Чуаньцилун
| Чуаньцилун Временной диапазон: ранний мел
| |
|---|---|
| |
| Образец голотипа | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Тиреофора |
| Клэйд : | † Анкилозаврия |
| Семья: | † Анкилозавриды |
| Род: | † Чуаньцилун Хан и др., 2014 г. |
| Типовой вид | |
| † Чуаньцилун чаоянгенсис Хан и др., 2014 г.
| |
Чуаньцилун (что означает «легендарный дракон») — моновидовой род базальных анкилозавридных динозавров из провинции Ляонин , Китай , который жил в раннем меловом периоде (от позднего баррема до аптского этапа, от 122,0 до 118,9 млн лет назад) на территории нынешней формации Цзюфотанг . Тип Chuanqilong и единственный вид, chaoyangensis , известен по почти полному скелету с черепом молодой особи. Его описали в 2014 году Фэнлу Хань, Вэньцзе Чжэн, Дунъюй Ху, Син Сюй и Пол М. Барретт. Chuanqilong имеет много общего с Liaoningosaurus и может представлять собой более позднюю онтогенетическую стадию таксона.
Чуаньцилун среднего размера был анкилозавром , предполагаемая длина которого составляла 4,5 метра (14,8 фута), хотя предполагалось, что он мог быть больше из-за незрелого возраста типового экземпляра. У него был треугольный череп и шея, защищенная полосами остеодерм, известными как шейные полукольца. Остальная часть тела была покрыта остеодермами и косточками различной формы и размера. В отличие от производных анкилозаврид, на конце его хвоста не было булавы . Как и другие анкилозавриды, он был четвероногим , с крепкими передними и задними конечностями .
Открытие и присвоение имени
[ редактировать ]
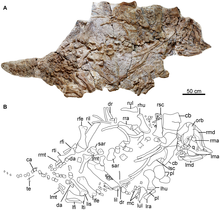
Почти полный скелет был собран местными фермерами из единственного карьера в провинции Ляонин , Китай . Скелет был извлечен из формации Цзюфотанг , которая датируется позднебарремским и аптским этапами раннего мелового периода, от 122,0 до 118,9 млн лет назад. Образец был назван и описан в 2014 году Фэнлу Ханем, Вэньцзе Чжэн, Дунъюй Ху, Син Сюй и Полом М. Барреттом. Образец голотипа , CJPM V001 , состоит из почти полного черепа и скелета , при этом отсутствует только дистальная часть хвостового ряда , и представляет собой молодую особь. Авторы отметили, что этот экземпляр находился на более поздней онтогенетической стадии, чем экземпляры симпатрического лиаонингозавра , исходя из большего размера типового экземпляра, размера глазницы и количества зубов . Образец сохранился в двухмерном виде, видна только брюшная сторона. Большая часть черепа сжата дорсовентрально, большая часть позвоночного столба разобщена, при этом конечности сохранены в сочленении. Типовой экземпляр в настоящее время хранится в Палеонтологическом музее Чаоян Цзизантанг, а слепок образца (IVPP FV 1978) хранится в Палеонтологическом музее Чаоян Цзизантанг. Институт палеонтологии и палеоантропологии позвоночных . [ 1 ]
Родовое китайских название Чуаньцилун происходит от слов « Чуаньци » (легендарный), что связано с обилием окаменелостей западного Ляонина, и « длинный » (дракон). Видовое название chaoyangensis относится к более широкой географической области , включающей типовую местность. [ 1 ]
В 2014 году в формации Цзюфотанг были описаны отпечатки лопаток и плечевой кости, принадлежащие неопределенному анкилозавру с предполагаемой длиной тела 6,0–8,6 метра (19,7–18,2 фута), и на тот момент это был первый анкилозавр, описанный из формации Цзюфотанг. формирование. [ 2 ] В абстрактной книге Канадского общества палеонтологии позвоночных, опубликованной в 2019 году, упоминается, что тип и единственный известный экземпляр Чуаньцилуна на самом деле представляет собой взрослую особь, в отличие от интерпретации Хана и др. (2014) и, возможно, был лиаонингозавром на другой, более поздней онтогенетической стадии. [ 3 ]
Описание
[ редактировать ]Размер и отличительные черты
[ редактировать ]Хан и др. (2014) оценили Чуаньцилуна длину в 4,5 метра (14,8 фута). Однако авторы предположили, что он мог достигать более крупных размеров, поскольку типовой экземпляр представляет собой молодую особь. [ 1 ]
Описывающие авторы указали на две отличительные черты. Оба они являются аутапоморфиями , уникальными производными персонажами. имеется В квадратной кости суставная ямка , расположенная на одном уровне с зубным рядом. Дистально суживающаяся седалищная кость сужена на средней длине. К другим отличительным признакам относятся наличие длинного ретроартикулярного отростка, наличие плечевой тонкой клиновидной слезной кости, соотношение длины кости к длине бедренной кости 0,88, ширина проксимального конца плечевой кости составляет половину длины плечевой кости. плечевой стержень и наличие подтреугольных когтей . [ 1 ]
