Мы узнали
| Мы узнали Временной диапазон: поздний мел , ~ | |
|---|---|
 | |
| Установленный голотип в Музее естественной истории Монголии в 2002 году (обратите внимание на неправильное количество хвостовых позвонков; в голотипе указано только четыре) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | † Орнитомимозаврия |
| Клэйд : | † Макрохейриформные |
| Суперсемейство: | † Орнитомимоидеа |
| Род: | † Мы узнали Барсболд , 1981 год. |
| Типовой вид | |
| † Мы коротконогие Барсболд, 1981 год. | |
Garudimimus (что означает « подражатель Гаруде ») — род орнитомимозавров , обитавший в Азии в позднем меловом периоде . Род известен по единственному экземпляру, обнаруженному в 1981 году советско-монгольской палеонтологической экспедицией в формации Баян-Шире и официально описанному в том же году Ринченом Барсболдом ; единственный вид — Garudimimus brevipes . несколько интерпретаций анатомических особенностей Garudimimus При задних исследованиях экземпляра было сделано , но большинство из них подверглись критике во время его всестороннего переописания в 2005 году. Обширные неописанные останки орнитомимозавра в типовом местонахождении Garudimimus могут представлять собой дополнительные экземпляры этого рода.
Единственным известным экземпляром гарудимимуса было животное среднего размера, длиной около 3,5 м (11,5 футов) и весом около 77,3–98 кг (170–216 фунтов). Это был орнитомимозавр со смесью базальных и производных черт; В отличие от примитивных орнитомимозавров, как верхняя, так и нижняя челюсти были беззубыми — черта, которая часто встречается у более продвинутых орнитомимид . У Гарудимимуса были относительно короткие и коренастые задние конечности, крепкие ступни и редуцированные подвздошные кости . На стопе было четыре пальца, первый из которых был сильно редуцирован, тогда как у орнитомимид было три пальца с отсутствующим первым пальцем. Беззубый череп имеет очень прямые челюсти, заканчивающиеся более закругленным кончиком морды, чем у других родов. Ранее считалось, что у этого примитивного орнитомимозавра был слезный «рог» наверху черепа, перед глазницей. Однако переописание единственного экземпляра показало, что эта структура представляла собой просто деформированную левую префронтальную кость . Другой ранней интерпретацией была реконструкция плюсны с помощью аркто-метатарзальное состояние.
При первом описании в 1981 году Garudimimus был идентифицирован как примитивный орнитомимозавр в своем собственном семействе. Однако при описании новых экземпляров Deinocheirus в 2014 году выяснилось, что последний является сестринским таксоном Garudimimus , группирующегося в составе Deinocheiridae — орнитомимозавров, не приспособленных к бегу или проворным движениям. Но такое размещение также оказалось маловероятным. Тазовый пояс и задние конечности Garudimimus показывают, что мускулатура ног была не так хорошо развита, как у быстро бегающих орнитомимид, что указывает, следовательно, на плохую к бегу способность . Как и другие представители Ornithomimosauria, Garudimimus был всеядным/травоядным животным с уменьшенной силой укуса, которая компенсировалась роговым клювом.
История открытия
[ редактировать ]
В 1981 году во время советско-монгольской палеонтологической экспедиции в пустыню Гоби относительно небольшой скелет теропода был обнаружен командой в Байши Цав в позднемеловой формации Баян Ширех , Юго-Восточная Монголия . Останки занесены в каталог как MPC-D 100/13 (Монгольский палеонтологический центр; первоначально ГИН 100/13) и представляют собой довольно полный и сочлененный скелет. кратко описан монгольским палеонтологом Ринченом Барсболдом как голотип рода В том же году этот экземпляр был формально и и вида Garudimimus brevipes . Общее название Garudimimus буддийской сочетает в себе отсылку к Гаруде , легендарным крылатым существам из монгольской мифологии , и латинскому mimus (что означает мимик). Видовое название происходит от латинского brevis (что означает короткий) и pes (что означает ступня), имея в виду короткую плюсну . Барсболд идентифицировал Garudimimus орнитомимозавров, как таксон но отметил, что он более примитивен, чем орнитомимиды , и выделил ему собственное семейство Garudimimidae. . [1] Барсболд описал дополнительные останки экземпляра голотипа в 1983 году, а затем в 1990 году вместе с Гальшкой Осмольской . [2] [3]
Голотип состоит из черепа целиком, 8 шейных позвонков (включая атлант и ось ), 9 спинных позвонков , 6 крестцовых позвонков и 4 хвостовых позвонков , некоторых ребер , подвздошных , лобковых , бедренных , большеберцовых , малоберцовых и практически полная левая стопа несовершеннолетней особи. [1] [4]
В 1988 году североамериканский автор Грегори С. Пол проиллюстрировал Гарудимимуса выступающим носовым рогом, в отличие от любого другого орнитомимозавра, считая эту особенность фактически сохранившейся. [5] Другая интерпретация была сделана также в 1988 году Филипом Дж. Карри и Дейлом Аланом Расселом, которые отнесли плюсну голотипа к Oviraptor sp. и реконструировали ее с помощью аркто-метатарзальной кости — состояния, при котором верхний конец третьей плюсневой кости сужен между окружающими плюсневыми костями — структурой. [6] В 1992 году Томас Р. Хольц последовал этой интерпретации и предположил, что плюсневая кость могла быть аркто-метатарзальной и была просто разобрана на сохранившуюся. [7] Подтверждая арктометатарзальное состояние, Карри и Дэвид А. Эберт в 1993 году утверждали, что часть материала Archaeornithomimus (орнитомимозавра из близлежащей формации Ирен-Дабасу ) принадлежала Garudimimus, основываясь на предполагаемом арктометатарзальном состоянии, наличии рудиментарного пальца I и пропорциях плюсневые кости II, III и IV. Они указали, что плюсневые кости раздавлены, а III плюсневая кость отодвинута назад от разгибательной поверхности плюсны. [8] В 1994 году Хольц предположил некоторое сходство между плюснами Garudimimus и Chirostenotes . [9]

