Гиганотозавр
| Гиганотозавр Временной диапазон: поздний мел ( сеноман ), | |
|---|---|
 | |
| Реконструированный скелет, возможно, с пропорционально слишком длинным черепом, Музей естественной истории Фернбанка. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Кархародонтозавриды |
| Племя: | † Гиганотозаврины |
| Род: | † Гиганотозавр Кориа и Сальгадо , 1995 г. [1] |
| Разновидность: | † Г. Каролинии |
| Биномиальное имя | |
| † Гиганотозавр Каролини Кориа и Сальгадо, 1995 г. | |
Гиганотозавр ( / ˌ ɡ ɪ ɡ ə ˌ n oʊ t ə ˈ s ɔː r ə s / GIG -ə- NOH -tə- SOR -es [2] ) — род теропод , динозавров- живших на территории современной Аргентины , в ранний сеноманский , позднего мела период примерно 99,6–95 миллионов лет назад. Образец голотипа был обнаружен в формации Канделерос в Патагонии в 1993 году и завершен почти на 70%. животное было названо Giganotosaurus carolinii В 1995 году ; Название рода переводится как «гигантская южная ящерица», а видовое название дано в честь первооткрывателя Рубена Каролини . Зубная кость , зуб и некоторые следы, обнаруженные до голотипа, позже были отнесены к этому животному. Этот род вызвал большой интерес и стал частью научных дискуссий о максимальных размерах динозавров-теропод.
Гиганотозавр был одним из крупнейших известных наземных хищников , но его точный размер было трудно определить из-за неполноты найденных к настоящему времени останков. Оценки наиболее полного экземпляра варьируются от длины от 12 до 13 м (от 39 до 43 футов), длины черепа от 1,53 до 1,80 м (от 5,0 до 5,9 футов) и веса от 4,2 до 13,8 т (от 4,6 до 15,2 тонны). тонн). Зубная кость, принадлежавшая предположительно более крупному человеку, была использована для экстраполяции длины 13,2 м (43 фута). Некоторые исследователи обнаружили, что животное крупнее тираннозавра , который исторически считался самым крупным тероподом, в то время как другие обнаружили, что они примерно равны по размеру, а оценки наибольшего размера гиганотозавра преувеличены. Череп был низким, с морщинистыми (грубыми и морщинистыми) носовыми костями и валикообразным гребнем на слезной кости перед глазом. Передняя часть нижней челюсти была уплощена и имела на конце выступающий вниз отросток (или «подбородок»). Зубы были сжаты вбок и имели зазубрины. Шея была сильной и грудной пояс пропорционально небольшой.
, член семейства Carcharodontosauridae , Гиганотозавр является одним из наиболее известных представителей группы, в которую входят другие очень крупные тероподы, такие как близкородственные Mapusaurus и Carcharodontosaurus . Считается, что гиганотозавр был гомеотермным (тип « теплокровности ») с метаболизмом между млекопитающим и рептилиями, что обеспечивало быстрый рост. Он был бы способен быстро смыкать челюсти, захватывать и сбивать добычу, нанося мощные укусы. «Подбородок», возможно, помогал противостоять стрессу при укусе добычи. Считается, что гиганотозавр был высшим хищником своей экосистемы и, возможно, питался молодыми динозаврами -зауроподами .
Открытие


В 1993 году аргентинский охотник за окаменелостями Рубен Дарио Каролини обнаружил большую берцовую кость (кость голени) динозавра-теропода , управляя багги для дюн в бесплодных землях недалеко от Вилья-Эль-Чокон , в провинции Неукен в Патагонии , Аргентина. Специалисты Национального университета Комауэ были отправлены на раскопки образца после того, как были уведомлены о находке. [3] [4] Об открытии объявили палеонтологи Родольфо Кориа и Леонардо Сальгадо на собрании Общества палеонтологии позвоночных в 1994 году, где научный писатель Дон Лессем предложил профинансировать раскопки после того, как его впечатлила фотография кости ноги. [3] [5] Частичный череп был разбросан на площади около 10 м2. 2 (110 кв. футов), посткраниальный скелет был разобран. Образец сохранил почти 70% скелета и включал большую часть позвоночного столба , грудные и тазовые пояса, бедренные кости, а также левые большеберцовую и малоберцовую кость. [6] [1]
В 1995 году этот экземпляр был предварительно описан Кориа и Сальгадо, которые сделали его голотипом нового рода и вида Giganotosaurus carolinii (части скелета в это время еще были замурованы в гипс). Родовое название происходит от древнегреческих слов gigas/ γίγας (что означает «гигант»), notos/ νότος (что означает «южный/южный», в зависимости от его происхождения) и -sauros/- σαύρος (что означает «ящерица»). Видовое название дано в честь Каролини, первооткрывателя. [1] [7] Скелет-голотип сейчас хранится в Палеонтологическом музее Эрнесто Бахмана (где он занесен в каталог как образец MUCPv-Ch1) на вилле Эль Чокон, который был открыт в 1995 году по просьбе Каролини. Этот образец является главной экспозицией музея и расположен на песчаном полу комнаты, посвященной животному, вместе с инструментами, использованными палеонтологами во время раскопок. В соседнем зале выставлена смонтированная реконструкция скелета. [4] [8]

Одной из особенностей тероподовых динозавров, вызвавшей наибольший научный интерес, является то, что в группу входят крупнейшие наземные хищники мезозойской эры . Этот интерес начался с открытия одного из первых известных динозавров, мегалозавра , названного в 1824 году из-за его большого размера. Более полувека спустя, в 1905 году, был назван тираннозавр , и он оставался крупнейшим известным динозавром-тероподом в течение 90 лет, хотя были известны и другие крупные тероподы. Дискуссия о том, какой теропод был самым крупным, возобновилась в 1990-х годах благодаря новым открытиям в Африке и Южной Америке . [1] В своем первоначальном описании Кориа и Сальгадо считали гиганотозавра как минимум самым крупным динозавром-тероподом южного полушария , а возможно, и самым крупным в мире. Они признали, что сравнение с тираннозавром было затруднительно из-за разрозненного состояния черепных костей гиганотозавра , но отметили, что при высоте 1,43 м (4,7 фута) бедренная кость гиганотозавра была на 5 см (2 дюйма) длиннее, чем у « Сью ». самый крупный известный экземпляр тираннозавра , и что кости гиганотозавра оказались более крепкими, что указывает на более тяжелое животное. По их оценкам, череп имел длину около 1,53 м (5 футов), длину всего животного — 12,5 м (41 фут) и вес от 6 до 8 т (от 6,6 до 8,8 коротких тонн). [1]