Череп
[ редактировать ]
При взгляде снизу череп треугольной формы. Верхняя челюсть имеет неглубокий и уплощенный щечный край с выемчатым краем, а анторбитальное окно в каудодорсальной области имеется . Тонкая клиновидная слезная кость образует ростральный край глазницы , а длинная супраорбитальная кость контактирует со слезной ветвью ростровентрально. Кость, которая может состоять из чешуйчатой и заглазничной костей, имеет субпрямоугольный контур и субпараллельные бороздки. Левый квадрат имеет прямоугольную головку и прямой, а стержень образует широкую и неглубокую впадину под головкой квадрата. В отличие от нодозаврид , квадратная кость не слита с чешуйчатой. Крыловидный отросток имеет поперечно расширенный вентральный конец, состоящий из двух мыщелков нижней челюсти , и имеет субпрямоугольный контур. Как и у большинства других анкилозавров , медиальный мыщелок шире в поперечнике и простирается дальше к нижней стороне, чем латеральный мыщелок. Как и у большинства анкилозавров, за исключением Ankylosaurus , левая верхняя челюсть имеет не менее 20 альвеолы . Сохранившиеся ростральные зубы верхней челюсти меньше хвостовых зубов, их коронки столь же высоки, как и широки, а основания вздуты и имеют слабую поясную кость . Зубы лишены серповидной поясной извилины. Ростральная коронка верхнего зуба имеет небольшие зубцы и бугорки. Некоторые зубы имеют зубцы, которые сужаются и имеют круглое поперечное сечение у основания. [ 1 ]
Нижняя челюсть похожа на таковую у других базальных анкилозаврид, поскольку она длинная и неглубокая. Однако в отличие от других базальных анкилозаврид у него отсутствует остеодерма на нижнем крае. Отсутствие остеодермы могло быть результатом неполной сохранности, поскольку она могла не слиться с костью нижней челюсти из-за незрелого возраста голотипа. Если бы остеодерма присутствовала, она могла бы быть ограничена боковым углом нижней челюсти. Зубной ряд не так сильно синусоидальный, как у производных анкилозавров, и прямой. В зубной кости имеется не менее 20 альвеол, при этом большая часть зубов отсутствует. Сохранившиеся зубы аналогичны зубам верхней челюсти. Симфиз правой зубной кости слегка загнут вниз, поперечное сечение субтреугольное. Как и у нодозаврид, венечный возвышение выступает над уровнем зубного ряда. Ниже венечного возвышения находится большая приводящая ямка . Засуставной отросток длинный и тонкий, а суставной маленький. Гленоидная ямка отличается от таковой у других анкилозавров, поскольку расположена на одном уровне с зубным рядом. [ 1 ]
Посткрания
[ редактировать ]
Центры катышкообразные шейных спинных и позвонков . Шейные центры короче, чем ширина, а спинные центры длиннее, чем высота. а крестцовый центр шире, чем его длина. Все центральные части спинных позвонков лишены вентрального киля. Крестцовые ребра имеют гантелеобразную форму и крепкие. На хвостовых позвонках имеются глубокие продольные бороздки. Центр среднего хвостового позвонка при виде сбоку имеет квадратный контур. В верхней части боков центра имеется поперечный отросток, редуцированный до небольшого узловатого отростка. Нервные отростки имеют дугообразный контур и удлинены. Фасетки презигапофиза обращены краниомедиально, а постзигапофиза — каудолатерально. Хвостовые позвонки имеют нервные отростки, которые соединяются вместе с постзигапофизами , образуя каудальный отросток, который заканчивается краниально к середине следующего позвонка. Уменьшение размеров постзигапофизов совпадает с уменьшением презигапофизов. В отличие от других анкилозаврид, у Chuanqilong не было хвостовой булавы, поскольку у него отсутствуют модифицированные рукояткообразные позвонки в дистальной части хвоста. [ 1 ] [ 4 ]