В 1994 году Бернардино П. Перес-Морено с коллегами описал примитивного орнитомимозавра Pelecanimimus и выявил наличие гребня у экземпляра голотипа. есть аналогичная черта Они утверждали, что у Гарудимимуса , представленная носовым рогом. [10] В ходе всестороннего переописания, проведенного в 2005 году Ёсицугу Кобаяши и Барсболдом, многие предыдущие утверждения были опровергнуты. Предполагаемый орбитальный рог на самом деле представляет собой разъединенную левую префронтальную кость, и было подтверждено, что плюсневая кость не претерпела тафономических (изменений во время распада и окаменения) искажений и не является арктометатарзальной. Более того, Кобаяши ранее показал, что помимо фактического отсутствия аркто-метатарзального состояния, соотношения плюсневых костей отличаются от Archaeornithomimus . [4]
Хотя Garudimimus известен только из MPC-D 100/13, дополнительные экземпляры потенциально могут присутствовать среди большого костного пласта, состоящего как минимум из пяти особей в Байши Цав и нескольких неописанных орнитомимозавров из других местонахождений в формации Баян Шире. [11] [12] [13]
Описание
[ редактировать ]
Гарудимимус (на основе голотипа) оценивается в рост около 3,5 м (11,5 футов) и вес примерно 77,3–98 кг (170–216 фунтов). [14] [15] [16] Несколько несросшихся позвонков указывают на то, что голотип, вероятно, был еще растущим полувзрослым животным. [4] Некоторые из отличительных черт этого таксона включают сочленение челюсти, расположенное более назад, чем заглазничная перемычка (выгнутая кость вокруг глазницы , также называемая заглазничной ), углубления на верхней поверхности надзатылочной кости в задней части черепа, парные углубления. на латеральных поверхностях нервных отростков передних хвостовых позвонков и глубокая борозда у верхнего конца латеральной поверхности III-1 и III-2 педальных фаланг (первой и второй фаланг третьего пальца стопы). [4]
Судя по отпечаткам перьев нескольких экземпляров Ornithomimus , у Garudimimus, вероятно, развилось мохнатое оперение, в основном похожее на оперение крупных палеогнатных птиц, таких как бескилевые . [17] [18]
Череп
[ редактировать ]
Гарудимимус имел довольно маленький череп по сравнению с общим скелетом , а обе челюсти были беззубыми (лишенными зубов). Череп имел длину около 25 см (250 мм) и ширину 4,6 см (46 мм). Глазница представляла собой большую полость высотой 6,1 см (61 мм) и шириной. Радиус склеротического кольца (костных пластинок, поддерживающих глазное яблоко ) составлял примерно 2,86 см (28,6 мм) в длину внутри и 4,12 см (41,2 мм) снаружи. Ширина каждой пластины составляла около 0,8 см (8,0 мм). [4] [19] Подвисочное окно (отверстие за глазницей) имело неправильную, почти овальную форму. Перед глазницей располагалось предглазничное окно — большое отверстие довольно неправильной формы. Верхняя челюсть представляла собой удлиненную кость с шероховатой боковой поверхностью, что было обычным явлением у орнитомимозавров. Как промаксиллярные, так и верхнечелюстные фенестры (небольшие отверстия в верхней челюсти) были плохо развиты — почти отсутствовали — у Garudimimus . Нижние края предчелюстной кости тонкие и резко развитые. На латеральных сторонах этой кости многочисленные отверстия располагались (небольшие отверстия), что более характерно для производных орнитомимид . [4] Беззубое состояние как предчелюстной, так и зубной кости с добавлением многочисленных отверстий на их поверхности указывает на то, что выступающая рамфотека (роговой клюв). при жизни присутствовала [4] [19] Ноздри . умеренно длинные и обращены вверх Они были окружены и в основном образованы носовой костью , имевшей узкий конец, выступавший вверх над глазницами. [4]

Слезная кость имела полуТ-образную форму и располагалась перед глазом. Префронтальная кость представляла собой тонкую и нежную кость, которая определяла общую форму глазницы. На виде сверху лобные кости имели треугольную форму и образовывали верхние границы глазницы. Оба фронтала вместе образовали единую куполообразную структуру. Задний край глазницы образован заглазничной костью , представлявшей собой резко развитую кость хорошо распределённой толщины в боковой проекции. имела Скуловая кость треугольную основную часть с очень тонкостенными отростками (костными расширениями); оно также придало форму нижней границе глазницы. Квадратоскуловая кость представляла собой большую кость треугольной формы, контрастирующую с L-образной квадратноскуловой костью большинства орнитомимозавров. [4] На задней поверхности квадратной кости имелось большое углубление овальной формы, расположенное на середине ее высоты. небольшое пневматическое отверстие. В нижней части впадины также имелось [20] Дентарий был самым длинным элементом нижней челюсти длиной 18,3 см (183 мм) и утолщался к концу. Передняя часть обеих зубных костей имела лопатовидную форму, с хорошо развитыми режущими краями. На латеральных поверхностях кончика зуба развиты многочисленные отверстия. было Надугольное отверстие вторым по величине элементом нижней челюсти и образовывало верхнюю границу нижнечелюстного окна (отверстия в нижней челюсти). По сравнению с двумя последними костями угловая была значительно меньше и образовывала нижнюю границу этого окна. [4]
Посткраниальный скелет
[ редактировать ]
Полный позвоночник Гарудимимуса . неизвестен Межцентральная часть и дуга атланта нервная были направлены вниз. Центральная длина оси была довольно короткой: примерно 3 см (30 мм) в длину и 1,8 см (18 мм) в высоту — вероятно, примитивное состояние у орнитомимозавров. Его задняя суставная поверхность была вогнутой, ее высота превышала ширину. Его нервный отросток был высоким и очень округлым, а нервная дуга имела пневматическую ямку (впадину) посередине центра ( тела). Нервные отростки большинства шейных позвонков были узкими и направленными к концу. Спинные позвонки постепенно увеличивались в размерах к концу, и вместе с размером нервные отростки становились выше в задних спинных позвонках и имели почти одинаковую длину. При виде сбоку все шипы были слегка наклонены к концу. Дорсальные центры не имели пневматизации, были крупнее в более задних позвонках и имели амфицельные (сильно вогнутые) фасетки. [4]

