Рукавицы
| Рукавицы Временной диапазон: ранний мел. ~ 125–120 млн лет назад - поздний | |
|---|---|
 | |
| Гипотетическая реконструкция жизни трёх киннаримимов | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | Манирапториформы |
| Клэйд : | † Орнитомимозаврия |
| Род: | † Воспоминание о перчатках Баффетаут и др. , 2009 г. |
| Типовой вид | |
| † Киннаримимус хонкаененсис Баффетаут и др. , 2009 г. | |
| Синонимы | |
| |
Kinnareemimus ( / ˌ k ɪ n ə r i ˈ m ɑː i m ə s / , что означает « подражатель киннари », в честь фигуры из тайского фольклора) — вымерший род орнитомимозавровых . - динозавров теропод , который был обнаружен в в раннем меловом периоде формации Сан Хуа на территории современного Таиланда . Род содержит только вид типовой K. khonkaenensis . Видовой эпитет взят из провинции Кхон Каен , которая находится на северо-востоке Таиланда, где были обнаружены останки животного. [1]
Открытие и присвоение имени
[ редактировать ]
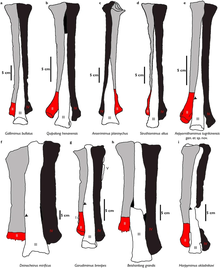
орнитомимозавров Окаменелости тел и ихнофоссилии были обнаружены на Таиланде в плато Корат еще в 1997 году в публикации Эрика Буффето и его коллег. В то время только два рода динозавров: Siamotyrannus и Phuwiangosaurus в стране были названы . В их отчет включен рисунок плюсневых костей орнитомимозавра, обнаруженного в местности под названием «Фу Вианг 5». Эти плюсневые кости ясно демонстрировали производное арктометатазальное состояние, подобное орнитомимозаврам, известным из Ларамидии . Однако этот экземпляр не был идентифицирован под каким-либо другим именем, кроме «Тайского орнитомимозавра». [2]
Очередное упоминание о «тайском орнитомимозавре» [3] была в первом выпуске журнала Dinosaur Press, вышедшего в Японии в 2000 году. [3] В этой публикации таксон упоминается под именем nomen nudum «Ginnareemimus» Рюичи Канеко, который написал в журнале статью, содержащую недавнюю информацию о палеонтологии в Таиланде . [4] [5]
Kinnareimimus Официальное описание вида получило в 2009 году , когда Эрик Баффетаут, Варавуд Сутитхорн и Хайян Тонг опубликовали анатомическую монографию в бюллетене Лондонского геологического общества . Все кости, относящиеся к орнитомимозаврам, найденным на Фу Вианге 5, были разрознены, и поэтому невозможно точно узнать, сколько отдельных животных было окаменело на этом месте. По этой причине, когда Kinnareemimus был описан в 2009 году, только одна частичная плюсневая кость III, обозначенная PW5A-100, была обозначена как голотип . Оставшимся экземплярам были присвоены уникальные обозначения (от 101 до 131) и каждый из них был отнесен к новому роду. Баффетаут и Сютитхорн в своем описании отмечают, что они уверены в принадлежности всего материала к одному виду, но консервативно не включили анатомию этих костей в диагностику вида. [1]
Когда он был описан, авторы назвали род в честь Киннари , существа из фольклора тайского народа , имеющего голову и туловище женщины, а также ноги птицы и живущего в легендарном лесу Химмапан. Баффетот и Сютитхорн отмечают, что выбор Киннари был сделан в отношении птичьих ног Киннаримимуса . [1]
Описание
[ редактировать ]Поскольку голотип Kinnareimimus . настолько фрагментарен (состоит только из одной плюсневой кости ), оценки размера животного всегда были весьма спекулятивными и неопределенными Авторы, описавшие род, Эрик Бюффето и его коллеги, не дали оценку массы животного. Они заметили, что соотношение длин голени и третьей плюсневой кости у Kinnareemimus наиболее сходно с Garudimimus . Они интерпретируют это как означающее, что пропорции его тела могли быть более похожими на этот род, чем на производные роды, такие как Gallimimus или Struthiomimus . [1] Впоследствии Молина-Перес и Ларраменди подсчитали, что его длина составляла около 1,8 метра (5,9 футов) и, возможно, весила около 13 килограммов (29 фунтов). [6]
Голотип
[ редактировать ]| Образец | Включает |
|---|---|
| ПВ5А-100 | дистальная часть III левой плюсневой кости |
