.1666840 . ISSN 0891-2963 . S2CID 208570844 .
- ^ Перейти обратно: а б Купер, Дж.А.; Пимьенто, К.; Феррон, ХГ; Бентон, MJ (2020). «Размеры тела вымершей гигантской акулы Otodus megalodon : 2D-реконструкция» . Научные отчеты . 10 (14596): 14596. Бибкод : 2020NatSR..1014596C . дои : 10.1038/s41598-020-71387-y . ПМЦ 7471939 . ПМИД 32883981 .
- ^ деГрюи, Майкл (2006). Идеальная акула (сериал). Би-би-си (Великобритания).
- ^ Перес, Виктор; Ледер, Ронни; Бадо, Тедди (2021). «Оценка длины тела неогеновых макрофагов пластинчатообразных акул (Carcharodon и Otodus), полученная на основе связанных с ними ископаемых зубных рядов» . Электронная палеонтология . 24 (1): 1–28. дои : 10.26879/1140 .
- ^ Пимьенто, К.; Макфадден, Би Джей; Клементс, CF; Варела, С.; Харамильо, К.; Велес-Хуарбе, Дж.; Силлиман, БР (2016). «Географические закономерности распространения Carcharocles megalodon с течением времени открывают подсказки о механизмах вымирания». Журнал биогеографии . 43 (8): 1645–1655. Бибкод : 2016JBiog..43.1645P . дои : 10.1111/jbi.12754 . S2CID 55776834 .
- ^ Пимьенто, К.; Балк, Массачусетс (2015). «Тенденции размеров тела вымершей гигантской акулы Carcharocles megalodon : глубокий взгляд на морских хищников» . Палеобиология . 41 (3): 479–490. Бибкод : 2015Pbio...41..479P . дои : 10.1017/pab.2015.16 . ПМЦ 4541548 . ПМИД 26321775 .
- ^ Ренц, Марк (2002). Мегалодон: Охота на охотника . Лихай-Эйкерс, Флорида: PaleoPress. стр. 100-1 1–159. ISBN 978-0-9719477-0-2 . OCLC 52125833 .
- ^ Портелл, Роджер; Хьюбелл, Гордон; Донован, Стивен; Грин, Джереми; Харпер, Дэвид ; Пикерилл, Рон (2008). «Миоценовые акулы в формациях Кендис и Гранд-Бэй на Карриаку, Гренадины, Малые Антильские острова» (PDF) . Карибский научный журнал . 44 (3): 279–286. дои : 10.18475/cjos.v44i3.a2 . S2CID 87154947 . Архивировано из оригинала (PDF) 20 июля 2011 года.
- ^ Готфрид, доктор медицины; Компаньо, LJV; Боуман, Южная Каролина (1996). «Размер и анатомия скелета гигантской большезубой акулы Carcharodon megalodon ». В Климли; Эйнли (ред.). Большие белые акулы: биология Carcharodon carcharias . Сан-Диего, Калифорния: Academic Press. стр. 55–89. ISBN 978-0-12-415031-7 .
- ^ Готфрид, доктор медицинских наук, Фордайс Р.Э. (2001). «Связанный экземпляр Carcharodon angustidens (Chondrichthyes, Lamnidae) из позднего олигоцена Новой Зеландии с комментариями о взаимоотношениях Carcharodon ». Журнал палеонтологии позвоночных . 21 (4): 730–739. doi : 10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2 . S2CID 86092645 .
- ^ Андрес, Лутц. «Окаменелости мегазубов, найденные в скалах Калверт в Мэриленде» . Архивировано из оригинала 26 августа 2022 года . Проверено 10 сентября 2022 г.
- ^ Амальфитано, Якопо; Далла Веккья, Фабио Марко; Карнавал, Джорджио; Форначари, Элиана; Роги, Гвидо; Джусберти, Лука (10 мая 2022 г.). «Морфология и палеобиология позднемеловой крупной акулы Cretodus crassidens (Dixon, 1850) (Neoselachii; Lamniformes)» (PDF) . Журнал палеонтологии . 96 (5): 1166–1188. Бибкод : 2022JPal...96.1166A . дои : 10.1017/jpa.2022.23 . ISSN 0022-3360 . S2CID 248702856 .
- ^ Фредериксон, Дж.А.; Шефер, С.Н.; Дусетт-Фредериксон, JA (2015). «Гигантская акула из нижнемеловой формации Дак-Крик в Техасе» . ПЛОС ОДИН . 10 (6): e0127162. Бибкод : 2015PLoSO..1027162F . дои : 10.1371/journal.pone.0127162 . ПМЦ 4454486 . ПМИД 26039066 .
- ^ Кент, BW (1999). «Предположения о размере и морфологии вымершей ламноидной акулы Parotodus benedeni (le Hon)» (PDF) . Мозазавр . 6 :11–15.
- ^ Уорд, диджей; Кент, BW (2015). Новый гигантский вид акулы-молотилки из миоцена США (Отчет). Музей естественной истории. дои : 10.13140/RG.2.1.1723.0969 .
- ^ Пимьенто, Каталина; Канталапьедра, Хуан Л.; Симада, Кенсю; Филд, Дэниел Дж.; Смаерс, Йерун Б. (2019). «Эволюционные пути к гигантизму акул и скатов» . Эволюция . 73 (3): 588–599. дои : 10.1111/evo.13680 . ISSN 1558-5646 . ПМИД 30675721 . S2CID 59224442 .
- ^ Себастьян Штумпф, Фавиэль А. Лопес-Ромеро, Рене Киндлиманн, Фредерик Лакомба, Буркхард Пол, Юрген Кривет. Уникальный гибодонтиформный скелет дает новое представление о мезозойской жизни хондрихтиев. 2021. https://doi.org/10.5061/dryad.fqz612jr5
- ^ Майш, Майкл В.; Мацке, Андреас Т. (1 июня 2016 г.). «Новая акула-гибодонтид (Chondrichthyes, Hybodontiformes) из нижнеюрской формации Posidonienschiefer в Доттернхаузене, юго-запад Германии» . Новогодний ежегодник по геологии и палеонтологии-трактаты . 280 (3): 241–257. дои : 10.1127/njgpa/2016/0577 .
- ^ Кац, Б. (4 февраля 2020 г.). «Челюсть акулы возрастом 330 миллионов лет обнаружена в пещере Кентукки» .
- ^ Вильялобос-Сегура, Э.; Кривет, Дж.; Вулло, Р.; Штумпф, С.; Уорд, диджей; Андервуд, CJ (2021). «Скелетные останки эвригалинного склероринхоида † Onchopristis (Elasmobranchii) из среднего мела и их палеонтологическое значение». Зоологический журнал Линнеевского общества . 193 (2): 746–771. doi : 10.1093/zoolinnean/zlaa166 .
