Динозавр
| Динозавры Временной диапазон: (возможная запись среднего триаса) | ||
|---|---|---|
 Herrerasaurus ischigualastensis (хищный базальный динозавр) | ||
| Научная классификация | ||
| Домен: | Эукариоты | |
| Королевство: | животное | |
| Тип: | Хордовые | |
| Клэйд : | Зауропсида | |
| Клэйд : | Архозаврия | |
| Клэйд : | Авеметатарсалия | |
| Клэйд : | Орнитодира | |
| Клэйд : | Динозавроморфа | |
| Клэйд : | Динозаврообразные | |
| Клэйд : | Дракохоры | |
| Клэйд : | Динозаврия Оуэн , 1842 г. | |
| Основные группы | ||
| ||






Динозавры — разнообразная группа рептилий. [примечание 1] из клады Dinosauria . Впервые они появились в триасовый период , между 243 и 233,23 миллионами лет назад (млн лет назад), хотя точное происхождение и сроки эволюции динозавров являются предметом активных исследований. Они стали доминирующими наземными позвоночными после триасово-юрского вымирания, произошедшего 201,3 млн лет назад, и их доминирование продолжалось на протяжении юрского и мелового периодов. Летопись окаменелостей показывает, что птицы — это пернатые динозавры , которые произошли от более ранних теропод в эпоху поздней юры и являются единственной известной линией динозавров, пережившей мел-палеогеновое вымирание примерно 66 млн лет назад. Таким образом, динозавров можно разделить на птичьих динозавров (птиц) и вымерших нептичьих динозавров , которые все являются динозаврами, кроме птиц.
Динозавры разнообразны с таксономической , морфологической и экологической точек зрения. Птицы, насчитывающие более 11 000 ныне живущих видов , относятся к числу самых разнообразных групп позвоночных. Используя ископаемые свидетельства, палеонтологи идентифицировали более 900 различных родов и более 1000 различных видов нептичьих динозавров. Динозавры представлены на каждом континенте как современными видами (птицами), так и ископаемыми остатками. В первой половине 20-го века, до того как птицы были признаны динозаврами, большая часть научного сообщества считала, что динозавры были медлительными и хладнокровными . Однако большинство исследований, проведенных с 1970-х годов , показали, что динозавры были активными животными с повышенным метаболизмом и многочисленными адаптациями к социальному взаимодействию. Некоторые были травоядными , другие плотоядными . Имеющиеся данные свидетельствуют о том, что все динозавры откладывали яйца и что строительство гнезд было чертой, присущей многим динозаврам, как птичьим, так и нептичьим.
Хотя динозавры изначально были двуногими , многие вымершие группы включали четвероногих видов, и некоторые из них могли переключаться между этими позициями. Сложные демонстрационные структуры, такие как рога или гребни, являются общими для всех групп динозавров, а у некоторых вымерших групп появились модификации скелета , такие как костная броня и шипы . В то время как современные сохранившиеся виды птиц (птиц) динозавров, как правило, невелики из-за ограничений полета, многие доисторические динозавры (нептичьи и птичьи) имели крупное тело - самые крупные динозавры- зауроподы , по оценкам, достигали длины 39,7 дюйма. метров (130 футов) и высотой 18 м (59 футов) и были самыми крупными наземными животными всех времен. Заблуждение о том, что нептичьи динозавры были одинаково гигантскими, отчасти основано на предвзятом отношении к сохранности , поскольку большие, крепкие кости с большей вероятностью сохранятся до тех пор, пока не окаменеют. Многие динозавры были довольно маленькими, некоторые имели длину около 50 сантиметров (20 дюймов).
Первые окаменелости динозавров были обнаружены в начале 19 века, а название «динозавр» (что означает «ужасная ящерица») было придумано сэром Ричардом Оуэном в 1842 году для обозначения этих «великих ископаемых ящериц». [7] [8] [9] С тех пор скелеты ископаемых динозавров стали главной достопримечательностью музеев по всему миру, а динозавры стали неотъемлемой частью массовой культуры . Крупные размеры некоторых динозавров, а также их, казалось бы, чудовищная и фантастическая природа обеспечили их регулярное появление в бестселлерах и фильмах, таких как франшиза «Парк Юрского периода ». Постоянный общественный энтузиазм по поводу животных привел к значительному финансированию науки о динозаврах, а новые открытия регулярно освещаются в средствах массовой информации.
Определение
Согласно филогенетической номенклатуре , динозавры обычно определяются как группа, состоящая из самого недавнего общего предка (MRCA) трицератопсов и современных птиц (неорнитов) и всех его потомков. [10] Также было предложено определить Dinosauria относительно MRCA Megalosaurus и Iguanodon , поскольку это были два из трех родов, упомянутых Ричардом Оуэном, когда он признал Dinosauria. [11] Оба определения охватывают одни и те же известные роды: Dinosauria = Ornithischia + Saurischia . Сюда входят такие основные группы, как анкилозавры (бронированные травоядные четвероногие), стегозавры (пластинчатые травоядные четвероногие), цератопсы (двуногие или четвероногие травоядные с оборками на шее ), пахицефалозавры (двуногие травоядные с толстым черепом), орнитоподы (двуногие или четвероногие травоядные, включая утиных ). клювы »), тероподы (в основном двуногие хищники и птицы) и зауроподоморфы (в основном крупные травоядные четвероногие с длинной шеей и хвостом). [12]
Птицы — единственные выжившие динозавры. В традиционной систематике птицы считались отдельным классом , произошедшим от динозавров, отдельным надотрядом . Однако большинство современных палеонтологов отвергают традиционный стиль классификации, основанный на анатомическом сходстве, в пользу филогенетической таксономии, основанной на установленном происхождении, в которой каждая группа определяется как все потомки данного рода-основателя. [13] Птицы относятся к подгруппе динозавров Maniraptora , которые являются целурозаврами , которые являются тероподами, которые являются ящеротазами. [14]
Исследования Мэтью Дж. Барона, Дэвида Б. Нормана и Пола М. Барретта в 2017 году предложили радикальный пересмотр систематики динозавров. Филогенетический анализ, проведенный Baron et al. обнаружил, что Ornithischia ближе к тероподам, чем к зауроподоморфам, в отличие от традиционного союза теропод с зауроподоморфами. Это привело бы к тому, что зауроподы и их родственники выпали бы за пределы традиционных динозавров, поэтому они переопределили Dinosauria как последнего общего предка Triceratops horridus , Passer Domesticus и Diplodocus carnegii , а также всех их потомков, чтобы гарантировать, что зауроподы и их родственники останутся включенными в число динозавров. Они также возродили кладу Ornithoscelida, чтобы обозначить группу, содержащую Ornithischia и Theropoda. [15] [16]
Общее описание