| ПВ5А-101 | левая плюсневая кость II |
| ПВ5А-102 | проксимальная часть левой плюсневой кости IV |
| ПВ5А-103 | средняя часть III левой плюсневой кости |
| ПВ5А-104 | проксимальная часть III левой плюсневой кости |
| ПВ5А-105 | большая часть левой плюсневой кости II |
| ПВ5А-106 | правая плюсневая кость IV |
| ПВ5А-107 | дистальная часть III правой плюсневой кости |
| ПВ5А-108 | проксимальная часть IV плюсневой кости |
| ПВ5А-109 | проксимальная часть IV плюсневой кости |
| ПВ5А-110 | правая большеберцовая кость |
| PW5A-111 | левая большеберцовая кость |
| PW5A-112 | проксимальная часть левой малоберцовой кости |
| ПВ5А-113 | проксимальная часть правой лобковой кости |
| PW5A-114 | проксимальная часть левого лобка |
| PW5A-115–121 | педальные фаланги |
| ПВ5А-122 | частичная педальная фаланга с когтевой костью |
| ПВ5А-123 | спинной позвоночный центр |
| ПВ5А-124 | частичный хвостовой позвонок |
| ПВ5А-125 | средний каудальный центр позвонка |
| PW5A-126–129 | дистальные хвостовые позвонки |
| ПВ5А-130 | проксимальный каудальный центр позвонка |
| ПВ5А-131 | проксимальная часть III правой плюсневой кости |
Голойтпе Киннаримимуса (PW5A-100) представляет собой частично полную третью плюсневую кость , сохраняющую суставную поверхность с левой стороны. Неполнота образца помешала Бюффето и его коллегам оценить полный размер кости. Однако другой экземпляр (PW5A-101) представляет собой вторую плюсневую кость такого же размера и прочности, но гораздо более полную. Авторы, описывающие его, полагают, что он мог принадлежать одному и тому же человеку, и поэтому они использовали его, чтобы оценить, что полная длина третьей плюсневой кости при жизни составляла примерно 145 миллиметров (5,7 дюйма). [1]
Этот образец использовался для диагностики Kinnareemimus в отличие от всех других орнитомимозавров на основании следующих апоморфий : полуарктометатарзальное состояние, при котором третья плюсневая кость все еще видна в проксимальном виде и сужается в дистальном направлении, а затем расширяется до более широкого треугольного поперечного сечения. на самом дистальном конце. [1] Самати изменил этот диагноз, В 2024 году включив в него уплощенные вентрально когти стоп со сгибательными ямками, что подтверждает включение Kinnareemimus в состав орнитомимозаврии. [7]
Рекомендуемые образцы
[ редактировать ]- Осевой скелет
Единственные сохранившиеся части осевого скелета Kinnareimimus — это восемь позвонков из различных отделов позвоночника (один предполагаемый спинной позвонок и семь хвостовых позвонков ). Единственный некаудальный позвонок (PW5A-124) был предположен как спинной позвонок, поскольку у него отсутствуют суставные грани для шевронов , хотя это точно не известно. Нервная дуга позвонка не сохранилась, а отсутствующая часть кости соответствует расположению несросшихся швов у неполовозрелой особи, что позволило авторам предположить, что образец принадлежал молодой особи. [1]
Хвостовые позвонки, отнесенные к Kinnareemimus, принадлежат нескольким разным особям. Центры у имеют форму песочных часов и полые, без поперечных отростков, что напоминает состояние, наблюдаемое Gallimimus . Некоторые из сохранившихся позвонков имеют разную форму поперечного сечения и, вероятно, происходят из разных отделов хвоста. По крайней мере один (PW5A-130) может быть первым хвостовым позвонком. Семь позвонков, сохранившихся от других частей хвоста, имеют различную степень сращения, что указывает на то, что эти экземпляры относятся к разновозрастному комплексу. [1]
- Аппендикулярный скелет
Большинство останков, обнаруженных на Фу Вианг 5, представляют собой кости задних конечностей или бедер различных людей. две частичные лобковые кости Сохранились , которые очень тонкие по сравнению с лобковыми костями такого же размера у орнитомимозавров такого же размера . индивидуума Баффето и его коллеги отмечают, что это может быть не настоящая апоморфия, а скорее артефакт онтогенеза . [1]