В 1996 году палеонтолог Пол Серено и его коллеги описали новый череп родственного рода Carcharodontosaurus из Марокко, теропода, описанного в 1927 году, но ранее известного только по фрагментарным останкам (большая часть его окаменелостей была уничтожена во время Второй мировой войны ). По их оценкам, череп имел длину 1,60 м (5 футов) и был похож на череп гиганотозавра , но, возможно, превосходил череп тираннозавра «Сью» с черепом длиной 1,53 м (5 футов). Они также отметили, что у кархародонтозавров были пропорционально большие черепа, но у тираннозавра, по-видимому, были более длинные задние конечности. [9] В интервью для статьи 1995 года, озаглавленной «Новый зверь узурпирует тиранозавра как короля-хищника», Серено отметил, что эти недавно открытые тероподы из Южной Америки и Африки конкурировали с тираннозавром как крупнейшими хищниками и помогут в понимании позднего мелового периода динозавров . фауны, которые в остальном были очень «ориентированы на Северную Америку». [10] В том же выпуске журнала, в котором был описан кархародонтозавр , палеонтолог Филип Дж. Карри предупредил, что еще предстоит определить, какое из двух животных было крупнее, и что размер животного менее интересен палеонтологам, чем размер животного. например, адаптации, отношения и распространение. Он также нашел примечательным то, что эти два животных были найдены с разницей в год и были близкими родственниками, несмотря на то, что были найдены на разных континентах. [11]

В интервью 1997 года Кориа оценил гиганотозавра длину от 13,7 (45 футов) до 14,3 (47 футов) м и вес от 8 до 10 т (от 8,8 до 11,0 коротких тонн), основываясь на новом материале, который был больше, чем у кархародонтозавра . Серено возразил, что было бы трудно определить диапазон размеров вида на основе нескольких неполных экземпляров, и оба палеонтологи согласились, что другие аспекты этих динозавров более важны, чем урегулирование «соревнования по размерам». [12] В 1998 году палеонтолог Хорхе О. Кальво частичную левую зубную кость (часть нижней челюсти), содержащую несколько зубов (MUCPv-95) и Кориа приписали гиганотозавру . Он был собран Кальво возле Лос-Канделерос в 1988 году (найден в 1987 году), который кратко описал его в 1989 году, отметив при этом, что он, возможно, принадлежал к новому таксону теропод . Кальво и Кориа обнаружили, что зубная кость идентична голотипу, но на 8% больше и составляет 62 см (24 дюйма). Хотя его задняя часть неполная, они предположили, что череп экземпляра-голотипа имел длину 1,80 м (6 футов), и оценили, что череп более крупного экземпляра имел длину 1,95 м (6,4 фута), самый длинный из них. череп любого теропода. [13] [14] [15]
В 1999 году Кальво отнес неполный зуб (MUCPv-52) к гиганотозавру ; этот экземпляр был обнаружен возле озера Эсекьель-Рамос-Мексиа в 1987 году А. Дельгадо и, следовательно, является первым известным ископаемым этого рода. Кальво далее предположил, что некоторые следы теропод и отдельные следы (которые он положил в основу ихнотаксона Abelichnus astigarrae в 1991 году) принадлежали гиганотозаврам , исходя из их большого размера. Самые большие дорожки имеют длину 50 см (20 дюймов) и темп 130 см (51 дюйм), а самые маленькие - 36 см (14 дюймов) в длину и темп 100 см (39 дюймов). Следы трехпалые (трехпалые), с большими и грубыми пальцами с заметными отпечатками когтей. Отпечатки цифр занимают большую часть длины следа, причем один след имеет тонкую пятку. Хотя следы были найдены на более высоком стратиграфическом уровне, чем основные окаменелости гиганотозавра , они были из тех же слоев , что и единственный зуб и некоторые динозавры- зауроподы , которые также известны из тех же слоев, что и гиганотозавр . [14]
Продолжение оценок размера
В 2001 году ученый-врач Фрэнк Сибахер предложил новый полиномиальный метод расчета оценки массы тела динозавров (с использованием длины, глубины и ширины тела) и обнаружил, что гиганотозавр весил 6,6 т (7,3 коротких тонны) (на основе первоначальная оценка длины 12,5 м (41 фут). [16] В своем описании черепной коробки гиганотозавра в 2002 году Кориа и Карри дали оценку длины черепа-голотипа в 1,60 м (5 футов) и рассчитали вес в 4,2 т (4,6 коротких тонны), экстраполируя из 520 мм (20 дюймов). ) окружность диафиза бедренной кости. В результате коэффициент энцефализации (показатель относительного размера мозга) составил 1,9. [6] В 2004 году палеонтолог Херардо В. Маццетта и его коллеги отметили, что, хотя бедренная кость голотипа гиганотозавра была больше, чем у «Сью», большеберцовая кость была на 8 см (3 дюйма) короче и составляла 1,12 м (4 фута). Они обнаружили, что экземпляр голотипа по размеру был равен тираннозавру и весил 8 т (8,8 коротких тонн) (немного меньше, чем «Сью»), но что более крупная зубная кость могла представлять собой животное весом 10 т (11 коротких тонн), если бы геометрически похож на образец голотипа. Используя уравнения многомерной регрессии , эти авторы также предложили альтернативный вес 6,5 т (7,2 коротких тонны) для голотипа и 8,2 т (9,0 коротких тонн) для более крупного экземпляра, и что последний, следовательно, был самым крупным известным наземным хищником. [17]