- ^ Перейти обратно: а б «Пилозубые левиафаны курсировали по древним морям» . Eartharchives.org . Архивировано из оригинала 22 августа 2021 года.
- ^ Перейти обратно: а б Stanislav Drobyshevsky (2022). Палеонтология антрополога. Книга 1. Докембрий и палеозой (Paleontology of anthropologist. Volume 1. Precambrian and Paleozoic) . Eksmo . ISBN 978-5-04-209523-8 . Проверено 31 августа 2022 г.
- ^ Тапанила, Л.; Прюитт, Дж.; Вилга, CD; Прадел, А. (2020). «Пилы, ножницы и акулы: позднепалеозойские эксперименты с симфизарным зубным рядом» . Анатомическая запись . 303 (2): 363–376. дои : 10.1002/ar.24046 . ПМИД 30536888 . S2CID 54478736 .
- ^ Перейти обратно: а б Кавин, Лайонел; Пьюз, Андре; Ферранте, Кристоф; Гино, Гийом (3 июня 2021 г.). «Гигантские мезозойские целаканты (Osteichthyes, Actinistia) демонстрируют значительную разницу в размерах тела, не связанную с таксическим разнообразием» . Научные отчеты . 11 (1): 11812. Бибкод : 2021NatSR..1111812C . дои : 10.1038/s41598-021-90962-5 . ISSN 2045-2322 . ПМЦ 8175595 . ПМИД 34083600 .
- ^ Пауло М. Брито; Дэйвид. М. Мартилль; Ян Ивз; Рой Э. Смит; Сэмюэл Э.А. Купер (июнь 2021 г.). «Морской целакант позднемелового (маастрихтского) периода из Северной Африки» . Меловые исследования . 122 : 104768. Бибкод : 2021CrRes.12204768B . дои : 10.1016/j.cretres.2021.104768 . S2CID 233551515 .
- ^ Симада, Кенсю; Киркланд, Джеймс И. (2011). «Загадочная мезозойская двоякодышащая рыба королевских размеров из Северной Америки». Труды Канзасской академии наук . 114 (1/2): 135–141. дои : 10.1660/062.114.0114 . ISSN 0022-8443 . JSTOR 41309634 . S2CID 84698238 .
- ^ Джеффри, Дж. Э. (2003). «Нижние челюсти ризодонтид: анатомия, функции и эволюция внутри стеблевой группы четвероногих» . Труды Королевского общества Эдинбурга: Науки о Земле . 93 (3): 255–276. дои : 10.1017/S0263593300000432 . S2CID 129517553 .
- ^ Джеффри, Джонатан Э. (1 сентября 2012 г.). «Краниальная морфология каменноугольного ризодонтида Screbinodus ornatus (Osteichthyes: Sarcopterygii)». Журнал систематической палеонтологии . 10 (3): 475–519. Бибкод : 2012JSPal..10..475J . дои : 10.1080/14772019.2011.595961 . ISSN 1477-2019 . S2CID 84810001 .
- ^ Холланд, Тимоти (14 декабря 2010 г.). «Останки остихтиана верхнего девона из реки Генуя, Виктория, Австралия» . Мемуары музея Виктории . 67 : 35–44. дои : 10.24199/j.mmv.2010.67.04 .
- ^ Хорнунг, Ян Дж.; Сакс, Свен (22 января 2020 г.). «Первая находка Gyrosteus mirabilis (Actinopterygii, Chondrosteidae) из тоара (нижняя юра) Балтийского региона» . ПерДж . 8 : е8400. дои : 10.7717/peerj.8400 . ISSN 2167-8359 . ПМК 6982408 . ПМИД 32002331 .
- ^ Несов, Л.А. (1997). Меловые неморские позвоночные Северной Евразии. СПб.: Институт земной коры СПбГУ, 218 с.
- ^ Гранде, Лэнс; Бемис, Уильям Э. (1991). «Остеология и филогенетические взаимоотношения ископаемых и современных веслоносов (Polyodontidae) с комментариями о взаимоотношениях Acipenseriformes» . Мемуары (Общество палеонтологии позвоночных) . 1 : II–121. дои : 10.2307/3889328 . ISSN 1062-161X . JSTOR 3889328 .
- ^ Листон Дж., Ньюбри М., Чалландс Т. и Адамс К., 2013, «Рост, возраст и размер юрской пахикормиды Leedsichthyspromaticus (Osteichthyes: Actinopterygii) в: Arratia, G., Schultze, Х. и Уилсон, М. (ред.) Мезозойские рыбы 5 – Глобальное разнообразие и эволюция . Верлаг, д-р Фридрих Пфайль, Мюнхен, Германия, стр. 145–175.
- ^ Мартилл, Д.М., 1988, « Leedsichthyspromaticus , гигантская костистая рыба-фильтратор из юрского периода Англии и Франции», Новогоднего справочника по геологии и палеонтологии Ежемесячные выпуски , 1988 (11): 670–680.
- ^ [Ред.] «Поймай 100-футовую рыбу», The Mirror , Лондон, Англия, 18 сентября 2003 г.
- ^ «В Мордене происходит крупное открытие окаменелостей» (PDF) . Канадский центр открытия ископаемых. 16 июля 2010 г. Пресс-релиз.
- ^ Эверхарт, Майкл Дж. (11 сентября 2017 г.). Океаны Канзаса: естественная история западного внутреннего моря . Издательство Университета Индианы. п. 110. ИСБН 978-0-253-02715-3 . Проверено 26 августа 2022 г.
- ^ Кривет, Юрген (2008). «Зубной ряд загадочной рыбы-пикнодонта Athrodon wittei (Fricke, 1876) (Neopterygii, Pycnodontiformes; поздняя юра; северо-запад Германии)» (PDF) . Ископаемый рекорд . 11 (2): 61–66. Бибкод : 2008FossR..11...61K . дои : 10.1002/mmng.200800002 .
- ^ Грандстафф, бакалавр наук; Смит, Дж. Б.; Ламанна, MC ; Лаковара, Кей Джей; Абдель-Гани, MS (2012). « Bawitius , gen. nov., гигантский полиптерид (Osteichthyes, Actinopterygii) из верхнемеловой формации Бахария в Египте» Журнал палеонтологии позвоночных . 32 (1): 17–26. Бибкод : 2012JVPal..32...17G . дои : 10.1080/02724634.2012.626823 . S2CID 140547157 .
- ^ Готфрид, Майкл Д.; Фордайс, Р. Юэн; Раст, Сиборн (2006). « Megalampris keyesi , гигантская рыба-луна (Teleostei, Lampridiformes) из позднего олигоцена Новой Зеландии» Журнал палеонтологии позвоночных . 6 (3): 544–551. doi : 10.1671/0272-4634(2006)26[544:MKAGMT]2.0.CO;2 . S2CID 85849900 .
