Галлимим
| Галлимим Временной диапазон: поздний мел ( маастрихт ), | |
|---|---|
 | |
| Реконструированный скелет (на основе голотипа взрослой особи и молодого экземпляра), Музей естественной истории . | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Клэйд : | † Орнитомимозаврия |
| Семья: | † Орнитомимиды |
| Род: | † Галлимим Осмольска и др. , 1972 г. |
| Разновидность: | † Г. буллатус |
| Биномиальное имя | |
| † Галлимимус буллатос Осмольска и др. , 1972 г. | |
| Синонимы | |
| |
Галлимимус ( / ˌ ɡ æ l ɪ ˈ m aɪ m ə s / GAL -im - EYE -məs ) — род динозавров теропод - , живших на территории современной Монголии в позднемеловой период , около семидесяти миллионов лет назад (млн лет назад). . Несколько окаменелостей на разных стадиях роста были обнаружены польско-монгольскими экспедициями в пустыне Гоби в Монголии в 1960-х годах; большой скелет, обнаруженный в этом регионе, стал голотипом нового рода и вида Gallimimus Bullatus в 1972 году. Родовое название означает « имитатор курицы », что указывает на сходство между его шейными позвонками и позвонками Galliformes . Видовое название происходит от слова «булла» , золотой капсулы, которую носил римский юноша, в связи с выпуклой структурой у основания черепа Галлимима . На момент своего названия окаменелости Gallimimus представляли собой наиболее полный и наиболее хорошо сохранившийся материал орнитомимидов (« страусиных динозавров»), который когда-либо был обнаружен, и этот род остается одним из самых известных представителей группы.
Галлимимус — крупнейший известный орнитомимид ; взрослые особи имели длину около 6 метров (20 футов), высоту в бедрах 1,9 метра (6 футов 3 дюйма) и весили около 400–490 кг (880–1080 фунтов). Как свидетельствует его родственник Орнитомим , у него были перья . Голова была маленькая и легкая, с большими глазами, обращенными в стороны. Морда была длинной по сравнению с другими орнитомимидами, хотя на кончике она была шире и более закругленной, чем у других видов. Галлимим был беззубым, с ороговевшим (роговым) клювом и нежной нижней челюстью. Многие позвонки имели отверстия, указывающие на то, что они были пневматическими (наполненными воздухом). Шея была пропорционально длинной по отношению к туловищу. Руки были пропорционально самыми короткими среди всех орнитомимозавров, и на каждой было по три пальца с изогнутыми когтями. Передние конечности были слабыми, а задние - пропорционально длинными. Семейство Ornithomimidae входит в группу Ornithomimosauria . Ансеримимус , также родом из Монголии, считается ближайшим родственником Галлимим .
Как орнитомимид, Галлимимус был быстрым (или беглым ) животным, использующим свою скорость, чтобы избежать хищников; его скорость оценивается в 42–56 км/ч (29–34 миль в час). Возможно, у него было хорошее зрение и интеллект, сравнимый с бескилевыми птицами. Галлимимы , возможно, жили группами, судя по открытию нескольких экземпляров, сохранившихся в костном слое . Были предложены различные теории относительно диеты галлимима и других орнитомимид. Очень подвижная шея, возможно, помогала находить мелкую добычу на земле, но она также могла быть всеядным животным . Также было высказано предположение, что он использовал небольшие столбчатые структуры в своем клюве для фильтрации воды, хотя вместо этого эти структуры могли представлять собой гребни, используемые для питания жестким растительным материалом, что указывает на травоядную диету. Галлимимус — наиболее часто встречающийся орнитомимозавр в формации Немегт , где он жил вместе со своими родственниками Ансеримимусом и Дейнохейрусом . Галлимимус был показан в фильме « Парк Юрского периода» . , в сцене, которая сыграла важную роль в истории спецэффектов , а также в формировании общего представления о динозаврах как о птицеподобных животных.
История открытия
[ редактировать ]
В 1963–1965 годах Польская академия наук и Монгольская академия наук организовали польско-монгольские палеонтологические экспедиции в пустыню Гоби в Монголии. Среди останков динозавров, обнаруженных в песчаных пластах бассейна Немегта, были многочисленные орнитомимиды на разных стадиях роста из местонахождений Немегт, Цааган Хушуу, Алтан Ула IV и Наран Булак. Были собраны три частично целых скелета, два с черепами, а также множество фрагментарных останков. Самый крупный скелет (позже ставший голотипом Gallimimus Bullatus ) был обнаружен палеонтологом Зофией Килан-Яворовской в Цааган Хушуу в 1964 году; он сохранился лежащим на спине, а под его тазом был найден череп. В том же году один небольшой экземпляр был найден в Цааган-Хушуу, а еще один небольшой экземпляр — в местонахождении Немегт. Небольшой скелет без передних конечностей был обнаружен в 1967 году монгольской палеонтологической экспедицией в Буген-Цаве за пределами бассейна Немегта. Окаменелости находились в Монголии, Польше и Академия наук СССР . [1] [2] [3] Польско-монгольские экспедиции примечательны тем, что их возглавляли женщины, некоторые из которых были одними из первых женщин, давших имена новым динозаврам. Окаменелости, обнаруженные в ходе этих экспедиций, проливают новый свет на обмен фауной между Азией и Северной Америкой в меловой период . [4] [5] [6] Некоторые из скелетов были выставлены в Варшаве в 1968 году в высоких полувертикальных позах, что было принято в то время, хотя сегодня предпочтение отдается более горизонтальным позам. [6]
В 1972 году палеонтологи Хальшка Осмолска , Ева Роневич и Ринхен Барсболд назвали новый род и вид Gallimimus Bullatus , используя самый большой собранный скелет, экземпляр IGM 100/11 (из Цаагана Хушуу, ранее называвшийся GINo.DPS 100/11 и MPD 100). /11), как голотип . Родовое название происходит от латинского Gallus , «курица», и греческого mimos , «имитирующий», в отношении передней части шейных позвонков, которые напоминали таковые у Galliformes . Видовое название происходит от латинского «булла» — золотой капсулы, которую римские юноши носили на шее, отсылая к выпуклой капсуле на парасфеноиде у основания черепа динозавра. Такая особенность в то время не была описана у других рептилий и считалась необычной. Голотип состоит из почти полного скелета с деформированной мордой, неполной нижней челюстью, позвоночным рядом, тазом, а также некоторыми отсутствующими костями рук и ног. [1] [7]

Остальные частично полные скелеты были молодыми; ZPAL MgD-I/1 (из Цааган Хушуу) имеет размозженный череп с отсутствующим кончиком, поврежденными позвонками, фрагментированными ребрами, грудным поясом и передними конечностями, неполную левую заднюю конечность, ZPAL MgD-I/94 (из местонахождения Немегт) у него отсутствуют череп, атлас, кончик хвоста, грудной пояс и передние конечности, а у самого маленького экземпляра, IGM 100/10 (из Бугин Цав), отсутствуют грудной пояс, передние конечности и несколько позвонков и ребер. Осмольска и ее коллеги перечислили всего двадцать пять известных экземпляров, девять из которых были представлены только отдельными костями. [1] [8]
