Дирозавриды
| Дирозавриды | |
|---|---|
| |
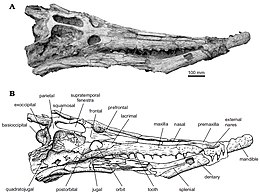
| Череп дирозаврида Arambourgisuchus khouribgaensis | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Рептилии |
| Клэйд : | Архозаврия |
| Клэйд : | Псевдозухия |
| Клэйд : | Крокодиломорфа |
| Клэйд : | Крокодилиформные |
| Подотряд: | † Тетисуха |
| Семья: | † Дирозавриды де Стефано, 1903 год. |
| Роды | |
| |
Dyrosauridae — семейство вымерших , неозуховых крокодилиформ живших от кампана до эоцена . Окаменелости дирозавридов распространены по всему миру: они были найдены в Африке, Азии, Европе, Северной Америке и Южной Америке. В настоящее время известно более дюжины видов, сильно различающихся по общему размеру и форме черепа. Большинство из них были водными, некоторые наземными, а другие полностью морскими (см. передвижение ниже), причем виды обитали как в пресноводной, так и в морской среде. Обитавшие в океане дирозавриды были одними из немногих морских рептилий, переживших мел-палеогеновое вымирание .
Дирозавриды представляли собой группу преимущественно морских четвероногих, похожих на крокодилов, с длинной челюстью и длиной до 6 метров (20 футов). [1] Самым крупным дирозавридом, вероятно, был фосфатозавр, длина которого оценивалась в 9 м (30 футов). [2] [3] На основании данных о костной ткани была выдвинута гипотеза, что они были медленно растущими. [4] прибрежные морские животные со сомкнутыми челюстями, [4] умеют плавать и ходить по суше. Внешние ноздри на заднем конце морды и внутренняя ноздря на крыловидной кости указывали на привычку охотиться во время плавания, при этом верхняя часть головы находилась над водой, что позволяло ему дышать во время преследования добычи. [1]
Обзор
[ редактировать ]
Известно, что дирозавриды имеют очень характерную форму черепа с длинной и тонкой мордой, составляющей примерно 68% от общей длины черепа. Самая передняя часть черепа и морды — это наружная ноздря, за которой сзади следуют две предчелюстные кости, пока они не достигают двух верхнечелюстных костей, разделенных одной носовой костью . [1]
Типичным для дирозавридов является единственный носовой элемент с характерным набором мелких ямок и постоянной шириной, пока он не расширяется до контакта с носом. слезные кости, затем на небольшом расстоянии сужаются, пока не достигнут границы лобных и префронтальных костей. [1]

У дирозавров есть предчелюстная кость с неглубокими ямками, простирающимися кзади от третьих верхнечелюстных альвеол. Есть две предчелюстные кости, узкие по сравнению с верхнечелюстными костями и продолжающиеся в двух длинных верхнечелюстных костях, разделенных единственной носовой костью. Последняя предчелюстная и первая верхняя челюсти широко разделены четвертым зубным зубом. Альвеолы широко расставлены спереди и пространство между ними уменьшается кзади от пятнадцатой альвеолы, сохраняя постоянный диаметр. [1]
Верхняя челюсть длинная (примерно в два с половиной раза длиннее скуловой кости ) и образует большую часть латерального края черепа. [5] По данным Жува и Барбозы и, возможно, в зависимости от возраста животного, на каждой верхней челюсти имеется 13–19 зубов. [5]
Важной особенностью зубного ряда дирозавридов являются глубокие окклюзионные ямки, особенно в задней части верхних челюстей, которые становятся менее выраженными спереди. Ямки указывают на сомкнутую закрытую челюсть дирозаврид, потому что ямки дают возможность чередоваться верхней и нижней челюстям. Хотя они все еще присутствуют у dyrosaurus phosphaticus, эта особенность глубоких окклюзионных ямок становится менее развитой и широкой. [5] Зубы дирозавра гомодонтные, конические, длинные и тонкие, с асимметричной лабиальной и лингвальной поверхностями. Задние зубы короче и более сжаты, чем передние, что указывает на уменьшение размера зуба спереди по отношению к заднему.