- ^ Перейти обратно: а б Санки, Джулия; Бивер, Джейкоб; Басуга, Янис; Паласиос, Франциско; Вагнер, Хью (2016). «Гигантский зубастый лосось Oncorhynchus rastrosus и «река Прото-Туолумн» (ранний плиоцен) Центральной Калифорнии» . ПалеоБиос . 33 : 1–16. дои : 10.5070/P9331033123 .
- ^ Стирли, РФ и Г.Р. Смит. 2016. Лососевые рыбы из мио-плиоценовых озерных отложений на равнине Западной Снейк-Ривер и Большой Бассейн. в У. Л. Финке и Н. Карпентере (ред.). Рыбы мио-плиоценовой равнины и окрестностей Западной реки Снейк . Разное. Паб. Музейная зоология, Мичиганский университет 204:1–45.
- ^ Грегорова, Рузена; Шульц, Ортвин; Харцхаузер, Матиас; Кро, Андреас; Чорич, Степан (12 июня 2009 г.). «Гигантская солнечная рыба раннего миоцена из Северо-Альпийского предгорья (Австрия) и ее значение для филогении молд». Журнал палеонтологии позвоночных . 29 (2): 359–371. Бибкод : 2009JVPal..29..359G . дои : 10.1671/039.029.0201 . ISSN 0272-4634 . S2CID 54774567 .
- ^ Тройер, Эмили М.; Бетанкур-Р, Рикардо; Хьюз, Лили С.; Вестнит, Марк; Карневале, Джорджо; Уайт, Уильям Т.; Погоноски, Джон Дж.; Тайлер, Джеймс С.; Болдуин, Кэрол С.; Орти, Гильермо; Бринкворт, Эндрю; Клавель, Жюльен; Арсила, Дахиана (11 июля 2022 г.). «Влияние палеоклиматических изменений на эволюцию размеров тела морских рыб» . Труды Национальной академии наук . 119 (29): e2122486119. Бибкод : 2022PNAS..11922486T . дои : 10.1073/pnas.2122486119 . ISSN 0027-8424 . ПМЦ 9308125 . ПМИД 35858316 .
- ^ Виньола Лопес, Лазаро В.; Карр, Ричард; Лоренцо, Логель (20 октября 2020 г.). «Первое появление ископаемых балистов (Tetradontiformes: Balistidae) из миоцена Кубы с описанием нового вида и пересмотром ископаемых балистов». Историческая биология . 32 (9): 1290–1299. Бибкод : 2020HBio...32.1290V . дои : 10.1080/08912963.2019.1580278 . ISSN 0891-2963 . S2CID 92003143 .
- ^ Мишо, Марк (2012). «Stratodontidae, Маастрихтен-дю-Нигер, часть 1» . HAL Открытая наука . Архивировано из оригинала 27 мая 2022 года.
- ^ Гесс, Ганс (2010). «Онлайн-трактат, № 16, Часть T, переработанный, Том 1, Глава 19: Палеоэкология пелагических криноидей» . Трактат онлайн . дои : 10.17161/to.v0i0.4086 . ISSN 2153-4012 .
- ^ Блейк, Дэниел Б. (1 июня 2009 г.). «Переоценка девонского семейства Helianthasteridae Gregory, 1899 (Asteroidea: Echinodermata)». Палеонтологическая газета . 83 (2): 293–308. Бибкод : 2009PalZ...83..293B . дои : 10.1007/s12542-009-0020-x . ISSN 1867-6812 . S2CID 131480746 .
- ^ Дэвид К. Лойделл, Роберт Ф. Ловеридж (23 апреля 2001 г.). «Самый длинный граптолит в мире?». Геологический журнал . 36 (1): 55–57. Бибкод : 2001GeolJ..36...55L . дои : 10.1002/gj.875 . S2CID 129566263 .
- ^ Фу, Дунцзин, Дай, Лю, Юнин, Цуй, Линьхао; Юн, У, Юй; Вэньруй; Роберт Р.; Чжан, Синлян (22 марта 2019 г.). — ископаемое Лагерштетте типа сланцев Берджесс из раннего кембрия» . Science «Биота Цинцзян 363 (6433): 1338–1342 . : 363.1338F . doi : 10.1126 . ISSN 0036-8075 . PMID 30898931 . / science.aau8800 2019Sci ...
- ^ « В Китае обнаружена «умопомрачительная» партия окаменелостей возрастом более 500 миллионов лет» . Хранитель . 21 марта 2019 г.
- ^ Перейти обратно: а б Винтер, Джейкоб; Поррас, Луи; Янг, Флетчер Дж.; Бадд, Грэм Э.; Эджкомб, Грегори Д. (2016). «Ротовой аппарат кембрийского жаберного лобопода Pambdelurion Whittingtoni» . Палеонтология . 59 (6): 841–849. Бибкод : 2016Palgy..59..841V . дои : 10.1111/пала.12256 . hdl : 1983/16da11f1-5231-4d6c-9968-69ddc5633a8a . ISSN 1475-4983 . S2CID 88758267 .
- ^ Ван Рой, Питер; Дейли, Эллисон С.; Бриггс, Дерек Э.Г. (2015). «Гомология конечностей туловища аномалокаридид, выявленная гигантским фильтратором с парными створками» . Природа . 522 (7554): 77–80. Бибкод : 2015Natur.522...77V . дои : 10.1038/nature14256 . ISSN 1476-4687 . ПМИД 25762145 . S2CID 205242881 .
- ^ Лерози-Обрил, Руди; Пейтс, Стивен (14 сентября 2018 г.). «Новый радиодонт, питающийся взвесью, предполагает эволюцию микропланктивных животных в кембрийском макронектоне» . Природные коммуникации . 9 (1): 3774. Бибкод : 2018NatCo...9.3774L . дои : 10.1038/s41467-018-06229-7 . ISSN 2041-1723 . ПМК 6138677 . ПМИД 30218075 .
- ^ Бергстрем, Ян; Штюрмер, Вильгельм; Винтер, Герхард (1 июня 1980 г.). «Palaeoisopus, Palaeopantopus и Palaeothea, членистоногие-пикногониды из нижнедевонского сланца Хунсрюк, Западная Германия» . Палеонтологический журнал . 54 (1–2): 7–54. Бибкод : 1980PalZ...54....7B . дои : 10.1007/BF02985882 . S2CID 86746066 .
- ^ Андерсон, Лайалл И.; Пошманн, Маркус; Браукманн, Карстен (1998). «Об эмсских (нижнедевонских) членистоногих Рейнских Сланцевых гор: 2. Синзифосуриновая Willwerathia » . Палеонтологическая газета . 72 (3–4): 325–336. Бибкод : 1998PalZ...72..325A . дои : 10.1007/BF02988363 . ISSN 0031-0220 . S2CID 128464147 .