две голени В типовом местонахождении сохранились . Они одинаковы по длине: один из правой ноги, другой из левой ноги. Однако считается, что они принадлежат разным особям из-за различий в устойчивости между ними, которые, возможно, являются результатом онтогенеза. Общая форма голеней напоминает примитивных орнитомимозавров с небольшими отличительными чертами. Единственная малоберцовая кость сохранившаяся лишь частично цела и сохраняет широкую и глубокую бороздку вдоль стержня, напоминающую состояние Галлимима . Сохранившиеся фаланги по форме схожи с Гарудимимусом и Гарпимимусом , но они более тонкие. В единственном сохранившемся частичном когте пальца нет бугорка-сгибателя, аналогичного третьему когтю у Struthiomimus . [1]
Большинство останков Kinnareemimus представляют собой плюсневые кости, которые происходят из нижней части ноги чуть выше стопы. Известны плюсневые кости нескольких человек, что позволяет реконструировать плюсну с достаточной точностью. Примечательной особенностью является наличие аркто-метатарзального состояния , сходного с таковым у производных орнитомимид и тираннозавроидов . Однако третья плюсневая кость не была полностью окружена проксимальными концами второй и четвертой плюсневых костей, поэтому это не была настоящая арктометатарзальная кость. [1]
Классификация
[ редактировать ]Хотя Kinnareemimus является относительно фрагментарным, имеющиеся данные реконструкций плюсневых костей позволяют предположить его относительное эволюционное положение в составе Ornithomimosauria . Развитие арктометатарзуса примерно линейно в рамках филогенетической топологии орнитомимозавров. Из этого был сделан вывод, что Kinnareemimus находится в более производном положении, чем Garudimimus , но менее производном, чем Ornithomimidae . Однако Kinnareimimus геологически намного старше Garudimimus , а также Pelecanimimus и Harpymimus (также базальных орнитомимозавров), что позволяет предположить, что эволюция арктометатарзуса у орнитомимозавров не была хронологически линейной. Баффето и его коллеги используют это, чтобы предположить, что большая часть эволюции орнитомимозавров еще не исследована. [1] Они не проводят филогенетический анализ, включая Kinnareemimus , в свое описание [1] но последующие авторы пытались это сделать. [8] [9]
В своей филогенетической работе над тероподами в 2014 году . Стив Брусатте и его коллеги составили филогенетическую матрицу, используя 152 таксона, закодированных по 853 дискретным признакам Эта матрица включала данные по Kinnareemimus , но таксон пришлось удалить из окончательных деревьев, созданных для статьи. Консенсусное дерево максимальной экономии, полученное в дополнительных материалах к этой статье, включало Kinnareemimus , и это дерево не смогло выявить большинство манирапторанов клад как монофилетические . [8] Последующие анализы с использованием этой матрицы и других матриц сократили число Kinnareimimus по тем же причинам. [10] [11]
Первая научная статья, в которой Киннаримимус был включен в опубликованные деревья со значительным разрешением, была в 2024 году написана Андреа Кау . Дополнительные материалы из этой статьи включали десятки консенсусных деревьев, некоторые из которых включали Kinnareimimus . Сокращенная версия деревьев, опубликованная Кау, показана ниже. [9]



Вскоре после публикации анализа теропод Кау в 2024 году Адун Самати опубликовал статью, в которой анализировалось филогенетическое положение Kinnareimimus . было очень мало поддержки Kinnareimimus как орнитомимозавра В своем анализе он отмечает, что в основных филогенетических анализах . Брусатте и его коллеги (2014) отметили, что это, вероятно, был базальный целурозавр . Хартман и его коллеги (2019) нашли его тираннозавроидом , но также подтвердили, что он может представлять собой альваресзавроида . Чтобы устранить неопределенность в литературе до этого момента, Самати сгруппировала анатомические данные Kinnareemimus в трех независимых матрицах теропод. Это были матрицы Шуаньера и его коллег (2012 г.), Макфитерса и его коллег (2016 г.) и Хаттори и его коллег (2023 г.). [7]