Коракоид , что может представлять собой онтогенетическую особенность , не окостенен с лопаткой как это также известно у других молодых экземпляров анкилозавров. Лопатка имеет ромбовидный контур с прямым дорсальным краем и вогнутым вентральным краем. Самое узкое место лопатки находится в направлении головки суставной ямки . Вентральный край лопатки не имеет четко выраженного энтезиса, что также может представлять собой онтогенетическую особенность. Гленоидная ямка имеет овальные очертания и имеет большие размеры. короткая Плечевая кость , с большим дельтопекторальным гребнем . Проксимальный конец бедренной кости имеет ширину, значительно превышающую ширину дистального конца. Лучевой мыщелок более выражен, чем медиальный мыщелок локтевой кости, а латеральный надмыщелковый гребень недоразвит. Локтевая кость имеет клиновидный локтевой отросток, как и у других неполовозрелых экземпляров анкилозавров, и может представлять собой онтогенетическую характеристику. Лучевая кость тонкая, стержнеобразная, с дистальным концом, который шире в поперечном направлении, чем проксимальный конец. радиус . только левая кисть кисти Известна пястных костей , состоящая из четырех тонких . Из всех пястных костей III пястная кость самая длинная, а IV пястная кость короче. Остальные пястные кости примерно одинаковой длины. Самая крепкая пястная кость — это I пястная кость, а II и IV пястные кости — самые тонкие. Дистальный и проксимальный концы, если все пястные кости расширены. Ногтевые фаланги имеют треугольный очертание с заостренным кончиком, а вентральные поверхности уплощены. [ 1 ]

Предацетабулярный отросток подвздошной кости длинный и ротирован к середине, а постацетабулярный отросток ротирован в аппозиции. Предацетабулярный отросток расходится вбок от позвоночного столба и имеет прямой боковой край. Постацетабулярный отросток имеет подтреугольное очертание и короче вертлужной впадины. Лобковая ножка имеет полуокруглый и развитый профиль, тогда как седалищная ножка неразвита. Седалищная кость длинная, не имеет запирательного отростка и имеет тонкий стержень, слегка изогнутый к нижней стороне. Средняя часть седалищной кости узкая и расширяется к дистальному концу, а затем сужается дистально. Седалищная кость имеет проксимальный конец, прямой при взгляде сбоку, в отличие от выпуклой и веерообразной седалищной кости Анкилозавра и вогнутой проксимальной седалищной кости Струтиозавра . [ 1 ]
Как это типично для других анкилозавров, бедренная кость крепкая и прямая. Головка бедренной кости образует суставную поверхность округлой формы. Бедренная кость имеет как краниальный, так и большой вертлуги , которые отделены от головки бедренной кости затяжкой. Краниальный вертел отделен от большого вертела, что наблюдается у молодых экземпляров анкилозавров, но не у большинства взрослых анкилозавров. Однако это скорее может представлять собой плезиоморфный признак анкилозаврид, а также признак, находящийся под онтогенетическим контролем у некоторых анкилозавров, поскольку краниальный вертел также присутствует у некоторых нодозаврид. На бедренной кости имеется неглубокая краниальная межмыщелковая ямка . Chuanqilong имеет такое же соотношение длины плечевой кости к длине бедренной кости, как и у анкилозавра , но ниже, чем у других молодых экземпляров анкилозавра и хунгарозавров . Большеберцовая кость короче бедренной и крепкая, ее проксимальный конец имеет более слабое поперечное расширение, чем дистальный конец большеберцовой кости. Немного короче большеберцовой кости находится малоберцовая кость, она тонкая и имеет стержень овального сечения, а также относительно равный размер. Правая нога сохраняет плюсневые кости II, III и IV в сочленении. Самая длинная и крепкая плюсневая кость — это III плюсневая кость, тогда как II и IV плюсневые кости примерно одинаковой длины. Все сохранившиеся плюсневые кости имеют расширенные проксимальный и дистальный концы. Когтевые кости имеют субтреугольный контур с закругленными дистальными концами и похожи на таковые у Диоплозавра . [ 1 ]
Броня
[ редактировать ]Единственное сохранившееся шейное полукольцо Чуаньцилуна . состоит из соединительной перемычки, сросшейся в единую пластинку, но сжатой к дорсальной и вентральной сторонам и также сегментированной на четыре отдела Из этих секций правые три имеют субпрямоугольный контур и выгнуты вверх, а левая часть имеет подтреугольный профиль и сужается каудолатерально. В плечевой области сохраняются две пластинки остеодермы, крупные, плоские, утолщенные, имеющие подпрямоугольное очертание. Самая большая из этих пластинок остеодермы в два раза длиннее другой, и обе они похожи на остеодермы шейного полукольца, но также могут представлять собой отдельные пластинки шейной остеодермы. Между проксимальным концом левой локтевой кости и лучевой костью находится небольшая треугольная остеодерма, имеющая широкое основание и сужающаяся дистально. Около левой седалищной кости имеется овальная остеодерма с резким килем по средней линии. По всему телу сохранилось множество остеодерм и косточек, маленьких и неровных. [ 1 ]
Классификация