На момент своего названия окаменелости Gallimimus представляли собой наиболее полный и наиболее хорошо сохранившийся материал орнитомимид, когда-либо обнаруженный, и этот род остается одним из самых известных представителей группы. Орнитомимиды ранее были известны в основном из Северной Америки, причем Archaeornithomimus был единственным известным ранее представителем из Азии (хотя и без черепа). С момента первых открытий в ходе дальнейших международных экспедиций под руководством Монголии было обнаружено больше экземпляров. [1] [8] [5] [6] Три скелета галлимима (включая голотип) позже стали частью передвижной выставки окаменелостей монгольских динозавров, которая путешествовала по разным странам. [9] [10]

Браконьерство в отношении ископаемых стало серьезной проблемой в Монголии в 21 веке, и несколько экземпляров Gallimimus были разграблены. В 2017 году Ханг-Джэ Ли и его коллеги сообщили об обнаруженных в 2009 году ископаемых следах, связанных со сжатой ступней галлимима (образец MPC-D100F/17). Остальная часть скелета, по-видимому, была ранее удалена браконьерами вместе с несколькими другими экземплярами Gallimimus (на что указывают пустые ямы для раскопок, мусор и разбросанные сломанные кости в карьере). Необычно находить следы, тесно связанные с окаменелостями тела; некоторые следы соответствуют стопам орнитомимид, а другие принадлежат другим динозаврам. [11] [12] В 2014 году плита с двумя экземплярами галлимима была репатриирована в Монголию вместе с другими скелетами динозавров после контрабанды в США. [13]
В 1988 году палеонтолог Грегори С. Пол пришел к выводу, что черепа орнитомимид более похожи друг на друга, чем считалось ранее, и переместил большинство видов в один и тот же род Ornithomimus , в результате чего возникла новая комбинация O. Bullatus . [14] В 2010 году он вместо этого перечислил его как « Galimimus (или Struthiomimus ) Bullatus », но в 2016 году вернулся к использованию только названия рода Gallimimus . [15] [16] Другие авторы обычно выделяли рассматриваемые виды в отдельные роды. [17] Об орнитомимидном позвонке из Японии, неофициально названном «Санхузавр», сообщалось в журнале 1988 года, но он был отнесен к Gallimimus sp. (неопределенного вида) палеонтологом Дун Чжимином и его коллегами в 1990 году. [7] В 2000 году палеонтолог Филип Дж. Карри предположил, что , но это было Anserimimus, который известен только по одному скелету из Монголии, был младшим синонимом Gallimimus отвергнуто Кобаяши и Барсболдом, которые указали на несколько различий между ними. Барсболд отметил некоторые морфологические различия среди новых экземпляров Gallimimus , хотя это никогда не публиковалось. Барсболд неофициально назвал почти полный скелет (IGM 100/14) «Gallimimus mongoliensis», но, поскольку он отличается от Gallimimus в некоторых деталях, Ёсицугу Кобаяши и Барсболд в 2006 году предположили, что он, вероятно, принадлежит к другому роду. [8]
Описание
[ редактировать ]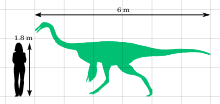
Gallimimus — самый крупный известный представитель семейства Ornithomimidae . Взрослый голотип (образец IGM 100/11) имел длину около 6 метров (20 футов) и высоту в бедрах 1,9 метра (6,2 фута); его череп имел длину 330 миллиметров (1,08 фута), а длина бедренной кости (бедренная кость) - 660 миллиметров (2,17 фута). Он весил бы около 400–490 кг (880–1080 фунтов). [14] [15] [18] [19] Для сравнения, один молодой экземпляр (ZPAL MgD-I/94) имел длину около 2,15 метра (7,1 фута), высоту в бедрах 0,79 метра (2,6 фута) и весил около 26–30,2 кг (57–67 фунтов). [14] [18] На основе окаменелостей родственных им орнитомимов известно, что орнитомимозавры (« страусиные динозавры») были пернатыми , а взрослые особи имели крыловидные структуры, о чем свидетельствует наличие перьевых выступов на . локтевой кости предплечья, шишек которые указывают, где могли бы прикрепиться перья. [20]
Череп
[ редактировать ]
Голова Галлимима была очень маленькой и легкой по сравнению с позвоночником. Из-за длины морды череп был длинным по сравнению с другими орнитомимидами, а верхний профиль морды имел пологий выпуклый наклон. Боковой профиль рыла отличался от других орнитомимид тем, что не сужался к передней половине, а нижний передний край предчелюстной кости в передней части верхней челюсти приподнимался вверх, а не был горизонтальным. При взгляде сверху морда была почти лопатообразной (ложкообразной), широкой и закругленной на кончике (или U-образной ), тогда как у североамериканских орнитомимид она была острой (или V-образной). Орбиты . (глазницы) были большими и обращены вбок, как и у других орнитомимид Височная область сбоку черепа за глазами была глубокой, а подвисочное окно (нижнее отверстие за глазницей) было почти треугольным и меньшим, чем у родственного Struthiomimus . У него были глубокие мышечные рубцы в задней части крыши черепа, вдоль теменной кости. . Парасфеноид (кость черепной коробки в нижней части основания черепа) был тонкостенным, полым и имел грушевидную выпуклую структуру. В конструкции была неглубокая борозда, открывавшаяся вперед. ( Внутренние ноздри внутренние отверстия для носового хода) были большими и располагались далеко назад на небе из-за наличия обширного вторичного неба , свойственного орнитомимидам. [1] [17] [21] [22]
Нежная нижняя челюсть, состоящая из тонких костей, была тонкой и неглубокой спереди, углубляющейся к задней части. Передняя часть нижней челюсти имела лопатообразную форму, в результате чего между кончиками челюстей в закрытом состоянии образовывался зазор. Лопаткообразная форма была похожа на форму обыкновенной чайки , а нижняя часть клюва, возможно, имела форму, подобную форме этой птицы. Засуставной отросток в задней части челюсти (место прикрепления челюстных мышц, открывающих клюв) был хорошо развит и состоял преимущественно из угловой кости . кость Надугольная представляла собой самую крупную кость нижней челюсти, обычную для теропод. Нижнечелюстное окно , обращенное в сторону отверстие в нижней челюсти, было удлиненным и сравнительно небольшим. Нижняя челюсть не имела венечного отростка и надзубной кости, отсутствие которых является общей чертой клюворылых теропод (орнитомимозавров, овирапторозавров , теризинозавров и птиц), но необычной среди теропод в целом. [1] [23] Челюсти Галлимима были беззубыми (беззубыми), а передняя часть при жизни была бы покрыта ороговевшей рамфотекой (роговым клювом). Клюв, возможно, занимал меньшую площадь, чем у североамериканских родственников, из-за отсутствия питательных отверстий на верхней челюсти. Внутренняя сторона клюва имела небольшие, плотно упакованные и равномерно расположенные столбчатые структуры (их точная природа дискуссионна), которые были наиболее длинными спереди и укорачивались к задней части. [21] [23] [24]
Посткраниальный скелет
[ редактировать ]