В 2005 году палеонтолог Криштиану Даль Сассо и его коллеги описали новый материал черепа (морду) спинозавра (исходные окаменелости которого также были уничтожены во время Второй мировой войны) и пришли к выводу, что этот динозавр имел длину от 16 до 18 м (от 52 до 59 м). футов) в длину и весом от 7 до 9 т (от 7,7 до 9,9 коротких тонн), что превышает максимальный размер всех других теропод. [18] В 2006 году Кориа и Карри описали большого теропода Mapusaurus из Патагонии; он был тесно связан с гиганотозавром и имел примерно такой же размер. [19] В 2007 году палеонтологи Франсуа Терриен и Дональд М. Хендерсон обнаружили, что гиганотозавр достигал 13 м (43 футов) в длину и 13,8 т (15,2 коротких тонны) в весе, тогда как кархародонтозавр приближался к 13,3 м (44 фута) в длину. и 15,1 т (16,6 коротких тонн) в весе (превышая тираннозавра ), а длина черепа голотипа гиганотозавра оценивалась в 1,56 м (5 футов). Они предупредили, что эти измерения зависят от того, были ли неполные черепа этих животных реконструированы правильно, и что для более точных оценок необходимы более полные образцы. , предложенная Далем Сассо и его коллегами, Они также обнаружили, что реконструкция спинозавра была слишком большой, и вместо этого оценили его длину в 14,3 м (47 футов), вес 20,9 т (23,0 коротких тонны) и, возможно, всего 12,6 м (41 фут). ) в длину и 12 т (13 коротких тонн) в весе. Они пришли к выводу, что эти динозавры достигли верхнего биомеханического предела размеров, достижимого для строго двуногих животных. [20] В 2010 году палеонтолог Грегори С. Пол предположил, что черепа кархародонтозавров в целом были реконструированы как слишком длинные. [21]
В 2012 году палеонтолог Мэтью Т. Каррано и его коллеги отметили, что, хотя гиганотозавр привлек много внимания из-за своего огромного размера и несмотря на то, что голотип был относительно полным, он еще не был описан подробно, за исключением черепной коробки. Они отметили, что многие контакты между костями черепа не сохранились, что приводит к неоднозначности общей длины черепа. Вместо этого они обнаружили, что черепа гиганотозавра и кархародонтозавра были точно такого же размера, как и черепа тираннозавра . Они также измерили длину бедренной кости голотипа гиганотозавра как 1,365 м (4 фута), в отличие от первоначального измерения, и предположили, что масса тела в целом была меньше. [22] В 2013 году палеонтолог Скотт Хартман опубликовал в своем блоге оценку массы методом графической двойной интеграции (на основе нарисованных реконструкций скелета), в которой он обнаружил, что тираннозавр («Сью») в целом был крупнее гиганотозавра . По его оценкам, голотип гиганотозавра весил 6,8 т (7,5 коротких тонн), а более крупный экземпляр - 8,2 т (9,0 коротких тонн). По оценкам, тираннозавр весил 8,4 т (9,3 коротких тонны), и Хартман отметил, что у него было более широкое туловище, хотя при виде сбоку они казались похожими. Он также отметил, что зубная кость гиганотозавра , которая предположительно была на 8% больше, чем у образца голотипа, скорее была бы на 6,5% больше или просто могла принадлежать животному такого же размера с более крепкой зубной костью. Он признал, что, поскольку известен только один хороший экземпляр гиганотозавра потребовалось почти столетие, чтобы найти «Сью» . тираннозавра , вполне возможно, что будут найдены более крупные особи, поскольку после открытия [23]
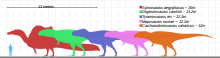
В 2014 году палеонтолог Низар Ибрагим и его коллеги оценили длину спинозавра более 15 м (49 футов) путем экстраполяции нового экземпляра, увеличенного в масштабе, чтобы соответствовать морде, описанной Далем Сассо и его коллегами. [24] Это сделало бы спинозавра крупнейшим известным плотоядным динозавром. [25] В 2019 году палеонтолог У. Скотт Персонс и его коллеги описали экземпляр тираннозавра (по прозвищу «Скотти») и оценили его как более массивный, чем другие гигантские тероподы, но предупредили, что пропорции бедренных костей кархародонтозавридов Giganotosaurus и Tyrannotitan указывают на массу тела. крупнее других взрослых тираннозавров . Они отметили, что этих теропод было известно гораздо меньше экземпляров, чем тираннозавра , и что будущие находки могут обнаружить экземпляры крупнее, чем «Скотти», о чем свидетельствует большая зубная кость гиганотозавра . Хотя у «Скотти» была самая большая окружность бедра, длина бедра гиганотозавра была примерно на 10% больше, но авторы заявили, что сравнивать пропорции между крупными кладами теропод было трудно. [26] [27]
В 2021 году палеонтолог Матиас Реолид и его коллеги собрали различные оценки массы теропод (включая гиганотозавра ) для расчета среднего значения, но не включили оценки Терриена и Хендерсона 2007 года по карнотаврам и гиганотозаврам , посчитав их выбросами. В результате масса тела гиганотозавра составила от 5,5 до 8,5 т (от 6,1 до 9,4 коротких тонны), в среднем 6,75 т (7,44 коротких тонны). Они также применили соотношение длины черепа и длины тела, предложенное Терриеном и Хендерсоном, и реконструировали различные цифровые 3D-модели теропод для измерения распределения массы тела и объема, в результате чего масса гиганотозавра длиной 13 м (43 фута) достигла 7,2 т. 7,9 коротких тонн). Эти исследователи обнаружили, что оценки соответствуют значениям, предложенным в предыдущих исследованиях. [28] В 2022 году Хуан И. Канале и его коллеги описали крупного кархародонтозавра Meraxes , у которого наиболее хорошо известен череп кархародонтозавра, предполагаемая длина которого составляет 1,27 м (4,2 фута). Экстраполируя этот череп, они оценили длину черепа гиганотозавра в 1,634 м (5,36 фута), что делает его одним из крупнейших известных черепов теропод. [29] В 2023 году Хендерсон предположил, что существует тесная связь между размерами области таза и размером тела теропод, что позволило оценить размеры неполных экземпляров. Основываясь на этой идее, он обнаружил, что длина гиганотозавра составляла 12,5 м (41 фут), что соответствует оценке, предложенной в описании 1995 года. [30]
Описание
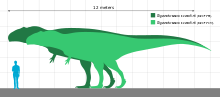
