Анкилозавриды
| Анкилозавриды Временной диапазон: ранний мел – поздний мел ,
| |
|---|---|

| |
| Установленный скелет Scolosaurus thronus , Королевский палеонтологический музей Тиррелла. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Тиреофора |
| Клэйд : | † Анкилозаврия |
| Клэйд : | † Эуанкилозаврия |
| Семья: | † Анкилозавриды Браун , 1908 год. |
| Типовой вид | |
| † Анкилозавр магнивентрис Браун, 1908 год.
| |
| Подгруппы | |
| |
| Синонимы | |
| |
Ankylosauridae ( / ˌ æ ŋ k ɪ l oʊ ˈ s ɔː r ɪ d iː / ) — семейство панцирных динозавров в составе Ankylosauria , родственной группе Nodosauridae . Самые старые известные анкилозавриды появились примерно 122 миллиона лет назад и вымерли 66 миллионов лет назад во время мел-палеогенового вымирания . [1] Эти животные были в основном травоядными и представляли собой облигатные четвероногие животные с листовидными зубами и крепким, покрытым щитками телом. Анкилозавриды обладают отчетливо куполообразной и короткой мордой, клиновидными остеодермами на черепе, щитками вдоль туловища и хвостовой булавой. [2]
Ankylosauridae известны исключительно из северного полушария, экземпляры встречаются в западной части Северной Америки, Европе и Восточной Азии. Первые открытия этого семейства были сделаны Питером Кайзером и Барнумом Брауном в Монтане в 1906 году. [3] В 1908 году Браун назвал Ankylosauridae и подсемейство Ankylosaurinae .
Анатомия
[ редактировать ]
Анкилозавриды — толстые, крепко сложенные, бронированные динозавры. Они обладают дополнительными оссификациями на костях черепа, которые закрывают некоторые отверстия черепа и образуют клиновидные, роговидные структуры. Вдоль туловища анкилозаврид расположены ряды щитков, заполненных более мелкими косточками , образующими сросшийся панцирный щит. [2] На шее можно обнаружить только два воротника панцирных пластин, в отличие от сестринской группы нодозаврид, у которых их три. [1] Nodosauridae и Ankylosauridae также обладают уникальным свойством обилия структурных волокон как в первичной, так и во вторичной кости. [4] Анкилозавриды также имеют S-образный носовой ход. [1]
Наиболее отличительной особенностью анкилозаврид является наличие хвостовой булавы . Он состоит из модифицированных переплетенных дистальных хвостовых позвонков и увеличенных луковичных остеодерм. [5] «Ручка» хвостовой булавы затрагивает позвонки и требует, чтобы удлиненные презигапофизы перекрывали по крайней мере половину длины предшествующего центра позвонка. [5] Эти дистальные хвостовые позвонки также лишены поперечных отростков и нервных отростков, поэтому их длина обычно превышает ширину; обратное верно для проксимальных хвостовых позвонков. [5] Производные анкилозаврид имеют слияние задних спинных, крестцовых, а иногда и передних хвостовых позвонков, которое образует единую структуру, называемую «синсакрум-комплекс». Наблюдается полное слияние центров, нервных дуг, зигапофизов и иногда нервных отростков. [6]
В 2017 году Виктория М. Арбор и Дэвид К. Эванс описали новый род анкилозаврин, у которого сохранились обширные мягкие ткани по всему телу. Это животное, названное Зуулом в честь его сходства с монстром из Охотников за привидениями , также является первым анкилозавром из формации реки Джудит. [7]
История обучения
[ редактировать ]
Барнум Браун и Питер Кайсен обнаружили первый род анкилозаврид, Ankylosaurus , в 1906 году в пластах Хелл-Крик в Монтане. [3] Ископаемый материал, который они нашли, представлял собой часть черепа, два зуба, несколько позвонков, деформированную лопатку, ребра и более тридцати остеодерм. [3] Реконструкция экземпляра первоначально была встречена со скептицизмом теми, кто считал его, по крайней мере, очень близким к роду Stegopelta или полностью его частью , а сам Браун поместил его в подотряд Stegosauria. [3]
Ранее было интерпретировано, что вариации формы хвостовой булавы анкилозаврид обусловлены половым диморфизмом , который предполагает, что морфология хвостовой булавы имеет сцепленную с полом внутривидовую функцию. [6] Это возможно, если хвостовая дубинка использовалась для агонистического поведения. Однако теория полового диморфизма предсказывает примерно равное количество особей с хвостовыми булавами двух разных размеров. Очевидный половой диморфизм не задокументирован, но если булавы одного пола намного больше, то будет иметь место предвзятость к сохранению и открытию в сторону этого пола. [6] [8]
Филогения
[ редактировать ]
В 1978 году У.П. Кумбс-младший отнес почти все действующие виды Ankylosauria к Nodosauridae или Ankylosauridae. [9] Это было ключевое исследование, в котором были описаны многие признаки анкилозавров в самых ранних филогенетических анализах этой группы.