галлимима было 64–66 позвонков В позвоночнике , меньше, чем у других орнитомимид. Центры (или тела) позвонков были платицелозными, с плоской передней поверхностью и вогнутой задней поверхностью, за исключением первых шести хвостовых (хвостовых) позвонков, у которых задняя поверхность также была плоской, и позвонков на конце хвоста. — которые были амфиплатианскими с плоскими обеими поверхностями. Многие центры имели отверстия (отверстия, которые также называли « плевроцелями ») и поэтому, вероятно, были пневматическими (их полые камеры заполнены воздушными мешками ). Шея состояла из 10 шейных позвонков , все они были длинными и широкими, за исключением атланта (первого позвонка, соединяющегося с задней частью черепа). Атлант отличался от такового у других орнитомимид тем, что передняя поверхность его межцентра была наклонена вниз к спине, а не была вогнутой и обращена вверх для поддержки затылочного мыщелка . Шея, по-видимому, была пропорционально длиннее туловища, чем у других орнитомимид. Шея была разделена на две отдельные части: шейные позвонки спереди имели центры, которые при виде сбоку были почти треугольными и сужались к спине, а также низкие невральные дуги и короткие, широкие зигапофизы ( отростки , сочленяющиеся между позвонками); шейные позвонки сзади имели бочкообразные центры, постепенно становившиеся выше, и длинные тонкие зигапофизы. Пневматические отверстия здесь были маленькими и овальными, а нервные отростки, выступающие наружу из центра, образовывали длинные, низкие и острые гребни, за исключением самых задних шейных позвонков. [1] [25]

На спине Галлимима было 13 спинных позвонков с бочкообразными центрами, которые были короткими, но имели тенденцию становиться глубже и длиннее к спине. Их поперечные отростки (отростки, сочленяющиеся с ребрами) немного увеличивались в длине к спине. Два первых спинных центра имели глубокие пневматические отверстия, тогда как остальные имели только неглубокие ямки (впадины), а нервные отростки были выступающими и имели несколько треугольную или прямоугольную форму. Крестец крестцовых (сросшиеся позвонки между костями таза) состоял из пяти позвонков примерно одинаковой длины. Центры здесь имели катушкообразную форму, уплощены по бокам и имели ямки, которые, по-видимому, у некоторых экземпляров продолжали оставаться глубокими отверстиями. Нервные отростки здесь были прямоугольными, широкими и выше, чем у спинных позвонков. Они были выше или равны по высоте верхнему краю лопатки подвздошной кости и были раздельными, тогда как у других орнитомимид они срослись между собой. В хвосте было 36–39 хвостовых позвонков, центральная часть передних позвонков имела форму катушки, а задняя часть была почти треугольной и удлиненной в поперечнике. Нервные отростки здесь были высокими и плоскими, но уменьшались назад и становились гребневидными. Единственным признаком пневматичности хвоста были глубокие ямки между нервными отростками и поперечным отростком двух первых хвостовых позвонков. Все позвонки перед крестцом несут ребер, за исключением атланта и последнего спинного позвонка. [1] [25]
Лопатка (лопатка) была короткой и изогнутой , тонкой на переднем конце и толстой сзади. Он сравнительно слабо был связан с клювовидным отростком , который был большим и глубоким сверху донизу. В целом передние конечности мало чем отличались от конечностей других орнитомимид, все они были сравнительно слабыми. ( Плечевая кость кость плеча), имевшая почти круглое поперечное сечение, была длинной и искривленной. Дельтопекторальный гребень в верхней передней части плечевой кости был сравнительно небольшим и поэтому обеспечивал мало места для прикрепления мышц плеча. Локтевая кость была тонкой, длинной и слабо изогнутой, с почти треугольным стержнем. Олекранон ( выступ локтя) был заметным у взрослых, но недостаточно развит у молодых. ( Лучевая кость другая кость предплечья) была длинной и тонкой, с более расширенным верхним концом по сравнению с нижним. Рука (кисть) была пропорционально короткой по сравнению с таковыми у других орнитомимозавров, имея наименьшее соотношение длины кисти и плечевой кости среди всех членов группы, но в остальном была схожа по структуре. У него было три пальца одинакового развития; первый («большой палец») был самым сильным, третий — самым слабым, а второй — самым длинным. когтевые кости (когтевые кости) были крепкими, несколько изогнутыми (наиболее изогнутым был указательный палец) и сжатыми вбок с глубокими бороздками на каждой стороне. Когтевые кости были развиты так же, хотя третий был немного меньше. [1] [8] [17]

( Лобковая кость лобковая кость) была длинной и тонкой и заканчивалась лобковым ботинком, расширявшимся вперед и назад, что является общей чертой орнитомимозавров. Задние конечности мало отличались от таковых у других орнитомимид и были пропорционально длиннее, чем у других теропод. Бедренная кость была почти прямой, длинной и тонкой, со сплюснутым вбок стержнем. Голень мыщелками прямая, длинная, с двумя хорошо развитыми ( закругленным концом кости) на верхнем конце и уплощенным нижним концом. голени Малоберцовая кость плоская, тонкая и широкая на верхнем конце, сужающаяся к нижнему концу. Нижняя половина третьей плюсневой кости была широкой, если смотреть с конца, частично закрывая две прилегающие плюсневые кости с каждой стороны, но резко сужалась на середине длины, вклинивалась между этими костями и исчезала ( арктометатарзальная структура стопы). Третий палец ноги был пропорционально короче по отношению к конечности, чем у других орнитомимид. Как и у других орнитомимид, на ступне не было большого пальца стопы (или прибылого пальца, первого пальца у большинства других теропод). Когти пальцев ног были плоскими с нижней стороны; два внешних пальца слегка отклонились наружу от своих пальцев. [1] [17]
Классификация
[ редактировать ]Осмолска и ее коллеги отнесли Gallimimus к семейству Ornithomimidae в 1972 году, причем североамериканский Struthiomimus был ближайшим родственником, при этом сетуя на тот факт, что сравнение между таксонами было затруднено, поскольку другие орнитомимиды, известные в то время, либо плохо сохранились, либо неадекватно описаны. [1] В 1975 году Килан-Яворовска заявила, что, хотя многие динозавры из Азии были отнесены к тем же семействам, что и их североамериканские родственники, эта категория классификации, как правило, была более всеобъемлющей, чем та, которая использовалась для современных птиц. Она подчеркнула, что в то время как у галлимима был закругленный клюв (похожий на гусиный или утиный), у североамериканских орнитомимид клюв был заостренным, что в противном случае заставило бы систематиков отнести современных птиц к разным семействам. [5] В 1976 году Барсболд поместил Ornithomimidae в новую группу Ornithomimosauria. В 2003 году Кобаяши и Джун-Чанг Лю обнаружили, что , образуя производную ( Anserimimus был сестринским таксоном Gallimimus или « продвинутую») кладу с североамериканскими родами, что было подтверждено Кобаяши и Барсболдом в 2006 году. [8] [21]
На следующей кладограмме показано место Gallimimus среди Ornithomimidae по мнению палеонтолога Брэдли Макфитерса и его коллег, 2016: [26]


| Орнитомимиды |
| ||||||||||||||||||||||||||||||||||||