Используя одно из приведенных выше определений, динозавров можно в целом описать как архозавров , задние конечности которых находятся прямо под телом . [17] Другие доисторические животные, в том числе птерозавры , мозазавры , ихтиозавры , плезиозавры и диметродоны , хотя их часто принято считать динозаврами, таксономически не классифицируются как динозавры. [18] Птерозавры имеют отдаленное родство с динозаврами, являясь представителями клады Орнитодиры . Другие упомянутые группы, подобно динозаврам и птерозаврам, являются членами Sauropsida (клада рептилий и птиц), за исключением Dimetrodon (который является синапсидом ). Ни у одного из них не было вертикального положения задних конечностей, характерного для настоящих динозавров. [19]
Динозавры были доминирующими наземными позвоночными мезозойской эры , особенно юрского и мелового периодов. Другие группы животных были ограничены в размерах и нишах; млекопитающие , например, редко превышали размер домашней кошки и обычно были плотоядными животными размером с грызуна и мелкой добычей. [20] Динозавров всегда признавали чрезвычайно разнообразной группой: уверенно идентифицировано более 900 нептичьих родов динозавров (2018 г.) и 1124 вида (2016 г.). По оценкам, общее количество родов динозавров, сохранившихся в летописи окаменелостей, относится к 1850 году, почти 75% из них до сих пор не обнаружены. [21] [22] [23] и число, которое когда-либо существовало (в летописи окаменелостей или за ее пределами), составляло 3400. [24] По оценкам 2016 года, количество видов динозавров, живших в мезозое, составляло 1543–2468 человек. [25] [26] по сравнению с численностью современных птиц (птичьих динозавров) на уровне 10 806 видов. [27]
Вымершие динозавры, а также современные птицы включают роды травоядных и плотоядных, в том числе семеноядных, рыбоядных, насекомоядных и всеядных. Хотя динозавры изначально были двуногими (как и все современные птицы), некоторые из них превратились в четвероногих, а другие, такие как анхизавр и игуанодон , могли так же легко ходить на двух или четырех ногах. Черепные модификации, такие как рога и гребни, являются обычными чертами динозавров, а у некоторых вымерших видов была костная броня. Хотя самые известные виды отличаются большими размерами, многие мезозойские динозавры были размером с человека или меньше, а современные птицы, как правило, имеют небольшие размеры. Динозавры сегодня населяют все континенты, и окаменелости показывают, что они достигли глобального распространения самое позднее в раннеюрскую эпоху. [28] Современные птицы населяют большинство доступных местообитаний, от наземных до морских, и есть свидетельства того, что некоторые нептичьи динозавры (такие как Микрораптор ) могли летать или хотя бы планировать, а другие, такие как спинозавриды , имели полуводный образ жизни. [29]
Отличительные анатомические особенности
Хотя недавние открытия затруднили представление общепризнанного списка их отличительных особенностей, почти все обнаруженные к настоящему времени динозавры имеют определенные модификации скелета предков архозавров или явно являются потомками более старых динозавров, демонстрирующих эти модификации. Хотя некоторые более поздние группы динозавров имели дальнейшие модифицированные версии этих черт, они считаются типичными для Dinosauria; они были у самых ранних динозавров и передали их своим потомкам. Такие модификации, происходящие от самого недавнего общего предка определенной таксономической группы, называются синапоморфиями такой группы. [30]

Подробная оценка взаимоотношений архозавров, проведенная Стерлингом Несбиттом. [31] подтвердили или обнаружили следующие двенадцать однозначных синапоморфий, некоторые из которых известны ранее:
- В черепе перед надвисочным окном имеется надвисочная ямка (выемка) , главное отверстие в задней части крыши черепа.
- Эпипофизы , косо направленные назад отростки на задних верхних углах передних (передних) шейных позвонков позади атланта и оси , первых двух шейных позвонков.
- Вершина дельтопекторального гребня (выступ, к которому прикрепляются дельтопекторальные мышцы), расположенная на 30% или более длины плечевой кости (плечевая кость)
- Лучевая кость , кость предплечья, короче 80% длины плечевой кости.
- Четвертый вертел (выступ, где хвостатобедренная мышца прикрепляется к внутренней задней диафизу) на бедренной кости (бедренной кости) представляет собой острый фланец.
- Четвертый вертел асимметричный, с дистальным нижним краем, образующим более крутой угол с диафизом.
- На таранной и пяточной костях , верхней части лодыжки, проксимальной суставной фасетке, верхней соединительной поверхности, для малоберцовой кости занимает менее 30% поперечной ширины элемента.
- Экзоципитали (кости задней части черепа) не встречаются по средней линии на дне эндокраниальной полости, внутреннего пространства черепной коробки.
- В тазу проксимальные суставные поверхности седалищной кости с подвздошной и лобковой костью разделены большой вогнутой поверхностью (на верхней стороне седалищной кости между контактами с лобковой костью и лобковой костью располагается часть открытого тазобедренного сустава). подвздошная кость)
- Кнемиальный гребень на большеберцовой кости (выступающая часть верхней поверхности большеберцовой кости) дугой передне-латерально (загибается к передней и наружной стороне)
- Отчетливый проксимодистально ориентированный (вертикальный) гребень присутствует на задней поверхности дистального конца большеберцовой кости (задняя поверхность нижнего конца большеберцовой кости).
- Вогнутая суставная поверхность малоберцовой кости пяточной кости (верхняя поверхность пяточной кости в месте соприкосновения с малоберцовой костью имеет полый профиль)
Несбитт обнаружил ряд дополнительных потенциальных синапоморфий и исключил ряд ранее предложенных синапоморфий. Некоторые из них также присутствуют у силезавридов , которых Несбитт обнаружил как родственную группу Dinosauria, включая большой передний вертел, II и IV плюсневые кости примерно одинаковой длины, уменьшенный контакт между седалищной и лобковой костью, наличие кнемиального гребня на большеберцовой кости и восходящего отростка на таранной кости и многие другие. [10]