Крестец . состоял из шести крестцовых позвонков, из которых первый и шестой имели крестцовые ребра, прикрепленные к внутренней поверхности подвздошной кости Общая длина крестца (31 см (310 мм)) была почти равна длине подвздошной кости. Большинство сакральных центров были примерно одинаковой длины, за исключением четвертого. В отличие от других орнитомимозавров, крестцовые позвонки не имели пневматизации (воздушного пространства). Хвостовые нервные отростки становились все короче и имели прямые и горизонтальные верхние границы. Хвостовые позвонки имели парные выступы по обе стороны от борозд (щелей) для шевронного сочленения. [4] Предполагаемые большие отверстия и глубокие ямки (впадины) присутствовали на передних нервных дугах, что может указывать на каудальную пневматичность у Garudimimus . [21]
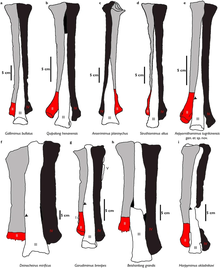
Ребра ( имели короткий бугорок (верхняя часть головы) и длинную головку (нижняя головка), а гастралии брюшные ребра) были сегментированы на латеральную и внутреннюю части. Большинство крестцовых ребер развиты слабо. Тазовый пояс образован подвздошной, лобковой и седалищной костями . Хотя последнее неизвестно Гарудимимусу . представляла Подвздошная кость собой большую кость, но короткую по сравнению с орнитомимидами, с крючкообразным передним концом. Лобок представлял собой крупный элемент длиной 39 см (390 мм). У него было почти прямое древко и выступающий лобковый ботинок (большой выступ на конце). Бедренная кость была относительно прямой и имела длину 37,1 см (371 мм). У него была хорошо развитая головка бедра и уплощенное вбок тело. И голень , и малоберцовая кость были почти равны по длине бедренной кости (36 см (360 мм) и 38,8 см (388 мм) соответственно). Большеберцовая кость была более уплощенной, чем другие элементы конечности, а стержень был очень тонким. Таранная кость и пяточная кость прикреплялись к нижним сторонам большеберцовой кости. [4]
По формуле pes (ступня) гарудимимус сильно отличался от состояния орнитомимида наличием пяти плюсневых костей . Первая плюсневая кость была наименьшим элементом плюсны и имела более уплощенную и защемленную форму. Вторая и четвертая плюсневые кости были примерно одинаковой длины, а третья плюсневая кость была самой длинной (22,9 см (229 мм) в длину). Последний не был защемлен на верхнем конце, что указывает на неаркто -метатарзальное состояние у Garudimimus . [4] Эта черта частично свойственна крупным Beishanlong и Deinocheirus . [22] Пятая плюсневая кость представляла собой очень редуцированный и тонкий элемент, не имевший функционального значения и прикреплявшийся к задней поверхности четвертой плюсневой кости. На ступнях было четыре примитивных пальца ( пальца ), первый из которых был самым маленьким. Большинство фаланг были очень сходны по форме, расширены на переднем и заднем концах с закругленными латеральными мыщелками и имели вогнутые передние суставные поверхности. Они становятся все короче от второго до четвертого пальца. [4] педалей Когтевые кости ( когтевые кости) имели загнутую форму, отличающуюся от прямой у орнитомимид, и выраженные бугорки сгибателей ( прикрепления сухожилий сгибателей ). [4] [22]
Классификация
[ редактировать ]
В 1981 году Барсболд отнес Garudimimus к отдельному Garudimimidae, учитывая необычные черты этого таксона. Однако он определил их как явно примитивные. [1] После переописания голотипа в 2005 году больше диагностических признаков гарудимимуса было предоставлено . Кобаяши и Барсболд отметили, что этот таксон имел общие черты с примитивными и производными орнитомимозаврами, такие как четырехпалые лапы и беззубые челюсти. анализ выявил Ornithomimidae как сестринский таксон Garudimimus Их филогенетический . [4] новые и довольно полные экземпляры давно загадочного дейнохейруса В 2014 году Юонг-Нам Ли и его коллеги описали . Останки показали, что Deinocheirus на самом деле был таксоном орнитомимозавров, но с отклонениями от нормы, а также позволили определить более старые Deinocheiridae как семейство, включающее все таксоны, имеющие более недавнего общего предка с Deinocheirus, чем с Ornithomimus . Ли и его команда провели большой филогенетический анализ Deinocheiridae, и в этой группе были обнаружены три члена: Beishanlong , Garudimimus и Deinocheirus . Их взаимоотношения в основном были представлены различными анатомическими особенностями конечностей, но последние два были сильно связаны, поскольку у них обоих были остатки черепа и тела. Полученная кладограмма позволила предположить, что орнитомимозавровые динозавры в раннем меловом периоде разделились на две основные линии: Deinocheiridae и Ornithomimidae. В отличие от быстро бегущих орнитомимид, дейнохейриды не были созданы для бега. Анатомические особенности дейнохейруса по сравнению с другими орнитомимозаврами можно объяснить заметно большими размерами, достигнутыми этим таксоном. [22]
Совсем недавно, в 2019 году, Хартман и его коллеги описали нового паравиана Hesperornithoides, проведя обширный филогенетический анализ целурозаврии . В случае орнитомимозавров Garudimimidae были обнаружены как семейство, включающее Garudimimus и его близких родственников. Garudimimus был обнаружен в довольно производном положении как сестринский таксон Beishanlong . Этот анализ показал, что тесная связь между Deinocheirus и Garudimimus маловероятна, поскольку первый был обнаружен у ближайшего основания Ornithomimosauria . [23]
Левая кладограмма Ли и его коллег в 2014 г. [22] правая кладограмма по Хартману и коллегам в 2019 году: [23]
|
|
В 2020 году Серрано-Браньяс и его команда описали и назвали новый род дейнохейрид Paraxenisaurus из формации Серро-дель-Пуэбло . Это первый член группы, обнаруженный за пределами Азии, и его открытие может указывать на то, что дейнохейриды возникли в Лавразии (северный суперконтинент во время мезозоя ) или что они рассеялись по полярным регионам в Северном полушарии ; Известно, что аналогичный обмен произошел и с другими группами динозавров, имеющими азиатское происхождение, в кампанско - маастрихтский период. Команда обнаружила, что Harpymimus — довольно примитивный дейнохейрид, однако Бэйшаньлун был помещен за пределы группы. Garudimimus и Deinocheirus были снова обнаружены как близкородственные таксоны с добавлением Paraxenisaurus в политомии . [24]