Считается, что гиганотозавр был одним из крупнейших динозавров-теропод, но неполнота его останков не позволила достоверно оценить его размер. Поэтому невозможно с уверенностью определить, был ли он крупнее , например, тираннозавра , который исторически считался самым крупным тероподом. Несколько исследователей пришли к разным оценкам размеров на основе различных методов и в зависимости от того, как были реконструированы недостающие части скелета. Оценочная длина образца голотипа варьировалась от 12 до 13 м (от 39 до 43 футов), длина черепа составляла от 1,53 до 1,80 м (от 5,0 до 5,9 футов), длина бедренной кости (бедренной кости) - от 1,365 до 1,43 м (4,48 и 5,9 футов). 4,69 футов) в длину и вес от 4,2 до 13,8 т (от 4,6 до 15,2 коротких тонн). [6] [1] [13] [20] Сращение швов (суставов) черепной коробки указывает на то, что образец голотипа был зрелой особью. [6] Второй образец, состоящий из зубной кости предположительно более крупного человека, был использован для экстраполяции длины 13,2 м (43 фута), черепа длиной 1,95 м (6,4 фута) и веса 8,2 т (9,0 коротких тонн). ). Некоторые авторы сочли оценки наибольших размеров обоих экземпляров преувеличенными. [13] [31] [23] [22] Гиганотозавра сравнивают с увеличенной версией известного рода Аллозавров . [10]
Череп

Череп гиганотозавра , хотя и не полностью известен, по-видимому, был низким. Верхняя челюсть . имела зубной ряд длиной 92 см (36 дюймов), была глубокой сверху вниз, а ее верхний и нижний края были почти параллельны челюсть имела выраженный отросток (выступ) под ноздрей и небольшое, эллипсовидное Верхняя окно (отверстие), как у аллозавра и тираннозавра . была Носовая кость очень морщинистой (шероховатой и морщинистой), и эти морщинистости продолжались назад, покрывая всю верхнюю поверхность этой кости. перед Слезная кость глазом имела выступающий морщинистый гребень (или рог), направленный вверх под углом назад. Гребень имел гребневую форму и имел глубокие бороздки. Заглазничная кость позади глаза имела направленный вниз и назад скуловой отросток, который выступал в глазницу (глазное отверстие), как это наблюдается у тираннозавра , абелизавра и карнотавра . Надглазничная кость над глазом, соприкасающаяся между слезной и заглазничной костями, имела карнизную форму и подобна кости глазницы. Абелизавр . Квадратная кость в задней части черепа имела длину 44 см (17 дюймов) и имела два пневматических (заполненных воздухом) отверстия (отверстия) на внутренней стороне. [1] [19]


Крыша черепа (образованная лобной и теменной костями ) была широкой и образовывала «полку», которая нависала над короткими надвисочными отверстиями в верхней задней части черепа. Челюсть сочленяется далеко позади затылочного мыщелка (где шея прикрепляется к черепу) по сравнению с другими тероподами. Мыщелок широкий и низкий, с воздухоносными полостями. У гиганотозавра не было сагиттального гребня на верхушке черепа, а челюстные мышцы не заходили на крышу черепа, в отличие от большинства других теропод (из-за выступа над надвисочными окнами). Вместо этого эти мышцы были бы прикреплены к нижним боковым поверхностям полки. Мышцы шеи, поднимавшие голову, должны были прикрепляться к выступающим надзатылочным костям на верхней части черепа, которые функционировали как гребень тираннозавров затылочный . [6] Латексная эндоскопия полости головного мозга гиганотозавра показала, что мозг был похож на мозг родственного рода Carcharodontosaurus , но крупнее. Эндокаст имел длину 29 мм (1 дюйм), ширину 64 мм (3 дюйма) и объем 275 мл (9,7 имп. жидких унций). [32]
Зубная кость нижней челюсти расширялась по высоте вперед (за счет нижнечелюстного симфиза , где соединялись две половины нижней челюсти), где она также была уплощена и имела выступ вниз на кончике (что было отнесено к как «подбородок»). Нижняя сторона зубной кости была вогнутой, наружная сторона при виде сверху выпуклой, по ней проходила бороздка, поддерживающая отверстия, питавшие зубы. Внутренняя сторона зубной кости имела ряд межзубных пластинок , где каждый зуб имел отверстие. меккелевская борозда По нижней границе проходила . Кривизна зубной кости показывает, что пасть гиганотозавра была широкой. Возможно, что каждая зубная кость имела двенадцать альвеол (зубных лунок). Большинство альвеол имели длину около 3,5 см (1,3 дюйма) спереди назад. Зубы зубной кости были одинаковой формы и размера, за исключением первого, который был меньше. Зубы были сжаты вбок, имели овальное в поперечном сечении, с зазубринами на переднем и заднем краях, что характерно для теропод. [13] [33] Зубы имели сигмовидную форму, если смотреть спереди и сзади. [34] Один зуб имел от девяти до двенадцати зубцов на мм (0,039 дюйма). [14] Боковые зубы гиганотозавра имели изогнутые валики эмали , а самые крупные зубы предчелюстной кости (передняя часть верхней челюсти) имели выраженные морщины (с наиболее высоким их рельефом вблизи зубцов). [35]
Посткраниальный скелет

Шея гиганотозавра была сильной, а осевая кость (шейный позвонок , сочленяющийся с черепом) была прочной. Задние шейные (шейные) позвонки имели короткие уплощенные центры («тела» позвонков) с почти полусферическими сочленениями (контактами) спереди и плевроцели (полые впадины), разделенные пластинками (пластинками). Задние (спинные) позвонки имели высокие невральные дуги и глубокие плевроцели. Хвостовые (каудальные) позвонки имели удлиненные спереди назад нервные отростки и крепкие центральные части. Поперечные отростки хвостовых позвонков были длинными спереди назад, а шевроны спереди имели лезвие. Грудной пояс был пропорционально короче, чем у тираннозавра , при этом соотношение между лопаткой (лопаткой) и бедренной костью составляло менее 0,5. Лопатка имела параллельные края и сильный бугорок для прикрепления трехглавой мышцы . Коракоид . был небольшим и крючкообразным [1]