Динозавры имеют и ряд других особенностей скелета. Однако, поскольку они либо являются общими для других групп архозавров, либо не присутствовали у всех ранних динозавров, эти особенности не считаются синапоморфиями. Например, как диапсиды , динозавры изначально имели две пары подвисочных окон (отверстий в черепе за глазами), а как представители диапсидной группы Archosauria, имели дополнительные отверстия в морде и нижней челюсти. [32] Кроме того, теперь известно, что некоторые характеристики, которые когда-то считались синапоморфиями, появились до динозавров или отсутствовали у самых ранних динозавров и независимо развивались различными группами динозавров. К ним относятся удлиненная лопатка , или лопатка; крестец , состоящий из трех и более сросшихся позвонков (у некоторых других архозавров встречаются три, а у Herrerasaurus — только два ); [10] и перфорированная вертлужная впадина , или тазобедренная впадина, с отверстием в центре ее внутренней поверхности (закрытой у Saturnalia tupiniquim ). , например, [33] [34] Еще одна трудность определения отчетливых особенностей динозавров заключается в том, что ранние динозавры и другие архозавры эпохи позднего триаса часто плохо известны и во многом были похожи; эти животные иногда ошибочно идентифицировались в литературе. [35]
Динозавры стоят с выпрямленными задними конечностями, подобно большинству современных млекопитающих , но отличаются от большинства других рептилий, чьи конечности раскинуты в обе стороны. [36] Эта поза обусловлена развитием обращенного латерально углубления в тазу (обычно открытой лунки) и соответствующей обращенной внутрь отчетливой головки на бедренной кости. [37] Их прямая поза позволяла ранним динозаврам легко дышать во время движения, что, вероятно, обеспечивало выносливость и уровень активности, превосходивший уровень «расползающихся» рептилий . [38] Прямые конечности, вероятно, также способствовали эволюции больших размеров за счет уменьшения изгибающих напряжений на конечностях. [39] Некоторые нединозавровые архозавры, в том числе райзухии , также имели прямые конечности, но достигали этого за счет «вертикальной» конфигурации тазобедренного сустава, при которой вместо выступа бедренной кости в гнезде на бедре верхняя тазовая кость был повернут, образуя нависающую полку. [39]
История обучения
Донаучная история
Окаменелости динозавров были известны на протяжении тысячелетий, хотя их истинная природа не была признана. Китайцы считали их костями дракона и документировали их как таковые. Например, Хуаянг Го Чжи ( 華陽國志 ), географический справочник, составленный Чан Цюем ( 常璩 ) во времена династии Западная Цзинь (265–316), сообщил об открытии костей дракона в Учэне в провинции Сычуань . [40] Жители деревень в центральном Китае уже давно раскопали окаменелые «кости дракона» для использования в традиционной медицине . [41] В Европе окаменелости динозавров обычно считали останками гигантов и других библейских существ. [42]
Ранние исследования динозавров

Научные описания того, что сейчас называют костями динозавров, впервые появились в конце 17 века в Англии. Часть кости, которая, как теперь известно, была бедренной костью мегалозавра . [43] был извлечен из известнякового карьера в Корнуэлле недалеко от Чиппинг-Нортона , Оксфордшир, в 1676 году. Фрагмент был отправлен Роберту Плоту , профессору химии Оксфордского университета и первому хранителю Эшмоловского музея , который опубликовал описание в своей книге «Естественная история». Оксфордшира (1677 г.). [44] Он правильно идентифицировал кость как нижнюю конечность бедренной кости крупного животного и признал, что она слишком велика, чтобы принадлежать какому-либо известному виду. Поэтому он пришел к выводу, что это бедренная кость огромного человека, возможно, Титана или другого типа гиганта, фигурирующего в легендах. [45] [46] Эдвард Люйд , друг сэра Исаака Ньютона , опубликовал Lithophylacii Britannici ichnographia (1699), первое научное исследование того, что теперь будет признано динозавром, когда он описал и назвал зуб зауропода « Rutellum imicatum ». [47] [48] который был найден в Касвелле, недалеко от Уитни , Оксфордшир. [49]

Между 1815 и 1824 годами преподобный Уильям Бакленд , первый преподаватель геологии в Оксфордском университете, собрал еще больше окаменевших костей мегалозавра и стал первым человеком, описавшим нептичьего динозавра в научном журнале . [43] [50] Второй идентифицированный род нептичьих динозавров, Iguanodon , согласно легенде, был открыт в 1822 году Мэри Энн Мантелл – женой английского геолога Гидеона Мантелла, которому на самом деле потребовались останки несколькими годами ранее. Гидеон Мантелл признал сходство между своими окаменелостями и костями современных игуан . Он опубликовал свои открытия в 1825 году. [51] [52]
Изучение этих «огромных ископаемых ящериц» вскоре вызвало большой интерес у европейских и американских учёных, а в 1842 году английский палеонтолог сэр Ричард Оуэн ввёл термин «динозавр», используя его для обозначения «отдельного племени или подотряда динозавров». Ящероподобные рептилии», которые тогда были признаны в Англии и во всем мире. [7] [8] [9] [53] [54] Этот термин происходит от древнегреческого δεινός (deinos) «ужасный, могущественный или ужасно великий» и σαῦρος (sauros) «ящерица или рептилия». [53] [55] Хотя таксономическое название часто интерпретировалось как отсылка к зубам, когтям и другим устрашающим характеристикам динозавров, Оуэн хотел, чтобы оно также напоминало об их размерах и величии. [56] Оуэн признал, что останки игуанодона , мегалозавра и гилеозавра , которые были найдены до сих пор, имеют общие отличительные черты, и поэтому решил представить их как отдельную таксономическую группу. Как пояснил британский геолог и историк Хью Торренс, Оуэн представил презентацию об ископаемых рептилиях Британской ассоциации содействия развитию науки в 1841 году, но отчеты того времени показывают, что Оуэн не упоминал слово «динозавр» и не признавал динозавров. как отдельную группу рептилий в его адрес. Он представил динозавров только в исправленной текстовой версии своего выступления, опубликованной в апреле 1842 года. [7] [8] При поддержке принца Альберта , мужа королевы Виктории , Оуэн основал Музей естественной истории в Лондоне для демонстрации национальной коллекции окаменелостей динозавров и других биологических и геологических экспонатов. [57]
Открытия в Северной Америке