Три матрицы дали разные результаты. Матрица Шуаньера и его коллег, которая также была модифицирована Самати для включения Deinocheirus , выявила Kinnareemimus как базального члена Deinocheiridae . Матрица Макфитерса и его коллег определила его как немакрохейриформного орнитомимозавра , и этот результат также был получен с помощью матрицы Хаттори и его коллег. Самати в конечном итоге не смог прийти к уверенной позиции в отношении этого таксона, кроме того, что он определенно не является членом Ornithomimidae . Окончательное дерево, опубликованное в работе Самати на основе матрицы Хаттори, показано ниже. [7]

Палеобиология
[ редактировать ]
В своем описании киннаримима Бюффето и его коллеги попытались оценить возможные беглые адаптации животного в жизни. Сохранившиеся кости позволили авторам реконструировать большую часть задней конечности на основе оценки размера костей взрослого человека, которые, как предполагалось, принадлежали молодым особям. Соотношение длин голени и плюсны было ранее установлено как метод определения беглости у других орнитомимозавров . Предполагаемая длина предплюсны у Киннаримима была сопоставима с известными останками Гарудимима и была значительно меньше, чем соотношения, известные у Струтиомима и Галлимима , которые, как полагают, были хорошо приспособлены для бега. [1]
Однако авторы также отметили, что плюсневые кости Kinnareemimus были гораздо более тонкими, чем у близкородственного Archaeornithomimus , что может свидетельствовать о том, что либо животное не было полностью взрослым, либо оно могло приобрести педоморфные черты. Они не выдвигают каких-либо конкретных гипотез относительно беговых способностей Киннаримимуса . [1]
Палеоэкология
[ редактировать ]Палеосреда
[ редактировать ]В формации Сан-Кхуа до сих пор были обнаружены только континентальные окаменелости, без каких-либо свидетельств морских окаменелостей или морских отложений . [12] Исторически были сообщения о зубах ихтиозавров и плезиозавров из формации Сан Хуа, но теперь они идентифицированы как принадлежащие крокодилиформным в основном не были морскими , которые в меловой период . [13] [14] Отложения формации Сан Хуа состоят из красных глин , аргиллитов , песчаников , алевролитов и конгломератных пород, что указывает на речную среду , в которой также были озера, поймы и разветвленные каналы . [12] [15]
Современная фауна
[ редактировать ]
Из всех мезозойских образований на северо-востоке Таиланда Сан-Кхуа имеет наиболее обильную и разнообразную окаменелостей позвоночных . летопись [16] В частности, известно, что в местонахождении Фувианг 5 обитали небольшие, возможно, молодые особи зауропода Phuwiangosaurus . Спинозаврид . сиамозавр также известен из этой местности по зубам [1]
В этом регионе обитало множество других теропод, хотя достоверно неизвестно, сосуществовали ли какие-либо из них конкретно с Киннаримимусом . В их число входили симпатрические мегарапторы Phuwiangvenator и Vayuraptor , [17] большой теропод Siamotyrannus , безымянный спинозаврид (иногда называемый «спинозаврид B Фу Вианг») и безымянный кархародонтозаврид . [13] [18] В этом районе также обитали другие мелкие тероподы, в том числе безымянный орнитомимозавр , безымянный компсогнатид и, возможно, орнитурины . [19] [20] В этом районе также известны останки зауроподов , которые, как полагают, отличаются от Phuwiangosaurus , но ни один из них не был официально описан или назван. Однако каких-либо птицетазовых животных . в этой формации до сих пор не обнаружено [13]
Пресноводная жизнь в формации Сан Хуа изобилует. К ним относятся неозухи Siamosurus , Sunosurus и Theriosuchus , многочисленные виды черепах , амиообразные рыбы, несколько гибодонтиформных акул и пресноводные двустворчатые моллюски . [15] Из этой формации также известен неопределенный птерозавр . [21]
См. также
[ редактировать ]- 2009 г. по палеонтологии архозавров.