таза Подвздошная кость имела . выпуклый верхний край, низкую поствертлужную лопасть (позади вертлужной впадины ) и узкую бревис-полку (выступ, где прикрепляются хвостовые мышцы) Лобковая . стопа выражена и короче спереди, чем сзади Седалищная кость была прямой, расширенной назад и заканчивалась лопастной формой. Бедренная кость имела сигмовидную форму, имела очень крепкую, направленную вверх головку с глубокой бороздкой (бороздкой). Малый вертел головки бедренной кости имел крыловидную форму и располагался под большим вертелом , который был коротким. Четвертый вертел был большим и выступал назад. Большеберцовая кость голени была расширена на верхнем конце, ее суставная фасетка (в месте сочленения с бедренной костью) была широкой, а ее тело сжато спереди назад. [1]
Классификация
Кориа и Сальгадо первоначально обнаружили, что гиганотозавры более тесно группируются с кладой теропод Tetanurae , чем с более базальными (или «примитивными») тероподами, такими как цератозавры , из-за общих особенностей ( синапоморфий ) в ногах, черепе и тазе. Другие особенности показали, что он находился за пределами более развитой (или «продвинутой») клады Coelurosauria . [1] В 1996 году Серено и его коллеги обнаружили, что Giganotosaurus , Carcharodontosaurus и Acrocanthosaurus тесно связаны внутри надсемейства Allosauroidea , и сгруппировали их в семейство Carcharodontosauridae . Общие черты этих родов включают слезные и заглазничные кости, образующие широкую «полку» над глазницей, а также квадратный передний конец нижней челюсти. [9]
По мере того, как было обнаружено больше кархародонтозавридов, их взаимоотношения становились яснее. и его коллеги определили эту группу как всех аллозавроидов, более близких к кархародонтозаврам, чем к аллозаврам или синрапторам . палеонтолог Томас Р. Хольц В 2004 году [36] В 2006 году Кориа и Карри объединили Giganotosaurus и Mapusaurus кархародонтозаврид в подсемейство Giganotosaurinae на основании общих особенностей бедренной кости, таких как слабый четвертый вертел и неглубокая широкая бороздка на нижнем конце. [19] В 2008 году Серено и палеонтолог Стивен Л. Брусатте объединили гиганотозавра , мапузавра и тираннотитана в трибу гиганотозаврини . [37] В 2010 году Пол перечислил гиганотозавра как « Giganotosaurus (или Carcharodontosaurus ) carolinii » без каких-либо подробностей. [21] Гиганотозавр — один из наиболее полных и информативных представителей Carcharodontosauridae. [36]


На следующей кладограмме показано место Giganotosaurus среди Carcharodontosauridae по мнению палеонтолога Андреа Кау , 2024 г.: [38]
| Кархародонтозавриды |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эволюция
Кориа и Сальгадо предположили, что конвергентная эволюция гигантизма у теропод могла быть связана с общими условиями в их среде обитания или экосистемах . [1] Серено и его коллеги обнаружили, что присутствие кархародонтозавридов в Африке ( кархародонтозавр ), Северной Америке ( акрокантозавр ) и Южной Америке ( гиганотозавр ) показало, что группа имела трансконтинентальное распространение к раннему меловому периоду . Пути распространения между северным и южным континентами, по-видимому, были прерваны океанскими барьерами в позднем меловом периоде, что привело к появлению более четкой провинциальной фауны, предотвращая обмен. [9] Ранее считалось, что меловой мир был биогеографически разделен: на северных континентах доминировали тираннозавриды, в Южной Америке — абелизавриды , а в Африке — кархародонтозавриды. [11] [39] Подсемейство Carcharodontosaurinae, к которому принадлежит Giganotosaurus , по-видимому, было ограничено южным континентом Гондвана (образованным Южной Америкой и Африкой), где они, вероятно, были высшими (высшими) хищниками . [36] Южноамериканское племя Giganotosaurini, возможно, было отделено от своих африканских родственников в результате наместничества , когда Гондвана распалась в апт - альбский период раннего мела. [34]
Палеобиология

В 1999 году палеонтолог Риз Э. Баррик и геолог Уильям Дж. Шоуерс обнаружили, что кости гиганотозавра и тираннозавра имели очень похожий состав изотопов кислорода и одинаковое распределение тепла в организме. Эти закономерности терморегуляции указывают на то, что эти динозавры имели промежуточный метаболизм между метаболизмом млекопитающих и рептилий и, следовательно, были гомойотермными (со стабильной основной температурой тела, типом « теплокровности »). Метаболизм гиганотозавра весом 8 т (8,8 коротких тонн) был бы сравним с метаболизмом плотоядного млекопитающего весом 1 т (1,1 тонны) и способствовал бы быстрому росту. [40]
В 2001 году физик Рудемар Эрнесто Бланко и Маццетта оценили беговую (беговую) способность гиганотозавра . Они отвергли гипотезу Джеймса О. Фарлоу о том, что риск травм, связанных с падением таких крупных животных во время бега, будет ограничивать скорость крупных теропод. Вместо этого они заявили, что ограничивающим фактором будет дисбаланс, вызванный увеличением скорости. Подсчитав время, которое потребуется ноге, чтобы обрести равновесие после отвода противоположной ноги, они обнаружили, что верхний кинематический предел скорости бега составляет 14 м/с (50 км/ч; 31 миля в час). Они также обнаружили, что сравнение беговых способностей гиганотозавра и птиц, таких как страус, на основе прочности их костей ног имеет ограниченную ценность, поскольку тероподы, в отличие от птиц, имели тяжелые хвосты, чтобы уравновесить их вес. [41]
Биомеханическое исследование беговых способностей тираннозавра, проведенное в 2017 году биологом Уильямом И. Селлерсом и его коллегами, показало, что скелетные нагрузки слишком велики, чтобы позволить взрослым людям бегать. Относительно длинные конечности, которые, как долгое время считалось, указывали на хорошие способности к бегу, вместо этого механически ограничивали его походкой, и поэтому он не мог быть хищником, преследующим на высокой скорости . Они предположили, что эти результаты также применимы к другим гигантским тероподам с длинными конечностями, таким как гиганотозавр , мапузавр и акрокантозавр . [42]
Кормление