В 1858 году Уильям Паркер Фоулк обнаружил первого известного американского динозавра в мергелевых карьерах в небольшом городке Хэддонфилд, штат Нью-Джерси . (Хотя окаменелости находили и раньше, их природа не была правильно определена.) Существо назвали Hadrosaurus foulkii . Это была чрезвычайно важная находка: Гадрозавр был одним из первых почти полных скелетов динозавров, найденных ( первый был обнаружен в 1834 году в Мейдстоне, Англия ), и он явно был двуногим существом. Это было революционное открытие, поскольку до этого момента большинство учёных считали, что динозавры, как и другие ящерицы, ходили на четырёх ногах. Открытия Фоулка вызвали волну интереса к динозаврам в Соединенных Штатах, известную как динозавромания. [58]
Мания динозавров была иллюстрирована ожесточенным соперничеством между Эдвардом Дринкером Коупом и Отниэлем Чарльзом Маршем , оба из которых стремились первыми найти новых динозавров в так называемых Войнах костей . Эта борьба между двумя учеными длилась более 30 лет и закончилась в 1897 году, когда Коуп умер, потратив все свое состояние на охоту на динозавров. Многие ценные экземпляры динозавров были повреждены или уничтожены из-за грубых методов этой пары: например, их землекопы часто использовали динамит , чтобы раскопать кости. Современные палеонтологи сочли бы такие методы грубыми и неприемлемыми, поскольку взрывные работы легко уничтожают ископаемые и стратиграфические свидетельства. Несмотря на несовершенство методов, вклад Коупа и Марша в палеонтологию был огромен: Марш раскопал 86 новых видов динозавров, а Коуп открыл 56, всего 142 новых вида. Коллекция Коупа сейчас находится в Американском музее естественной истории в Нью-Йорке, а коллекция Марша — в Музее естественной истории Пибоди в Йельском университете. . [59]
«Ренессанс динозавров» и не только

Вторая мировая война вызвала паузу в палеонтологических исследованиях; после войны внимание исследователей также все больше переключалось на ископаемых млекопитающих, а не на динозавров, которые считались медлительными и хладнокровными. [60] [61] Однако в конце 1960-х годов в области исследований динозавров произошел всплеск активности, который продолжается до сих пор. [62] К этой деятельности привели несколько плодотворных исследований. Сначала Джон Остром обнаружил птицеподобного -дромеозаврида теропода Deinonychus и описал его в 1969 году. Его анатомия указывала на то, что это был активный хищник, который, вероятно, был теплокровным, что резко контрастировало с преобладавшим в то время представлением о динозаврах. [60] Одновременно Роберт Т. Баккер опубликовал серию исследований, в которых также приводились доводы в пользу активного образа жизни динозавров, основанные на анатомических и экологических данных (см. § Физиология ). [63] [64] которые впоследствии были обобщены в его книге «Ереси динозавров» 1986 года . [65]

Новые открытия были подкреплены увеличением количества открытий динозавров. Основные новые открытия динозавров были сделаны палеонтологами, работающими в ранее неисследованных регионах, включая Индию, Южную Америку, Мадагаскар, Антарктиду и, что особенно важно, Китай. Среди теропод, зауроподоморфов и птицетазовых количество названных родов начало расти в геометрической прогрессии в 1990-х годах. [21] По состоянию на 2008 год [update] Ежегодно давали названия более 30 новым видам динозавров. [66] По крайней мере, у зауроподоморфов в 2010-х годах наблюдалось дальнейшее увеличение количества названных видов: в период с 2009 по 2020 год ежегодно было названо в среднем 9,3 новых вида. Как следствие, в период с 1990 по 2020 год было названо больше зауроподоморфов, чем за все предыдущие годы. годы вместе взятые. [67] Эти новые местоположения также привели к улучшению общего качества образцов: новые виды все чаще назывались не по разрозненным окаменелостям, а по более полным скелетам, иногда принадлежащим нескольким особям. Улучшение образцов также привело к тому, что новые виды стали реже признаваться недействительными. [66] В азиатских местонахождениях были обнаружены наиболее полные экземпляры теропод. [68] в то время как в местонахождениях Северной Америки были обнаружены наиболее полные экземпляры зауроподоморфов. [67]
До эпохи Возрождения динозавров динозавры в основном классифицировались с использованием традиционной ранговой системы таксономии Линнея . Возрождение также сопровождалось все более широким применением кладистики , более объективного метода классификации, основанного на происхождении и общих чертах, который оказался чрезвычайно полезным при изучении систематики и эволюции динозавров. Кладистический анализ, среди других методов, помогает компенсировать часто неполные и фрагментарные летописи окаменелостей. [69] [70] Справочники, обобщающие состояние исследований динозавров, такие как « Дэвида Б. Вейшампеля и его коллег Динозаврия» , сделали знания более доступными. [71] и стимулировал дальнейший интерес к исследованиям динозавров. Выпуск первого и второго изданий «Динозаврии» в 1990 и 2004 годах, а также обзорной статьи Пола Серено в 1998 году сопровождался увеличением количества опубликованных филогенетических деревьев динозавров. [72]
Мягкие ткани и молекулярная консервация