У дирозавров длина морды до черепа составляет около 68%, а роды Rhabdognathus и Atlantosuchus , Dyrosaurus и Arambourgisuchus имеют самые большие пропорции морды среди всех дирозавридов. Длина морды ранее использовалась для установления взаимоотношений дирозавридов, при этом удлинение морды считалось «более развитым» признаком. Это не согласовывалось с выводом Жува, который заключался в том, что самая длинная морда на самом деле является примитивным состоянием, поэтому более короткая или длинная морда появляется независимо по крайней мере четыре раза в эволюции дирозавридов. [6]
Палеобиогеография
[ редактировать ]Дирозавриды когда-то считались африканской группой, но открытия, сделанные начиная с 2000-х годов, указывают на то, что они населяли большинство континентов. [7] Фактически, базальные формы позволяют предположить, что их колыбелью могла быть Северная Америка.
Общий
[ редактировать ]| Род | Статус | Возраст | Расположение | Описание | Изображения |
|---|---|---|---|---|---|
| Ахеронтисух | Действительный | Палеоцен | Крупнотелый и длиннорылый пресноводный дирозаврид из формации Серрехон. | ||
| Эгиалозух | Действительный | Кампанский период | Морской дирозаврид из бассейна Кристианстад. | ||
| Антракозух | Действительный | Палеоцен | Короткомордый пресноводный дирозаврид из формации Серрехон. | ||
| Арамбургизух | Действительный | Палеоцен | Длиннорылый морской дирозаврид. | ||
| Атлантозух | Действительный | Палеоцен | Длиннорылый морской дирозаврид с самой длинной мордой по отношению к размеру тела среди всех дирозавридов. | ||
| Брахиозух | Действительный | Поздний мел | Длиннорылый и длиннорукий дирозаврид с самыми длинными руками среди всех дирозавридов. | ||
| Церрехонисух | Действительный | Палеоцен | Небольшой пресноводный дирозаврид с короткой мордой из формации Серрехон. | ||
| Ченанисух | Действительный | Маастрихт – Палеоцен | Род охватывает границу K – Pg. | 
| |
| Конгозавр | Действительный | Палеоцен | |||
| Дирозавр | Действительный | эоцен | Крупный морской дирозаврид с длинной мордой. | 
| |
| Гуаринизух | Действительный | Палеоцен | вероятный младший синоним Hyposaurus derbianus | 
| |
| Гипозавр | Действительный | Маастрихт – Палеоцен | Было названо пять видов, больше всех из всех родов дирозавров; род охватывает границу K – Pg. | ||
| Фосфатозавр | Действительный | эоцен | Крупный морской дирозаврид с длинной мордой, тупыми зубами и ложкообразным кончиком морды. | ||
| Рабдогнат | Действительный | Маастрихт – Палеоген | Крупный морской дирозаврид с длинной мордой; род охватывает границу K – Pg. | ||
| Сокотозавр | Младший синоним | Младший синоним гипозавра | |||
| Сокотозух | Действительный | Маастрихт | Длиннорылый морской дирозаврид. | ||
| Тилемсизух | Действительный | эоцен |
Филогения
[ редактировать ]Жув и др. (2005) диагностируют Dyrosauridae как кладу на основании следующих семи синапоморфий или общих признаков:
- Заднемедиальное крыло ретроартикулярного отростка дорсально расположено вентрально на ретроартикулярном отростке.
- Затылочные бугры маленькие
- Экзоципитальная часть в основном участвует в работе затылочного мыщелка.
- Надвисочные окна сильно удлинены в переднезаднем направлении.
- Симфиз примерно такой же ширины, как и высота
- Квадратоскуловая мышца в основном участвует в краниальном мыщелке, обеспечивая сочленение с челюстью.