В 2002 году Кориа и Карри обнаружили, что различные особенности задней части черепа (такие как наклон затылка вперед и низкий и широкий затылочный мыщелок) указывают на то, что гиганотозавр имел хорошую способность перемещать череп вбок относительно тела. передние шейные позвонки. Эти особенности также могли быть связаны с увеличением массы и длины челюстных мышц; челюстное сочленение гиганотозавра и других кархародонтозавридов было смещено назад, чтобы увеличить длину челюстной мускулатуры, что позволило быстрее смыкать челюсти, тогда как тираннозавры увеличили массу мускулатуры нижней челюсти, чтобы увеличить силу укуса. [6]
В 2005 году Терриен и его коллеги оценили относительную силу укуса теропод и обнаружили, что гиганотозавр и родственные ему таксоны имели приспособления для захвата и падения добычи путем нанесения мощных укусов, тогда как тираннозавры имели приспособления для сопротивления скручивающему напряжению и дроблению костей. Оценки в абсолютных величинах, подобных ньютонам, были невозможны. Сила укуса гиганотозавра была слабее, чем у тираннозавра , а сила уменьшалась назад по зубному ряду. Нижние челюсти были приспособлены для режущих укусов, и, вероятно, он захватывал добычу и манипулировал ею передней частью челюстей. Эти авторы предположили, что гиганотозавр и другие аллозавры могли быть универсальными хищниками, которые питались широким спектром добычи, меньшей, чем они сами, например, молодыми зауроподами. Вентральный отросток (или «подбородок») нижней челюсти, возможно, был приспособлением для сопротивления растягивающему напряжению, когда мощный укус наносился передней частью челюстей против добычи. [43]
Первые известные окаменелости близкородственного Mapusaurus были обнаружены в костном ложе, состоящем из нескольких особей на разных стадиях роста. В своем описании рода в 2006 году Кориа и Карри предположили, что, хотя это могло быть связано с длительным или случайным накоплением туш, наличие разных стадий роста одного и того же таксона указывало на то, что агрегация не была случайной. [19] В статье National Geographic 2006 года Кориа заявил, что костное ложе, вероятно, возникло в результате катастрофического события и что присутствие в основном особей среднего размера, с очень небольшим количеством молодых и старых, является нормальным для животных, образующих стаи. Следовательно, говорит Кориа, крупные тероподы могли охотиться группами, что было бы выгодно при охоте на гигантских зауроподов. [44]
Палеосреда

Гиганотозавр был обнаружен в формации Канделерос , которая отложилась в раннем сеноманском периоде позднего мела, примерно 99,6–97 миллионов лет назад. [45] [46] [31] Это образование является самым низким подразделением в группе Неукен , в котором оно является частью подгруппы Рио-Лимай . Формация сложена крупнозернистыми и среднезернистыми песчаниками, отложенными в речной среде (связанной с реками и ручьями) и в эоловых условиях (под воздействием ветра). палеопочвы (погребенная почва), алевролиты и аргиллиты Присутствуют , некоторые из которых представляют собой болотные условия. [47]
Гиганотозавр, вероятно, был высшим хищником в своей экосистеме. Он делил свою среду обитания с травоядными динозаврами, такими как титанозавр -зауропод Andesaurus и реббахизаврид- зауропод Limaysaurus и Nopcsaspondylus . Другие тероподы включают абелизаврида Ekrixinatosaurus , дромеозаврида Buitreraptor и альваресзауроида Alnashetri . Другие рептилии включают крокодилообразных Araripesuchus , сфенодонтов , змей и черепаху Prochelidella . Другие позвоночные включают кладотерийных млекопитающих, пипоидную лягушку и цератодонтиформных рыб. указывают на присутствие крупных орнитопод и птерозавров . Следы также [47] [36]
Ссылки
- ^ Перейти обратно: а б с д и ж г час я дж к Кориа, РА; Сальгадо, Л. (1995). «Новый гигантский плотоядный динозавр из мелового периода Патагонии». Природа . 377 (6546): 224–226. Бибкод : 1995Natur.377..224C . дои : 10.1038/377224a0 . S2CID 30701725 .
- ^ Хейнс, Т.; Чемберс, П. (2007). Полное руководство по доисторической жизни . Италия: Firefly Books Ltd., стр. 116–117 . ISBN 978-1-55407-181-4 .
- ^ Перейти обратно: а б Хаек, Д. (2015). «Профинансирование раскопок динозавров и обнаружение гиганта: гиганотозавра » . NPR.org . Проверено 3 августа 2016 г.
- ^ Перейти обратно: а б Понс, М. «Музей динозавров Эрнесто Бахмана – Эль Чокон» . Добро пожаловать, Аргентина . Проверено 3 августа 2016 г.
- ^ Кориа, РА; Сагадо, Л. (1994). «Гигантский теропод из среднего мела Патагонии, Аргентина». Журнал палеонтологии позвоночных . 14 (3): 22А. дои : 10.1080/02724634.1994.10011592 . JSTOR 4523584 .
- ^ Перейти обратно: а б с д и ж Кориа, РА; Карри, Пи Джей (2002). «Мозговая черепная коробка Giganotosaurus carolinii (Dinosauria: Theropoda) из верхнего мела Аргентины». Журнал палеонтологии позвоночных . 22 (4): 802–811. doi : 10.1671/0272-4634(2002)022[0802:TBOGCD]2.0.CO;2 . S2CID 85678725 .
- ^ Перенасыщение, Д.Ф. (1997). Динозавры: Энциклопедия . Джефферсон, Северная Каролина: McFarland & Co. п. 438 . ISBN 978-0-89950-917-4 .
- ^ Понс, М. «Музей динозавров Эль Чокона» . ИнтерПатагония . Проверено 14 ноября 2016 г.
- ^ Перейти обратно: а б с Серено, ПК; Дютейл, Д.Б.; Ярошене, М.; Ларссон, HCE; Лион, GH; Магвене, премьер-министр; Сидор, Калифорния; Варриккио, диджей; Уилсон, Дж. А. (1996). «Хищные динозавры Сахары и дифференциация фауны позднего мела» (PDF) . Наука . 272 (5264): 986–991. Бибкод : 1996Sci...272..986S . дои : 10.1126/science.272.5264.986 . ПМИД 8662584 . S2CID 39658297 .
- ^ Перейти обратно: а б Монастерский, Р. (1995). «Новый зверь узурпирует Ти-рекса как короля-хищника». Новости науки . 148 (13): 199. дои : 10.2307/3979427 . JSTOR 3979427 .
- ^ Перейти обратно: а б Карри, Пи Джей (1996). «Из Африки: мясоеды-динозавры, бросающие вызов тираннозавру рексу ». Наука . 272 (5264): 971–972. Бибкод : 1996Sci...272..971C . дои : 10.1126/science.272.5264.971 . JSTOR 2889575 . S2CID 85110425 .