Позже в 1998 году Пол Серено официально определил Ankylosauridae как всех анкилозавров, более тесно связанных с Ankylosaurus, чем с Panoplosaurus . [10] Анкилозавры, не имеющие хвостовой булавы, были включены в филогению Ankylosauridae Кеннета Карпентера в 2001 году. [11]
В исследовании, проведенном в 2004 году Викариусом и др., Minmi были зарегистрированы как Gargoyleosaurus, Gastonia и базальные анкилозавриды, а остальные анкилозавриды были заполнены Gobisaurus , Shamosaurus и анкилозаврами из Китая, Монголии и Северной Америки. [12]
В 2012 году Томпсон и др. предпринял анализ почти всех известных на тот момент действительных анкилозавров и таксонов внешней группы. [13] В основу своей полученной филогении они положили признаки, представляющие краниальную, посткраниальную и остеодермальную анатомию, а также детали синапоморфий для каждой обнаруженной клады. В этом исследовании Gargoyleosaurus и Gastonia были помещены в базальные Nodosauridae, а Cedarpelta и Liaoningosaurus - в базальные анкилозавриды. [13]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

В 2016 году Арбор и Карри представили филогенетический анализ анкилозаврид, состоящих из Gastonia , Cedarpelta , Chuanqilong , других базальных анкилозаврид и ряда производных анкилозаврид. Их филогения включает некоторые неопределенные филогенетические отношения между анкилозавром , анодонтозавром , сколозавром и зиапельтой . [14]
| |||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]Поза и передвижение
[ редактировать ]Анкилозавриды, вероятно, были очень медлительными животными. У всех Ankylosauria малоберцовая кость более тонкая, чем большеберцовая кость , что позволяет предположить, что большеберцовая кость несла большую часть веса животного, в то время как малоберцовая кость служила местом прикрепления мышц. [15] Мышцы задних конечностей Euoplocephalus были восстановлены, а расположение нескольких мышц, прикрепляющихся к бедренной кости, имеет очень короткий момент плеч . Мышцы, прикрепляющиеся к большеберцовой и малоберцовой костям, имеют более длинные моментные плечи. Такое расположение мышц-втягивателей указывает на слоновью походку, соответствующую столбчатой позе. [15]
Восстановление передних конечностей Euoplocephalus демонстрирует сходство с мускулатурой передних конечностей крокодила. [16] Наиболее хорошо развитые мышцы грудной области выполняли скорее опорную функцию, чем вращательную. Также было высказано предположение, что запястные и пястные кости имеют сходство с таковыми четвероногих, имеющих роющую (роющую) привычку. [16]