Окаменелости динозавров не ограничиваются костями, но включают также отпечатки или минерализованные остатки кожных покровов, органов и других тканей. Из них кожные покровы на основе кератиновых легче всего сохраняются сшитой белков из-за их гидрофобной молекулярной структуры. [73] Окаменелости кожных покровов на основе кератина или костных кожных покровов известны у большинства основных групп динозавров. Окаменелости динозавров с чешуйчатыми отпечатками кожи находили с 19 века. Сэмюэл Беклс обнаружил в 1852 году переднюю конечность зауропода с сохранившейся кожей, которую ошибочно приписали крокодилу; он был правильно приписан Маршем в 1888 году и подвергнут дальнейшему изучению Реджинальдом Хули в 1917 году. [74] Среди птицетазовых в 1884 году Джейкоб Вортман обнаружил отпечатки кожи на первом известном экземпляре Edmontosaurus annectens , которые были в значительной степени уничтожены во время раскопок экземпляра. [75] Оуэн и Хули впоследствии описали отпечатки кожи гипсилофодона и игуанодона в 1885 и 1917 годах. [74] С тех пор отпечатки чешуи чаще всего находили среди гадрозаврид, причем отпечатки известны почти со всего тела у нескольких экземпляров. [76]


Начиная с 1990-х годов крупные открытия исключительно сохранившихся окаменелостей в отложениях, известных как консервационные Лагерштеттены, способствовали исследованиям мягких тканей динозавров. [77] [78] В основном среди них были породы, из которых образовались Джэхол (ранний мел) и Янляо (средний и поздний юрский период) биоты на северо-востоке Китая, из которых были обнаружены сотни экземпляров динозавров, несущих отпечатки перьевых структур (как тесно связанных с птицами, так и другими видами). см. § Происхождение птиц ) были описаны Син Сюй и его коллегами. [79] [80] клеточные структуры, хранящие пигмент, известные как меланосомы . У современных рептилий и млекопитающих за окраску частично отвечают [81] [82] Как химические следы меланина , так и меланосомы характерной формы были обнаружены в перьях и чешуе динозавров Джехол и Янляо, включая как теропод, так и птицетазовых. [83] Это позволило несколько раз полностью реконструировать окраску динозавров , например, Sinosauropteryx. [84] и Пситтакозавр [85] Якобом Винтером и его коллегами, и аналогичные методы были также распространены на окаменелости динозавров из других мест. [81] (Однако некоторые исследователи также предполагают, что окаменелые меланосомы представляют собой остатки бактерий. [86] [87] ) Содержимое желудка некоторых динозавров Джэхол и Янляо, тесно связанных с птицами, также дало косвенные указания на анатомию диеты и пищеварительной системы (например, сельскохозяйственные культуры ). [88] [89] Более конкретные свидетельства внутренней анатомии были обнаружены у Сципионикса из Пьетрароя Платтенкалк в Италии. Он сохраняет части кишечника, толстой кишки, печени, мышц и трахеи. [90]

Одновременно с этим направление работы под руководством Мэри Хигби Швейцер , Джека Хорнера и коллег сообщило о различных случаях сохранения мягких тканей и белков в окаменелостях костей динозавров. Различные минерализованные структуры, которые, вероятно, представляли собой эритроциты и коллагеновые волокна, были обнаружены Швейцером и другими в костях тираннозаврид еще в 1991 году. [91] [92] [93] Однако в 2005 году Швейцер и его коллеги сообщили, что в бедренной кости тираннозавра сохранились мягкие, гибкие ткани внутри, включая кровеносные сосуды , костный матрикс и соединительную ткань (костные волокна), которые сохранили свою микроскопическую структуру. [94] Это открытие позволило предположить, что исходные мягкие ткани могут сохраняться в течение геологического времени. [73] при этом было предложено несколько механизмов. [95] Позже, в 2009 году, Швейцер и его коллеги сообщили, что бедренная кость брахилофозавра сохранила сходные микроструктуры, а иммуногистохимические методы (основанные на связывании антител ) продемонстрировали присутствие таких белков, как коллаген, эластин и ламинин . [96] В обоих образцах были обнаружены последовательности белков коллагена, пригодные для молекулярно-филогенетического анализа , который, как и следовало ожидать, сгруппировал их с птицами. [96] [97] Сообщалось также об экстракции фрагментарной ДНК для обеих этих окаменелостей. [98] вместе с экземпляром Гипакрозавра . [99] В 2015 году Серджио Бертаццо и его коллеги сообщили о сохранении коллагеновых волокон и эритроцитов у восьми экземпляров динозавров мелового периода, которые не показали никаких признаков исключительной сохранности, что указывает на то, что мягкие ткани могут сохраняться чаще, чем считалось ранее. [100] Предположения, что эти структуры представляют собой бактериальные биопленки. [101] были отклонены, [102] однако остается возможность перекрестного заражения, которую трудно обнаружить. [103]
Эволюционная история
Происхождение и ранняя эволюция

