Литронакс
| Литронакс Временной диапазон: поздний мел ,
| |
|---|---|

| |
| Реконструированный скелет, Общественный музей Милуоки. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Тираннозавриды |
| Подсемейство: | † Тираннозавры |
| Клэйд : | † Тератофонеини |
| Род: | † Литронакс Лоуэн и др. , 2013 |
| Типовой вид | |
| † Литронакс аргестес Лоуэн и др. , 2013
| |
Литронакс ( LYE -thro-nax ) — род динозавров тираннозаврид - , обитавших в Северной Америке около 81,9–81,5 миллионов лет назад в позднемеловой период . Единственный известный экземпляр был обнаружен в штате Юта в формации Вахвип национального памятника Большая лестница-Эскаланте в 2009 году и состоит из частичного черепа и скелета. В 2013 году он стал основой нового рода и вида Lythronax argestes ; Родовое название Lythronax означает «король крови», а видовое название argestes происходит от названия греческого поэта Гомера , обозначающего юго-западный ветер, что указывает на географическое происхождение этого экземпляра в Северной Америке.
Оценочные размеры литронакса варьируются от 5 до 8 м (от 16 до 26 футов) в длину и от 0,5 до 2,5 т (от 1100 до 5500 фунтов) по весу. Это был тираннозаврид тяжелого телосложения, и как член этой группы он имел маленькие двупалые передние конечности, сильные задние конечности и очень крепкий череп. Задняя часть черепа Литронакса , по-видимому, была очень широкой, с глазницами, обращенными вперед так же, как у тираннозавра . У Литронакса было 11 зубных лунок в ; верхней челюсти у большинства тираннозаврид было больше. Передние зубы были самыми большими, самый длинный имел длину почти 13 см (5 дюймов). Другие детали черепа и скелета, которые отличали Литронакса от других тираннозаврид, включали S-образный внешний край верхней челюсти и отросток таранной кости лодыжки, выступ, который расширялся дальше вверх по сравнению с его родственниками.
Голотип был обнаружен в пачке Рейнольдс- Пойнт формации Вахвип, которая датируется кампанским этапом мелового периода. Таким образом, литронакс является старейшим известным членом семейства Tyrannosaurus и считается более базальным , чем тираннозавр . Из-за своего возраста Латронакс важен для понимания эволюционного происхождения тираннозаврид, включая развитие их анатомических специализаций. Направленные вперед глаза Литронакса давали ему восприятие глубины , что могло быть полезно во время преследования или нападения хищников .
Открытие и присвоение имени
[ редактировать ]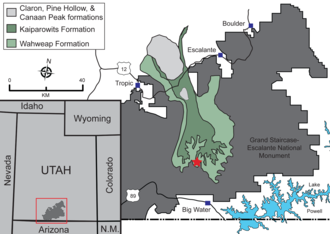
В 2009 году Скотт Ричардсон из Бюро землеустройства США искал окаменелости (BLM) вместе с коллегой в формации Вахвип Национального памятника Большая лестница-Эскаланте на юге Юты, когда они наткнулись на ногу и носовую кость динозавр теропод - в районе Ниппл-Бьютт. Ричардсон связался с командой палеонтологов из Университета Юты , которые были взволнованы, но поначалу были настроены скептически, поскольку окаменелости теропод в этом районе ранее не обнаруживались. Им прислали фотографию носовой кости, по которой они определили, что она принадлежит тираннозавру , который, вероятно, был новым видом , поскольку он произошел из эпохи, когда еще не было известных представителей этой группы. Ископаемые останки были тщательно раскопаны в течение года совместной командой BLM и Музея естественной истории штата Юта (UMNH). Местность, являющаяся государственной землей , получила обозначение UMNH VP 1501. [1] [2] [3] [4] До официального описания динозавра его называли «тираннозавром Ниппель-Бьютт» или «тираннозавридом Вахвепа». [5] [4]
Образец UMNH VP 20200 (с префиксом, обозначающим его хранение в UMNH), был сделан голотипом нового рода и вида Lythronax argestes палеонтологом Марком А. Лоуэном и его коллегами в 2013 году . Родовое название происходит от греческих слов литрон (λύθρον), что означает «запекшаяся кровь», и анакс (ἄναξ), что означает «царь». Видовое название argestes (ἀργεστής) — это греческое название, использованное поэтом Гомером для обозначения ветра с юго-запада в связи с тем, где этот экземпляр был найден в Северной Америке. [6] Полностью научное название можно перевести как «король крови (или «король крови») с юго-запада». Лоуэн заявил, что суффикс, означающий «король» в имени Литронакса, был предназначен для намека на его более позднего, похожего родственника Тираннозавра Рекса . Префикс, означающий «запекшуюся кровь», был выбран, чтобы проиллюстрировать «его предполагаемый образ жизни хищника с головой, покрытой кровью мертвого животного». [2] [3] [7]

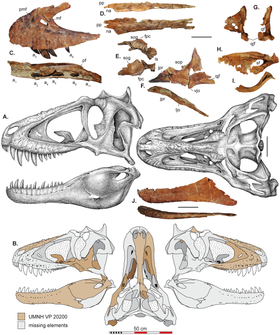