Палеобиология
[ редактировать ]Ежедневная активность
[ редактировать ]
Пропорции конечностей, морфология педальных когтей и удлинение подвздошной кости у Garudimimus позволяют предположить, что он не был приспособлен для скорости, как у более беглых орнитомимид , для которых характерны мощные бедра и прямые когти. [4] [22] В 2011 году Ларс Шмитц и Рёсуке Мотани измерили размеры склеротического кольца и глазницы у ископаемых экземпляров динозавров и птерозавров, а также некоторых ныне живущих видов. Они отметили, что в то время как дневные (фотопические) животные имеют меньшие склеротические кольца, ночные (скотопические) животные, как правило, имеют более крупные кольца. Катемерные (мезопические) животные, которые нерегулярно активны в течение дня и ночи, находятся между этими двумя диапазонами. Шмитц и Мотани разделили экологические и филогенетические факторы и, исследовав 164 ныне живущих вида, заметили, что измерения глаз довольно точны при определении дневного, катемерного или ночного образа жизни четвероногих . Результаты показали, что гарудимимус был катемерным травоядным животным, а другие хищные тероподы вели в основном ночной образ жизни. Было обнаружено, что крупные травоядные динозавры вели катемерный образ жизни, скорее всего, из-за длительного периода кормления и факторов терморегуляции . Птицы и птерозавры в основном вели дневной образ жизни, за некоторыми ночными исключениями. Они пришли к выводу, что экологическая ниша была основной движущей силой развития диэлитной деятельности. [25]
Однако в том же году этот анализ подвергся критике со стороны Холла и его коллег, выступавших против методов, использованных Шмитцем и Мотани, поскольку существует значительная степень сходства в анатомии склеры между дневными, катемерными и ночными животными. Более того, мягкие ткани , такие как роговица, необходимы для определения того, сколько света может попасть в глаз, чего явно нет у ископаемых образцов. [26] Шмитц и Мотани ответили на этот комментарий, подтвердив, что их исследования и методология правильно классифицировали поведение вымерших динозавров на основе существующих видов. Они назвали внешнюю границу склерального кольца надежным источником при реконструкции зрительных способностей. [27]
Чувства
[ редактировать ]В 2019 году Грэм М. Хьюз и Джон А. Финарелли проанализировали соотношение обонятельных луковиц у современных птиц и сохранили черепа нескольких вымерших видов динозавров, чтобы предсказать, сколько генов могло быть задействовано в обонятельной силе этих вымерших видов. Их анализ показал, что у большинства орнитомимозавров было относительно слабое обоняние; У гарудимимуса было примерно 417 генов, кодирующих его обонятельные рецепторы, а соотношение обонятельных луковиц составляло 28,8, что указывает на плохо развитое обоняние. Учитывая, что эти значения были более выражены у более крупных видов динозавров, таких как тираннозавриды , Хьюз и Финарелли отметили, что по мере того, как линии динозавров становились крупнее, размер обонятельной луковицы также увеличивался, что может указывать на то, что обоняние является доминирующей сенсорной модальностью у нептичьих видов. динозавры. [28]
Привычки питания
[ редактировать ]
Кобаяши и Барсболд заявили в 2005 году, что отсутствие зубных рядов на нижней челюсти и их изогнутая форма указывают на то, что гарудимимусы были гораздо больше похожи на орнитомимид при потреблении пищи, чем на других примитивных орнитомимозавров. Как и у орнитомимид, передние части верхней и нижней челюстей Garudimimus были покрыты клювом и, возможно, использовались для ощипывания пищи. Кроме того, острые края средних частей челюстей помогали резать пищу. [4] Более того, Garudimimus и его родственники широко считаются травоядными или, по крайней мере, всеядными динозаврами из-за их особых анатомических особенностей, включая наличие клюва. [29] [25]
В 2015 году Эндрю Р. Кафф и Эмили Дж. Рэйфилд выполнили компьютерную томографию искаженных тафономически образцов черепов Гарудимима , Орнитомима и Струтиомима с целью ретродеформации — метода, используемого для получения исходного состояния деформированного объекта. Сканирование и относительная полнота образцов позволили реконструировать приводящую (индуцирующую движение) мускулатуру нижней челюсти, которая затем была нанесена на карту на черепах. Кроме того, размеры рамфотеки (клюва) были реконструированы на мелкие (на основе бескилевых птиц и страуса ) и крупные морфы. Среди трех изученных родов Garudimimus имел самую низкую силу укуса: было обнаружено, что он производит 19 Н на кончике клюва и 23,9 Н на его средней длине. Однако у этого таксона было наиболее модифицированное механическое преимущество мышц и плечо момента — мера эффективности участия мышцы в определенном движении в ряде конфигураций. С учетом этого наиболее значительная мышечная разница между Было обнаружено, что у гарудимимуса и двух орнитомимид большинство мышц в последних двух более выгодны с механической точки зрения; это, вероятно, вызвано более длинным черепом у Garudimimus . Кафф и Рэйфилд отметили, что сила укуса Гарудимимуса может быть ограничена из-за необходимости использовать нижнюю челюсть Струтиомима при реконструкциях или что MPC-D 100/13 является несовершеннолетним человеком. Предварительно они согласились с тем, что относительно небольшая сила укуса в сочетании с рамфотекой может использоваться для удержания растительного материала во время кормления орнитомимозавров, и все это в то время, как мускулатура шеи обеспечивает достаточную силу для выдергивания растительности . [19]