Динозавры отделились от своих предков-архозавров в эпоху среднего и позднего триаса, примерно через 20 миллионов лет после разрушительного пермско-триасового вымирания, уничтожившего примерно 96% всех морских видов и 70% видов наземных позвоночных примерно 252 миллиона лет назад. [104] [105] Самые старые окаменелости динозавров, известные по существенным останкам, относятся к карнийской эпохе триасового периода и были обнаружены в основном в формациях Ишигуаласто и Санта-Мария в Аргентине и Бразилии, а также в формации Галечная Аркоза в Зимбабве. [106]
Формация Ишигуаласто ( возраст радиометрически 231-230 миллионов лет). [107] ) произвел раннего ящеротазового Eoraptor , первоначально считавшегося членом Herrerasauridae. [108] но теперь считается ранним зауроподоморфом, наряду с герреразавридами Herrerasaurus и Sanjuansaurus , а также зауроподоморфами Chromogisaurus , Eodromaeus и Panphagia . [109] с Вероятное сходство эораптора общим предком всех динозавров позволяет предположить, что первые динозавры были маленькими двуногими хищниками . [110] [111] [112] Формация Санта-Мария (радиометрически датирована более древней, возрастом 233,23 миллиона лет). [113] ) произвел герреразавридов Gnathovorax и Staurikosaurus , а также зауроподоморфов Bagualosaurus , Buriolestes , Guaibasaurus , Macrocollum , Nhandumirim , Pampadromaeus , Saturnalia и Unaysaurus . [109] В формации Пеббли-Аркос, возраст которой неизвестен, но, вероятно, был сопоставим с двумя другими, образовалась зауроподоморфа Mbiresaurus , а также безымянный герреразаврид. [106]
Менее хорошо сохранившиеся останки зауроподоморфов Jaklapallisaurus и Nambalia , а также раннего ящеротазового Alwalkeria известны из формаций Верхнего Малери и Нижнего Малери Индии. [114] карнийского возраста В формации Чаньярес в Аргентине сохранились примитивные, похожие на динозавров орнитодиры, такие как Lagosurus и Lagerpeton в Аргентине , что делает ее еще одним важным местом для понимания эволюции динозавров. Эти орнитодиры поддерживают модель ранних динозавров как маленьких двуногих хищников. [109] [115] Динозавры, возможно, появились еще в анизийскую эпоху триаса, примерно 243 миллиона лет назад, то есть в эпоху Ньясазавра из формации Манда в Танзании. Однако его известные окаменелости слишком фрагментарны, чтобы идентифицировать его как динозавра или только его близкого родственника. [116] Отнесение формации Манда к анизийскому отложениям также неясно. Тем не менее, динозавры существовали рядом с орнитодиранами, не являющимися динозаврами, в течение определенного периода времени, по оценкам, от 5 до 10 миллионов лет. [117] до 21 миллиона лет. [113]
Когда появились динозавры, они не были доминирующими наземными животными. Наземные местообитания занимали различные виды архозавроморф и терапсидов , например цинодонты и ринхозавры . Их главными конкурентами были псевдозухии , такие как этоозавры , орнитозухиды и рауизухии, которые добились большего успеха, чем динозавры. [118] Большинство этих других животных вымерло в триасе в результате одного из двух событий. различные базальные архозавроморфы, в том числе проторозавры Во-первых, около 215 миллионов лет назад вымерли . За этим последовало триасово-юрское вымирание (около 201 миллиона лет назад), которое привело к исчезновению большинства других групп ранних архозавров, таких как этоозавры, орнитозухиды, фитозавры и рауизухаи. Ринхозавры и дицинодонты выжили (по крайней мере, в некоторых областях), по крайней мере, в ранне- средненорийском и поздненорийском или самом раннем ретском периодах соответственно. [119] [120] и точная дата их исчезновения неизвестна. Эти потери оставили после себя наземную фауну крокодиломорфов , динозавров, млекопитающих, птерозавров и черепах . [10] Первые несколько линий ранних динозавров диверсифицировались на карнийском и норийском этапах триаса, возможно, заняв ниши вымерших групп. [12] наблюдался повышенный темп вымирания Также примечательно, что во время Карнийского плювиального события . [121]
Эволюция и палеобиогеография

Эволюция динозавров после триаса следовала за изменениями растительности и расположения континентов. В позднем триасе и ранней юре континенты были соединены в единый массив суши Пангея , и существовала всемирная фауна динозавров, в основном состоящая из целофизоидных хищников и ранних травоядных зауроподоморфов. [122] Голосеменные растения (особенно хвойные ), потенциальный источник пищи, распространились в позднем триасе. Ранние зауроподоморфы не имели сложных механизмов переработки пищи во рту, и поэтому, должно быть, использовали другие способы расщепления пищи на дальнейших этапах пищеварительного тракта. [123] Общая однородность фауны динозавров сохранялась и в средней и поздней юре, где в большинстве местонахождений обитали хищники, состоящие из цератозавров , мегалозавроидов и аллозавроидов , и травоядные животные, состоящие из стегозавров-орнитисхий и крупных зауроподов. Примеры этого включают формацию Моррисон в Северной Америке и пласты Тендагуру в Танзании. Динозавры в Китае демонстрируют некоторые различия: специализированные тероподы -метриакантозавриды и необычные зауроподы с длинной шеей, такие как Mamenchisaurus . [122] Анкилозавры и орнитоподы также становились все более распространенными, но примитивные зауроподоморфы вымерли. хвойные и птеридофиты Наиболее распространенными растениями были . Зауроподы, как и более ранние зауроподоморфы, не были пероральными процессорами, но птицетазовые животные разработали различные способы обращения с пищей во рту, включая потенциальные щекоподобные органы для удержания пищи во рту и движения челюстей для измельчения пищи. [123] Другим заметным эволюционным событием юрского периода стало появление настоящих птиц, произошедших от манирапторовых целурозавров. [14]
К раннему меловому периоду и продолжающемуся распаду Пангеи динозавры стали сильно дифференцироваться по суше. распространились В самый ранний период этого времени анкилозавры, игуанодонты и брахиозавриды по Европе, Северной Америке и Северной Африке . Позднее они были дополнены или заменены в Африке крупными спинозавридами и кархародонтозавридами- тероподами, а также реббахизавридами и титанозаврами -зауроподами, также обнаруженными в Южной Америке . В Азии манирапторовые целурозавры, такие как дромеозавриды, троодонтиды и овирапторозавры, стали обычными тероподами, а анкилозавриды и ранние цератопсы, такие как пситтакозавры, стали важными травоядными животными. Между тем, Австралия была домом для фауны базальных анкилозавров, гипсилофодонтов и игуанодонтов. [122] Стегозавры, по-видимому, вымерли в какой-то момент позднего раннего или раннего позднего мела . Главным изменением раннего мела, которое усилилось в позднем мелу, стала эволюция цветковых растений . В то же время несколько групп травоядных динозавров развили более сложные способы пероральной обработки пищи. Цератопсы разработали метод нарезки зубов, сложенных друг на друга в батареи, а игуанодонты усовершенствовали метод измельчения зубными батареями , доведенный до крайности у гадрозаврид. [123] У некоторых зауроподов также развились зубные батареи, лучшим примером которых является реббахизаврид Нигерзавр . [124]
В позднем мелу существовало три основные фауны динозавров. На северных континентах Северной Америки и Азии основными тероподами были тираннозавриды и различные типы более мелких теропод-манирапторов с преимущественно птицетазовыми травоядными животными, состоящими из гадрозавридов, цератопсов, анкилозаврид и пахицефалозавров. На южных континентах, составлявших ныне расколовшийся суперконтинент Гондвана , абелизавриды были обычными тероподами, а титанозавровые зауроподы - обычными травоядными. Наконец, в Европе были распространены дромеозавриды, рабдодонтиды игуанодонты, нодозавриды анкилозавры и титанозавры зауроподы. [122] Цветущие растения сильно светились, [123] первые травы появились к концу мелового периода. [125] Измельчение гадрозавридов и стрижка цератопсов стали очень разнообразными в Северной Америке и Азии. Тероподы также были травоядными или всеядными , при этом теризинозавры и орнитомимозавры стали обычным явлением. [123]
Мел-палеогеновое вымирание, произошедшее примерно 66 миллионов лет назад в конце мелового периода, вызвало вымирание всех групп динозавров, за исключением неорнитиновых птиц. Некоторые другие группы диапсид, в том числе крокодилы , дирозавры , себекозухи , черепахи, ящерицы , змеи , сфенодонты и хористодеры , также пережили это событие. [126]
Сохранившиеся линии неорнитиновых птиц, включая предков современных бескилевых , уток и кур , а также различных водоплавающих птиц , быстро диверсифицировались в начале палеогенового периода , заняв экологические ниши, оставшиеся вакантными в результате вымирания мезозойских групп динозавров, таких как древесные. энантиорнитины , водные гесперорнитины и даже более крупные наземные тероподы (в виде гасторнисов , эогруид , баторнитид , бескилевых, гераноидид , михирунгов и « ужасных птиц »). Часто утверждают, что млекопитающие вытеснили неорнитинов за доминирование в большинстве наземных ниш, но многие из этих групп сосуществовали с богатой фауной млекопитающих на протяжении большей части кайнозойской эры. [127] Ужасные птицы и баторнитиды занимали плотоядные гильдии наряду с хищными млекопитающими. [128] [129] бескилевые все еще довольно успешны как травоядные среднего размера; эогруиды аналогичным образом существовали от эоцена до плиоцена и вымерли совсем недавно, после более чем 20 миллионов лет сосуществования со многими группами млекопитающих. [130]
Классификация