[ редактировать ]
Хан и др. (2014) первоначально обнаружили, Chuanqilong является базальным анкилозавридом, который был сестринским таксоном Liaoningosaurus что . Авторы отметили, что только две однозначные синапоморфии подтверждают его близкое родство с Liaoningosaurus , а именно наличие предглазничной ямки и гленоида лопатки, ориентированного вентрально. Авторы также отметили, что, хотя оба таксона представлены молодыми экземплярами и являются сестринскими, Chuanqilong можно отличить от Liaoningosaurus по ряду характеристик, таких как различия в соотношении длины плюсны к длине пясти и морфологии коронки щечного зуба. . [ 1 ] Кладистический анализ, проведенный Arbor & Currie (2015), выявил, что Chuanqilong либо находится в политомии с другими базальными анкилозавридами, либо как сестринский таксон Cedarpelta , положение, также подтвержденное Arbor et al. (2016). [ 5 ] [ 6 ] Анализы, проведенные Arbor & Evans (2017), Zheng et al. (2018) и Парк и др. (2019) аналогичным образом поместили Chuanqilong в политомию с другими базальными анкилозавридами, хотя включение определенных таксонов, таких как Aletopelta, во внутренний узел варьировалось. [ 7 ] [ 8 ] [ 9 ] Ривера-Сильва и др. (2018) также обнаружили его в политомии в дереве строгого консенсуса, но он также был обнаружен как родственный таксон Cedarpelta в дереве с правилом 50% большинства. В отличие от других исследований, Frauenfelder et al. (2022) обнаружили, что Chuanqilong находится в кладе, включающей Liaoningosaurus и Cedarpelta, за пределами Ankylosauridae и Nodosauridae. [ 10 ] В 2019 году в реферате было высказано предположение о возможности того, что Chuanqilong и Liaoningosaurus могут представлять один и тот же вид, но на разных стадиях онтогенетического развития. [ 3 ]
Ниже воспроизведен филогенетический анализ Arbor & Currie (2015). [ 5 ]
Палеосреда
[ редактировать ]
Останки Чуаньцилуна были обнаружены в формации Цзюфотан группы Джехол. Формация состоит из аргиллитов , алевролитов , сланцев , песчаников и туфов и перекрывает формацию Исянь . Недавний анализ циркона U-Pb методом вторичной ионной масс-спектрометрии (SIMS) позволяет предположить, что образование датируется позднебарремским - аптским этапами раннего мела , ок. 122,0–118,9 млн лет назад. [ 11 ] Формации Исянь и Цзюфотан представляют собой пресноводные озерные среды, в которых отсутствовали реки , а также многие другие переменные особенности пресноводных условий, и которые сезонно колебались между полузасушливыми и мезическими условиями. В формации Цзюфотан вулканическая активность наблюдалась нечасто , тогда как в более молодой формации Исянь активность наблюдалась чаще. Сочетание регионального вулканизма и наличия множества мелководных озер позволило обеспечить исключительную сохранность окаменелостей и сохранение отпечатков покровов, хрящей и кератина . [ 12 ]
Разнообразные формы Euornithines (такие как Mengciusornis , [ 13 ] ты ел рыбу [ 14 ] Парахонгшанорнис , [ 15 ] и Янорнис [ 16 ] ) и энантиорнитины (такие как Cuspirostrisornis , [ 17 ] Лонгиптерикс , [ 18 ] Рапаксавис , [ 19 ] Sinornis, [ 20 ] и Юанчуавис [ 21 ] ) присутствуют в формации Цзюфотанг. многочисленные птерозавры Из формации также известны , в том числе чаоянкоптериды Chaoyangopterus , [ 22 ] Эоаждарчо , [ 23 ] Джидаптерус [ 24 ] и Шэньцхоуптерус , [ 25 ] гребневик Forfexopterus , [ 26 ] анхангериды Гуидрако [ 27 ] и Лиаонингоптерус, [ 22 ] лонходраконид Икрандрако , [ 28 ] истиодактилиформы Hongshanopterus , [ 29 ] Лиаоксиптерус , [ 30 ] Линлонгоптерус [ 31 ] и Нургахий , [ 32 ] [ 33 ] тапежарид Sinopterus , [ 34 ] анурогнатид Vesperopterylus [ 35 ] и неопределенный птеродактилоид Pangupterus . [ 36 ] Другая присутствующая фауна включает jeholornithiforms Jeholornis. [ 37 ] и Компсорнис , [ 38 ] омнивороптеригиды [ 39 ] и Сапеорнис , [ 40 ] овирапторозавр Similicaudipteryx , дромеозаврид Microraptor , [ 41 ] тираннозавроид синотираннус , [ 42 ] цератопсийский Psittacosaurus , млекопитающие Fossiomanus [ 43 ] и Лиаоконодон , [ 44 ] и хористодеры Филидрозавры , [ 45 ] Икехозавр [ 46 ] и Лиаоксизавр . [ 47 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л Хан, Ф.; Чжэн, В.; Ху, Д.; Сюй, Х.; Барретт, премьер-министр (2014). «Новый базальный анкилозаврид (Dinosauria: Ornithischia) из нижнемеловой формации Цзюфотанг провинции Ляонин, Китай» . ПЛОС ОДИН . 9 (8): е104551. Бибкод : 2014PLoSO...9j4551H . дои : 10.1371/journal.pone.0104551 . ПМК 4131922 . ПМИД 25118986 .