В 2019 году Дэвид Дж. Баттон и Линдси Э. Занно провели большой филогенетический анализ, основанный на биомеханических характеристиках черепа, предоставленных 160 видами мезозойских динозавров, чтобы проанализировать множественные проявления травоядных среди нептичьих динозавров. Их результаты показали, что травоядные динозавры в основном придерживались двух различных способов питания: либо переработка пищи в кишечнике, характеризующемся относительно изящными черепами и низкой силой укуса, либо рот, который характеризовался особенностями, связанными с интенсивной переработкой, такими как высокая сила укуса и крепкая челюстная мускулатура. Было обнаружено, что Garudimimus и Deinocheirus , наряду с динозаврами-ценагнатидами , диплодокоидами , орнитомимидами , теризинозаврами и титанозаврами , относятся к первой категории, что указывает на то, что Garudimimus действительно обладал низкой силой укуса и в основном полагался на свой желудок для переработки пищи. Большие размеры тела были достигнуты независимо у дейнохейрид и орнитомимид, а более крупные размеры дают преимущества травоядным животным, такие как повышенная устойчивость к голодание и большее количество приема пищи . Следовательно, эти привычки могут указывать на более травоядный образ жизни дейнохейрид и орнитомимид. Однако Баттон и Занно отметили, что взаимосвязь между травоядностью и размером не всегда связана, и не существует прямой тенденции к увеличению массы у орнитомимозавров. Более того, определенные ниши для большинства орнитомимозавровых динозавров использовались плохо, и только дейнохейрус демонстрировал оппортунистическое всеядное существование. Баттон и Занно пришли к выводу, что стратегии питания у неродственных травоядных динозавров с течением времени эволюционировали конвергентно. [30]
В том же году Али Набавизаде изучил мускулатуру челюстей травоядных динозавров и пришел к выводу, что некоторые из них в основном питались ортально — двигали челюстями вверх и вниз — в том числе тероподы , зауроподоморфы , примитивные птицетазовые животные и некоторые стегозавры . Набавизаде отметил, что орнитомимозавры питались ортально и поднимали челюсти изогнатно, при этом верхние и нижние зубы каждой стороны смыкались (контактировали друг с другом) одновременно. Deinocheirus и Garudimimus имеют уникальные места прикрепления мышц, открывающих и закрывающих челюсти, которые состоят из расширенного вниз треугольного комплекса приводящих мышц, прикрепляющегося к расширяющейся вперед надугольной кости . Это, в сочетании с расширенным скуловым фланцем, обеспечивает большую полость для приводящей мышцы. При такой удлиненной морде сила укуса у обоих дейнохейрид была низкой с меньшим механическим преимуществом, однако, если бы эта мышца простиралась дальше вперед, это добавило бы силы височной мышце . [31]
Палеосреда
[ редактировать ]
Известные останки Гарудимимуса были обнаружены в позднемеловой формации Баян Шире в местонахождении Байши Цав. [1] Эта свита сложена преимущественно аргиллитами и песчаниками с конкрециями и характеризуется аргиллитами и желтовато-коричневыми среднезернистыми песчаниками. Наличие калише, речных и озерных отложений в различных местах указывает на то, что это образование формировалось извилистыми реками и другими крупными водоемами на аллювиальной равнине (равнине, состоящей из отложений горных рек) в условиях полузасушливого климата. [32] [33] Предполагается, что сам Байши Цав отложился в речных условиях. [34] На основе кальцита U-Pb измерений возраст формации Баян Шире был датирован примерно от 95,9 ± 6,0 миллиона до 89,6 ± 4,0 миллиона лет назад на сеноман - сантонском этапе. [35]
Многочисленные чешуйчатые поперечные наслоения в местонахождении Бейншир являются индикаторами крупных меандров, и подавляющее большинство этих водоемов, возможно, дренировали восточную часть пустыни Гоби . [33] Наличие такой влажной среды подтверждается открытием водных рептилий, таких как черепахи , крокодиломорфы и остракоды . [36] [37] На основании сходных комплексов ископаемых Баян Шире и Ирен Дабасу . многие авторы предлагали корреляцию между формациями [32] [8] [38] Кроме того, формация Ирен-Дабасу датируется примерно 95,8 ± 6,2 миллиона лет назад. [39]
Гарудимимус разделял свою среду обитания с многочисленными динозаврами фауны , такими как травоядные птицетазовые Gobihadros , Graciliceratops и Talarurus . [40] [41] Остальные тероподы представлены хищником некрупной добычи Ахиллобатором , [42] и теризинозавриды Erlikosaurus и Segnosaurus . [43] Эркету, возможно, был самым высоким травоядным животным в фауне. [44] Несколько видов были разделены путем дифференциации ниш , например, эрликозавр и большой сегнозавр . [43] или травоядный Таларурус и браузер Цагантегия , еще один анкилозаврид из формации. [41] С точки зрения биоразнообразия , теризинозавры были одними из самых многочисленных теропод с множеством описанных и неописанных экземпляров по всей формации. [37] Однако останки гадрозавроидов, особенно в Бейншире, также многочисленны и хорошо известны. [34] [45] Хотя Garudimimus в настоящее время является единственным названным таксоном орнитомимозавров, формация Баян Шире привела к появлению местных многочисленных орнитомимозавров в Байши Цав. [46] [47] [11]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Barsbold, R. (1981). "Беззубые хищные динозавры Монголии" [Toothless carnivorous dinosaurs of Mongolia]. Transactions, Joint Soviet–Mongolian Palaeontological Expedition (in Russian). 15 : 28−39. S2CID 195060070 . Translated paper
- ^ Barsbold, R. (1983). "Хищные динозавры мела Монголии" [Carnivorous dinosaurs from the Cretaceous of Mongolia] (PDF) . Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 19 : 1−120. Translated paper
- ^ Барсболд, Р.; Осмольска, Х. (1990). «Орнитомимозаврия». В Вейшампеле, Д.Б.; Осмольская, Х.; Додсон, П. (ред.). Динозаврия (1-е изд.). Беркли: Издательство Калифорнийского университета. стр. 225−244. ISBN 9780520067271 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Кобаяши, Ю.; Барсболд, Р. (2005). «Повторное исследование примитивного орнитомимозавра Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda) из позднего мела Монголии» (PDF) . Канадский журнал наук о Земле . 42 (9): 1501–1521. Бибкод : 2005CaJES..42.1501K . дои : 10.1139/e05-044 . hdl : 2115/14579 .