Орнитомимозавры принадлежали к кладе Maniraptoriformes целурозавровых теропод , в которую входят и современные птицы. У ранних орнитомимозавров были зубы, которые были потеряны у более продвинутых представителей группы. [27] В 2004 году Маковицки, Кобаяши и Карри предположили, что большая часть ранней эволюционной истории орнитомимозавров произошла в Азии, где было обнаружено большинство родов, включая самые базальные (или «примитивные») таксоны, хотя они и признали, что присутствие базальный Pelecanimimus в Европе представляет собой сложность в классификации. Эта группа, должно быть, один или два раза рассеялась из Азии в Северную Америку через Берингию, чтобы объяснить обнаруженные там позднемеловые роды. Как видно из некоторых других групп динозавров, орнитомимозавры были в основном ограничены Азией и Северной Америкой после того, как Европа была отделена от Азии Тургайским проливом . [17]
В 1994 году палеонтолог Томас Р. Хольц сгруппировал орнитомимозавров и троодонтид в кладу на основе общих черт, таких как наличие выпуклой капсулы на парасфеноиде. Он назвал кладу Bullatosauria, основываясь на видовом названии Gallimimus Bullatus , которое также относилось к капсуле. [28] Вместо этого в 1998 году Хольц обнаружил, что троодонтиды были базальными манирапторами, а это означает, что все члены этой клады относятся к Bullatosauria, которая, следовательно, стала младшим синонимом Maniraptoriformes, и с тех пор эта клада вышла из употребления. [29] [30]
Палеобиология
[ редактировать ]
Шейные позвонки Галлимима указывают на то, что он держал шею наклонно, склоняясь вверх под углом 35 градусов. Осмольска и ее коллеги обнаружили, что руки Галлимима не были цепкими (или неспособными хватать), а большой палец не был противопоставляемым. Они также предположили, что руки были слабыми по сравнению, например, с таковыми у орнитомимозавра Deinocheirus . Они согласились с интерпретациями биологии орнитомимид, сделанными палеонтологом Дейлом Расселом ранее в 1972 году, в том числе с тем, что они были очень быстрыми (или беглыми ) животными, хотя и менее проворными, чем крупные современные наземные птицы, и использовали бы свою скорость, чтобы спастись от хищников. . Рассел также предположил, что у них было хорошее зрение и интеллект, сравнимый с интеллектом современных бескилевых птиц. Поскольку их хищники могли обладать цветовым зрением , он предположил, что это повлияло на их окраску, что, возможно, привело к маскировке . [1] [31] В 1982 году палеонтолог Ричард А. Талборн подсчитал, что Галлими мог бегать со скоростью 42–56 км/ч (29–34 миль в час). Он обнаружил, что орнитимимиды не были бы такими быстрыми, как страусы, которые могли достигать скорости 70–80 км/ч (43–49 миль в час), отчасти из-за того, что их руки и хвосты увеличивали их вес. [32]

В 1988 году Пол предположил, что глазные яблоки орнитомимид были уплощены и имели минимальную подвижность внутри глазниц, что требовало движения головы для просмотра объектов. Поскольку их глаза смотрели в сторону больше, чем у некоторых других птицеподобных теропод, их бинокулярное зрение было бы более ограниченным, что является адаптацией у некоторых животных, улучшающей их способность видеть хищников позади себя. Пол считал относительно короткие хвосты, которые уменьшали вес, и отсутствие пальцев орнитомимид адаптацией к скорости. Он предположил, что они могли бы защитить себя, клевая и пиная, но для побега в основном полагались бы на свою скорость. [14] что вместе с дейнохейрусом и археорнитомимом галлимимус В 2015 году Акинобу Ватанабэ и его коллеги обнаружили , имел наиболее пневматизированный скелет среди орнитомимозавров. Считается, что пневматизация полезна для полета современных птиц, но ее функция у нептичьих динозавров достоверно не известна. Было высказано предположение, что пневматизация использовалась для уменьшения массы крупных костей, что она была связана с высоким обменом веществ , балансом во время передвижения или использовалась для терморегуляции . [25]
В 2017 году Ли и его коллеги предположили различные возможные тафономические обстоятельства (изменения во время распада и окаменения), чтобы объяснить, как ступня Галлимима , обнаруженная в 2009 году, была связана со следами. Тропа сохранилась из песчаника, а подножие сохранилось из аргиллита , простираясь на 20 сантиметров (7,9 дюйма) ниже слоя с путями. Возможно, окаменелость представляет собой животное, которое умерло на своем пути, но глубина ступни в грязи может быть слишком небольшой, чтобы она могла завязнуть. Возможно, он также погиб в результате наводнения, после чего был похоронен в пруду. Однако слои грязи и песка указывают не на наводнение, а, вероятно, на засушливую среду, а разрушенные отложения вокруг окаменелости указывают на то, что животное было живо, когда оно прибыло в этот район. Таким образом, авторы предположили, что следы были оставлены в течение длительного периода времени и периода высыхания, и что, вероятно, ни один из них не был оставлен человеком, которому принадлежала ступня. Возможно, животное шло по дну пруда, пробивая следами слой наносов, пока оно было мокрым от дождя или содержало воду. Животное могло умереть в таком положении от жажды, голода или по другой причине, и грязь затем оседала на песке, тем самым покрывая и сохраняя следы и тушу. Нога могла сжаться и разъединиться по мере разложения, что сделало сухожилия сгибаются, и позже на него наступили тяжелые динозавры. Этот район мог представлять собой один слой костей (исходя из возможного количества браконьерских экземпляров), что свидетельствует о массовой смертности Gallimimus , возможно, из-за засухи или голода. Тот факт, что животные, по-видимому, погибли одновременно (пустые ямы были стратиграфически идентичными), может указывать на то, что Gallimimus был стадным человеком (жил группами), что предполагалось и для других орнитомимид. [12]
Кормление и диета
[ редактировать ]
Осмольска и ее коллеги отметили, что передняя часть шеи Gallimimus должна была быть очень подвижной (задняя часть была более жесткой), нервные дуги в позвонках этой области были похожи на куриные и другие Galliformes, что указывает на схожие привычки питания. Они обнаружили, что клюв Галлимима похож на клюв утки или гуся и что он питался маленькой живой добычей, которую проглатывал целиком. Подвижность шеи была бы полезна при обнаружении добычи на земле, поскольку глаза располагались по бокам черепа. Они предположили, что все орнитомимиды имели схожие пищевые привычки, и отметили, что Рассел сравнил клювы орнитомимид с клювами насекомоядных птиц. Осмольска и его коллеги предположили, что Галлимимус был способен к краниальному кинезису (из-за, казалось бы, слабого соединения между некоторыми костями задней части черепа), особенности, которая позволяет отдельным костям черепа двигаться относительно друг друга. Они также предположили, что он использовал свои короткие передние конечности не для того, чтобы подносить еду ко рту, а для того, чтобы разгребать или рыть землю в поисках еды. [1] руки Галлимима могли быть слабее, чем, например, у Струтиомима , который вместо этого мог использовать свои руки для крючков и захватов. Согласно статье палеонтологов Элизабет Л. Николлс и Энтони П. Рассела, опубликованной в 1985 году, [33]