- ^ Перейти обратно: а б Ламсделл, Джеймс К. (1 января 2013 г.). «Пересмотренная систематика палеозойских «мечехвостов» и миф о монофилетической меченосце» . Зоологический журнал Линнеевского общества . 167 (1): 1–27. дои : 10.1111/j.1096-3642.2012.00874.x . ISSN 0024-4082 .
- ^ Эджкомб, Грегори Д. (25 ноября 1998 г.). «Ранние многоногие членистоногие из Австралии: Мальдибулакия из девона Нового Южного Уэльса» . Записи Австралийского музея . 50 (3): 293–313. дои : 10.3853/j.0067-1975.50.1998.1288 .
- ^ Гайяр, Кристиан (1 декабря 2011 г.). «Гигантская тропа лимулидов (Kouphichnium Lithographicum) из литографических известняков Серина (поздний кимеридж, Франция): этологическое и экологическое значение» . Швейцарский журнал геонаук . 104 (1): 57–72. дои : 10.1007/s00015-010-0032-2 . ISSN 1661-8734 . S2CID 140543244 .
- ^ Ламсделл, Джеймс К.; Гандерсон, Джеральд О.; Мейер, Рональд К. (8 января 2019 г.). «Обыкновенное членистоногое из позднеордовикского периода Big Hill Lagerstätte (Мичиган) обнаруживает неожиданное экологическое разнообразие внутри Chasmataspidida» . Эволюционная биология BMC . 19 (1): 8. Бибкод : 2019BMCEE..19....8L . дои : 10.1186/s12862-018-1329-4 . ISSN 1471-2148 . ПМК 6325806 . ПМИД 30621579 .
- ^ Брэдди, Саймон Дж; Пошманн, Маркус; Тетли, О. Эрик (23 февраля 2008 г.). «Гигантский коготь обнаружил самое большое членистоногое в истории » Письма по биологии . 4 (1): 106–109. дои : 10.1098/rsbl.2007.0491 . ПМЦ 2412931 . ПМИД 18029297 .
- ^ Перейти обратно: а б с Ламсделл, Джеймс К.; Брэдди, Саймон Дж. (2009). «Правило Коупа и теория Ромера: закономерности разнообразия и гигантизма у эвриптерид и палеозойских позвоночных» . Письма по биологии . 6 (2): 265–9. дои : 10.1098/rsbl.2009.0700 . ПМК 2865068 . ПМИД 19828493 . Дополнительная информация . Архивировано из оригинала 28 февраля 2018 года . Проверено 5 сентября 2022 г.
- ^ Перейти обратно: а б Ламсделл, Джеймс К.; Бриггс, Дерек Э.Г.; Лю, Хуайбао; Витцке, Брайан Дж.; Маккей, Роберт М. (1 сентября 2015 г.). «Самый старый описанный эвриптерид: гигантский среднеордовикский (дарривильский) мегалограптид из Виннешик-Лагерштетте в Айове» . Эволюционная биология BMC . 15 (1): 169. Бибкод : 2015BMCEE..15..169L . дои : 10.1186/s12862-015-0443-9 . ПМК 4556007 . ПМИД 26324341 .
- ^ Джерам, Эндрю Дж. (1993). «Скорпионы из Визеа Восточного Кирктона, Западный Лотиан, Шотландия, с ревизией инфраотряда Mesoscorpionina» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 84 (3–4): 283–299. Бибкод : 1993EETR..84..283J . дои : 10.1017/S0263593300006106 . ISSN 1755-6929 . S2CID 131416804 .
- ^ Кьеллесвиг-Варинг, Эрик Н. (1972). «Brontoscorpio anglicus: гигантский нижнепалеозойский скорпион из Центральной Англии». Журнал палеонтологии . 46 (1): 39–42. ISSN 0022-3360 . JSTOR 1302906 .
- ^ Кьеллесвиг-Варинг, EN (1986). «Повторное исследование ископаемых скорпионов мира» . Палеонтография Американа . 55 : 1–287.
- ^ Селден, Пенсильвания; Ши, СК; Рен, Д. (2011). «Золотой паук-круготкач (Araneae: Nephilidae: Nephila ) из средней юры Китая» . Письма по биологии . 7 (5): 775–8. дои : 10.1098/rsbl.2011.0228 . ПМК 3169061 . ПМИД 21508021 .
- ^ Селден, Пол А.; Да Коста Касадо, Фабио; Вианна Мескита, Мариса (2006). «Пауки-мигаломорфы (Araneae: Dipluridae) из нижнего мела Crato lagerstätte, бассейн Арарипе, северо-восток Бразилии: МЕЛОВЫЕ ПАУКИ ИЗ КРАТО, БРАЗИЛИЯ» . Палеонтология . 49 (4): 817–826. дои : 10.1111/j.1475-4983.2006.00561.x . hdl : 1808/8355 . S2CID 3845623 .
- ^ Дэвид М. Мартилл; Гюнтер Бехли; Роберт Ф. Ловеридж (2007). Ископаемые пласты Крато в Бразилии: окно в древний мир . Издательство Кембриджского университета. п. 124. ИСБН 978-1-139-46776-6 . Проверено 22 августа 2022 г.
- ^ Сантана, Уильям; Пиньейру, Эллиссон П.; Сильва, Тьяго Андраде; Пять, Дэниел (3 января 2024 г.). «Описание новой окаменелости Thelyphonida (Arachnida, Uropygi) и дальнейшая запись Cratosolpuga wunderlichi Selden в Селдене и Шире, 1996 г. (Arachnida, Solifugae) из формации Крато (апт/альб), бассейн Арарипе, Бразилия » ПерДж . 12 : е16670. дои : 10.7717/peerj.16670 . ISSN 2167-8359 . ПМЦ 10771091 . ПМИД 38188171 .
- ^ Джейсон А. Данлоп, Дэвид М. Мартилл. Первый паук-кнут (Arachnida: Amblypygi) и три новых паука-хлыста (Arachnida: Thelyphonida) из нижнемеловой формации Крато в Бразилии . Труды Королевского общества Эдинбурга: Науки о Земле, 92, 325–334, 2002.
- ^ Уэлен, Найл; Селден, Пол (2021). «Новый гигантский рицинулеид (Arachnida, Ricinulei) из Пенсильвании штата Иллинойс и идентификация нового, онтогенетически стабильного, диагностического признака» . Журнал палеонтологии . 95 (3): 601–612. Бибкод : 2021JPal...95..601W . дои : 10.1017/jpa.2020.104 .
- ^ Дж. А. Данлоп, К. Франерт и Дж. Макол. 2018. Гигантский клещ в меловом бирманском янтаре . Ископаемая запись 21: 285–290
- ^ Перейти обратно: а б Росслер, Ронни и Данлоп, Джейсон А. (1997). «Переописание крупнейшего паукообразного тригонотарбида - Kreischeria wiedei Geinitz 1882 из верхнего карбона Цвикау, Германия». Палеонтологический журнал . 71 (3–4): 237–245. Бибкод : 1997PalZ...71..237R . дои : 10.1007/BF02988493 . S2CID 129447249 .