- ^ Цзи, Шу-ан; Чжан, Лицзюнь; Чжан, Шудун; Ханг, Шан (2014). «Крупноразмерный анкилозавр (динозавр) из нижнемеловой формации Цзюфотан в Западном Ляонине, Китай». Акта Геологика Синика . 88 (4): 1060–1065. Бибкод : 2014AcGlS..88.1060J . дои : 10.1111/1755-6724.12273 . S2CID 140573975 .
- ^ Jump up to: а б Ли, Х.; Рейс, Р.Р. (10–13 мая 2019 г.). Раннемеловой анкилозавр Liaoningosaurus из Западного Ляонина, Китай; Прогресс и проблемы . 7-е ежегодное собрание Канадского общества палеонтологии позвоночных. стр. 31–32.
{{cite conference}}: CS1 maint: дата и год ( ссылка ) - ^ Арбур, Виктория М.; Карри, Филип Дж. (01 октября 2015 г.). «Хвостовые булавы динозавров-анкилозаврид развились путем постепенного приобретения ключевых особенностей» . Журнал анатомии . 227 (4): 514–523. дои : 10.1111/joa.12363 . ISSN 1469-7580 . ПМК 4580109 . ПМИД 26332595 .
- ^ Jump up to: а б Арбур, ВМ; Карри, Пи Джей (2015). «Систематика, филогения и палеобиогеография динозавров-анкилозаврид». Журнал систематической палеонтологии . 14 (5): 1–60. Бибкод : 2016JSPal..14..385A . дои : 10.1080/14772019.2015.1059985 . S2CID 214625754 .
- ^ Арбур, ВМ; Занно, Ле; Гейтс, Т. (2016). «Палеоэкологические ассоциации анкилозавровых динозавров находились под влиянием истребления, колебаний уровня моря и георассредоточения». Палеогеография, Палеоклиматология, Палеоэкология . 449 : 289–299. Бибкод : 2016PPP...449..289A . дои : 10.1016/j.palaeo.2016.02.033 .
- ^ Арбур, Виктория М.; Эванс, Дэвид К. (2017). «Новый динозавр-анкилозавр из формации Джудит-Ривер в Монтане, США, основанный на исключительном скелете с сохранением мягких тканей» . Королевское общество открытой науки . 4 (5): 161086. Бибкод : 2017RSOS....461086A . дои : 10.1098/rsos.161086 . ПМК 5451805 . ПМИД 28573004 .
- ^ Вэньцзе Чжэн; Синшэн Цзинь; Ёичи Адзума; Цюнъин Ван; Казунори Мията; Син Сюй (2018). «Самый базальный анкилозавровый динозавр из альба-сеномана Китая, имеющий значение для эволюции хвостовой булавы» . Научные отчеты . 8 (1): Артикул 3711. Бибкод : 2018НатСР...8.3711Z . дои : 10.1038/s41598-018-21924-7 . ПМЦ 5829254 . ПМИД 29487376 .
- ^ Пак, JY; Ли, Ю.Н.; Карри, Пи Джей; Кобаяши, Ю.; Коппельхус, Э.; Барсболд, Р.; Матеус, О.; Ли, С.; Ким, SH (2019). «Дополнительные черепа Talarurus plicatospineus (Dinosauria: Ankylosauridae) и значение для палеобиогеографии и палеоэкологии панцирных динозавров». Меловые исследования . 108 : 104340. Бибкод : 2020CrRes.10804340P . дои : 10.1016/j.cretres.2019.104340 . S2CID 212423361 .