Голотип и единственный известный экземпляр Lythronax состоит из частичного черепа и скелета, который включает правую верхнюю челюсть , обе носовые кости, правую лобную часть , левую скуловую кость , левую квадратную кость , правый латеросфеноид , правое небное , левую зубную кость , левая сплениальная , левая надугловая , левая предсуставная , дорсальное ребро , каудальный шеврон , обе лобковые кости , левая большеберцовая и малоберцовая кости , а также левая вторая и четвертая плюсневые кости . [6] В статье, названной Lythronax , авторы также описали новый экземпляр геологически более молодого тираннозавра Teratophoneus (который получил имя в 2011 году); этот род известен из формации Кайпаровиц в Гранд-Стэйркейс-Эскаланте, и два тираннозавра использовались для исследования эволюционного и географического происхождения семейства Tyrannosauridae . [6] [8] Основываясь на выводах статьи, UMNH на своем веб-сайте назвала Литронакса «двоюродным дедушкой» тираннозавра . [3]
В 2017 году правительство США объявило о планах уменьшить Большую лестницу-Эскаланте (чуть более чем вдвое) и памятники «Медвежьи уши» , чтобы обеспечить добычу угля и другие виды развития энергетики на земле; это было крупнейшее сокращение национальных памятников США в истории. [9] [10] Сам Литронакс был одним из двух динозавров из бывшего памятника, упомянутого в президентской декларации , наряду с Диаблоцератопсом . [11] Американский палеонтолог Скотт Д. Сэмпсон (соавтор литронакса ), который курировал большую часть ранних исследований памятника, выразил опасение, что такой шаг поставит под угрозу дальнейшие открытия. [10] [6] Средства массовой информации подчеркнули важность открытия ископаемых в этом районе, в том числе более 25 новых таксонов , в то время как некоторые выделили литронакс как одну из важных находок. [12] [13] [14] Впоследствии на правительство США подала в суд группа ученых, защитников окружающей среды и коренных американцев ; в 2021 году памятник был восстановлен в прежнем размере последующей администрацией. [9] [14] [15]
Описание
[ редактировать ]
На момент анонса Литронакса новостные сайты сообщали об оценочных размерах около 7,3–8 м (24–26 футов) в длину и около 2,5 т (5500 фунтов) в весе на основе сравнения с гораздо более крупным родственником Тираннозавра ; Лоуэн заявил, что он, возможно, стал еще больше. [1] [2] Американский палеонтолог Грегори С. Пол в 2016 году дал более низкую оценку: длину 5 м (16 футов) и вес всего 500 кг (1100 фунтов). [16] В 2019 году на основе объемного анализа размер голотипа оценивался в 6,8 м (22 фута) в длину, 2,3 м (7,5 фута) в высоту по бедрам и 1,4 т (1,5 коротких тонны) в массе тела. [17] Литронакс был относительно крепким тираннозавридом. Как и другие члены группы, он обладал маленькими двупалыми передними конечностями, большими и сильными задними конечностями, широкими челюстями и очень прочным черепом. [16] Tyrannosauroidea с небольшим телом Хотя более ранние представители надсемейства обладали протоперьями , их наличие могло варьироваться в зависимости от вида или возраста особи. [4]

Литронакс имел относительно короткую морду и широкий череп (ширина более 40% длины), как и у других тираннозаврид. Носовые кости в верхней части морды были намного шире спереди, чем посередине, в отличие от других тираннозаврид. При взгляде сверху внешние края черепа (образованные верхней и скуловой костями) имели сильно сигмовидную (или s-образную) форму. Наряду с шириной лобной кости (кость в верхней части черепа), это, по-видимому, сделало заднюю часть Литронакса черепа очень широкой, с орбитами (глазницами), обращенными почти вперед. В остальном эти особенности известны только у Тарбозавра и Тираннозавра ; у ранее расходящихся тираннозаврид орбиты были меньше обращены вперед, а задняя часть их черепов была уже. [6]
Литронакс отличался еще и тем, что поверхности лобной кости, соприкасающиеся с предлобной и заглазничной костями на ее передней и задней сторонах, были разделены лишь узкой бороздкой. Максиллы Lythronax были крепкими и сильно выпуклыми по внешнему краю, как и у всех других известных тираннозаврид, но отличались сигмовидными краями. У Литронакса было 11 альвеол (зубных лунок) на каждой верхней челюсти, и этот признак не характерен ни для одного тираннозавра, кроме Тератофонеуса и Бистахиевёрсора (у других тираннозавров было 12 или более верхнечелюстных альвеол). Зубы верхней челюсти были гетеродонтными (дифференцированными), первые пять были значительно крупнее последующих. [6] Некоторые из самых передних зубов были почти 13 см (5 дюймов) в длину. [1] Зубы по форме напоминали бананы, крепкие и зазубренные . [18] Как и у тираннозавра , нёбная полка была хорошо развита. [6]