Несколько мышц задней части анкилозаврид (dorsalis caudae, подвздошно-каудалис, копчико-бедренная короткая, копчико-бедренная длинная, подвздошно-большеберцовая и седалищная каудальная) использовались для движения хвоста и хвостовой булавы. [15] Анкилозавриды имеют тенденцию иметь горизонтальную, а не наклонно-вертикальную ориентацию зигапофизарных сочленений свободных хвостовых позвонков хвоста. Такое расположение наиболее эффективно для поперечной, а не вертикальной мобильности. [6] Отсутствие мускулатуры, позволяющей поднимать хвост, и такая ориентация зигапофизов позволяют предположить, что хвост и его булава двигались параллельно земле и немного над ней. [6]
Биогеография
[ редактировать ]
Географическое происхождение анкилозаврид в настоящее время установить затруднительно. Существует смесь базальных анкилозаврид как из Северной Америки, так и из Азии, что подтверждается общепринятыми кладистическими анализами. [17] По-видимому, в середине мела азиатские нодозавриды были заменены анкилозавриновыми анкилозавридами. [14] Некоторые исследователи постулируют, что анкилозаврины мигрировали в Северную Америку из Азии между альбом и кампаном , где они диверсифицировались в кладу анкилозаврин, характеризующуюся изогнутыми мордами и плоскими черепными костными пластинами (caputegulae). [14] Нет никаких свидетельств существования анкилозаврид в Гондване . [14]
Вариация
[ редактировать ]У Ankylosauridae существует множество индивидуальных и межвидовых различий в выраженности панциря. Однако наиболее изученным аспектом панциря анкилозаврид является хвостовая булава. значительная онтогенетическая В морфологии этого признака обнаружена входило по меньшей мере 16 хвостовых позвонков и индивидуальная изменчивость. В рукоятку хвостовой булавы Pinacosaurus grangeri , а у Euoplocephalus примерно 9–11 окостеневших хвостовых позвонков. [6]
Вариации формы, толщины и длины хвостового выступа объясняются индивидуальными вариациями, таксономией или представлением различных фаз роста. [6] Однако с этим последним аспектом возникают трудности, поскольку известные булавы не соответствуют единой серии роста, хотя некоторые различия должны быть онтогенетическими и аллометрическими . [6] [8]
Образ жизни
[ редактировать ]Зубы большинства анкилозаврид имели листовидную форму, что предполагает в основном травоядную диету. Их зубы могли быть гладкими или рифлеными, а также различаться на губной и язычной поверхностях. [18] Euoplocephalus tutus имеет гребни и бороздки на зубах, не имеющие никакого отношения к их краевым буграм. [18] Учитывая, что шея и голова обращены вниз, анкилозавриды вполне вероятно питаются пастбищным способом. [1]
Однако у некоторых видов были замешаны нетравоядные привычки. Предполагается, что пинакозавр был насекомоядным, похожим на муравьеда с длинным языком. [19] в то время как Лиаонингозавр был предложен как рыбоядное животное . И то, и другое было бы исключительным свидетельством плотоядности среди птицетазовых .