- ^ Г. Фрауэнфельдер, Тимоти; Р. Белл, Фил; Бруэм, Том; Дж. Бевитт, Джозеф; Округ Колумбия Бикнелл, Рассел; П. Кир, Бенджамин; Горе, Стивен; Э. Кампионе, Николас (2022). «Новые черепные остатки анкилозавров из формации Тулебук нижнего мела (верхний альб) в Квинсленде, Австралия» . Границы в науках о Земле . 10 :1–17. дои : 10.3389/feart.2022.803505 .
- ^ Ю, Чжицян; Ван, Мин; Ли, Юцзюань; Дэн, Чэнлун; Хэ, Хуайю (01 декабря 2021 г.). «Новые геохронологические ограничения для нижнемеловой формации Цзюфотанг в бассейне Цзяньчан, северо-восток Китая, и их последствия для биоты позднего Джехол» . Палеогеография, Палеоклиматология, Палеоэкология . 583 : 110657. Бибкод : 2021PPP...583k0657Y . дои : 10.1016/j.palaeo.2021.110657 . ISSN 0031-0182 . S2CID 239406222 .
- ^ Чжоу, Чжунхэ; М. Барретт, Пол; Хилтон, Джейсон (2003). «Исключительно сохранившаяся экосистема нижнего мела». Природа . 421 (6925): 807–814. Бибкод : 2003Natur.421..807Z . дои : 10.1038/nature01420 . ПМИД 12594504 . S2CID 4412725 .
- ^ Ван, М.; О'Коннор, Дж. К.; Чжоу, С.; Чжоу, З. (2019). «Новая зубастая птица-орнитуроморф раннего мела обнаруживает внутрикладовое разнообразие в характере потери зубов». Журнал систематической палеонтологии . 18 (8): 631–645. дои : 10.1080/14772019.2019.1682696 . S2CID 209575088 .
- ^ Чжоу, С.; Чжоу, З.; О'Коннор, Дж. (2013). «Новый рыбоядный орнитуроморф из биоты Джехол». Историческая биология . 26 (5): 608–618. дои : 10.1080/08912963.2013.819504 . S2CID 67854494 .
- ^ Ли, Ли; Ван, Цзин-Ци; Хоу, Ши-Лин (2011). «Новая птица-орнитурин (Hongshanornithidae) из формации Цзюфотан в Чаояне, Ляонин, Китай» (PDF) . Позвоночные Палазиатские . 49 (2): 195–200.
- ^ Чжоу, З.; Кларк, Дж.А.; Чжан, Ф. (2002). « археораптора » Лучшая половина . Природа . 420 (6913): 285. Бибкод : 2002Natur.420..285Z . дои : 10.1038/420285a . ПМИД 12447431 . S2CID 4423242 .
- ^ Чжоу З. и Ван Ю. (2010). «Разнообразие позвоночных биоты Джехол по сравнению с другими лагерштеттенами». Наука Китая: Науки о Земле , 53 (12): 1894–1907. doi : 10.1007/s11430-010-4094-9 [1] Архивировано 29 октября 2013 г. на Wayback Machine.
- ^ Чжан, Фучэн; Чжоу, Чжунхэ; Хоу, Ляньхай; Гу, Банда (июнь 2001 г.). «Ранняя диверсификация птиц: данные о новой противоположной птице». Китайский научный бюллетень . 46 (11): 945–949. Бибкод : 2001ЧСБу..46..945З . дои : 10.1007/bf02900473 . S2CID 85215328 .
- ^ Моршхаузер, Э.М.; Варриккио, диджей; Гао С.; Лю Дж.; Ван Х.; Ченг С. и Мэн Ц. (2009). «Анатомия раннемеловой птицы Rapaxavis pani , нового вида из провинции Ляонин, Китай». Журнал палеонтологии позвоночных . 29 (2): 545–554. Бибкод : 2009JVPal..29..545M . дои : 10.1671/039.029.0210 . S2CID 84643293 .
- ^ Серено, ПК, и Рао, К. (1992). «Ранняя эволюция птичьего полета и сидения: новые данные из нижнего мела Китая». Наука , 255(5046), 845.
- ^ Ван М., О'Коннор Дж. К., Чжао Т., Пан Ю, Чжэн Х, Ван Х, Чжоу Z (16 сентября 2021 г.). «Раннемеловая энантиорнитиновая птица с шилохвостом» . Современная биология . 31 (21): 4845–4852.e2. Бибкод : 2021CBio...31E4845W . дои : 10.1016/j.cub.2021.08.044 . ISSN 0960-9822 . ПМИД 34534442 .
- ^ Jump up to: а б Ван Сяо-Линь; Чжоу Чжун-Хе (2003). «Два новых птеродактилоидных птерозавра из раннемеловой формации Цзюфотан в Западном Ляонине, Китай». Позвоночные Палазиатские . 41 (1): 34–41.