В 1988 году Пол не согласился с тем, что орнитомимиды были всеядными и питались мелкими животными и яйцами, а также растениями, как предполагалось ранее. Он указал, что страусы и эму в основном травоядные и бродячие животные , и что черепа орнитомимид были наиболее похожи на черепа вымерших моа , которые были достаточно сильны, чтобы откусывать ветки, о чем свидетельствует содержимое их кишок. Он также предположил, что орнитомимиды были хорошо приспособлены к кормлению жестких растений и использовали свои руки, чтобы подносить ветки к своим челюстям. [14] В 2001 году палеонтолог Йорн Хурум предположил, что из-за схожего строения челюстей галлимимы , возможно, имели оппортунистическую и всеядную диету, как чайки. Он также заметил, что плотный внутричелюстной сустав предотвращает любое движение между передней и задней частями нижней челюсти. [23]

В 2001 году палеонтологи Марк А. Норелл , Маковицки и Карри сообщили о черепе галлимима (IGM 100/1133) и черепе орнитомима , в которых сохранились структуры мягких тканей на клюве. Внутренняя сторона клюва Gallimimus имела столбчатые структуры, которые, как обнаружили авторы, похожи на пластинки в клювах гусеобразных птиц, которые используют их для манипуляций с пищей, процеживания осадков, фильтрации путем отделения пищевых продуктов от другого материала, а также для резки растений. во время выпаса. Они обнаружили , что северная лопатка , питающаяся растениями, моллюсками, остракодами и фораминиферами , является современным гусеобразным видом со структурами, наиболее похожими по анатомии на структуры Gallimimus . Авторы отметили, что орнитомимиды, вероятно, не использовали свои клювы для охоты на крупных животных и были в изобилии в мезической среде, но реже в более засушливых средах, что позволяет предположить, что они, возможно, зависели от водных источников пищи. [24] Маковицки, Кобаяши и Карри отметили, что, если эта интерпретация верна, Галлимим был бы одним из крупнейших известных наземных фильтраторов. [17]
В 2005 году палеонтолог Пол Барретт отметил, что пластинчатые структуры Gallimimus, похоже, не представляли собой гибкие щетинки, как у птиц, питающихся фильтратором (поскольку нет никаких признаков перекрытия или разрушения этих структур), а вместо этого были более сродни тонким, равномерно расположенным вертикальным гребням на клювах черепах и динозавров -гадрозаврид . Считается, что у этих животных такие гребни связаны с травоядным рационом, используемым для выращивания жесткой растительности. Барретт предположил, что гребни на клюве Галлимима представляют собой естественный слепок внутренней поверхности клюва, что указывает на то, что животное было травоядным, питавшимся материалом с высоким содержанием клетчатки. Обнаружение множества гастролитов ( желудочных камней) у некоторых орнитомимид указывает на наличие желудочной мельницы и, следовательно, указывает на травоядный рацион, так как они используются для измельчения пищи животных, не имеющих необходимого жевательного аппарата. Барретт также подсчитал, что 440 кг (970 фунтов) Галлимиму потребовалось бы от 0,07 до 3,34 килограмма (от 0,15 до 7,36 фунта) пищи в день, в зависимости от того, имел ли он эндотермический или экзотермический («теплый» или «хладнокровный») метаболизм. невозможно, если бы это был фильтратор. Он также обнаружил, что орнитомимиды были в изобилии не только в формациях, представляющих мезическую среду, но и в засушливых средах, где было недостаточно воды для поддержания диеты, основанной на фильтрационном питании. [22] В 2007 году палеонтолог Эспен М. Кнутсен написал, что форма клюва орнитомимид по сравнению с формой клюва современных птиц соответствует всеядности или травоядности с высоким содержанием клетчатки. [34]
Дэвид Дж. Баттон и Занно обнаружили в 2019 году, что травоядные динозавры в основном придерживались двух различных способов питания: либо переработка пищи в кишечнике, характеризующаяся изящными черепами и низкой силой укуса, либо рот, характеризующийся особенностями, связанными с интенсивной переработкой. Было обнаружено , что орнитомимиды-орнитомимозавры, дейнохейрусы , диплодокоидные и титанозавровые зауроподы, сегнозавры и ценугнатиды относятся к первой категории. Эти исследователи предположили, что орнитомимиды-орнитомимозавры, такие как Gallimimus и дейнохейриды, вторглись в эти ниши по отдельности, конвергентно достигнув относительно больших размеров. Преимущества большой массы тела у травоядных включают повышенную скорость потребления пищи и устойчивость к голоданию, и, следовательно, эти тенденции могут указывать на то, что орнитомимиды и дейнохейриды были более травоядными, чем другие орнитомимозавры. Они предупредили, что корреляция между травоядностью и массой тела непроста и что у клада не наблюдается направленной тенденции к увеличению массы. Более того, рацион большинства орнитомимозавров малоизвестен. [35]
Разработка
[ редактировать ]
Форма и пропорции черепа существенно менялись в процессе роста. Задняя часть черепа и глазницы уменьшились в размерах, а морда стала относительно длиннее; аналогичные изменения происходят и у современных крокодилов . Череп также был пропорционально больше у более молодых экземпляров, а наклон верхнего профиля морды был менее выражен. Ребра на шее срослись с позвонками только у взрослых. Передние конечности, по-видимому, стали пропорционально длиннее в процессе роста, тогда как пропорциональная длина костей задних конечностей изменилась очень мало. [1] [23] В 2012 году палеонтолог Дарла К. Зеленицкая и ее коллеги пришли к выводу, что, поскольку у взрослых орнитомимозавров на руках были крылья, а у молодых - нет (о чем свидетельствуют образцы Ornithomimus ), эти структуры изначально были вторичными половыми признаками , которые можно было использовать для репродуктивного поведения, такого как ухаживание , демонстрация и вынашивание. [20]
Исследование 1987 года, проведенное биологами Романом Павлицким и П. Болечалой, показало возрастные различия в содержании кальция и фосфора (важных компонентов формирования костей) у Gallimimus особей . Они обнаружили, что это соотношение было самым высоким у животных молодого и среднего возраста и уменьшалось с возрастом. [36] В 1991 году они сообщили, что кости старых особей содержат наибольшее количество свинца и железа, а кости молодых животных — меньше. [37] Исследование гистологии костей различных динозавров, проведенное в 2000 году биологами Джоном М. Ренсбергером и Махито Ватабе, показало, что канальцы (каналы, соединяющие костные клетки) и пучки коллагеновых волокон галлимима и других орнитомимид больше похожи на таковые у птиц, чем у птиц. млекопитающих, в отличие от птицетазовых динозавров, которые были больше похожи на млекопитающих. Эти различия могли быть связаны с процессом и скоростью формирования кости. [38]
Палеосреда
[ редактировать ]