Существует несколько преобладающих теорий функции хвостовой булавы анкилозаврид. Первый — агонистическое поведение внутри вида. [6] У большинства позвоночных, включая динозавров, такое поведение сопровождается структурами для демонстрации или боя. Некоторые исследователи полагают, что это явление было бы неправдоподобным, учитывая, что не существует современного аналога четвероногих, использующего для этой цели хвост. Вместо этого эти палеонтологи предполагают, что анкилозавриды использовали свой широкий плоский череп для бодания головами между людьми. [6]
Вторая теория функции хвостовой булавы заключается в защите от хищников. Было высказано предположение, что дубинка будет наиболее эффективна против плюсневых костей атакующего теропода. [6] [15]
Кости, образующие черепные украшения, имеют физиологические затраты, поэтому их производство просто для защиты от хищников было бы неэффективно. Поэтому была выдвинута теория, что эти клиновидные остеодермы могут поддерживать интерпретацию, частично выбранную половым путем . [8]
Хронология родов
[ редактировать ]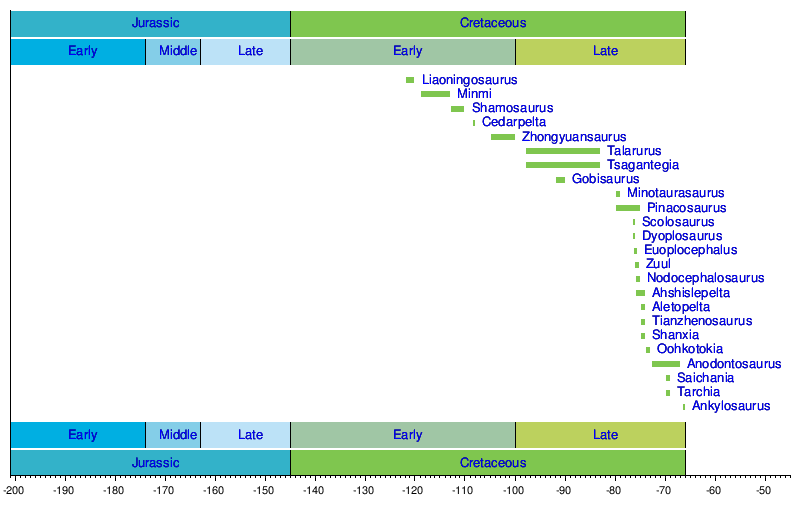
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Фастовский, Дэвид Э.; Вейшампель, Дэвид Б. (2012). Динозавры: краткая естественная история (2-е изд.). Кембридж: Издательство Кембриджского университета.
- ^ Jump up to: а б Серено, Пол К. (25 июня 1999 г.). «Эволюция динозавров». Наука . 284 (5423): 2137–2147. дои : 10.1126/science.284.5423.2137 . ISSN 0036-8075 . ПМИД 10381873 .
- ^ Jump up to: а б с д Барнум., Браун; К., Кайсен, Питер (1 января 1908 г.). «Ankylosauridae, новое семейство панцирных динозавров из верхнего мела. Бюллетень AMNH; т. 24, статья 12». hdl : 2246/1435 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Штейн, Мартина; Хаяси, Сёдзи; Сандер, П. Мартин (24 июля 2013 г.). «Гистология длинных костей и закономерности роста анкилозавров: значение для истории жизни и эволюции» . ПЛОС ОДИН . 8 (7): e68590. Бибкод : 2013PLoSO...868590S . дои : 10.1371/journal.pone.0068590 . ISSN 1932-6203 . ПМЦ 3722194 . ПМИД 23894321 .
- ^ Jump up to: а б с Арбур, Виктория М.; Карри, Филип Дж. (01 октября 2015 г.). «Хвостовые булавы динозавров-анкилозаврид развились путем постепенного приобретения ключевых особенностей» . Журнал анатомии . 227 (4): 514–523. дои : 10.1111/joa.12363 . ISSN 1469-7580 . ПМК 4580109 . ПМИД 26332595 .
- ^ Jump up to: а б с д и ж г час я дж к Кумбс, Уолтер П. младший (1 июля 1995 г.). «Хвостовые булавы анкилозавров среднего кампанского и раннего маастрихтского периода из западной части Северной Америки, с описанием крошечной булавы из Альберты и обсуждением ориентации хвоста и функции хвостовой булавы». Канадский журнал наук о Земле . 32 (7): 902–912. Бибкод : 1995CaJES..32..902C . дои : 10.1139/e95-075 . ISSN 0008-4077 .
- ^ Свитек, Брайан (08 мая 2017 г.). «Представляем Зуула, анкилозавра, от которого действительно могут болеть лодыжки» . Смитсоновский институт . Проверено 15 января 2018 г.