- 4 предчелюстных зуба
Жув и др. 2020 г. представлен комплексный анализ взаимоотношений дирозавридов, показанный ниже. Обратите внимание, что бывшие дирозавриды Sabinosuchus и Fortignathus обнаружены как фолидозавриды и пейрозавриды соответственно. [8]
| Дирозавриды |
| ||||||||||||||||||
Палеобиология
[ редактировать ]Рост
[ редактировать ]
Доказательства полуводной жизни дирозавридов получены в результате тщательного анализа структуры костей. У водных четвероногих могут наблюдаться два типа структурной организации костей: остеопоротическая и пахиостотическая . Остеопоротическая кость является губчатой и пористой, тогда как пахиостотическая предполагает увеличение скелетной массы. Губчатая/пористая кость, такая как остеопороз, связана с более быстрым плаванием и лучшей маневренностью в воде из-за уменьшения количества костной ткани. Многие современные китообразные и морские черепахи имеют кости, пораженные остеопорозом, что позволяет им быть хорошими пловцами. Пахиостоз кости — это общее/локальное увеличение скелетной массы, которое может быть вызвано остеосклерозом (внутренним уплотнением кости), пахиостозом (гиперплазией компактных кортикальных слоев) или пахеосклерозом (сочетанием этих двух факторов). Исследования костей дирозавра, проведенные Рафаэлем Сезаром Лимой Педросо де Андраде и Джулианой Мансо Саяо, показали, что у этой семьи была остеопорозная костная ткань, что указывает на быстрое плавание, а также на некоторые остеосклероз , который является компонентом пахиостотической костной ткани. Остеопороз связан с полностью водным образом жизни, тогда как пахиостоз не является полностью водным, а связан с экологией быстрого плавания. Таким образом, дирозавры — полуводные быстрые пловцы, о чем свидетельствует их костная структура. Другим свидетельством прибрежного полуводного образа жизни является то, где находят окаменелости, часто в переходных морских отложениях. [4] - использование плавания на осевой частоте (которое используется в основном современными крокодилами) с большей волнообразностью движений и частотой хвоста из-за высокоразвитой мускулатуры, позволяющей делать более мощный толчок вперед. [4]
Дирозавриды имеют структуру тканей, указывающую на медленно растущее животное, что было установлено путем тщательного анализа правого бедра и левой большеберцовой кости . В левой большеберцовой кости кора имела пластинчатую зональную кость с пятью линиями задержки роста (ЛАГ), расположенными на расстоянии 300 мм друг от друга, а также четкую сосудистую сеть первичных остеонов, плотность которых уменьшалась по направлению к мембране (периостиально). В правом бедре позже появились двойные LAG и EFS, а также вторичные остеоны, возникающие в глубокой коре и губчатой оболочке. Такая модель роста тканей является общей характеристикой медленно растущих животных. [4]
среда обитания
[ редактировать ]Дирозавриды встречаются в переходных морских отложениях от позднего мела до нижнего эоцена. [4] Это семейство известно главным образом из отложений маастрихта в Нью-Джерси и пород от позднего мела до раннего палеогена из моря Тетис в северной и западной Африке. [4] Окаменелости также были обнаружены в палеоценовых и эоценовых слоях Пакистана, а также в Южной Америке, Бразилии, Индии, Южной Азии, а также в прибрежных районах. Обычно дирозавриды обнаруживаются в прибрежных и устьевых отложениях Северной Африки и Ближнего Востока, что подтверждает их существование как полуводных животных. [9]
Наибольшее таксономическое разнообразие Dyrosauridae имело в раннем палеогене , но похоже, что клада смогла получить более широкое и широкое географическое распространение в позднем мелу. Самые ранние находки дирозавридов находятся либо в Африке, либо вблизи нее, с фрагментарными находками из сеномана Судана и Португалии и несколькими другими находками в домаастрихтском и позднем меловом периоде в Египте. Позже, к маастрихту Северной Америки, летопись дирозавридов стала более полной, установив широкое распространение, которое, по-видимому, сохранялось на протяжении палеоцена и эоцена. [10]
Дирозавриды также были обнаружены в неморских отложениях. В северном Судане дирозавриды известны из речных отложений, что указывает на то, что они жили в речных условиях. [11] были обнаружены во внутренних отложениях Пакистана Кости неопределенных дирозавридов также . Некоторые дирозавриды, например, из формации Умм-Химар в Саудовской Аравии , обитали в устьевой зоне вблизи побережья. Недавно названные дирозавриды Cerrejonisuchus и Acherontisuchus были обнаружены в формации Серрехон на северо-западе Колумбии , которая, как полагают, представляет собой переходную морско-пресноводную среду, окруженную тропическими лесами, расположенными дальше от суши, чем устьевая среда формации Умм-Химар. [12] Cerrejonisuchus и Acherontisurus жили в неотропических условиях в то время, когда глобальные температуры были намного выше, чем сегодня. [13] [14]
Воспроизведение
[ редактировать ]В 1978 году было высказано предположение, что дирозавриды во взрослом состоянии жили в океане, но размножались во внутренних пресноводных средах. Останки мелких дирозавридов из Пакистана были интерпретированы как молодые особи. Их присутствие во внутренних отложениях рассматривалось как свидетельство того, что дирозавриды вылупились вдали от океана. [15] В нижнемэоценовом бассейне Улад-Абдун встречается очень мало молодых дирозавридов, но много взрослых особей такого же размера. Это способствовало предположению, что молодые особи жили в пресноводной среде, а взрослые особи — в морской. [16] Однако недавно крупные и полностью зрелые дирозавриды формации Серрехон показали, что некоторые дирозавриды всю свою жизнь прожили во внутренних средах, никогда не возвращаясь на побережье. [17]
Передвижение
[ редактировать ]Исследование Cerrejonisuchus предполагает, что этот род был более наземным, чем другие дирозавриды, а также показывает, что современные крокодилы не являются хорошими функциональными аналогами Dyrosauridae. [18]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Жув, Стефан (март 2005 г.). «Новое описание черепа Dyrosaurus phosphaticus (Thomas, 1893) (Mesoeucrocodylia: Dyrosauridae) из нижнего эоцена Северной Африки» . Канадский журнал наук о Земле . 42 (3): 323–337. Бибкод : 2005CaJES..42..323J . дои : 10.1139/e05-008 . ISSN 0008-4077 . Проверено 22 марта 2021 г.