Gallimimus известен из формации Нэмегт в пустыне Гоби на юге Монголии. Это геологическое образование никогда не датировалось радиометрически , но фауна, присутствующая в летописи окаменелостей, указывает на то, что оно, вероятно, отложилось на раннем маастрихтском этапе, в конце позднего мела, около 70 миллионов лет назад. [39] [40] [41] Отложения Галлимимус местонахождения типа Цааган Хушуу состоят из алевритов , алевролитов , аргиллитов, песков, а также реже маломощных прослоев песчаников. [3] горных Фации пород формации Немегт предполагают наличие речных каналов, илистых отмелей, мелких озер и пойм в среде, аналогичной дельте Окаванго в современной Ботсване . [42] Крупные речные русла и почвенные отложения свидетельствуют о значительно более влажном климате, чем тот, который обнаружен в более старых формациях Барун-Гойот и Джадохта , хотя отложения калише указывают на то, что случались периодические засухи. [43] Ископаемые кости из бассейна Немегта, в том числе Галлимима , более радиоактивны , чем окаменелости из прилегающих территорий, возможно, потому, что в костях накапливался уран , переносимый туда просачивающимися грунтовыми водами. [6] [44]
Реки Немегта, где жил Галлимим , были домом для множества организмов. Случайные окаменелости моллюсков , а также множество других водных животных, таких как рыбы, черепахи, [39] и крокодиломорфы , в том числе Shamosuchus , были обнаружены в этом регионе. [45] Окаменелости млекопитающих редки в формации Немегт, но было обнаружено множество птиц, в том числе энантиорнитин Gurilynia , гесперорнитиформный Judinornis , а также Teviornis , возможный гусеобразный вид. Травоядные динозавры, обнаруженные в формации Немегт, включают анкилозаврид, таких как Tarchia , пахицефалозавров Prenocephale , крупных гадрозавридов, таких как Saurolophus и Barsboldia , и зауроподов, таких как Nemegtosaurus и Opisthocoelicaudia . [39] [46] Хищные тероподы, которые жили рядом с Галлимимом, включают тираннозавроидов , таких как Тарбозавр , Алиорам и Багараатан , и троодонтид, таких как Бороговия , Тохизавр и Занабазар . К травоядным или всеядным тероподам относятся теризинозавры, такие как Therizinosaurus , а также овирапторозавры, такие как Elmisaurus , Nemegtomaia и Rinchenia . [47] другие орнитомимозавры, в том числе Anserimimus и Deinocheirus Встречаются и Gallimimus . , но наиболее распространенным представителем группы в Немегте является [8]
Культурное значение
[ редактировать ]
Галлимимус был показан в фильме 1993 года « Парк Юрского периода» режиссера Стивена Спилберга ; в аналогичной сцене в оригинальном романе 1990 года вместо этого фигурировали гадрозавры. Спилберг хотел, чтобы в фильме была сцена давки со стадами животных, но не знал, как этого добиться, и изначально ее планировалось визуализировать с помощью покадровой анимации . В то время мало кто верил в создание животных с помощью компьютерной анимации , но по производству визуальных эффектов компания Industrial Light and Magic получила от продюсеров фильма добро на изучение возможностей. ILM создала скелет Галлимима на компьютере и анимировала тест, показывающий стадо бегущих скелетов, а затем тираннозавра, преследующего полностью визуализированное стадо Галлимима . Производственная группа была полна энтузиазма, поскольку ничего подобного ранее не было достигнуто, и Спилберга убедили вписать эту сцену в сценарий, а также использовать компьютерную графику для других кадров с динозаврами в фильме вместо покадровой анимации. [48] [49] Галлимимы кадры , были анимированы путем отслеживания кадров со страусами, а на которых пасутся газели . также ссылались на [50] Килан-Яворовска, обнаружившая образец голотипа, назвала это «красивой сценой». [6] Динозавры в фильме были одним из наиболее широко разрекламированных применений компьютерных изображений в кино и считались более реалистичными, чем то, что ранее достигалось с помощью спецэффектов . [51]
Подчеркивание птичьего стадного поведения стада галлимимов было важным моментом в истории Парка Юрского периода , поскольку они должны были представлять предшественников птиц. Стадо было показано движущимся в целом, а не отдельные животные, бегающие вокруг, а меньшие Галлимимы были показаны в середине группы, как будто они были под защитой. [48] Во время этой сцены палеонтолог Алан Грант говорит, что стадо движется «равномерно меняя направление, точно так же, как стая птиц, уклоняющаяся от хищника», наблюдая за движениями быстрого и изящного Галлимима . Это контрастировало с тем, как динозавры традиционно изображались в средствах массовой информации как неуклюжие, волочащие хвост животные, и фильм помог изменить общепринятое представление о динозаврах. Эта и другие сцены отражают недавние на тот момент теории эволюции птиц, выдвинутые научным консультантом фильма, палеонтологом Джоном Р. Хорнером , идеи, которые в то время все еще вызывали споры. [52] [53] [54] Несмотря на такие теории, Галлимим и другие динозавры в фильме были изображены без перьев, отчасти потому, что в то время было неизвестно, насколько широко они были распространены среди этой группы. [52] [55]
Утверждалось, что следы карьера Ларк (одна из крупнейших в мире концентраций следов динозавров) в Квинсленде , Австралия, послужили источником вдохновения и «научной основой» для сцены панического бегства Галлимима в Парке Юрского периода ; Первоначально эти следы были интерпретированы как изображение панического бегства динозавров, вызванного прибытием хищника-теропода. Идея о том, что следы представляют собой давку, с тех пор оспаривается («теропод» мог быть травоядным, похожим на муттабурразавра ), а консультант « Парка Юрского периода» отрицает, что следы послужили источником вдохновения для фильма. [56] [57] [58]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н Осмольская, Х.; Роневич, Э.; Барсболд, Р. (1972). «Новый динозавр Gallimimus Bullatus n. gen., n. sp. (Ornithomimidae) из верхнего мела Монголии» (PDF) . Палеонтология Полоника . 27 : 103–143.
- ^ Килан-Яворовска, З.; Довчин, Н. (1968). «Рассказ о польско-монгольских палеонтологических экспедициях 1963–1965 годов» (PDF) . Палеонтологическая Полоника : 7–30.
- ^ Jump up to: а б Градзихски Р.; Казмерчак Дж.; Лефельд, Дж. (1969). «Географические и геологические данные польско-монгольских палеонтологических экспедиций» (PDF) . Палеонтологическая Полоника . 19 : 33–82.
- ^ Додсон, П. (1998). Рогатые динозавры: естественная история . Принстон: Издательство Принстонского университета. п. 9. ISBN 978-0-691-05900-6 .
- ^ Jump up to: а б с Килан-Яворовска, З. (1975). «Млекопитающие и динозавры позднего мела из пустыни Гоби: окаменелости, раскопанные польско-монгольскими палеонтологическими экспедициями 1963–71 годов, проливают новый свет на примитивных млекопитающих и динозавров, а также на обмен фауной между Азией и Северной Америкой». Американский учёный . 63 (2): 150–159. Бибкод : 1975AmSci..63..150K . JSTOR 27845359 .
- ^ Jump up to: а б с д и Килан-Яворовска, З. (2013). В поисках ранних млекопитающих . Жизнь прошлого. Блумингтон: Издательство Университета Индианы. стр. 47–48, 50–51. ISBN 978-0-253-00824-4 .