- ^ Монастерский, Р. (1997). « Ти-рекс побежден аргентинским зверем». Новости науки . 151 (21): 317. дои : 10.2307/4018414 . JSTOR 4018414 .
- ^ Перейти обратно: а б с д Кальво, Джо; Кориа, РА (1998). «Новый экземпляр Giganotosaurus carolinii (Coria & Salgado, 1995) подтверждает, что он является самым крупным тероподом, когда-либо найденным». Гайя . 15 : 117–122.
- ^ Перейти обратно: а б с Кальво, ДЖО (1999). «Динозавры и другие позвоночные животные района озера Эсекьель-Рамос-Мексиа, Неукен-Патагония, Аргентина». Монографии Национального музея науки . 15 :13–45.
- ^ Кальво, ДЖО (1990). «Гигантский теропод из пачки Канделерос (альб-сеноман) формации Рио-Лимай, Патагония, Аргентина». VII Аргентинская конференция по палеонтологии позвоночных. Амегиниана (на испанском языке). 26 :241.
- ^ Сибахер, Ф. (2001). «Новый метод расчета аллометрических отношений длины и массы динозавров» (PDF) . Журнал палеонтологии позвоночных . 21 (1): 51–60. CiteSeerX 10.1.1.462.255 . doi : 10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2 . ISSN 0272-4634 . S2CID 53446536 .
- ^ Маццетта, Г.В.; Кристиансен, П.; Фаринья, РА (2004). «Гиганты и причуды: размер тела некоторых меловых динозавров южной и южной Америки» (PDF) . Историческая биология . 16 (2–4): 71–83. Бибкод : 2004HBio...16...71M . CiteSeerX 10.1.1.694.1650 . дои : 10.1080/08912960410001715132 . S2CID 56028251 .
- ^ Сассо, К. Даль; Магануко, С.; Бюффето, Э.; Мендес, Массачусетс (2005). «Новая информация о черепе загадочного теропода спинозавра с замечаниями о его размере и сходстве» . Журнал палеонтологии позвоночных . 25 (4): 888–896. doi : 10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2 . S2CID 85702490 .
- ^ Перейти обратно: а б с д Кориа, РА; Карри, Пи Джей (2006). «Новый кархародонтозаврид (Dinosauria, Theropoda) из верхнего мела Аргентины». Геодиверситас . 28 (1): 71–118.
- ^ Перейти обратно: а б Терриен, Ф.; Хендерсон, DM (2007). «Мой теропод больше вашего… или нет: оцениваем размер тела теропод по длине черепа». Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . ISSN 0272-4634 . S2CID 86025320 .
- ^ Перейти обратно: а б Пол, GS (2010). Принстонский полевой справочник по динозаврам . Издательство Принстонского университета. стр. 97–98 . ISBN 978-0-691-13720-9 .
- ^ Перейти обратно: а б Каррано, Монтана; Бенсон, РБЖ; Сэмпсон, SD (2012). «Филогения столбняков (Dinosauria: Theropoda)». Журнал систематической палеонтологии . 10 (2): 211–300. Бибкод : 2012JSPal..10..211C . дои : 10.1080/14772019.2011.630927 . S2CID 85354215 .
- ^ Перейти обратно: а б Хартман, С. (2013). «Массовые оценки: сокращение Севера и Юга» . Skeletal Drawing.com Скотта Хартмана . Проверено 24 августа 2013 г.
- ^ Ибрагим, Н.; Серено, ПК; Даль Сассо, К.; Магануко, С.; Фаббри, М.; Мартилл, DM; Зухри, С.; Мирвольд, Н.; Юрино, Д.А. (2014). «Полуводные адаптации гигантского хищного динозавра» . Наука . 345 (6204): 1613–1616. Бибкод : 2014Sci...345.1613I . дои : 10.1126/science.1258750 . ПМИД 25213375 . S2CID 34421257 .
- ^ Балтер, М. (2014). «Гигантский динозавр был ужасом на водных путях мелового периода». Наука . 345 (6202): 1232. Бибкод : 2014Sci...345.1232B . дои : 10.1126/science.345.6202.1232 . ПМИД 25214585 .
- ^ Лица, WS; Карри, Пи Джей; Эриксон, генеральный директор (2019). «Старый и исключительно крупный взрослый экземпляр тираннозавра рекса » . Анатомическая запись . 303 (4): 656–672. дои : 10.1002/ar.24118 . ПМИД 30897281 .
- ^ Грешко, М. (2019). самый большой в мире тираннозавр « Обнаружен » . Нэшнл Географик. Архивировано из оригинала 26 марта 2019 года . Проверено 29 марта 2019 г.
- ^ Реолид, М.; Карденаль, Ф.Дж.; Реолид, Дж. (2021). «Цифровые 3D-модели теропод для приближения к распределению массы и объема тела» . Журнал иберийской геологии . 47 (2): 599–624. Бибкод : 2021JIbG...47..599R . дои : 10.1007/s41513-021-00172-1 .
- ^ Канале, Дж.И.; Апестегия, С.; Галлина, Пенсильвания; Митчелл, Дж.; Смит, Северная Дакота; Каллен, ТМ; Шинья, А.; Халуза, А.; Джанекини, Ф.А.; Маковицкий, П.Дж. (7 июля 2022 г.). «Новый гигантский плотоядный динозавр демонстрирует конвергентные эволюционные тенденции в уменьшении рук теропод» . Современная биология . 32 (14): 3195–3202.e5. Бибкод : 2022CBio...32E3195C . дои : 10.1016/j.cub.2022.05.057 . ПМИД 35803271 .
- ^ Хендерсон, Дональд М. (2023). «Ограничения роста устанавливают верхний предел размера тела динозавра-теропода». Наука о природе . 110 (1): 4. Бибкод : 2023SciNa.110....4H . дои : 10.1007/s00114-023-01832-1 . ПМИД 36715746 . S2CID 256362332 .
- ^ Перейти обратно: а б Хольц, ТР (2011). «Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов, приложение, зима 2011 г.» (PDF) . Проверено 13 января 2012 г.