Скуловая кость (или «щечная» кость) была прочной и имела широкий заглазничный отросток (который выступал вверх от скуловой кости и контактировал с заглазничной костью), в отличие от других тираннозавров, за исключением Bistahieversor , Tyrannosaurus и Tarbosaurus . Передняя граница заглазничного отростка имела сильный выступ, что указывает на то, что у Lythronax был большой подглазничный фланец (выступ в нижнюю часть глазницы), непохожий на более мелкие у других тираннозаврид. Каждая ветвь зубной кости (половина зубной части нижней челюсти) была сильно вогнута к внешней стороне (выгнута внутрь по длине черепа). Это отражало контуры верхней челюсти и сильное расширение задней части черепа; он был похож на Бистахиевёрсора , Тираннозавра и Тарбозавра , но отличался от других тираннозавроидов. Зубная кость также была глубокой на заднем конце, что указывает на то, что следующая часть нижней челюсти была сравнима с Тарбозавром и Тираннозавром по глубине , но не с другими тираннозавридами. Как и другие тираннозавриды, надугольная кость позади зубной кости имела глубокий и хорошо развитый выступ прямо перед тем местом, где челюсть сочленялась с черепом, а литронакс был похож на тираннозавра тем, что этот выступ имел вогнутую верхнюю поверхность. [6]

Хотя посткраниальный скелет Lythronax малоизвестен, известные остатки лобковой кости (части таза) и задних конечностей демонстрируют черты, типичные для Tyrannosauridae. Лобковый ботинок, расширение на нижнем конце лобковой кости, имел большой, направленный вперед отросток, как и у всех тираннозаврид. У Lythronax лобковый ботинок был большим и сравнительно глубоким, наиболее похожим на таковой у Tarbosaurus и Tyrannosaurus , но отличался от менее расширенных лобковых ботинок Teratophoneus , Albertosaurus , Gorgosaurus и Daspletosaurus . Малоберцовая кость, кость голени, имела на верхнем конце глубокую срединную впадину, как и у других тираннозаврид. У Lythronax таранная кость голеностопного сустава имела восходящий отросток над сочленением со стопой, который был расширен дальше вверх по сравнению со своими родственниками. [6]
Классификация
[ редактировать ]
Lythronax argestes принадлежит к семейству Tyrannosauridae, семейству крупнотелых целурозавров ; большинство родов тираннозаврид известны из Северной Америки и Азии. [6] Судя по своему стратиграфическому положению, литронакс является старейшим тираннозавридом, обнаруженным на сегодняшний день. [2] [3] [6] До того, как Lythronax был официально назван, Занно и его коллеги в 2013 году отметили, что образец голотипа, вероятно, отличается от Teratophoneus и Bistahieversor , которые также происходят из южной Юты. присутствовало по крайней мере три рода тираннозаврид в Западном внутреннем бассейне Это означало бы, что на кампанском этапе . Филогенетический анализ, проведенный Занно и его коллегами, поместил все три таксона в одну группу тираннозаврид, исключив всех остальных членов группы. [4]
Подробный филогенетический анализ, проведенный Левеном и его коллегами в дополнение к их описанию Lythronax в 2013 году , основанный на 303 краниальных и 198 посткраниальных особенностях, поместил его и Teratophoneus в подсемейство Tyrannosaurinae . Lythronax был родственным таксоном группы, состоящей из маастрихтских таксонов Tarbosaurus и Tyrannosaurus и позднекампанского Zhuchengtyrannus . Он был более тесно связан с этой группой, чем другие таксоны, такие как Daspletosaurus и Teratophoneus , которые были моложе Lythronax , но старше этой группы. [6]
В 2017 году американские палеонтологи Стивен Брусатт и Томас Д. Карр опубликовали новый филогенетический анализ Tyrannosauroidea, включая более полный набор анатомических характеристик и таксонов, который не согласовывался с результатами Левена и его коллег. Хотя триба Alioramini в анализе Левена и его коллег не находилась за пределами Tyrannosauridae, Брусатте и Карр поместили эту группу как самую базальную (раннедивергентную или «примитивную») группу среди Tyrannosaurinae. И наоборот, Лоуэн и его коллеги обнаружили, что Bistahieversor является производным («продвинутым») тираннозавром, тесно связанным с аналогичным образом производными Teratophoneus и Lythronax , в то время как Брусатте и Карр поместили его в более базальное положение непосредственно за пределами Tyrannosauridae, причем как Teratophoneus , так и Lythronax являются базальными тираннозавринами. . Было высказано предположение, что оба этих результата возникли из-за чрезмерного придания большого значения некоторым характеристикам Лоуэном и его коллегами, что привело к исключению длиннорылых форм алиорамина из короткорылых тираннозаврин и размещению Бистахиеверсор и Литронакс ближе к тираннозаврам, чем в противном случае. [19] Результаты двух контрастирующих анализов показаны на кладограммах ниже: [6] [19] [20]
|
Топология 1: Лоуэн и коллеги, 2013 г. |
Топология 2: Брусатте и Карр, 2017 г.