- ^ Чжан, Маоинь; Лю, Ю; Хоу, Сянгуан; Ортега-Эрнандес, Хавьер; Май, Хуэйцзюань; Шмидт, Мишель; Мельцер, Роланд Р.; Го, Цзинь (2022). «Вентральная морфология нетрилобитных парнокопытных Retifacies ненормальной Hou, Chen & Lu, 1989, из раннекембрийской биоты Чэнцзян, Китай» . Биология . 11 (8): 1235. doi : 10.3390/biology11081235 . ISSN 2079-7737 . ПМК 9405172 . ПМИД 36009864 .
- ^ Перейти обратно: а б Николас Дж. Минтер, М. Габриэла Мангано и Жан-Бернар Карон (2011). «Скольжение по поверхности с помощью движения членистоногих Берджесс Шейл» . Труды Королевского общества B: Биологические науки . 279 (1733): 1613–1620. дои : 10.1098/рспб.2011.1986 . ПМЦ 3282348 . ПМИД 22072605 .
- ^ Гутьеррес-Марко, Хуан К.; Са, Артур А.; Гарсия-Беллидо, Диего К.; РАБО, Изабель; Валерио, Маноэль (2009). «Гигантские трилобиты и скопления трилобитов из ордовика Португалии». Геология . 37 (5): 443–446. Бибкод : 2009Geo....37..443G . дои : 10.1130/G25513A.1 .
- ^ Джордж Р. Макги-младший (12 ноября 2013 г.). Когда вторжение на сушу провалилось: наследие девонского вымирания . Издательство Колумбийского университета. ISBN 978-0-231-16057-5 .
- ^ «Волнение открытия» . Виртуальный музей Канады . Архивировано из оригинала 4 февраля 2012 года . Проверено 17 апреля 2006 г.
- ^ «На пляже Нортумберленда найдена самая крупная ископаемая многоножка в истории» . Новости Би-би-си . 21 декабря 2021 г. Проверено 21 декабря 2021 г.
- ^ Нил Дэвис; и др. (21 декабря 2021 г.). «Самое большое членистоногое в истории Земли: данные по недавно обнаруженным останкам артроплевры (серпуховская формация Стейнмор, Нортумберленд, Англия)» . Журнал Геологического общества . 179 (3). дои : 10.1144/jgs2021-115 . S2CID 245401499 .
- ^ Дзик, Ежи (2008). «Строение жабр и взаимоотношения триасовых циклоидных ракообразных» (PDF) . Журнал морфологии . 269 (12): 1501–1519. дои : 10.1002/jmor.10663 . ПМИД 18690662 . S2CID 17617567 .
- ^ Эмерсон, MJ; Шрам, Франция (1991). «Ремипедия, часть 2: палеонтология» . Труды Общества естественной истории Сан-Диего . 7 :1–52.
- ^ Ремипедия: Виды — робустус, Годзиллиус . Crustacea.net (2 октября 2002 г.)
- ^ Шааль, Стефан (27 января 2006 г.). «Мессель». ЭЛС . дои : 10.1038/npg.els.0004143 . ISBN 978-0-470-01617-6 .
- ^ Энгель, М.С. (2006). «Гигантская медоносная пчела Apislithohermaea Engel из миоцена Японии и геологическая история Apis (Hymenoptera: Apidae)» . Наука о медоносных пчелах – Университет Тамагава (Япония) (на японском языке). ISSN 0388-2217 .
- ^ Арчибальд, С.Б.; Расницын, АП (2015). «Новые раннеэоценовые Siricomorpha (Hymenoptera: Symphyta: Pamphiliidae, Siricidae, Cephidae) из высокогорья Оканаган, западная часть Северной Америки». Канадский энтомолог . 148 (2): 209–228. дои : 10.4039/tce.2015.55 . S2CID 85743832 .
- ^ Гао, Тайпин; Ши, Чунгкун; Расницын Александр П.; Рен, Донг (3 мая 2013 г.). «Hoplitolyda duolunica gen. et sp. nov. (Insecta, Hymenoptera, Praesiricidae), крупнейший на сегодняшний день пилильщик из мезозоя Китая» . ПЛОС ОДИН . 8 (5): е62420. Бибкод : 2013PLoSO...862420G . дои : 10.1371/journal.pone.0062420 . ISSN 1932-6203 . ПМЦ 3643952 . ПМИД 23671596 .
- ^ Чжан, Яньцзе; Ши, Чунгкун; Расницын, Александр; Рен, Донг; Гао, Тайпин (2020). «Новая блоха из раннего мела Китая» . Acta Palaeontologica Polonica . 65 . дои : 10.4202/app.00680.2019 . ISSN 0567-7920 . S2CID 213622677 .
- ^ Перейти обратно: а б «Лабидура» . Естественная история острова Святой Елены и Вознесения . Архивировано из оригинала 17 июля 2011 года . Проверено 13 октября 2022 г.
- ^ Уокер, Мэтт (17 ноября 2014 г.). «Самая большая в мире уховертка объявлена вымершей» . BBC Земля . Архивировано из оригинала 1 мая 2017 года . Проверено 13 октября 2022 г.
- ^ Ф. Э. Цойнер. 1962. Субфоссильный гигантский дермаптерон с острова Святой Елены. Труды Лондонского зоологического общества 138: 651–653.
- ^ «Ископаемое с идентификационным номером: 1780 – Chresmoda obscura ГЕРМАР, 1839 – Окаменелости Зольнхофена» . Solnhofen-fossilienatlas.de . Архивировано из оригинала 20 августа 2022 года . Проверено 13 октября 2022 г.
- ^ Шалюмо, Ф.; Брошье, Б. (2001). «Новая ископаемая форма Chiasognathinae: Protognathinus spielbergi (Coleoptera, Lucanidae)» (PDF) . Ламбиллионея . 101 : 593–595. Архивировано (PDF) из оригинала 20 сентября 2022 года.
- ^ Бретт Рэтклифф; Дена М. Смит; Дайан Мари Эрвин (2005). «Oryctoantiquus borealis, новый род и вид из эоцена Орегона, США, старейшая в мире ископаемая династия и крупнейший ископаемый скарабеид (Coleoptera: Scarabaeidae: Dynastinae)» . Бюллетень колеоптерологов . 59 : 127–135. doi : 10.1649/0010-065X(2005)059[0127:OBNGAS]2.0.CO;2 . S2CID 18230496 .
- ^ Пак Тэ-Юн С.; Ким, До Юн; Нам, Ги-Су; Ли, Миринаэ (6 мая 2022 г.). «Новый титаноптеран Magnatitan jongheoni n. gen. n. sp. с юго-запада Корейского полуострова» . Журнал палеонтологии . 96 (5): 1111–1118. Бибкод : 2022JPal...96.1111P . дои : 10.1017/jpa.2022.30 . ISSN 0022-3360 . S2CID 248592776 .