- ^ Баффето, Э. (1978). «Дирозавриды (Crocodylia, Mesosuchia) из нижнеэоценовых фосфатов Туниса: Dyrosaurus , Rhabdognathus , Phosphatosaurus ». Средиземноморская геология . 5 (2): 237–256. дои : 10.3406/geolm.1978.1046 .
- ^ Мартин, Джереми Э.; Антуан, Пьер-Оливье; Перье, Винсент; Добро пожаловать, Жан-Лу; Метаис, Грегуар; Мариво, Лоран (04 июля 2019 г.). «Большой крокодилоид из олигоцена холмов Бугти, Пакистан» (PDF) . Журнал палеонтологии позвоночных . 39 (4): e1671427. Бибкод : 2019JVPal..39E1427M . дои : 10.1080/02724634.2019.1671427 . ISSN 0272-4634 . S2CID 209439989 .
- ^ Перейти обратно: а б с д и ж г де Андраде, Рафаэль Сезар Лима Педросо; Саяо, Джулиана Мансо (17 июля 2014 г.). «Палеогистология и выводы об образе жизни дирозаврида (Archosauria: Crocodylomorpha) из бассейна Параиба (северо-восток Бразилии)» . ПЛОС ОДИН . 9 (7): e102189. Бибкод : 2014PLoSO...9j2189A . дои : 10.1371/journal.pone.0102189 . ISSN 1932-6203 . ПМК 4102515 . ПМИД 25032965 .
- ^ Перейти обратно: а б с Барбоза, Хосе Антонио; Келлнер, Александр Вильгельм Армин; Виана, Мария Сомали Продажи (2008). «Новый крокодиломорф дирозавридов и свидетельства круговорота фауны при переходе K – P в Бразилии» . Труды Королевского общества B: Биологические науки . 275 (1641): 1385–1391. дои : 10.1098/rspb.2008.0110 . ПМК 2602706 . ПМИД 18364311 .
- ^ Жув, Стефан; Буя, Баади; Амагзаз, Мбарек (март 2008 г.). «Длиннорылый дирозаврид (Crocodyliformes, Mesoeucrocodylia) из палеоцена Марокко: филогенетические и палеобиогеографические последствия» . Палеонтология . 51 (2): 281–294. Бибкод : 2008Palgy..51..281J . дои : 10.1111/j.1475-4983.2007.00747.x .
- ^ Жув и др. (2008)
- ^ Жув, Стефан; де Мюизон, Кристиан; Сеспедес-Пас, Рикардо; Сосса-Соруко, Виктор; Нолл, Стефан (19 октября 2020 г.). «Лонгиростриновые крокодилиформы из Боливии и их эволюция на границе мела и палеогена» . Зоологический журнал Линнеевского общества . 192 (злаа081): 475–509. doi : 10.1093/zoolinnean/zlaa081 . ISSN 0024-4082 .
- ^ Ламанна, Мэтью С.; Смит, Джошуа Б.; Аттиа, Юсри С.; Додсон, Питер (10 сентября 2004 г.). «От динозавров до дирозавридов (Crocodyliformes): удаление постсеноманской (позднемеловой) находки Ornithischia из Африки» (PDF) . Журнал палеонтологии позвоночных . 24 (3): 764–768. doi : 10.1671/0272-4634(2004)024[0764:FDTDCR]2.0.CO;2 . ISSN 0272-4634 . S2CID 16525132 .