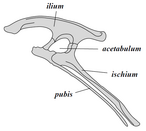

Динозавры принадлежат к группе, известной как архозавры, в которую также входят современные крокодилы. Внутри группы архозавров динозавры наиболее заметно отличаются походкой. Ноги динозавров простираются прямо под телом, тогда как ноги ящериц и крокодилов растягиваются в обе стороны. [30]
В совокупности динозавры как клада делятся на две основные ветви: Saurischia и Ornithischia. Saurischia включает таксоны, имеющие более недавнего общего предка с птицами, чем с Ornithischia, в то время как Ornithischia включает все таксоны, имеющие более недавнего общего предка с трицератопсом , чем с Saurischia. Анатомически эти две группы наиболее заметно различаются по строению таза . Ранние ящерицы — «ящерицетазовые», от греческого sauros ( σαῦρος ), означающего «ящерица», и ischion ( ἰσχίον ), означающего «тазобедренный сустав», — сохранили структуру бедра своих предков, с лобковой костью, направленной краниально или вперед. [37] Эта базовая форма была модифицирована путем поворота лобка назад в разной степени в нескольких группах ( Herrerasaurus , [131] теризинозавроиды, [132] дромеозавриды, [133] и птицы [14] ). Saurischia включает тероподов (исключительно двуногих и с самым разнообразным рационом питания) и зауроподоморфов (длинношеих травоядных, включающих развитые четвероногие группы). [29] [134]
Напротив, птицетазовые - «птичьи бедра» от греческого ornitheios (ὀρνίθειος), означающего «птица» и ischion (ἰσχίον), означающего «тазобедренный сустав», - имели таз, внешне напоминавший птичий таз: лобковая кость была ориентирована каудально (направляясь назад). В отличие от птиц, лобковая кость птицетазовых обычно имела дополнительный направленный вперед отросток. Ornithischia включает множество видов, которые в основном были травоядными.
Несмотря на термины «птичье бедро» (Ornithischia) и «бедро ящерицы» (Saurischia), птицы не являются частью Ornithischia. Вместо этого птицы принадлежат к Saurischia, динозаврам с «ящеричьими бедрами» - птицы произошли от более ранних динозавров с «ящеричьими бедрами». [30]
Таксономия
Ниже приводится упрощенная классификация групп динозавров, основанная на их эволюционных взаимоотношениях, а также классификация основных групп динозавров Theropoda, Sauropodomorpha и Ornithischia, составленная Джастином Твитом. [135] Более подробную информацию и другие гипотезы классификации можно найти в отдельных статьях.
- Динозаврия

- † Ornithischia («птицетазовые»; разнообразные двуногие и четвероногие травоядные животные)
- † Heterodontosauridae (мелкие травоядные/всеядные животные с выступающими клыкообразными зубами )
- † Геназаврии («щекие ящерицы»)
- † Тиреофора (бронированные динозавры; двуногие и четвероногие)
- † Eurypoda (тяжелые четвероногие тиреофоры)
- † Стегозаврия (шипы и пластины в качестве основной брони)
- † Huayangosauridae (маленькие стегозавры с боковыми остеодермами и хвостовыми булавами)
- † Stegosauridae (крупные стегозавры)
- † Анкилозаврия ( щитки как основной панцирь)
- † Паранкилозаврия (маленькие южные анкилозавры с хвостами, похожими на макуауитли )
- † Nodosauridae (в основном остроконечные анкилозавры без булавы)
- † Ankylosauridae (характеризуются плоскими щитками)
- † Ankylosaurinae (булавохвостые анкилозавриды)
- † Neornithischia («новые птицетазовые»)
- † Церапода («рогатые ноги»)
- † Маргиноцефалия (характеризуется краниальным ростом)