- ^ Пол, GS (1988). «Загадочные мимики страуса» . Хищные динозавры мира . Саймон и Шустер. п. 383. ИСБН 978-0-671-61946-6 .
- ^ Карри, Пи Джей; Рассел, Д.А. (1988). «Остеология и взаимоотношения Chirostenotes pergracilis (Saurischia, Theropoda) из формации реки Джудит (Олдман) в Альберте, Канада» Канадский журнал наук о Земле . 25 (7): 272–286. Бибкод : 1988CaJES..25..972C . дои : 10.1139/e88-097 .
- ^ Хольц, Т.Р. (1992). Необычное строение плюсны теропод (Archosauria: Dinosauria: Saurischia) мелового периода (кандидатская диссертация). Йельский университет. п. 347.
- ^ Jump up to: а б Карри, Пи Джей; Эберт, Д.А. (1993). «Палеонтология, седиментология и палеоэкология формации Ирен-Дабасу (верхний мел), Внутренняя Монголия, Китайская Народная Республика». Меловые исследования . 14 (2): 127–144. дои : 10.1006/cres.1993.1011 .
- ^ Хольц, Т.Р. (1994). «Аркто-метатарзальная нога, необычное строение плюсны мелового теропода (Dinosauria: Saurischia)» . Журнал палеонтологии позвоночных . 14 (4): 480–519. дои : 10.1080/02724634.1995.10011574 . JSTOR 4523590 .
- ^ Перес-Морено, BP; Санс, Дж.Л.; Бускалиони, AD; Мораталла, Джей-Джей; Ортега, Ф.; Раскин-Гутман, Д. (1994). «Уникальный многозубый динозавр-орнитомимозавр из нижнего мела Испании» (PDF) . Природа . 370 (6488): 363–367. Бибкод : 1994Natur.370..363P . дои : 10.1038/370363a0 . S2CID 4302245 .
- ^ Jump up to: а б Чинзориг, Т.; Кобаяши, Ю.; Санеоши, М.; Цогтбаатар, К.; Батамхатан, З.; Рюдзи, Т. (2017). «Мультитаксическое костное ложе двух новых орнитомимид (Theropoda, Ornithomimosauria) из верхнемеловой баянширской формации юго-восточной пустыни Гоби, Монголия». Журнал палеонтологии позвоночных . Программа и тезисы: 97.
- ^ Чинзориг, Т.; Кобаяши, Ю.; Цогтбаатар, К.; Карри, Пи Джей; Такасаки, Р.; Танака, Т.; Иидзима, М.; Барсболд, Р. (2018). «Орнитомимозавры из немегтинской свиты Монголии: морфологическая изменчивость и разнообразие кисти руки» . Палеогеография, Палеоклиматология, Палеоэкология . 494 : 91–100. Бибкод : 2018PPP...494...91C . дои : 10.1016/j.palaeo.2017.10.031 .
- ^ Цогтбаатар, К. (2019). Эволюция, разнообразие и неравенство орнитомимозавров (Dinosauria: Theropoda) верхнего мела Монголии (PDF) (кандидатская диссертация). Университет Хоккайдо. hdl : 2115/74432 .
- ^ Темпертон, Дж.; Роузуорн, Г.; Киндерсли, Д. (1993). Киндерсли, Д. (ред.). Визуальный словарь динозавров . Визуальные словари очевидцев. Университет штата Пенсильвания: ДК ДЕТИ. стр. 1−64. ISBN 9780751310122 . LCCN 92053446 . ОСЛК 38325967 .
- ^ Занно, Ле; Маковицкий, П.Дж. (2013). «Нет доказательств направленной эволюции массы тела травоядных динозавров-теропод» . Труды Королевского общества Б. 280 (1751): 20122526. doi : 10.1098/rspb.2012.2526 . ПМЦ 3574415 . ПМИД 23193135 .
- ^ Чинзориг, Цогтбаатар; Каллен, Томас; Филлипс, Джордж; Рольке, Ричард; Занно, Линдси Э. (19 октября 2022 г.). «Крупнотелые орнитомимозавры населяли Аппалачи в позднем меловом периоде Северной Америки» . ПЛОС ОДИН . 17 (10). е0266648. дои : 10.1371/journal.pone.0266648 . ПМЦ 9581415 .
- ^ Зеленицкий, ДК; Терриен, Ф.; Эриксон, генеральный директор; ДеБур, CL; Кобаяши, Ю.; Эберт, Д.А.; Хэдфилд, Ф. (2012). «Пернатые нептичьи динозавры из Северной Америки дают представление о происхождении крыльев». Наука . 338 (6106): 510–514. Бибкод : 2012Sci...338..510Z . дои : 10.1126/science.1225376 . ПМИД 23112330 . S2CID 2057698 .
- ^ Ван дер Реест, AJ; Вулф, AP; Карри, Пи Джей (2016). «Плотно оперенный орнитомимид (Dinosauria: Theropoda) из формации Парка динозавров верхнего мела, Альберта, Канада». Меловые исследования . 58 : 108–117. CiteSeerX 10.1.1.710.3240 . дои : 10.1016/j.cretres.2015.10.004 .
- ^ Jump up to: а б с Манжета, Арканзас; Рэйфилд, Э.Дж. (2015). «Ретродеформация и мышечная реконструкция черепов орнитомимозавров динозавров» . ПерДж . 3 : е1093. дои : 10.7717/peerj.1093 . ПМЦ 4512775 . ПМИД 26213655 .