- ^ Хосла, Ашу; Сертич, Джозеф JW; Прасад, Гунтупалли В.Р.; Верма, Омкар (12 декабря 2009 г.). «Останки дирозавридов из межтрапповых отложений Индии и позднемеловое распространение дирозавридов». Журнал палеонтологии позвоночных . 29 (4): 1321–1326. Бибкод : 2009JVPal..29.1321K . дои : 10.1671/039.029.0416 . ISSN 0272-4634 . S2CID 130987967 .
- ^ Бюффето, Э.; Буссерт, Р.; Бринкманн, В. (1990). «Новая неморская фауна позвоночных в верхнем мелу северного Судана». Берлинские геонаучные трактаты . 120 : 183-202.
- ^ Гастингс, АК; Блох, Дж.И.; Кадена, EA; Харамилло, Калифорния (2010). «Новый небольшой короткорылый дирозаврид (Crocodylomorpha, Mesoeucrocodylia) из палеоцена северо-востока Колумбии» . Журнал палеонтологии позвоночных . 30 (1): 139–162. Бибкод : 2010JVPal..30..139H . дои : 10.1080/02724630903409204 . S2CID 84705605 .
- ^ Руководитель, Джей-Джей; Блох, Дж.И.; Гастингс, АК; Борке, младший; Кадена, EA; Эррера, ФА; Полли, PD; Харамилло, Калифорния (2009). «Гигантская змея-боид из неотропов палеоцена обнаруживает более высокие температуры за экваториальными температурами». Природа . 457 (7230): 715–717. Бибкод : 2009Natur.457..715H . дои : 10.1038/nature07671 . ПМИД 19194448 . S2CID 4381423 .
- ^ Канапо, Б. (2 февраля 2010 г.). «Исследователи УФ: родственник древнего крокодила, вероятный источник пищи для Титанобоа» . Новости Университета Флориды . Архивировано из оригинала 8 июня 2010 года . Проверено 3 февраля 2010 г.
- ^ Баффето, Э. (1978). «Останки крокодилов из эоцена Пакистана». Новогодний альбом по геологии и палеонтологии, Трактаты . 156 : 262-283.
- ^ Жув, Стефани; Барде, Натали; Джалиль, Нур-Эддин; Супербиола, Хавьер Переда; Буя, Баади и Амагзаз, Мбарек. 2008. Самый старый африканский крокодил: филогения, палеобиогеография и дифференциальная выживаемость морских рептилий на границе мелового и третичного периодов. Журнал палеонтологии позвоночных 28 (2): 409–421, июнь 2008 г.
- ^ Гастингс, АК; Блох, Дж.; Харамилло, Калифорния (2011). «Новый лонгиростриновый дирозаврид (Crocodylomorpha, Mesoeucrocodylia) из палеоцена северо-восточной Колумбии: биогеографические и поведенческие последствия для дирозавридов нового мира». Палеонтология . 54 (5): 1095–116. Бибкод : 2011Palgy..54.1095H . дои : 10.1111/j.1475-4983.2011.01092.x .
- ^ Скавеццони, Изауре; Фишер, Валентин (2021). «Посткраниальный скелет Cerrejonisuchus improcerus (Crocodyliformes: Dyrosauridae) и необычная анатомия дирозавридов» . ПерДж . 9 : е11222. дои : 10.7717/peerj.11222 . ПМЦ 8117932 . ПМИД 34026348 .
- Баффето, Э. (1985). Эволюция крокодилов. Вымершие животные – За науку, Париж 109 .
- Жув, С.; Буя, Б.; Амагзаз, М. (2008). «Длиннорылый дирозаврид (Crocodyliformes, Mesoeucrocodylia) из палеоцена Марокко: филогенетические и палеогеографические значения» . Палеонтология . 51 (2): 281–294. Бибкод : 2008Palgy..51..281J . дои : 10.1111/j.1475-4983.2007.00747.x .
- Янг, С.; Ярошен, М.; Буя, Б.; Амагзаз, М. (2005). «Новый крокодилиформный дирозаврид из палеоцена Марокко и филогенетический анализ Dyrosauridae». Acta Palaeontologica Polonica . 50 (3): 581–594.