|
В популярной книге, опубликованной в 2016 году, Пол предположил, что Lythronax argestes может быть членом рода Tyrannosaurus , и отметил, что производные тираннозавриды «сильно чрезмерно разделены на уровне рода». [16] Последующие публикации, включая как таксономический, так и филогенетический анализ, сохранили этот вид в отдельном роде Lythronax . [19] [20] [21] [22] В 2023 году палеонтологи Чарли Р. Шерер и Кристиан Войкулеску-Холвад перенесли Lythronax в новую кладу Teratophoneini вместе с Teratophoneus и Dynamoterror . [23]
Палеобиогеография
[ редактировать ]
В течение позднего мелового периода (около 95 миллионов лет назад) Западный внутренний морской путь изолировал западную часть Северной Америки ( Ларамидия ) от восточной части Северной Америки ( Аппалачи ), а иногда изолировал осадочные бассейны друг от друга. [24] Это привело к развитию высокоэндемичных экосистем в Ларамидии; эти экосистемы также грубо разделены на северную и южную провинции, [6] [8] [25] но такое чистое разделение оспаривается. [19] [26] Как и многие линии ларамидских динозавров, эволюционная история тираннозаврид, распространение которых ограничено Азией и Ларамидией, характеризуется обменом фауной между двумя континентами. [27] Последовательность событий обмена, произошедших среди ларамидских тираннозаврид, неясна, а разнообразные тираннозавроиды, обнаруженные в южной Ларамидии (включая Lythronax , Teratophoneus и Bistahieversor ), еще больше усложнили их эволюционную историю. [6] [8] В частности, нерешенным остается вопрос, произошел ли тираннозавр от азиатских тираннозаврид или от южных ларамидских тираннозаврид. [20]
Основываясь на своих филогенетических результатах, Занно и его коллеги предположили, что тогда еще безымянный Lythronax обладал чертами, которые объединяли тираннозаврид из южной Ларамидии, исключая другие роды. [4] Хотя Левен и его коллеги не обнаружили уникальную группу южных таксонов, они решили, что все три тесно связаны друг с другом и являются базальными для группы более крупных, более поздних форм. [6] На основании этих результатов Лоуэн и его коллеги предположили, что существует значительное биогеографическое разделение между северными ларамидскими и южными ларамидскими формами с ограниченным взаимообменом. Кроме того, поскольку они обнаружили, что Alioramini относят к тираннозавридам, а азиатские роды Tarbosaurus и Zhuchengtyrannus относятся к группе, исключающей всех других тираннозаврид, Левен и его коллеги предположили, что произошел только один обмен тираннозавридами из Северной Америки в Азию. Они предположили, что обмен произошел в конце кампана, когда глобальный уровень моря упал, а тираннозавр произошел от североамериканских форм, существовавших до того, как произошла такая миграция. [6]

Из-за различных филогенетических результатов биогеографические выводы Левена и его коллег были оспорены Брусаттом и Карром. Поскольку Бистахиеверсор из южной Ларамидии был помещен за пределами Тираннозаврид, а Тератофонеус из Юты гнездился ближе всего к аляскинскому Нануксавру , Брусатте и Карр вместо этого предположили, что между северной и южной Ларамидией происходили динамичные и повторяющиеся обмены фауной тираннозаврид, и отвергли наличие эндемичных провинций. Азиатские таксоны Tarbosaurus , Zhuchengtyrannus , Qianzhousaurus и Alioramus также были отнесены к Tyrannosaurinae, среди североамериканских родов. Брусатте и Карр предположили, что произошли по крайней мере два континентальных обмена, где тираннозавры возникли в Азии и мигрировали в Северную Америку после дивергенции алиораминов, а затем снова вернулись в Азию с тарбозавром и жученгтиранносом . Другой возможный сценарий, предложенный Брусатте и Карром, заключался в том, что произошли две отдельные миграции в Азию, которые по отдельности дали начало алиораминам и более крупным, более поздним формам. В обоих сценариях Тираннозавр , гнездившийся среди азиатских таксонов, был «инвазивным мигрантским видом, распространившимся по Ларамидии» из Азии в Маастрихте. [19]
Гипотезы об азиатско-североамериканской миграции Брусатте и Карра были подтверждены более поздним анализом, проведенным канадским палеонтологом Джаредом Ворисом и его коллегами в 2020 году . Однако Ворис и его коллеги внесли поправки в первоначальный анализ, добавив роды Dynamoterror из южной Ларамидии ( Нью-Мексико ) и Thanatotheristes из северной Ларамидии ( Альберта ), и они смогли воспроизвести подразделения тираннозаврид с севера на юг, предложенные Левеном и его коллегами. . Южные таксоны Teratophoneus , Dynamoterror и Lythronax образовали исключительную группу (за исключением Nanuqsaurus , в отличие от Brusatte и Carr) коротко- и глубокорылых таксонов вне группы более производных северных ларамидских форм, а южные ларамидские формы также имел отдельный морфотип скелета. Ворис и его коллеги предположили, что эти морфологические различия возникли по экологическим причинам, возможно, включая состав добычи или стратегии питания. Поскольку основные группы добычи были одинаковыми в северной и южной Ларамидии, когда тираннозавриды жили в этих регионах, Ворис и его коллеги пришли к выводу, что различия в анатомии черепа возникли из-за различий в стратегиях питания. [20]