- ^ Маккеун, Кейт К. (15 мая 1937 г.). «Новые крылья ископаемых насекомых (Protohemiptera, семейство Mesotitanidae)» . Записи Австралийского музея . 20 (1): 31–37. дои : 10.3853/j.0067-1975.20.1937.565 . ISSN 0067-1975 . [ постоянная мертвая ссылка ]
- ^ Бечли, Г.; Макаркин В.Н. (2016). «Новый гигантский вид златоглазки (Insecta: Neuroptera) из нижнего мела Бразилии подтверждает появление Kalligrammatidae в Америке». Меловые исследования . 58 : 135–140. Бибкод : 2016CrRes..58..135B . дои : 10.1016/j.cretres.2015.10.014 .
- ^ Селлардс, Элиас Ховард (1 апреля 1903 г.). «Некоторые новые особенности строения палеозойских тараканов» . Американский научный журнал . с4-15 (88): 307–315. Бибкод : 1903AmJS...15..307S . дои : 10.2475/ajs.s4-15.88.307 . ISSN 0002-9599 .
- ^ Пасхальный день, Кэри Рэй (2004). Стратиграфия и палеонтология Кладбищенского холма (десмуанский-миссурийский период: верхний карбон), округ Колумбия, восточный Огайо (Диссертация). Университет штата Огайо. Архивировано из оригинала 20 сентября 2022 года . Проверено 20 августа 2022 г.
- ^ Шнайдер, Йорг; Шольце, Франк; Германн, Себастьян; Лукас, Спенсер (16 апреля 2021 г.). «ПОЗДНЕПЕНСИЛЬВАНСКАЯ ПРИБРЕЖНАЯ ФАУНА НАСЕКОМЫХ КАРЬЕРА КИННИ-БРИК, ИСКОПАЕМЫЕ БЕСПОЗВОНОЧНЫЕ И ПОЗВОНОЧНЫЕ, ЛАГЕРШТЕТ, НЬЮ-МЕКСИКО» . Бюллетень Американского музея естественной истории . 84 .
- ^ Род Ptiloteuthis Gabb: Notulae Naturae Академии естественных наук Филадельфии, вып. 9 . Академия естественных наук. ISBN 978-1-60483-009-5 .
- ^ Энгель, М.С.; Гросс, М. (2008). «Гигантский термит из позднего миоцена Штирии, Австрия (Isoptera)». Naturwissenschaften . 96 (2): 289–295. Бибкод : 2009NW.....96..289E . дои : 10.1007/s00114-008-0480-y . ПМИД 19052720 . S2CID 21795900 .
- ^ Ганд, Г.; Нел, АН; Флек, Г.; Гарруст, Р. (1 января 2008 г.). «Odonatoptera позднепермского бассейна Лодев (Insecta)» . Журнал иберийской геологии (на испанском языке). 34 (1): 115–122. ISSN 1886-7995 .
- ^ Чжан, Хайчунь; Чжэн, ДаРань; Ван, Бо; Фанг, Ян; Яржембовский, Эдмунд А. (1 мая 2013 г.). «Самый крупный известный донат в Китае: Сюфуа Чаой Чжан и Ван, род. и пр. ноябрь из средней юры Внутренней Монголии » Китайский научный бюллетень . 58 (13): 1579–1584. Бибкод : 2013ЧСБу..58.1579З дои : 10.1007/s11434-012-5567-3 . ISSN 1861-9541 . S2CID 95712438 .
- ^ Дереньокур, Изабель; Вапплер, Торстен; Андерсон, Джон; Бету, Оливье (2017). «Новое триадотипидное насекомое из позднего триаса Южной Африки» . Acta Palaeontologica Polonica . 62 . дои : 10.4202/app.00345.2017 . ISSN 0567-7920 . S2CID 115149987 .
- ^ Синищенкова, Нина (2012). «Самая большая ископаемая подёнка (Insecta: Ephemerida = Ephemeroptera) из верхнепермского местонахождения Исады, Север Европейской России» . Дальневосточный энтомолог .
- ^ Хуан, Цзяндун; Рен, Донг; Синищенкова Нина Д.; Ши, Чунгкун (2 ноября 2007 г.). «Новый род и вид Hexagenitidae (Insecta: Ephemeroptera) из формации Исянь, Китай» . Зоотакса . 1629 (1): 39–50. дои : 10.11646/zootaxa.1629.1.3 . ISSN 1175-5334 .
- ^ Дж. Кукалова-Пек. 1985. Эфемероидное жилкование крыльев на основе новых гигантских каменноугольных подёнок и основные морфологии, филогения и метаморфоз крыловидных насекомых (Insecta, Ephemerida) . Канадский журнал зоологии 63 (4): 933–955.
- ^ Срок, Павел; Станичек, Арнольд Х.; Бехли, Гюнтер (2 ноября 2015 г.). «Ревизия гигантского крыловидного насекомого Bojophlebia prokopi Kukalová-Peck, 1985 (Hydropaeoptera: Bojophlebiidae) из каменноугольного периода Чешской Республики, с первым кладистическим анализом ископаемых палеоптерообразных насекомых». Журнал систематической палеонтологии . 13 (11): 963–982. Бибкод : 2015JSPal..13..963S . дои : 10.1080/14772019.2014.987958 . ISSN 1477-2019 . S2CID 84037275 .
- ^ Кукалова-Пек, Ярмила; Ричардсон, Юджин С. (1983). «Новые Homoiopteridae (Insecta: Paleodictyoptera) с сочленениями крыльев из слоев верхнего карбона Мейзон-Крик, штат Иллинойс». Канадский журнал зоологии . 61 (7): 1670–1687. дои : 10.1139/z83-218 .
- ^ Браукманн, Карстен; СТАБО, КАРЛ (2007). «Подкруглое крыло насекомого из позднего карбона из Оснабрюка, Германия» . Клаусталер Геовиссеншафтен . 6 : 79–85.
- ^ Монтанья, Маттео; Хауг, Иоахим Т.; Страда, Лаура; Хауг, Кэролин; Фельбер, Маркус; Тинтори, Андреа (7 апреля 2017 г.). «Центральная нервная система и мышечные пучки сохранились у гигантского щетинистого хвоста возрастом 240 миллионов лет (Archaeognatha: Machilidae)» . Научные отчеты . 7 (1): 46016. Бибкод : 2017НатСР...746016М . дои : 10.1038/srep46016 . ISSN 2045-2322 . ПМК 5384076 . ПМИД 28387236 .