- ^ Jump up to: а б Перенасыщение, Д.Ф. (1997). Динозавры: Энциклопедия . Джефферсон: McFarland & Co., стр. 429–432 . ISBN 978-0-89950-917-4 .
- ^ Jump up to: а б с д и ж Кобаяши, Ю.; Барсболд, Р. (2006). «Орнитомимиды из немегтинской свиты Монголии» (PDF) . Журнал Палеонтологического общества Кореи . 22 (1): 195–207.
- ^ «Меловые монгольские динозавры» . DinoCasts.com. Архивировано из оригинала 2 февраля 2015 года.
- ^ Восток, СГ; Хансен, С.; Ост, Т. (1997). «Динозавры, яйца и детеныши – специальная выставка о затерянном мире» (PDF) . GeologiskNyt (на датском языке). 6 : 24–25. Архивировано из оригинала (PDF) 14 мая 2018 года . Проверено 13 мая 2018 г.
- ^ Фанти, Ф.; Белл, PR; Тайге, М.; Милан, Луизиана; Динелли, Э. (2018). «Геохимическая дактилоскопия как инструмент репатриации окаменелостей динозавров, добытых браконьерами в Монголии: тематическое исследование для местности Нэмэгт, пустыня Гоби». Палеогеография, Палеоклиматология, Палеоэкология . 494 : 51–64. Бибкод : 2018PPP...494...51F . дои : 10.1016/j.palaeo.2017.10.032 .
- ^ Jump up to: а б Ли, Х.-Дж.; Ли, Ю.-Н.; Адамс, ТЛ; Карри, Пи Джей; Кобаяши, Ю.; Джейкобс, LL; Коппельхус, Э.Б. (2018). «Следы теропода, связанные со скелетом стопы галлимима из формации Нэмэгт, Монголия». Палеогеография, Палеоклиматология, Палеоэкология . 494 : 160–167. Бибкод : 2018PPP...494..160L . дои : 10.1016/j.palaeo.2017.10.020 .
- ^ Ганнон, М. (2014). «Украденное «гнездо динозавров» вернулось в Монголию» . uk.news.yahoo.com . LiveScience.com . Проверено 21 мая 2019 г.
- ^ Jump up to: а б с д и Пол, GS (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. стр. 384–386, 393–394 . ISBN 978-0-671-61946-6 .
- ^ Jump up to: а б Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон: Издательство Принстонского университета. п. 131. ИСБН 978-0-691-16766-4 .
- ^ Пол, GS (2010). Принстонский полевой справочник по динозаврам (1-е изд.). Принстон: Издательство Принстонского университета. стр. 112–114 . ISBN 978-0-691-13720-9 .
- ^ Jump up to: а б с д и ж Маковицкий, П.Дж.; Кобаяши, Ю.; Карри, Пи Джей (2004). «Орнитомимозаврия». В Вейшампеле, Д.Б.; Додсон, П.; Осмольская, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 137–150 . ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б Чинзориг, Цогтбаатар; Каллен, Томас; Филлипс, Джордж; Рольке, Ричард; Занно, Линдси Э. (19 октября 2022 г.). «Крупнотелые орнитомимозавры населяли Аппалачи в позднем меловом периоде Северной Америки» . ПЛОС Один . 17 (10). е0266648. дои : 10.1371/journal.pone.0266648 . ПМЦ 9581415 .
- ^ Кристиансен, Пер (1998). «Значения показателей силы длинных костей теропод с комментариями к пропорциям конечностей и беговому потенциалу» (PDF) . Гайя . 15 : 241–255. ISSN 0871-5424 .
- ^ Jump up to: а б Зеленицкий, ДК; Терриен, Ф.; Эриксон, генеральный директор; ДеБур, CL; Кобаяши, Ю.; Эберт, Д.А.; Хэдфилд, Ф. (2012). «Пернатые нептичьи динозавры из Северной Америки дают представление о происхождении крыльев». Наука . 338 (6106): 510–514. Бибкод : 2012Sci...338..510Z . дои : 10.1126/science.1225376 . ПМИД 23112330 . S2CID 2057698 .
- ^ Jump up to: а б с Кобаяши, Ю.; Лю, Ж.-К. (2003). «Новый динозавр-орнитомимид со стадным поведением из позднего мела Китая» . Acta Palaeontologica Polonica . 48 (2): 235–259.
- ^ Jump up to: а б Барретт, премьер-министр (2005). «Питание динозавров-страусов (Theropoda: Ornihomimosauria)» . Палеонтология . 48 (2): 347–358. дои : 10.1111/j.1475-4983.2005.00448.x .
- ^ Jump up to: а б с д Хурум, Дж. (2001). «Нижняя челюсть Gallimimus Bullatus » . В Танке, DH; Карпентер, К.; Скрепник, М.В. (ред.). Жизнь мезозойских позвоночных . Блумингтон: Издательство Университета Индианы. стр. 34–41 . ISBN 978-0253339072 .
- ^ Jump up to: а б Норелл, Массачусетс; Маковицкий, П.Дж.; Карри, Пи Джей (2001). «Клювы страусиных динозавров». Природа . 412 (6850): 873–874. Бибкод : 2001Natur.412..873N . дои : 10.1038/35091139 . ПМИД 11528466 . S2CID 4313779 .
- ^ Jump up to: а б с Ватанабэ, А.; Евгения Леоне Голд, М.; Брусатте, СЛ; Бенсон, РБЖ; Шуаньер, Ж.; Дэвидсон, А.; Норелл, Массачусетс; Классенс, Л. (2015). «Пневматичность позвонков у орнитомимозавра Archaeornithomimus (Dinosauria: Theropoda), выявленная с помощью компьютерной томографии, и переоценка осевой пневматичности у орнитомимозаврии» . ПЛОС ОДИН . 10 (12): e0145168. Бибкод : 2015PLoSO..1045168W . дои : 10.1371/journal.pone.0145168 . ПМЦ 4684312 . ПМИД 26682888 .
- ^ Макфитерс, Б.; Райан, MJ; Шредер-Адамс, К.; Каллен, ТМ (2016). «Новый теропод-орнитомимид из формации Парка динозавров в Альберте, Канада». Журнал палеонтологии позвоночных . 36 (6): e1221415. Бибкод : 2016JVPal..36E1415M . дои : 10.1080/02724634.2016.1221415 . S2CID 89242374 .
- ^ Хендрикс, К.; Хартман, SA; Матеус, О. (2015). «Обзор открытий и классификации нептичьих теропод» . Журнал палеонтологии позвоночных PalArch . 12 (1): 1–73. Архивировано из оригинала 22 июня 2018 года . Проверено 16 апреля 2018 г.
- ^ Хольц, Т.Р. (1994). «Филогенетическое положение тираннозаврид: значение для систематики теропод». Журнал палеонтологии . 68 (5): 1100–1117. дои : 10.1017/S0022336000026706 . JSTOR 1306180 .
- ^ Хольц, Т.Р. (1998). «Новая филогения хищных динозавров». ГАЙЯ . 15 :5–61. CiteSeerX 10.1.1.211.3044 . ISSN 0871-5424 .