- Аптское вымирание
- Группа Корат
- Список азиатских динозавров
- Палеонтология в Таиланде
- Хронология исследований орнитомимозавров
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п Баффето, Эрик; Сутитхорн, Варавуд; Тонг, Хайян (2009). «Ранний страусиный динозавр» (Theropoda: Ornithomimosauria) из раннемеловой формации Сан Хуа на северо-востоке Таиланда». В Бюффето, Эрик; Куни, Г.; Ле Лефф, Жан; Сутитхорн, Варавуд (ред.). Позднепалеозойские и мезозойские экосистемы Юго-Восточной Азии . Геологическое общество Лондона. стр. 229–243. дои : 10.1144/SP315.16 . ISBN 978-1-86239-275-5 . S2CID 128633687 .
- ^ Баффето, Эрик; Сутитхорн, Варавуд; Тонг, Хайян; Чаймани, Яовалак; Хансубха, Сасидхорн (1997). «Новые открытия динозавров в юрском и меловом периоде северо-восточного Таиланда» (PDF) . Международная конференция по стратиграфии, тектонической эволюции Юго-Восточной Азии и южной части Тихого океана : 157–161.
- ^ Jump up to: а б Канеко, Рюичи (2000). «По следам динозавров в Таиланде». Дино Пресс Том. 1 . Япония: Aurora Oval Inc.
- ^ «Журнал Дино Пресс» . Архив списка рассылки ДИНОЗАВРОВ . 2000.
«По следам динозавров в Таиланде»: статья Рюичи Канеко об ихнологии динозавров с некоторым обсуждением фувиангозавра и сиамотиранна.
- ^ «Исправления списка родов динозавров № 150» . Архив списка рассылки ДИНОЗАВРОВ . 2000.
В статье Рюичи Канеко в первом выпуске Dino Press приводится название еще неописанного орнитомимида из Таиланда, Ginnareemimus (название вида не указано).
- ^ Молина-Перес, Рубен; Ларраменди, Азиер (2019). Факты и цифры о динозаврах: тероподы и другие динозаврообразные Перевод Коннолли, Дэвид; Рамирес Кросс, Гонсало Анхель. Иллюстрировано Андреем Атучиным и Санте Маццеи. Издательство Принстонского университета. ISBN 978-0691180311 .
- ^ Jump up to: а б с Самати, Адун (2024). «Филогенетическое положение Kinnareimimus khonkaenensis (Dinosauria: Theropoda: Ornithomimosauria) из нижнего мела Таиланда» Зоотаксы 5448 : 67–84. дои : 10.11646/zootaxa.5448.1.4 .
- ^ Jump up to: а б Брусатте, Стивен Л.; Ллойд, Грэм Т.; Ван, Стив С.; Норелл, Марк А. (2014). «Постепенная сборка плана тела птиц привела к быстрым темпам эволюции при переходе от динозавра к птице» (PDF) . Современная биология . 24 (20): 2386–2392. дои : 10.1016/j.cub.2014.08.034 . ПМИД 25264248 . S2CID 8879023 .
- ^ Jump up to: а б Кау А. (2024). Единая основа макроэволюции хищных динозавров . Бюллетень Итальянского палеонтологического общества, 63(1): 1-19.
- ^ Ли Сюй; Ёсицугу Кобаяши; Цзюньчан Лю; Юнг-Нам Ли; Юнцин Лю; Кохей Танака; Синляо Чжан; Сонхай Цзя; Цзиминь Чжан (2011). «Новый динозавр-орнитомимид, имеющий североамериканское происхождение, из позднемеловой формации Цюпа в провинции Хэнань, Китай». Меловые исследования . 32 (2): 213–222. дои : 10.1016/j.cretres.2010.12.004 .
- ^ Серрано-Браньяс, Клаудия Инес; Эспиноса-Чавес, Белинда; Маккракен, С. Огаста; Гутьеррес-Бландо, Сирена; Де Леон-Давила, Клаудио; Вентура, Хосе Флорес (2020). « Paraxenisaurusnormalensis , крупный дейнохейрид-орнитомимозавр из формации Серро-дель-Пуэбло (верхний мел), Коауила, Мексика». Журнал южноамериканских наук о Земле . 101 . Бибкод : 2020JSAES.10102610S . дои : 10.1016/j.jsames.2020.102610 . S2CID 218968100 .