Палеобиология
[ редактировать ]

Литронакс отличался от большинства других тираннозаврид укороченным черепом с расширенной задней частью, а также направленными вперед орбитами (что было прямым следствием морфологии его черепа). Ни один другой тираннозавроид не имел таких направленных вперед орбит, за исключением тираннозавра и тарбозавра . [6] хотя более продвинутые тираннозавроиды обычно имели более крупные и более направленные вперед орбиты, чем базальные тираннозавроиды. [28] Открытие Литронакса позволяет предположить, что эти персонажи появились как минимум 80 миллионов лет назад. [6]
Направленные вперед орбиты Литронакса расширили бы поле зрения его бинокулярного зрения за счет увеличения расстояния между орбитами и сделав их линии зрения более параллельными друг другу (т. е. уменьшив расхождение оптических осей). [28] что дало бы Литронаксу глубокое восприятие . [2] [29] В 2006 году палеонтолог Кент Стивенс предположил, что подобные орбиты тираннозавра помогли бы либо преследовать хищников путем наблюдения за удаленной добычей и трехмерного обнаружения препятствий, либо устраивать засады хищников благодаря способности определять время и направление выпадов. [28]
Будучи тираннозавридом, Литронакс, вероятно, разделял бы другие специализации группы, связанные с хищническим образом жизни, включая большие размеры тела; большой череп с мощными челюстными мышцами и крепкими зубами; усиленные швы, скрепляющие кости черепа; и относительно небольшие передние конечности. [2] [30] Зубы и челюстные мышцы Литронакса способствовали созданию сильной силы укуса, позволяющей не только вырезать куски плоти, но и дробить кости. [7] [18] Напряжения и нагрузки от этих укусов были бы эффективно поглощены сросшимися, изогнутыми носовыми костями и усиленными швами. [31] [32]
Палеосреда
[ редактировать ]
Литронакс был обнаружен в наземных осадочных породах, принадлежащих нижней части пачки Рейнольдс-Пойнт формации Вахвип. Возраст пород, из которых образовался литронакс , оценивается в 81,49 млн лет назад с диапазоном неопределенности 81,86–81,45 млн лет назад. Возраст всей формации Вахвеап радиометрически датирован от 82,2 до 77,3 миллиона лет. [33] Во времена жизни Литронакса Западный внутренний морской путь находился в самом широком месте, почти полностью изолируя южную Ларамидию от остальной части Северной Америки. [2] Территория существования динозавров включала озера, поймы и реки, текущие на восток. Формация Вахвип является частью региона Большой Лестницы , огромной последовательности слоев осадочных пород, которые простираются на юг от национального парка Брайс-Каньон через национальный парк Зайон и в Гранд-Каньон . Среди других свидетельств наличие быстро отложившихся отложений предполагает влажный сезонный климат. [34]
Литронакс , вероятно, был крупнейшим хищником своей экосистемы. [2] Он разделял свою палеосреду с другими динозаврами, такими как гадрозавры Acristavus и Adelolophus . [35] цератопсовый диаблоцератопс , [2] [36] [37] и безымянные анкилозавры и пахицефалозавры . [38] Позвоночные животные, присутствовавшие в то время в формации Вахвип, включали пресноводных рыб , боуфинов , многочисленных скатов и акул , черепах, таких как компсемис , крокодилов , [39] и двоякодышащая рыба . [40] В этом регионе обитали многочисленные млекопитающие , в число которых входили несколько родов многотуберкулезных , кладотерий , сумчатых и плацентарных насекомоядных . [41] Млекопитающие были более примитивными, чем те, что обитали в более молодой формации Кайпаровиц. Следы окаменелостей относительно многочисленны в формации Вахвеап и предполагают присутствие крокодиломорфов , а также птицетазовых и тероподовых динозавров. [42] Доказательства активности беспозвоночных в этой формации варьировались от нор окаменевших насекомых в окаменелых бревнах. [43] окаменелостям моллюсков , крупных крабов , [44] и большое разнообразие брюхоногих моллюсков и остракод . [45]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Ралофф, Дж. (2013). «Король Гора» . Новости науки для студентов . Проверено 5 февраля 2020 г.
- ^ Перейти обратно: а б с д и ж г час я Вергано, Д. (6 ноября 2013 г.). «Новообретенный динозавр «Короля Гора» правил до Ти-рекса » . Нэшнл Географик . Архивировано из оригинала 8 ноября 2013 года . Проверено 22 ноября 2013 г.
- ^ Перейти обратно: а б с д « Кровавый король Юго-Запада», Lythronax argestes » . Музей естественной истории штата Юта . 6 ноября 2013 г. Проверено 22 ноября 2013 г.
- ^ Перейти обратно: а б с д и Занно, Ле; Лоуэн, Массачусетс; Фарке, А.А.; Ким, Г.-С.; Классенс, LPAM; МакГаррити, Коннектикут (2013). «Позднемеловые динозавры-тероподы Южной Юты». В Титусе, Алабама; Лоуэн, Массачусетс (ред.). На вершине парадной лестницы: поздний мел Южной Юты . Блумингтон: Издательство Университета Индианы. стр. 504–525. ISBN 978-0-253-00883-1 .