- ^ Хендрикс, К.; Араужо, Э.; Матеус, О. (2015). «Квадрат I нептичьего теропода: стандартизированная терминология с обзором анатомии и функций» . ПерДж . 3 : е1245. дои : 10.7717/peerj.1245 . ПМЦ 4579021 . ПМИД 26401455 .
- ^ Ватанабэ, А.; Золото, МЭЛГ; Брусатте, СЛ; Бенсон, РБЖ; Шуаньер, Ж.; Дэвидсон, А.; Норелл, Массачусетс (2015). «Пневматичность позвонков у орнитомимозавра Archaeornithomimus (Dinosauria: Theropoda), выявленная с помощью компьютерной томографии и переоценка осевой пневматичности у орнитомимозаврии» . ПЛОС ОДИН . 10 (12): e0145168. Бибкод : 2015PLoSO..1045168W . дои : 10.1371/journal.pone.0145168 . ПМЦ 4684312 . ПМИД 26682888 .
- ^ Jump up to: а б с д и Ли, Ю.-Н.; Барсболд, Р.; Карри, Пи Джей; Кобаяши, Ю.; Ли, Х.-Дж.; Годфруа, П.; Скалли, Ф.; Цогтбаатар, К. (2014). «Разгадка давней загадки гигантского орнитомимозавра Deinocheirus mirificus » Природа 515 (7526): 257–260. Бибкод : 2014Nature.515..257L . дои : 10.1038/nature13874 . ПМИД 25337880 . S2CID 2986017 .
- ^ Jump up to: а б Хартман, С.; Мортимер, М.; Валь, WR; Ломакс, ДР; Липпинкотт, Дж.; Лавлейс, DM (2019). «Новый паравианский динозавр из поздней юры Северной Америки подтверждает позднее приобретение птичьего полета» . ПерДж . 7 : е7247. дои : 10.7717/peerj.7247 . ПМК 6626525 . ПМИД 31333906 .
- ^ Серрано-Браньяс, CI; Эспиноза-Чавес, Б.; Маккракен, ЮАР; Гутьеррес-Бландо, К.; Леон-Давила, К.; Вентура, Дж. Ф. (2020). «Paraxenisaurusnormalensis, крупный дейнохейрид-орнитомимозавр из формации Серро-дель-Пуэбло (верхний мел), Коауила, Мексика». Журнал южноамериканских наук о Земле . http://dx.doi.org/10.1037/0021-843X.101.102610 Бибкод : 2020JSAES.101j2610S . дои : 10.1016/j.jsames.2020.102610 . S2CID 218968100 .
- ^ Jump up to: а б Шмитц, Л.; Мотани, Р. (2011). «Ночной образ жизни динозавров, выведенный на основании морфологии склерального кольца и орбиты». Наука . 332 (6030): 705–708. Бибкод : 2011Sci...332..705S . дои : 10.1126/science.1200043 . ПМИД 21493820 . S2CID 33253407 .
- ^ Холл, Мичиган; Кирк, ЕС; Камилар, Дж. М.; Каррано, Монтана (2011). «Комментарий к ночному образу жизни динозавров, сделанный на основе морфологии склерального кольца и орбиты» . Наука . 334 (6063): 1641. Бибкод : 2011Sci...334.1641H . дои : 10.1126/science.1208442 . ПМИД 22194558 .
- ^ Шмитц, Л.; Мотани, Р. (2011). «Ответ на комментарий о ночном образе жизни динозавров, выведенном на основании морфологии склерального кольца и орбиты» . Наука . 334 (6063): 1641. Бибкод : 2011Sci...334.1641S . дои : 10.1126/science.1208489 . ПМИД 22194558 .
- ^ Хьюз, генеральный менеджер; Финарелли, Дж. А. (2019). «Размер репертуара обонятельных рецепторов динозавров» . Труды Королевского общества Б. 286 (1904): 20190909. doi : 10.1098/rspb.2019.0909 . ПМК 6571463 . PMID 31185870 .
- ^ Барретт, премьер-министр (2005). «Питание динозавров-страусов (Theropoda: Ornithomimosauria)» . Палеонтология . 48 (2): 347–358. дои : 10.1111/j.1475-4983.2005.00448.x .
- ^ Баттон, диджей; Занно, LE (2019). «Повторяющаяся эволюция дивергентных способов травоядности у нептичьих динозавров» . Современная биология . 30 (1): 158−168.e4. дои : 10.1016/j.cub.2019.10.050 . ПМИД 31813611 . S2CID 208652510 .
- ^ Набавизаде, А. (2019). «Черепная мускулатура травоядных динозавров: обзор реконструированного анатомического разнообразия и механизмов питания» . Анатомическая запись . 303 (4): 1104–1145. дои : 10.1002/ar.24283 . ПМИД 31675182 . S2CID 207815224 .
- ^ Jump up to: а б Ежикевич, Т.; Рассел, Д.А. (1991). «Позднемезозойская стратиграфия и позвоночные животные бассейна Гоби». Меловые исследования . 12 (4): 345–377. дои : 10.1016/0195-6671(91)90015-5 .
- ^ Jump up to: а б Хикс, Дж. Ф.; Бринкман, Д.Л.; Николс, диджей; Ватабе, М. (1999). «Палеомагнитный и палинологический анализ отложений от альба до сантона в Байн Шире, Бурхант и Хурен Дух, восточная часть пустыни Гоби, Монголия» . Меловые исследования . 20 (6): 829–850. дои : 10.1006/cres.1999.0188 .
- ^ Jump up to: а б Исигаки, С.; Цогтбаатар, К.; Санеоши, М.; Майнбаяр, Б.; Аоки, К.; Ульзицерен, С.; Имаяма, Т.; Такахаши, А.; Тойота, С.; Баярдорж, К.; Буянтегш, Б.; Бацух Дж.; Пуревсурэн, Б.; Асаи, Х.; Цутанага, С.; Фуджи, К. (2016). «Отчет совместной экспедиции Научного университета Окаямы – Монгольского института палеонтологии и геологии в 2016 году» (PDF ) Вестник НИИ естествознания (42): 33−46.