- ^ Jump up to: а б с М, Арбур, Виктория (июнь 2014 г.). «Систематика, эволюция и биогеография динозавров-анкилозаврид» . Библиотеки Университета Альберты: Архив образования и исследований. дои : 10.7939/R31N7XW06 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кумбс, В.П. младший (1978). «Семейства отряда птицетазовых динозавров Ankylosauria». Палеонтология : 143–70.
- ^ Серено, Пол (1998). «Обоснование филогенетических определений с применением к таксономии динозавров более высокого уровня». Новогодний альбом по геологии и палеонтологии, статьи : 41–83. дои : 10.1127/njgpa/210/1998/41 .
- ^ Карпентер, К. (2001). «Филогенетический анализ анкилозаврии». В Карпентер, К. (ред.). Бронированные динозавры . Издательство Университета Индианы. стр. 455–483.
- ^ Викариус, Мэтью К.; Марианска, Тереза ; Вейшампель, Дэвид Б. (12 июня 2004 г.). Анкилозавр . Издательство Калифорнийского университета. стр. 363–392. дои : 10.1525/Калифорния/9780520242098.003.0020 . ISBN 9780520941434 .
- ^ Jump up to: а б Томпсон, Ричард С.; Пэриш, Джолион К.; Мейдмент, Сюзанна ЧР; Барретт, Пол М. (1 июня 2012 г.). «Филогения анкилозавровых динозавров (Ornithischia: Thyreophora)». Журнал систематической палеонтологии . 10 (2): 301–312. дои : 10.1080/14772019.2011.569091 . ISSN 1477-2019 . S2CID 86002282 .
- ^ Jump up to: а б с д Арбур, Виктория М.; Карри, Филип Дж. (3 мая 2016 г.). «Систематика, филогения и палеобиогеография динозавров-анкилозаврид». Журнал систематической палеонтологии . 14 (5): 385–444. дои : 10.1080/14772019.2015.1059985 . ISSN 1477-2019 . S2CID 214625754 .
- ^ Jump up to: а б с д Кумбс, В. П. младший (1979). «Остеология и миология задних конечностей при анкилозаврии (рептилии, орнитишии)». Журнал палеонтологии . 53 (3): 666–684. JSTOR 1304004 .
- ^ Jump up to: а б Кумбс, Уолтер П. (1 января 1978 г.). «Мышцы передних конечностей анкилозавров (рептилий, орнитиший)». Журнал палеонтологии . 52 (3): 642–657. JSTOR 1303969 .
- ^ Арбур, Виктория (23 августа 2015 г.). «Знай своих анкилозавров: в этом выпуске все вместе» . Псевдоплоцефалия .
- ^ Jump up to: а б Карпентер, Кеннет; Карри, Филип Дж. (1990). Систематика динозавров: подходы и перспективы . Издательство Кембриджского университета.
- ^ Хилл, Р., Д'Эмик, М., Бевер, Г., Норелл, М. 2015. Сложный гиобранхиальный аппарат мелового динозавра и древность птичьих параглоссалий. Зоологический журнал Линнеевского общества. дои: 10.1111/zoj.12293
- Динозавры и другие доисторические существа , под редакцией Ингрид Крэнфилд (2000), книги о саламандрах, стр. 250–257.
- Карпентер К. (2001). «Филогенетический анализ анкилозавров». В Карпентер, Кеннет (ред.). Бронированные динозавры . Издательство Университета Индианы. стр. 455–484. ISBN 978-0-253-33964-5 .
- Киркланд, Дж.И. (1996). Биогеография фаун среднего мела западной части Северной Америки - потеря европейских связей и первый великий азиатско-североамериканский обмен. Дж. Вертебр. Палеонтол. 16 (дополнение к 3): 45А
Внешние ссылки
[ редактировать ]- Семейное древо
- Веб-проект Ankylosauridae «Древо жизни»
- Pseudoplocephalus , Блог биолога-эволюциониста и палеонтолога позвоночных, специализирующегося на анкилозаврах.