- ^ Лю, Цзюньчан; Цян Цзи (2005). «Новый птерозавр аждархид из раннего мела западного Ляонина». Акта Геологика Синика . 79 (3): 301–307. Бибкод : 2005AcGlS..79..301L . дои : 10.1111/j.1755-6724.2005.tb00893.x . S2CID 128958556 .
- ^ Донг, З.; Сан, Ю.; Ву, С. (2003). «О новом птерозавре из нижнего мела бассейна Чаоян, Западный Ляонин, Китай». Глобальная геология . 22 (1): 1–7.
- ^ Люй Дж.; Д.М. Анвин; Сюй Л.; Чжан С. (2008). «Новый аждархоидный птерозавр из нижнего мела Китая и его значение для филогении и эволюции птерозавров». Naturwissenschaften . 95 (9): 891–7. Бибкод : 2008NW.....95..891L . дои : 10.1007/s00114-008-0397-5 . ПМИД 18509616 . S2CID 13458087 .
- ^ Цзян, С.; Ченг, X.; Может.; Ван, X. (2016). «Новый археоптеродактилоидный птерозавр из формации Цзюфотанг на западе Ляонина, Китай, со сравнением грудины у Pterodactylomorpha». Журнал палеонтологии позвоночных . 36 (6): e1212058. Бибкод : 2016JVPal..36E2058J . дои : 10.1080/02724634.2016.1212058 . S2CID 89481172 .
- ^ Сяолинь Ван; Александр В.А. Келлнер; Шуньсин Цзян; Синь Ченг (2012). «Новая зубастая летающая рептилия из Азии: близкое сходство между фауной птерозавров раннего мела из Китая и Бразилии». Naturwissenschaften . 99 (4): 249–57. Бибкод : 2012NW.....99..249W . дои : 10.1007/s00114-012-0889-1 . ПМИД 22354475 . S2CID 7323552 .
- ^ Сяолинь Ван; Таисса Родригес; Шуньсин Цзян; Синь Ченг; Александр В.А. Келлнер (2014). «Раннемеловой птерозавр с необычным нижнечелюстным гребнем из Китая и потенциально новой стратегией питания» . Научные отчеты . 4 : Артикул 6329. doi : 10.1038/srep06329 . ПМЦ 5385874 . ПМИД 25210867 .
- ^ Сяолинь Ван; Диоген де Алмейда Кампос; Чжунхэ Чжоу; Александр В.А. Келлнер (2008). «Примитивный истиодактилидный птерозавр (Pterodactyloidea) из формации Цзюфотанг (ранний мел), северо-восточный Китай». Зоотакса . 1813 : 1–18. дои : 10.11646/zootaxa.1813.1.1 .
- ^ Донг З. и Люй Дж. (2005). Новый ктенохазматидный птерозавр из раннего мела провинции Ляонин. Acta Geologica Sinica 79 (2): 164-167.
- ^ Родригес, Т.; Цзян, С.; Ченг, X.; Ван, X.; Келлнер, AWA (2015). «Новый зубатый птеранодонтоид (Pterosauria, Pterodactyloidea) из формации Цзюфотанг (нижний мел, апт) в Китае и комментарии к Liaoningopterus Гуй Ван и Чжоу, 2003». Историческая биология . 27 (6): 782–795. Бибкод : 2015HBio...27..782R . дои : 10.1080/08912963.2015.1033417 . S2CID 129062416 .
- ^ Чжоу X., Пегас Р.В., Леал МЭК и Бонд Н. 2019. « Nurhachius luei , новый истиодактилидный птерозавр (Pterosauria, Pterodactyloidea) из раннемеловой формации Цзюфотан города Чаоян, провинция Ляонин (Китай) и комментарии к Istiodactylidae. " ПирJ 7 :e7688
- ^ Сяолинь Ван, Келлнер, AWA, Чжоу Чжунхэ и де Алмейда Кампос, Д. (2005). Разнообразие птерозавров и круговорот фауны в меловых наземных экосистемах Китая. Природа 437 :875–879.
- ^ Ван, Сяолинь; Чжоу, Чжунхэ (июнь 2003 г.). «Новый птерозавр (Pterodactyloidea, Tapejaridae) из раннемеловой формации Цзюфотанг на западе Ляонина, Китай, и его значение для биостратиграфии». Китайский научный бюллетень . 48 (1): 16–23. Бибкод : 2003ЧСБу..48...16Вт . дои : 10.1007/bf03183326 . S2CID 43993745 .