- ^ ДеБлё, Д.Д.; Киркланд, Дж.И.; Гейтс, штат Калифорния; Итон, Дж. Г.; Гетти, Массачусетс; Сэмпсон, SD; Лоуэн, Массачусетс; Хайден, MC (2013). «Палеонтологический обзор и тафономия среднекампанской формации Вахвеап в национальном памятнике Большая лестница – Эскаланте». В Титусе, А.; Лоуэн, М. (ред.). На вершине парадной лестницы: поздний мел Южной Юты . Блумингтон: Издательство Университета Индианы. стр. 563–577. ISBN 978-0-253-00883-1 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Лоуэн, Массачусетс; Ирмис, РБ; Сертич, JJW; Карри, Пи Джей; Сэмпсон, SD (2013). Эванс, округ Колумбия (ред.). «Эволюция динозавров-тиранов отслеживает подъем и падение океанов позднего мела» . ПЛОС ОДИН . 8 (11): е79420. Бибкод : 2013PLoSO...879420L . дои : 10.1371/journal.pone.0079420 . ПМЦ 3819173 . ПМИД 24223179 .
- ^ Перейти обратно: а б Виегас, Дж. (6 ноября 2013 г.). «Зубастый динозавр терроризировал Юту раньше тираннозавра » . НОВОСТИ Дискавери . Архивировано из оригинала 8 мая 2015 года . Проверено 16 ноября 2013 г.
- ^ Перейти обратно: а б с Карр, Т.Д.; Уильямсон, TE; Бритт, BB; Штадтман, К. (2011). «Доказательства высокого таксономического и морфологического разнообразия тираннозаврид в позднем мелу (позднем кампане) на юго-западе Америки и нового короткочерепного тираннозаврид из формации Кайпаровиц в штате Юта». Naturwissenschaften . 98 (3): 241–246. Бибкод : 2011NW.....98..241C . дои : 10.1007/s00114-011-0762-7 . ПМИД 21253683 . S2CID 13261338 .
- ^ Перейти обратно: а б Гонсалес, Р.; Зиглер, К.; Дуайер, К. (4 декабря 2017 г.). «Трамп распорядился о крупнейшем сокращении национального памятника в истории США» . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . Архивировано из оригинала 5 июля 2019 года . Проверено 25 июня 2019 г.
- ^ Перейти обратно: а б Финнеган, М. (25 июня 2017 г.). «Выдающиеся открытия динозавров оказались под угрозой из-за плана Трампа по сокращению национального памятника в штате Юта, говорят ученые» . Лос-Анджелес Таймс . Архивировано из оригинала 25 июня 2019 года . Проверено 24 июня 2019 г.
- ^ Трамп, диджей (2017). «Президентская указа об изменении Национального памятника Большой лестницы – Эскаланте» . Белый дом . Проверено 6 февраля 2020 г. - из Национального архива .
- ^ Уайлс, Т. (30 января 2018 г.). «Как сокращение Трампом государственных земель угрожает будущим открытиям динозавров» . Хранитель . Проверено 6 февраля 2020 г.
- ^ Грэмлинг, К. (27 апреля 2017 г.). «Наука и политика сталкиваются из-за Медвежьих ушей и других национальных памятников» . Наука АААС . Архивировано из оригинала 26 июня 2019 года . Проверено 26 июня 2019 г.
- ^ Перейти обратно: а б Вей-Хаас, М. (18 декабря 2017 г.). «Что означает сокращение национальных памятников, богатых ископаемыми, для науки» . Смитсоновский институт . Архивировано из оригинала 24 июня 2019 года . Проверено 24 июня 2019 г.
- ^ «Общество дикой природы и др. Трамп и др. (Большая лестница – Эскаланте)» . НРДЦ . 10 апреля 2020 г. Проверено 19 апреля 2020 г.
- ^ Перейти обратно: а б с Пол, GS (2016). Принстонский полевой справочник по динозаврам (второе изд.). Издательство Принстонского университета. стр. 108–109, 113. ISBN. 978-0-691-16766-4 .
- ^ Молина-Перес, Р.; Ларраменди, А. (2019). Факты и цифры о динозаврах: тероподы и другие динозаврообразные . Издательство Принстонского университета. п. 267. ИСБН 9780565094973 .
- ^ Перейти обратно: а б Мохан, Г. (6 ноября 2013 г.). «Король кровавых динозавров был «плохим дедушкой» тираннозавров» . Лос-Анджелес Таймс . Проверено 8 ноября 2013 г.
- ^ Перейти обратно: а б с д и Брусатте, СЛ; Карр, ТД (2017). «Филогения и история эволюции тираннозавроидов» . Научные отчеты . 6 : 20252. Бибкод : 2016NatSR...620252B . дои : 10.1038/srep20252 . ПМЦ 4735739 . ПМИД 26830019 .
- ^ Перейти обратно: а б с д Ворис, Дж. Т.; Терриен, Ф.; Зеленицкий, ДК; Браун, СМ (2020). «Новый тираннозавр (Theropoda: Tyrannosauridae) из передовой кампанской формации Альберты, Канада, дает представление об эволюции и биогеографии тираннозаврид». Меловые исследования . X : 104388. Бибкод : 2020CrRes.11004388V . дои : 10.1016/j.cretres.2020.104388 .