- ^ Хоэлл, Х.В.; Дойен, Дж. Т. и Перселл, А. Х. (1998). Введение в биологию и разнообразие насекомых (2-е изд.). Издательство Оксфордского университета. п. 321. ИСБН 0-19-510033-6 .
- ^ Шир, Уильям А.; Кукалова-Пек, Ярмила (15 февраля 2011 г.). «Экология палеозойских наземных членистоногих: ископаемые свидетельства» . Канадский журнал зоологии . 68 (9): 1807–1834. дои : 10.1139/z90-262 .
- ^ Стокар, Рудольф. «Первая мезозойская запись о вымерших аптериготных насекомых рода Dasyleptus (Insecta: Archaeognatha: Monura: Dasyleptidae) из триаса Монте-Сан-Джорджио (Швейцария)» . Палеразнообразие . 4 .
- ^ Станичек, Арнольд Х.; Срок, Павел; Бехли, Гюнтер (2014). «Ни чешуйница, ни домашняя птица: загадочная каменноугольная Carbotriplura kukalovae Kluge, 1996 (Insecta: Carbotriplurida) — предполагаемая ископаемая сестринская группа крылатых насекомых (Insecta: Pterygota)» . Систематическая энтомология . 39 (4): 619–632. Бибкод : 2014SysEn..39..619S . дои : 10.1111/syen.12076 . S2CID 83792881 .
- ^ Пак Тэ-Юн С.; Нильсен, Мортен Лунде; Парри, Люк А.; Соренсен, Мартин Винтер; Ли, Миринаэ; Ким, Джи-Хун; Ан, Инхе; Парк, Чанкунь; Виво, Джачинто; Смит, М. Пол; Харпер, Дэвид А.Т.; Нильсен, Арне Т.; Винтер, Джейкоб (2024). «Гигантский хетогнат стеблевой группы» . Достижения науки . 10 (1):eadi6678. Бибкод : 2024SciA...10I6678P . дои : 10.1126/sciadv.adi6678 . ISSN 2375-2548 . ПМЦ 10796117 . ПМИД 38170772 .
- ^ « Гигантские» черви-хищники возрастом более полумиллиарда лет обнаружены в Северной Гренландии . Бристольский университет . 3 января 2024 г.
- ^ Перейти обратно: а б Эрикссон, Матс Э.; Парри, Люк А.; Рудкин, Дэвид М. (февраль 2017 г.). «Старейший на Земле «червь Боббит» - гигантизм у девонской полихеты-евникиды» . Научные отчеты . 7 (2): 239–260. Бибкод : 2017NatSR...743061E . дои : 10.1038/srep43061 . ПМК 5318920 . ПМИД 28220886 .
- ^ Цюрих, ETH-Библиотека. «Гигантские брюхоногие моллюски рода Campanile из карибского эоцена» . Электронная периодика (на немецком языке). Архивировано из оригинала 15 июня 2022 года.
- ^ Церитий гигантский или колокольчик гигантский . Архивная копия от 28 июля 2016 г.
- ^ Доминичи, Стефано; Форнасьеро, Мариагабриелла; Джусберти, Лука (14 декабря 2020 г.). «Самый крупный из известных каури и итеративная эволюция гигантских брюхоногих моллюсков» . Научные отчеты . 10 (1): 21893. Бибкод : 2020NatSR..1021893D . дои : 10.1038/s41598-020-78940-9 . ISSN 2045-2322 . ПМЦ 7736312 . ПМИД 33318588 .
- ^ Весселинг Ф.П. (2006). «Моллюски из миоценовой формации Пебас Перуанской и Колумбийской Амазонии». Scripta Geologica 133 : 19–290. Рисунок 274. аннотация , PDF
- ^ Журнал палеонтологии, январь 2007 г., Том. 81, № 1 (январь 2007 г.), стр. 64–81.
- ^ "Verdens største musling" . Архивировано из оригинала 18 августа 2022 года.
- ^ Асато, Кайто; Касе, Томоки; Оно, Теруо; Сашида, Кацуо; Агемацу, Сатико (2017). «Аберрантные пермские двустворчатые моллюски (Alatoconchidae: Ambonychioidea) из Японии» . Палеонтологические исследования . 21 (4): 358–379. дои : 10.2517/2017PR002 . ISSN 1342-8144 . S2CID 135126528 .
- ^ «Информационный бюллетень Музея окаменелостей может опубликовать № 25, посвященный окаменелостям крупных ракушек с горы Киншо» (PDF) . Музей окаменелостей Цзиньшэншань .
- ^ ЧИНЗЕЙ, К. (1986). «Строение раковины, рост и функциональная морфология удлиненной меловой устрицы» . Палеонтология . 29 (1): 139–154.
- ^ Перейти обратно: а б с Вермей, Гират Дж. (15 января 2016 г.). «Гигантизм и его последствия для истории жизни» . ПЛОС ОДИН . 11 (1): e0146092. Бибкод : 2016PLoSO..1146092V . дои : 10.1371/journal.pone.0146092 . ISSN 1932-6203 . ПМЦ 4714876 . ПМИД 26771527 .
- ^ Асато, Кайто; Касе, Томоки (2021). «Гигантские ладьевидные моллюски (Mollusca) из пермского известняка Акасака, центральная Япония» . Журнал палеонтологии . 95 (4): 748–762. Бибкод : 2021JPal...95..748A . дои : 10.1017/jpa.2021.3 . ISSN 0022-3360 . S2CID 233649005 .
- ^ Клюг, Кристиан; Баетс, Кеннет Де; Крёгер, Бьёрн; Белл, Марк А.; Корн, Дитер; Пейн, Джонатан Л. (2015). «Нормальные гиганты? Временные и широтные сдвиги палеозойского гигантизма морских беспозвоночных и глобальные изменения» . Летайя . 48 (2): 267–288. Бибкод : 2015Лета..48..267К . дои : 10.1111/лет.12104 . ISSN 1502-3931 .
- ^ Пейн, Джонатан Л.; Бойер, Элисон Г.; Браун, Джеймс Х.; Финнеган, Сет; Ковалевский, Михал; Краузе, Ричард А.; Лайонс, С. Кэтлин; Макклейн, Крейг Р.; Макши, Дэниел В.; Новак-Готтшалл, Филип М.; Смит, Фелиса А.; Стемпьен, Дженнифер А.; Ван, Стив С. (6 января 2009 г.). «Двухфазное увеличение максимального размера жизни на протяжении 3,5 миллиардов лет отражает биологические инновации и экологические возможности» . Труды Национальной академии наук . 106 (1): 24–27. Бибкод : 2009PNAS..106...24P . дои : 10.1073/pnas.0806314106 . ISSN 0027-8424 . ПМК 2607246 . ПМИД 19106296 .
- ^ Тейхерт, К. и Б. Куммель 1960. Размер эндоцероидных головоногих моллюсков . Музей сравнительной зоологии Бревиоры 128 : 1–7.