- ^ Паулина Карабахал, А.; Канале, JI (2010). «Краниальная эндокаст теропода кархародонтозаврида Giganotosaurus carolinii Coria & Salgado, 1995». Новогодний альбом по геологии и палеонтологии, Трактаты . 258 (2): 249–256. дои : 10.1127/0077-7749/2010/0104 .
- ^ Новас, FE; де Вале, С.; Викерс-Рич, П.; Рич, Т. (2005). «Большой меловой теропод из Патагонии, Аргентина, и эволюция кархародонтозавридов». Naturwissenschaften . 92 (5): 226–230. Бибкод : 2005NW.....92..226N . дои : 10.1007/s00114-005-0623-3 . hdl : 11336/103474 . ПМИД 15834691 . S2CID 24015414 .
- ^ Перейти обратно: а б Канале, Дж.И.; Новас, FE; Пол, Д. (2014). «Остеология и филогенетические взаимоотношения Tyrannotitan chubutensis Novas, de Valais, Vickers-Rich and Rich, 2005 (Theropoda: Carcharodontosauridae) из нижнего мела Патагонии, Аргентина». Историческая биология . 27 (1): 1–32. дои : 10.1080/08912963.2013.861830 . hdl : 11336/17607 . S2CID 84583928 .
- ^ Брусатте, СЛ; Бенсон, РБЖ; Карр, Т.Д.; Уильямсон, TE; Серено, ПК (2007). «Систематическая польза от морщин на эмали тероподов» . Журнал палеонтологии позвоночных . 27 (4): 1052–1056. дои : 10.1671/0272-4634(2007)27[1052:tsuote]2.0.co;2 . JSTOR 30117472 . S2CID 85615205 .
- ^ Перейти обратно: а б с д Новас, FE; Аньолин, Флорида; Эскурра, доктор медицины; Порфири, Дж.; Канале, Джи (2013). «Эволюция хищных динозавров в меловой период: данные из Патагонии». Меловые исследования . 45 : 174–215. Бибкод : 2013CrRes..45..174N . дои : 10.1016/j.cretres.2013.04.001 . hdl : 11336/102037 .
- ^ Брусатте, СЛ; Серено, ПК (2008). «Филогения Allosauroidea (Dinosauria: Theropoda): сравнительный анализ и разрешение» (PDF) . Журнал систематической палеонтологии . 6 (2): 155–182. Бибкод : 2008JSPal...6..155B . дои : 10.1017/S1477201907002404 . hdl : 20.500.11820/5f3e6d44-fea6-468d-81d3-769f8c2830dd . S2CID 86314252 .
- ^ Кау, Андреа (2024). «Единая основа макроэволюции хищных динозавров» . Бюллетень Итальянского палеонтологического общества . 63 (1): 1–19. дои : 10.4435/BSPI.2024.08 . ISSN 0375-7633 .
- ^ Кориа, Родольфо А.; Сальгадо, Леонардо (июнь 1996 г.). «Плотоядные динозавры Южной Америки» . Исследования и наука (на испанском языке) (237): 39–40.
- ^ Баррик, RE; Души, WJ (1999). «Термофизиология и биология гиганотозавра : сравнение с тираннозавром » . Электронная палеонтология . 2 (2). Архивировано из оригинала 17 мая 2011 года . Проверено 6 февраля 2016 г.
- ^ Бланко, Р. Эрнесто; Маццетта, Херардо В. (2001). «Новый подход к оценке способности к бегу гигантского теропода Giganotosaurus carolinii » . Acta Palaeontologica Polonica . 46 (2): 193–202.
- ^ Селлерс, Висконсин; Пруд, Южная Каролина; Брасси, Калифорния; Мэннинг, Польша; Бейтс, КТ (18 июля 2017 г.). «Исследование беговых способностей тираннозавра рекса с использованием динамического анализа нескольких тел с ограничениями по напряжению» . ПерДж . 5 : е3420. дои : 10.7717/peerj.3420 . ISSN 2167-8359 . ПМК 5518979 . ПМИД 28740745 .
- ^ Терриен, Ф.; Хендерсон, DM; Рафф, CB, 2005, «Укуси меня: биомеханические модели нижних челюстей теропод и их значение для питания». В: Карпентер К. Плотоядные динозавры. Жизнь прошлого . Издательство Университета Индианы. стр. 179–237
- ^ Оуэн, Дж. (2006). «Мясоедный динозавр был больше тираннозавра» . Nationalgeographic.com . Национальные географические новости. Архивировано из оригинала 30 сентября 2016 года . Проверено 27 августа 2016 г.
- ^ Райнольди, Алабама; Франкини, М.; Бофорт, Д.; Патриер, П.; Джузиано, А.; Импичини, А.; Понс, Дж. (2014). «Крупномасштабное обесцвечивание красных отложений, связанное с восходящей миграцией углеводородов: высота Лос-Чиуидос, бассейн Неукен, Аргентина». Журнал осадочных исследований . 84 (5): 373–393. Бибкод : 2014JSedR..84..373R . дои : 10.2110/jsr.2014.31 . hdl : 11336/34039 .
- ^ Санчес, МЛ; Асурменди, Э. (2015). «Стратиграфия и седиментология конечного конуса формации Канделерос (группа Неукен), нижний мел, бассейн Неукен, провинции Неукен и Мендоса, Аргентина» . Андская геология . 42 (3). дои : 10.5027/andgeoV42n3-a03 . hdl : 11336/69686 .
- ^ Перейти обратно: а б Леанза, HA; Апестегия, С.; Новас, FE; де ла Фуэнте, MS (2004). «Меловые наземные отложения из бассейна Неукен (Аргентина) и их комплексы четвероногих». Меловые исследования . 25 (1): 61–87. Бибкод : 2004CrRes..25...61L . дои : 10.1016/j.cretres.2003.10.005 .
Внешние ссылки
 Данные, относящиеся к гиганотозаврам , в Wikispecies
Данные, относящиеся к гиганотозаврам , в Wikispecies  СМИ, связанные с гиганотозавром, на Викискладе?
СМИ, связанные с гиганотозавром, на Викискладе? - Канадский музей природы: «Кто был настоящим динозавром? Гиганотозавр или Ти-рекс ?» - видео представлено Джорданом Мэллоном















| Базы данных органов управления : Национальные |
|---|