- ^ Мэллон, Джей Си; Бура, младший; Карри, Пи Джей (2019). «Проблемный скелет тираннозаврид (Dinosauria: Theropoda) и его значение для разнообразия тираннозаврид в формации каньона Подкова (верхний мел) в Альберте» . Анатомическая запись . 303 (4): 673–690. дои : 10.1002/ar.24199 . ПМК 7079176 . ПМИД 31254458 .
- ^ Макдональд, AT; Вульф, генеральный директор; Дули, AC младший (2018). «Новый тираннозаврид (Dinosauria: Theropoda) из верхнемеловой формации Менефи в Нью-Мексико» . ПерДж . 6 :6:e5749. дои : 10.7717/peerj.5749 . ПМК 6183510 . ПМИД 30324024 .
- ^ Шерер, Чарли Роджер; Войкулеску-Холвад, Кристиан (2023). «Повторный анализ набора данных опровергает утверждения об анагенезе у тираннозавров линии тираннозавров (Theropoda, Tyrannosauridae)» . Меловые исследования . 155 : 105780. doi : 10.1016/j.cretres.2023.105780 .
- ^ Хорнер, младший; Варриккио, диджей; Гудвин, МБ (1992). «Морские трансгрессии и эволюция меловых динозавров». Природа . 358 (6381): 59–61. Бибкод : 1992Natur.358...59H . дои : 10.1038/358059a0 . S2CID 4283438 .
- ^ Сэмпсон, SD; Лоуэн, Массачусетс; Фарке, А.А.; Робертс, EM; Форстер, Калифорния; Смит, Дж.А.; Титус, Алабама; Степанова, А. (2010). «Новые рогатые динозавры из Юты служат доказательством внутриконтинентального эндемизма динозавров» . ПЛОС ОДИН . 5 (9): e12292. Бибкод : 2010PLoSO...512292S . дои : 10.1371/journal.pone.0012292 . ПМЦ 2929175 . ПМИД 20877459 .
- ^ Леман, ТМ (2001). «Позднемеловая провинциальность динозавров». В Танке, Д.; Карпентер, К. (ред.). Жизнь мезозойских позвоночных . Блумингтон: Издательство Университета Индианы. стр. 310–328. ISBN 978-0-253-33907-2 .
- ^ Апчерч, П.; Ханн, Калифорния; Норман, Д.Б. (2002). «Анализ биогеографии динозавров: доказательства существования закономерностей викариантности и расселения, вызванных геологическими событиями» . Труды Королевского общества Б. 269 (1491): 613–621. дои : 10.1098/рспб.2001.1921 . ПМК 1690931 . ПМИД 11916478 .
- ^ Перейти обратно: а б с Стивенс, К.А. (2006). «Бинокулярное зрение тероподовых динозавров». Журнал палеонтологии позвоночных . 26 (2): 321–330. doi : 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 .
- ^ Уайт, М. (6 ноября 2013 г.). «Новый вид динозавров, Lythronax argestes , обнаружен в штате Юта» . Хаффингтон Пост . Проверено 8 ноября 2013 г.
- ^ Брусатте, СЛ; Норелл, Массачусетс; Карр, Т.Д.; Эриксон, генеральный директор; Хатчинсон-младший; Баланов, А.М.; Бевер, Г.С.; Шуаньер, Ж.Н.; Маковицкий, П.Дж.; Сюй, X. (2010). «Палеобиология тираннозавров: новые исследования древних образцов организмов» (PDF) . Наука . 329 (5998): 1481–1485. Бибкод : 2010Sci...329.1481B . дои : 10.1126/science.1193304 . hdl : 20.500.11820/fc52fb23-10e8-466d-a7e9-081260d166c6 . ПМИД 20847260 . S2CID 45978858 .
- ^ Снайвли, Э.; Хендерсон, DM; Филипс, Д.С. (2006). «Сросшиеся и сводчатые носовые кости динозавров-тираннозаврид: влияние на силу черепа и механику питания» (PDF) . Acta Palaeontologica Polonica . 51 (3): 435–454.
- ^ Рэйфилд, Э.Дж. (2004). «Черепная механика и питание тираннозавра рекса » . Труды Королевского общества Б. 271 (1547): 1451–1459. дои : 10.1098/rspb.2004.2755 . ПМК 1691752 . ПМИД 15306316 .
- ^ Беверидж, Теган Л.; Робертс, Эрик М.; Рамезани, Джахандар; Титус, Алан Л.; Итон, Джеффри Г.; Ирмис, Рэндалл Б.; Сертич, Джозеф JW (апрель 2022 г.). «Уточненная геохронология и пересмотренная стратиграфическая номенклатура верхнемеловой формации Вахвип, штат Юта, США, и возраст ранних кампанских позвоночных из южной Ларамидии» . Палеогеография, Палеоклиматология, Палеоэкология . 591 : 110876. Бибкод : 2022PPP...59110876B . дои : 10.1016/j.palaeo.2022.110876 . ISSN 0031-0182 .