- ^ Ифрим, Кристина; Стиннесбек, Вольфганг; Гонсалес, Артуро Х. Гонсалес; Шорндорф, Нильс; Гейл, Эндрю С. (10 ноября 2021 г.). «Онтогенез, эволюция и палеогеографическое распространение крупнейшего в мире аммонита Parapuzosia (P.) seppenradensis (Landois, 1895)» . ПЛОС ОДИН . 16 (11): e0258510. Бибкод : 2021PLoSO..1658510I . дои : 10.1371/journal.pone.0258510 . ISSN 1932-6203 . ПМЦ 8580234 . ПМИД 34758037 .
- ^ Иба, Ясухиро; Сано, Син-Ичи; Гото, Мичихару (январь 2015 г.). «Крупные белемниты уже были распространены в ранней юре — новые данные из Центральной Японии». Палеонтологические исследования . 19 (1): 21–25. дои : 10.2517/2014PR025 . S2CID 55001872 .
- ^ Фукс, Дирк; Иба, Ясухиро; Хейнг, Александр; Иидзима, Масая; Клюг, Кристиан; Ларсон, Нил Л.; Швайгерт, Гюнтер (2020). Брайард, Арно (ред.). «Muensterelloidea: филогения и эволюция характера мезозойских стволовых осьминогов» . Статьи по палеонтологии . 6 (1): 31–92. Бибкод : 2020PPal....6...31F . дои : 10.1002/spp2.1254 . ISSN 2056-2802 . S2CID 198256507 .
- ^ Танабэ, Казусигэ; Хикида, Ёсинори; Иба, Ясухиро (2006). «Две колеоидные челюсти из верхнего мела Хоккайдо, Япония» . Журнал палеонтологии . 80 (1): 138–145. doi : 10.1666/0022-3360(2006)080[0138:TCJFTU]2.0.CO;2 . ISSN 0022-3360 . S2CID 130583140 .
- ^ Танабэ, Казусигэ; Мисаки, Акихиро; Убуката, Такао (2014). «Позднемеловые находки крупных мягкотелых колеоидов на основе остатков нижней челюсти с Хоккайдо, Япония» . Acta Palaeontologica Polonica . дои : 10.4202/app.00052.2013 . ISSN 0567-7920 .
- ^ Мычко, Эдуард Васильевич; Фельдман, Родни М.; Швейцер, Кэрри Э.; Алексеев Александр Сергеевич (25 октября 2019 г.). «Новый род Cyclida (Crustacea) из нижнего карбона (Миссисипи, Визе) России и Англии и новые виды из Визеа России» . Новогодний ежегодник по геологии и палеонтологии-трактаты . 294 : 81–90. дои : 10.1127/njgpa/2019/0847 . S2CID 210640676 .
- ^ Джордж Р. МакГи-младший (2019). Конвергентная эволюция на Земле. Уроки поиска внеземной жизни . МТИ Пресс. п. 47. ИСБН 9780262354189 . Проверено 23 августа 2022 г.
- ^ Войтех Турек; Дж. Марек; Йозеф Бенеш (1989). Окаменелости мира. Комплексное практическое руководство по сбору и изучению окаменелостей . Арч Кейп Пресс. п. 146. ИСБН 9780517679043 . Проверено 23 августа 2022 г.
- ^ Хелен Маргарита Мьюир-Вуд, Густав Артур Купер (1960). Морфология, классификация и образ жизни Productoidea (Brachiopoda) . Геологическое общество Америки. п. 334. ИСБН 9780813710815 . Проверено 23 августа 2022 г.
- ^ Пил, Джон С.; Йохельсон, Эллис Л. (1984). «Пермские Toxeumorphorida из Гренландии: оценка класса моллюсков Xenoconchia» . Летайя . 17 (3): 211–221. Бибкод : 1984Лета..17..211П . дои : 10.1111/j.1502-3931.1984.tb01619.x . ISSN 0024-1164 .
- ^ Майор, Эдвард; Линан, Эладиус; Винтанед, Хосе Антонио Гамес; Мунис, Фернандо; Гозало, Родольфо (3 марта 2021 г.). «Выброшенная на мель медуза в самом нижнем кембрии (Кордубане) Испании » Испанский журнал палеонтологии . 19 (2): 191–198. дои : 10.7203/sjp.19.2.20531 . ISSN 2660-9568 . S2CID 126508499 .
- ^ Том Кларк. Желе откатывают время вспять . Природа (2002).
- ^ М. ЛАФЛАММ, ГМ НАРБОННА, К. ГРИНТРИ И ММ АНДЕРСОН. 2016. Морфология и тафономия эдиакарской ветви: Чарния с полуострова Авалон в Ньюфаундленде . Департамент геологических наук и геологической инженерии, Королевский университет, Кингстон, Онтарио, Канада K7L 3N6. Архивная копия от 18 августа 2022 года.
- ^ Данн, Фрэнсис С.; Уилби, Филип Р.; Кенчингтон, Шарлотта Г.; Гражданкин Дмитрий В.; Донохью, Филип CJ; Лю, Александр Г. (2019). «Анатомия эдиакарской рангеоморфы Charnia masoni » . Статьи по палеонтологии . 5 (1): 157–176. Бибкод : 2019PPal....5..157D . дои : 10.1002/spp2.1234 . ПМК 6472560 . ПМИД 31007942 .
- ^ Дженкинс, RJF (1992). «Функциональные и экологические аспекты эдиакарских комплексов» . В Липпсе, Дж.; Синьор, PW (ред.). Происхождение и ранняя эволюция Metazoa . Нью-Йорк, штат Нью-Йорк: Спрингер. стр. 131–176. ISBN 978-0-306-44067-0 . OCLC 231467647 .
- ^ Федонкин М.А.; Гелинг Дж.Г.; Грей К.; Нарбонн ГМ; Викерс-Рич П. (2007). Восстание животных. Эволюция и разнообразие царства животных . Издательство Университета Джонса Хопкинса. п. 326. ИСБН 978-0-8018-8679-9 .
- ^ Ригби, Дж. Кейт; Сеновбари-Дарьян, Баба (1996). «Gigantospongia, новый род, самая крупная известная пермская губка, капитанский известняк, горы Гуадалупе, Нью-Мексико». Журнал палеонтологии . 70 (3): 347–355. Бибкод : 1996JPal...70..347R . дои : 10.1017/S0022336000038294 . ISSN 0022-3360 . JSTOR 1306431 . S2CID 130222477 .
Источники
[ редактировать ]- Роуз, Кеннет Дэвид (2006). Начало эпохи млекопитающих . Балтимор: JHU Press. ISBN 978-0-8018-8472-6 .
- Дезохо, Джулия Бренда (2013). Анатомия, филогения и палеобиология ранних архозавров и их родственников . Лондон: Геологическое общество. ISBN 978-1-86239-361-5 .