- ^ Нэйш, Д.; Мартилл, DM; Фрей, Э. (2004). «Экология, систематика и биогеографические взаимоотношения динозавров, включая нового теропода, из формации Сантана (? Альб, ранний мел) Бразилии». Историческая биология . 16 (2–4): 57–70. CiteSeerX 10.1.1.394.9219 . дои : 10.1080/08912960410001674200 . S2CID 18592288 .
- ^ Рассел, Д.А. (1972). «Страусовые динозавры из позднего мела Западной Канады». Канадский журнал наук о Земле . 9 (4): 375–402. Бибкод : 1972CaJES...9..375R . дои : 10.1139/e72-031 .
- ^ Талборн, РА (1982). «Скорость и походка динозавров». Палеогеография, Палеоклиматология, Палеоэкология . 38 (3–4): 227–256. Бибкод : 1982PPP....38..227T . дои : 10.1016/0031-0182(82)90005-0 .
- ^ Николлс, Эл.; Рассел, AP (1985). «Строение и функции грудного пояса и передней конечности Struthiomimus altus (Theropoda: Ornithomimidae)» . Палеонтология . 28 (4): 64–677.
- ^ Мэдсен, ЭК (2007). «Морфология клюва современных птиц и влияние на морфологию клюва орнитомимид» . Det Matematisk-Naturvitenskapelige Fakultet – Диссертация : 1–21.
- ^ Баттон, диджей; Занно, LE (2019). «Повторяющаяся эволюция различных способов травоядности у нептичьих динозавров» . Современная биология . 30 (1): 158–168.e4. дои : 10.1016/j.cub.2019.10.050 . ПМИД 31813611 . S2CID 208652510 .
- ^ Павлицкий, Р.; Болечала, П. (1987). «Рентгеновский микроанализ костей ископаемых динозавров: возрастные различия в содержании кальция и фосфора в костях Gallimimus Bullatus ». Folia Histochemica et Cytobiologica . 25 (3–4): 241–244. ПМИД 3450541 .
- ^ Павлицкий, Р.; Болечала, П. (1991). «Рентгеновский микроанализ костей ископаемых динозавров: возрастные различия в содержании свинца, железа и магния». Folia Histochemica et Cytobiologica . 29 (2): 81–83. ПМИД 1804726 .
- ^ Ренсбергер, Дж. М.; Ватабе, М. (2000). «Тонкая структура костей динозавров, птиц и млекопитающих». Природа . 406 (6796): 619–622. Бибкод : 2000Natur.406..619R . дои : 10.1038/35020550 . ПМИД 10949300 . S2CID 4384549 .
- ^ Jump up to: а б с Ежикевич, Т.; Рассел, Д.А. (1991). «Позднемезозойская стратиграфия и позвоночные животные бассейна Гоби» . Меловые исследования . 12 (4): 345–377. дои : 10.1016/0195-6671(91)90015-5 .
- ^ Салливан, РМ (2006). «Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia)» (PDF) . В Лукасе, Спенсер Г.; Салливан, Роберт М. (ред.). Позднемеловые позвоночные Западного Внутреннего региона . Том. 35. Бюллетень Музея естественной истории и науки Нью-Мексико. стр. 347–366.
- ^ Градштейн, FM; Огг, Дж.Г.; Смит, АГ (2005). Геологическая шкала времени, 2004 г. Кембридж: Издательство Кембриджского университета. стр. 344–371 . дои : 10.1017/CBO9780511536045 . ISBN 978-0-521-78142-8 .
- ^ Хольц, ТР (2014). «Палеонтология: Тайна ужасных рук раскрыта». Природа . 515 (7526): 203–205. Бибкод : 2014Natur.515..203H . дои : 10.1038/nature13930 . ПМИД 25337885 . S2CID 205241353 .
- ^ Новачек, М. (1996). Динозавры Пылающих скал . Нью-Йорк: Якорь. п. 133 . ISBN 978-0-385-47775-8 .
- ^ Яворовский З.; Пенско, Дж. (1967). «Необычайно радиоактивные ископаемые кости из Монголии». Природа . 214 (5084): 161–163. Бибкод : 1967Natur.214..161J . дои : 10.1038/214161a0 . S2CID 4148855 .
- ^ Efimov, M. B. (1983). "Peresmotr iskopayemykh krokodilov Mongolii" [Revision of the fossil crocodiles of Mongolia]. Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya Trudy (in Russian). 24 : 76–96.
- ^ Хурум, Дж. Х.; Сабат, К. (2003). «Гигантские динозавры-тероподы из Азии и Северной Америки: сравнение черепов Tarbosaurus bataar и Tyrannosaurus rex » (PDF) . Acta Palaeontologica Polonica . 48 (2): 188.
- ^ Хольц, Т.Р. (2004). «Тираннозавроидея». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. п. 124 . ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б Шей, Д.; Дункан, Дж. (1993). Создание Парка Юрского периода . Нью-Йорк: Boxtree Ltd., стр. 48–52, 135–136. ISBN 978-1-85283-774-7 .
- ^ Крайтон, М. (1990). Парк Юрского периода . Лондон: Группа Random Century. стр. 137–138 . ISBN 978-0-394-58816-2 .
- ^ Бэрд, Р. (1998). «Анимализация динозавров Парка Юрского периода : схемы блокбастеров и межкультурное познание на сцене угроз». Киножурнал . 37 (4): 82–103. дои : 10.2307/1225728 . JSTOR 1225728 .
- ^ Принц, С. (1996). «Правдивая ложь: перцептивный реализм, цифровые изображения и теория кино». Film Quarterly (Представлена рукопись). 49 (3): 27–37. дои : 10.2307/1213468 . hdl : 10919/25811 . JSTOR 1213468 .
- ^ Jump up to: а б Монтанари, С. (2015). «Что мы узнали о динозаврах после «Парка Юрского периода выхода »» . Форбс . Проверено 7 мая 2018 г.
- ^ Кирби, Д.А. (2011). Лабораторные халаты в Голливуде: наука, ученые и кино . Массачусетс: MIT Press . стр. 127–129 . ISBN 978-0262294867 .
- ^ Кирби, Д.А. (2003). «Научные консультанты, художественные фильмы и научная практика». Социальные исследования науки . 33 (2): 231–268. дои : 10.1177/03063127030332015 . JSTOR 3183078 . S2CID 111227921 .
- ^ Филлипс, И. (2015). «Как динозавры из Мира Юрского периода выглядели в реальной жизни» . Бизнес-инсайдер . Проверено 3 декабря 2016 г.
- ^ Мартин, Эй Джей (2014). Динозавры без костей: жизнь динозавров, обнаруженная по их окаменелостям . Лондон: Книги Пегаса. стр. 66–67. ISBN 978-1605984995 .
- ^ Ромилио, А. (2015). «Побег динозавров остановился» . Австралазийская наука . 36 (2): 24–27. ISSN 1442-679X .
- ^ Шапиро, AD (2013). «Великая паника динозавров, которой никогда не было?» . Новости Би-би-си . Проверено 7 мая 2018 г.
Внешние ссылки
[ редактировать ] СМИ, связанные с Галлимимом, на Викискладе?
СМИ, связанные с Галлимимом, на Викискладе?















| Базы данных органов управления : Национальные |
|---|