- ^ Джинна, З.А. (2009). «Последовательность стратиграфического контроля на основе аллювиальной архитектуры речной системы верхнего мела - формация Вахвип, Южная Юта, США» (PDF) . Статья о поиске и открытии . п. 30088 . Проверено 10 декабря 2013 г.
- ^ Гейтс, штат Калифорния; Джинна, З.; Левитт, К.; Гетти, Массачусетс (2014). «Новые экземпляры гадрозавридов (Dinosauria, Ornithopoda) из нижней и средней кампанской формации Вахвип на юге Юты». В Эберте, Д.А.; Эванс, округ Колумбия (ред.). Гадрозавры: материалы Международного симпозиума по гадрозаврам . Издательство Университета Индианы. стр. 156–173. ISBN 978-0-253-01385-9 .
- ^ Лайвли, Дж. (2012). « Диаблоцератопс едятони » . Музей естественной истории штата Юта . Проверено 16 ноября 2013 г.
- ^ Гейтс, штат Калифорния; Хорнер, младший; Ханна, РР; Нельсон, ЧР (2011). «Новый гадрозаврин-гадрозаврид без украшений (Dinosauria, Ornithopoda) из кампана Северной Америки». Журнал палеонтологии позвоночных . 31 (4): 798. Бибкод : 2011JVPal..31..798G . дои : 10.1080/02724634.2011.577854 . S2CID 8878474 .
- ^ Киркланд, Дж.И.; ДеБлё, Д.Д. (2007). «Новые рогатые динозавры из формации Вахвип» (PDF) . Примечания к опросу . Геологическая служба Юты. стр. 4–5.
- ^ Томпсон, ЧР (2004). «Предварительный отчет о биостратиграфии пресноводных скатов мелового периода, формации Вахвип и члена формации Джона Генри формации Стрейт-Клифс, южная Юта» . Рефераты с программами Геологического общества Америки . 36 (5): 43. Архивировано из оригинала 17 декабря 2014 года . Проверено 10 декабря 2013 г.
- ^ Орсулак, М.; Симпсон; Вольф, Эл.; Симпсон, штат Вашингтон; Тиндалл, СС; Бернард, Дж.; Дженески, Т. (2007). «Нора двоякодышащей рыбы в верхнем меловом слое покрывающего песчаника в районе петушиного гребня формации Вахвип, Большая лестница – национальный памятник Эскаланта, Юта» . Рефераты с программами Геологического общества Америки . 39 (5): 43.
- ^ Итон, Дж. Г.; Чифелли, Р.Л. (2005). «Обзор палеонтологии млекопитающих мелового периода; Национальный памятник Большая лестница – Эскаланте, Юта». Рефераты с программами Геологического общества Америки . 37 (7): 115.
- ^ Тестер, Э.; Симпсон, Эл.; Вольф, Привет; Симпсон, штат Вашингтон; Тиндалл, СС; Бернард, Дж.; Дженески, Т. (2007). «Изолированные следы позвоночных из покрывающего песчаника верхнего мела формации Вахвип; Национальный памятник Большая лестница – Эскаланте, Юта» . Рефераты с программами Геологического общества Америки . 39 (5): 42. Архивировано из оригинала 17 декабря 2014 года . Проверено 10 декабря 2013 г.
- ^ ДеБлё, Д.Д. (2007). «Анализ местонахождения гадрозавров Джима; местонахождение динозавров в формации Вахвип среднего кампана (мелового периода) национального памятника Гранд-Стеркас-Эскаланте (GSENM), южная Юта» . Рефераты с программами Геологического общества Америки . 39 (5): 6. Архивировано из оригинала 17 декабря 2014 года . Проверено 10 декабря 2013 г.
- ^ Киркланд, Дж.И.; ДеБлё, Д.Д.; Хайден, М. (2005). «Инвентаризация палеонтологических ресурсов в нижней части формации Вахвеап (нижний кампан), южное плато Кайпаровиц, национальный памятник Большая лестница – Эскаланте, Юта» . Рефераты с программами Геологического общества Америки . 37 (7): 114. Архивировано из оригинала 17 декабря 2014 года . Проверено 10 декабря 2013 г.
- ^ Уильямс, Джей Джей; Лоренгель, CF (2007). «Предварительное исследование пресноводных брюхоногих моллюсков в формации Вахвип, национальный парк Брайс-Каньон, штат Юта» . Рефераты с программами Геологического общества Америки . 39 (5): 43. Архивировано из оригинала 17 декабря 2014 года . Проверено 10 декабря 2013 г.
Внешние ссылки
[ редактировать ]- Новое открытие динозавра: Lythronax argestes - видеопрезентация описателей Марка Лоуэна и Рэндалла Ирмиса
- Lythronax argestes : King of Gore — замедленное видео сборки скелета и модели головы















- Тираннозавриды
- Монотипические роды динозавров
- Позднемеловые динозавры Северной Америки.
- Кампанские роды
- Меловой период Юта
- Палеонтология в Юте
- Большая лестница – национальный памятник Эскаланте
- Ископаемые таксоны описаны в 2013 г.
- Таксоны, названные Филипом Дж. Карри
- Таксоны, названные Скоттом Д. Сэмпсоном