- ^ Лю, Дж.; Мэн, К.; Ван, Б.; Лю, Д.; Шен, К.; Чжан, Ю. (2017). «Краткая заметка о новом анурогнатидном птерозавре со свидетельствами его поведения на насесте из Цзяньчана, провинция Ляонин, Китай» (PDF) . Ин Хон, DWE; Уиттон, член парламента; Мартилл, Д.М. (ред.). Новые взгляды на палеобиологию птерозавров . Геологическое общество, Лондон, Специальные публикации. Том. 455. Лондон: Лондонское геологическое общество. стр. 95–104. дои : 10.1144/SP455.16 . S2CID 219196969 .
- ^ Лу, Дж.; Лю, К.; Пан, Л.; Шен, К. (2016). «Новый птеродактилоидный птерозавр из раннего мела западной части провинции Ляонин, северо-восточный Китай». Акта Геологика Синика . 90 (3): 777–782. Бибкод : 2016AcGlS..90..777L . дои : 10.1111/1755-6724.12721 . S2CID 132555691 .
- ^ О'Коннор, Дж. К.; Сан, К.; Сюй, Х.; Ван, X.; Чжоу, З. (2012). «Новый вид Jeholornis с полным хвостовым покровом». Историческая биология . 24 (1): 29–41. Бибкод : 2012HBio...24...29O . дои : 10.1080/08912963.2011.552720 . S2CID 53359901 .
- ^ Сюри Ван; Цзяндун Хуан; Мартин Кундрат; Андреа Кау; Сяоюй Лю; Ян Ван; Шубин Джу (2020). «Новый jeholornithiform демонстрирует самое раннее появление сросшихся грудины и таза в эволюции птичьих динозавров». Журнал азиатских наук о Земле . 199 : Артикул 104401. Бибкод : 2020JAESc.19904401W . doi : 10.1016/j.jseaes.2020.104401 . S2CID 219511931 .
- ^ Черкас, С.А. и Джи, К. (2002). «Предварительный отчет о всеядной летучей птице из северо-восточного Китая». В : Черкас, С.Дж. (редактор): Пернатые динозавры и происхождение полета. Журнал Музея динозавров 1 : 127–135. HTML-абстракт
- ^ Чжоу, Чжунхэ и Чжан, Фучэн (2003): Анатомия примитивной птицы Sapeornis chaoyangensis из раннего мела Ляонин, Китай. Канадский журнал наук о Земле 40 (5): 731–747. doi : 10.1139/E03-011 (аннотация в формате HTML)
- ^ Сюй, Х.; Норелл, Массачусетс (2006). «Окаменелости нептичьих динозавров из группы Джехоль нижнего мела на западе Ляонина, Китай». Геологический журнал . 41 (3–4): 419–437. Бибкод : 2006GeolJ..41..419X . дои : 10.1002/gj.1044 . S2CID 32369205 .
- ^ Джи, К.; Джи, С.-А.; Чжан, Л.-Ж. (2009). «Первый крупный теропод-тираннозавроид из раннемеловой биоты Джехоль на северо-востоке Китая». Геологический вестник Китая . 28 (10): 1369–1374.
- ^ Мао, Ф.; Чжан, К.; Лю, К.; Мэн, Дж. (2021). «Фоссориальность и эволюционное развитие двух млекопитающих мелового периода». Природа . 592 (7855): 577–582. Бибкод : 2021Natur.592..577M . дои : 10.1038/s41586-021-03433-2 . ПМИД 33828300 . S2CID 233183060 .
- ^ Мэн, Дж.; Ван, Ю.; Ли, К. (2011). «Переходное среднее ухо млекопитающих от нового эвтриконодонта Джехол мелового периода». Природа . 472 (7342): 181–185. Бибкод : 2011Natur.472..181M . дои : 10.1038/nature09921 . ПМИД 21490668 . S2CID 4428972 .
- ^ Гао, К.-К.; Фокс, RC (2005). «Новая хористодера (Reptilia: Diapsida) из нижнего мела западной провинции Ляонин, Китай, и филогенетические связи Monjurouchidae» . Зоологический журнал Линнеевского общества . 145 (3): 427–444. дои : 10.1111/j.1096-3642.2005.00191.x .
- ^ Новый вид икехозавра (Reptilia: Choristodera) из формации Цзюфутанг (ранний мел) города Чифэн, Внутренняя Монголия. Архивировано 4 сентября 2012 г. в Wayback Machine LÜ J.-C.; Кобаяши Ю.; Ли З.-Г.
- ^ Гао Чуньлин; Лу Цзюньчан; Лю Цзиньюань; Цзи Цян (2005). «Новые хористодеры из нижнемеловой формации Цзюфотан в районе Чаоян, Ляонин, Китай». Геологическое обозрение (на китайском языке). 51 (6): 694–697.