- ^ Jump up to: а б Такер, Райан Т.; Хайланд, Итан Г.; Гейтс, Терри А.; Кинг, М. Райан; Робертс, Эрик М.; Фоли, Эллиот К.; Берндт, Дэвид; Ханта, Раттанафорн; Хансубха, Саса-он; Асвасерилерт, Васини; Занно, Линдси Э. (2022). «Возраст, история отложений и палеоклиматическая обстановка скоплений динозавров раннего мела из формации Сан Хуа (группа Корат), Таиланд» . Палеогеография, Палеоклиматология, Палеоэкология . 601 : 111107. doi : 10.1016/j.palaeo.2022.111107 . S2CID 249702713 .
- ^ Jump up to: а б с Баффето, Эрик; Сутитхорн, Варавуд (1999). «Фауна динозавров формации Сан-Кхуа в Таиланде и начало меловой радиации динозавров в Азии» . Палеогеография, Палеоклиматология, Палеоэкология . 150 (1–2): 13–23. Бибкод : 1999PPP...150...13B . дои : 10.1016/S0031-0182(99)00004-8 . ISSN 0031-0182 .
- ^ Баффето, Эрик; Жиль, Кюни; Ле Лефф, Жан; Сутитхорн, Варавуд (2009). Позднепалеозойские и мезозойские континентальные экосистемы Юго-Восточной Азии . Лондон: Лондонское геологическое общество . стр. 2–3. дои : 10.1144/SP315 . ISBN 978-1862395633 .
- ^ Jump up to: а б Эрик, Бюффето; Варавуд, Сутитхорн (1998). Холл, Роберт; Холлоуэй, Джереми Д. (ред.). «Биогеографическое значение мезозойских позвоночных Таиланда» . Биогеография и геологическая эволюция Юго-Восточной Азии . Лейден, Нидерланды: Издательство Backhuys: 83–90.
- ^ Самати, Адун; Сандер, П. Мартин; Чантасит, Форнфен (2021). «Спинозаврид из Таиланда (формация Сан-Хуа, ранний мел) и повторная оценка Camarillasaurus cirugedae из раннего мела Испании» . Историческая биология . 33 (12): 3480–3494. дои : 10.1080/08912963.2021.1874372 . S2CID 233884025 .
- ^ Самати, А.; Чантасит, П.; Мартин Сандер, П. (май 2019 г.). «Два новых базальных целурозавровых теропода из формации Сан Хуа нижнего мела в Таиланде» . Acta Palaeontologica Polonica . 64 (2): 239–260. дои : 10.4202/app.00540.2018 .
- ^ Баффето, Эрик; Сутитхорн, Варавуд (январь 2012 г.). «Теропод кархародонтозаврид (Dinosauria, Saurischia) в формации Сан Хуа (ранний мел, баррем) Таиланда» . Тезисы докладов EAVP, 2012 г .: 27–30.
- ^ Баффето, Эрик; Сутитхорн, Варавуд; Ле Лефф, Жан; Хансубха, Саса-Он; Тонг, Хайян; Вонгко, К. (январь 2005 г.). «Фауна динозавров из формации Хок Круат (ранний мел) Таиланда» . Международная конференция по геологии, геотехнологии и минеральным ресурсам Индокитая (GEOINDO 2005) : 575–581.
- ^ Баффето, Эрик; Дайк, Гарет; Сутитхорн, Варавуд; Тонг, Хайян (1 декабря 2005 г.). «Первая запись ископаемой птицы из раннего мела Таиланда» . Comptes Рендус Палевол . 4 (8): 681–686. дои : 10.1016/j.crpv.2005.06.002 . ISSN 1631-0683 .
- ^ Кюни, Жиль; Сутитхорн, В. (1 января 2003 г.). «Зуб птеродактилоида из формации Сан Хуа (ранний мел) Таиланда» . Журнал Университета Махасаракхама . 22 : 92–98.
Внешние ссылки
[ редактировать ]- Краткое описание. Архивировано 17 июля 2011 г. на Wayback Machine в списке рассылки динозавров.