- ^ Курумада, Ю.; Аоки, С.; Аоки, К.; Като, Д.; Санеоши, М.; Цогтбаатар, К.; Уиндли, БФ; Исигаки, С. (2020). «Кальцитовый U – Pb возраст меловой формации Баин Шир, содержащей позвоночных, в восточной пустыне Гоби в Монголии: полезность каличе для определения возраста» . Терра Нова 32 (4): 246–252. Бибкод : 2020Ноябрь..32..246K дои : 10.1111/тер.12456 .
- ^ Данилов И.Г.; Хираяма, Р.; Суханов В.Б.; Сузуки, С.; Ватабе, М.; Витек, Н.С. (2014). «Меловые мягкопанцирные черепахи (Trionychidae) Монголии: новое разнообразие, записи и пересмотр». Журнал систематической палеонтологии . 12 (7): 799–832. дои : 10.1080/14772019.2013.847870 . S2CID 86304259 .
- ^ Jump up to: а б Ли, Ю.М.; Ли, HJ; Кобаяши, Ю.; Карабахал, АП; Барсболд, Р.; Фиорилло, Арканзас; Цогтбаатар, К. (2019). «Необычное поведение при передвижении, сохранившееся на крокодилообразных путях из верхнемеловой баянширской формации Монголии, и его палеобиологические последствия». Палеогеография, Палеоклиматология, Палеоэкология . 533 (109353): 2. Бибкод : 2019PPP...533j9239L . дои : 10.1016/j.palaeo.2019.109239 . S2CID 197584839 .
- ^ Аверьянов А.; Сьюс, Х. (2012). «Корреляция комплексов континентальных позвоночных позднего мела в Средней и Центральной Азии» (PDF) . Журнал стратиграфии . 36 (2): 462–485. S2CID 54210424 . Архивировано из оригинала (PDF) 7 марта 2019 г.
- ^ Го, ZX; Ши, Ю.П.; Ян, Ю.Т.; Цзян, SQ; Ли, Л.Б.; Чжао, ЗГ (2018). «Инверсия Эрлианского бассейна (северо-восток Китая) в раннем позднем мелу: последствия столкновения Охотоморского блока с Восточной Азией» (PDF) . Журнал азиатских наук о Земле . 154 : 49–66. Бибкод : 2018JAESc.154...49G . дои : 10.1016/j.jseaes.2017.12.007 . Архивировано из оригинала (PDF) 19 сентября 2020 г. Проверено 2 сентября 2020 г.
- ^ Цогтбаатар, К.; Вейшампель, Д.Б.; Эванс, округ Колумбия; Ватабе, М. (2019). «Новый гадрозавроид (Dinosauria: Ornithopoda) из позднемеловой байнширской формации пустыни Гоби (Монголия)» . ПЛОС ОДИН . 14 (4): e0208480. Бибкод : 2019PLoSO..1408480T . дои : 10.1371/journal.pone.0208480 . ПМК 6469754 . ПМИД 30995236 .
- ^ Jump up to: а б Пак, JY; Ли, Ю.Н.; Карри, Пи Джей; Кобаяши, Ю.; Коппельхус, Э.; Барсболд, Р.; Матеус, О.; Ли, С.; Ким, SH (2020). «Дополнительные черепа Talarurus plicatospineus (Dinosauria: Ankylosauridae) и значение для палеобиогеографии и палеоэкологии панцирных динозавров». Меловые исследования . 108 : 104340. doi : 10.1016/j.cretres.2019.104340 . S2CID 212423361 .
- ^ Пауэрс, Массачусетс; Салливан, К.; Карри, Пи Джей (2020). «Пересмотр соотношения предчелюстных и верхнечелюстных признаков у Eudromaeosauria (Dinosauria: Theropoda): расходящиеся тенденции в морфологии морды между азиатскими и североамериканскими таксонами». Палеогеография, Палеоклиматология, Палеоэкология . 547 (109704): 109704. Бибкод : 2020PPP...54709704P . дои : 10.1016/j.palaeo.2020.109704 . S2CID 216499705 .
- ^ Jump up to: а б Занно, Ле; Цогтбаатар, К.; Чинзориг, Т.; Гейтс, Т.А. (2016). «Специализации анатомии нижней челюсти и зубочелюстного аппарата Segnosaurus galbinensis (Theropoda: Therizinosauria) » ПерДж . 4 : е1885. дои : 10.7717/peerj.1885 . ПМЦ 4824891 . ПМИД 27069815 .
- ^ Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 223−224. ISBN 9780691167664 .
- ^ Словиак, Дж.; Щигельский, Т.; Гинтер, М.; Фостович-Фрелик, Л. (2020). «Непрерывный рост неполярного гадрозавра объясняет гигантизм среди утконосых динозавров». Палеонтология . 63 (4): 579–599. дои : 10.1111/пала.12473 . S2CID 213247742 .
- ^ Ватабе, М.; Сузуки, С. (2000). «Отчет о совместной палеонтологической экспедиции Японии и Монголии в пустыню Гоби, 1993 г.». Бюллетень исследований Музея естественных наук Хаясибары . 1 :19−29.
- ^ Ватабе, М.; Сузуки, С. (2000). «Отчет о совместной палеонтологической экспедиции Японии и Монголии в пустыню Гоби, 1994 г.». Бюллетень исследований Музея естественных наук Хаясибары . 1 :30−44.
Внешние ссылки
[ редактировать ] СМИ, связанные с Гарудимимусом, на Викискладе?
СМИ, связанные с Гарудимимусом, на Викискладе?  Данные, относящиеся к Гарудимимусу , в Wikispecies
Данные, относящиеся к Гарудимимусу , в Wikispecies - 3D-голотип черепа на Phenome10k
- 3D-реконструированный голотип черепа на Phenome10k
- Смонтированный голотип на YouTube