- † Пахицефалозаврия (двуногие животные с куполообразным или узловатым наростом на черепе)
- † Ceratopsia (двуногие и четвероногие; у многих были оборки на шее и рога)
- † Chaoyangsauridae (маленькие базальные цератопсы без оборочек)
- † Neoceratopsia («новые цератопсы»)
- † Leptoceratopsidae (почти без излишеств, безрогие, с крепкими челюстями)
- † Protoceratopsidae (базальные цератопсы с небольшими оборками и короткими рогами)
- † Ceratopsoidea (цератопсы с большими рогами)
- † Ceratopsidae (крупные цератопсы с искусным орнаментом)
- † Chasmosaurinae (цератопсиды с увеличенными надбровными рогами)
- † Centrosaurinae (цератопсиды, в основном характеризующиеся оборкой и носовым орнаментом)
- † Nasutoceratopsini (центрозаврины с увеличенной носовой полостью)
- † Центрозаврины (центрозаврины с увеличенными носовыми рогами)
- † Pachyrhinosaurini (чаще всего имели носовые выступы вместо рогов)
- † Орнитоподы (разных размеров; двуногие и четвероногие; развили метод жевания, используя гибкость черепа и многочисленные зубы)
- † Jeholosauridae (небольшие азиатские неорнитишии)
- † Thescelosauridae («чудесные ящерицы»)
- † Orodrominae (роющие)
- † Thescelosaurinae (крупные тецелозавриды)
- † Iguanodontia («зубы игуаны»; развитые орнитоподы)
- † Elasmaria (в основном южные орнитоподы с минерализованными пластинками вдоль ребер; могут быть тесцелозавриды)
- † Рабдодонтоморфа (с характерным зубным рядом)
- † Rhabdodontidae (европейские рабдодонтоморфы)
- † Dryosauridae (среднего размера, с маленькой головой)
- † Анкилополлексия (первые участники среднего размера, коренастые)
- † Стиракостерна («шипастая грудина »)
- † Hadrosauriformes (предки имели шип на большом пальце; крупные четвероногие травоядные животные, зубы слились в зубные батареи)
- † Hadrosauromorpha (гадрозавриды и их ближайшие родственники)
- † Hadrosauridae («утконосые динозавры»; часто с гребнями)
- † Saurolophinae (гадрозавриды с твердыми, маленькими гребнями без гребней)
- † Брахилофосаурин (с коротким гребнем)
- † Критозаурини (увеличенные, твердые носовые гребни)
- † Зауролофини (маленькие шипообразные гребни)
- † Эдмонтозаврины (плоскоголовые зауролофины)
- † Lambeosaurinae (гадрозавриды часто с полыми гребнями)
- † Аралозаурини (с цельным гребнем)
- † Цинтаосаурини (вертикальные трубчатые гребни)
- † Паразауролофины (длинные, загнутые назад гребни)
- † Ламбеозаурини (обычно округлые гребни)
- † Herrerasauridae (ранние двуногие хищники)

- † Sauropodmorpha (травоядные животные с маленькой головой, длинной шеей и длинным хвостом)
- † Unaysauridae (примитивные, строго двуногие «просауроподы»)
- † Платеозаврия (разнообразная: двуногие и четвероногие)
- † Massospondylidae (длинношеие примитивные зауроподоморфы)
- † Riojasauridae (крупные примитивные зауроподоморфы)
- † Sauroodiformes (тяжелые, двуногие и четвероногие)
- † Sauropoda (очень большой и тяжелый; четвероногий)
- † Lessemsauridae (гигантские, но без некоторых приспособлений для снижения веса)
- † Гравизаурия («тяжелые ящерицы»)
- † Eusauropoda («настоящие зауроподы»)
- † Туриазаврия (часто крупные и широко распространенные зауроподы)
- † Neosauropoda («новые зауроподы»; столбчатые конечности)
- † Diplodocoidea (череп и хвост удлиненные; зубы обычно узкие и карандашные)
- † Rebbachisauridae (диплодокоиды с короткой шеей и низким обзором, часто с высокой спиной)
- † Flagellicaudata (хлыстохвостый)
- † Dicraeosauridae (маленькие диплодокоиды с короткой шеей и увеличенными шейными и спинными позвонками)
- † Diplodocidae (очень длинношеие)
- † Apatosaurinae (крепкие шейные позвонки)
- † Diplodocinae (длинные, тонкие шеи)
- † Макронария (коробчатые черепа; зубы в форме ложки или карандаша)
- † Titanosauriformes («формы титановой ящерицы»)
- † Brachiosauridae (макронарии с длинной шеей и длинными руками)
- † Сомфоспондилы («пористые позвонки»)
- † Euhelopodidae (коренастые, в основном азиатские)
- † Диамантиназаврия (лошадиные черепа; ограничены южным полушарием; могут быть титанозаврами)
- † Титанозаврия (разнообразная; коренастая, с широкими бедрами; наиболее распространена в позднем мелу южных континентов)
- Тероподы (плотоядные)
- Neotheropoda («новые тероподы»)
- † Coelophysoidea (ранние тероподы; включает целофизов и близких родственников)
- † «Неотероподы класса дилофозавров» (более крупные динозавры с изогнутой мордой)
- Аверостра («птичья морда»)
- † Цератозаврия (как правило, хищные животные с рогами, существовавшие от юрского до мелового периода, первоначально включали Coelophysoidea)
- † Ceratosauridae (цератозавры с большими зубами)
- † Abelisauroidea (цератозавры, на примере уменьшенных рук и кистей)
- † Abelisauridae (крупные абелизавроиды с короткими руками и часто сложным орнаментом на лице)
- † Noasauridae (разнообразные, обычно легкие тероподы; могут включать несколько малоизвестных таксонов)
- † Elaphrosaurinae (птицеподобные; всеядны в молодом возрасте, но травоядны во взрослом возрасте)
- † Noasaurinae (мелкие хищники)
- Тетануры (жесткохвостые динозавры)
- † Megalosauroidea (ранняя группа крупных хищников)
- † Piatnitzkysauridae (небольшие базальные мегалозавроиды, эндемичные для Америки)
- † Мегалозавриды (крупные мегалозавроиды с мощными руками и кистями)
- † Spinosauidae (крокодилоподобные полуводные хищники)
- Avetheropoda («птицы-тероподы»)
- † Карнозаврия (крупные плотоядные динозавры; иногда включают мегалозавроидов)
- † Metriacanthosauridae (примитивные азиатские аллозавроиды)
- † Allosauridae ( аллозавр и его ближайшие родственники)
- † Кархародонтозаврия (крепкие аллозавроиды)
- † Carcharodontosauridae (включает некоторых из крупнейших чисто наземных хищников)
- † Neovenatoridae («новые охотники»; могут включать мегарапторов)
- Целурозаврия (пернатые тероподы с разными размерами тела и нишами)
- † Мегараптора ? (тероподы с большими когтями на руках; потенциально тираннозавроиды или неовенаториды)
- † «Связь базальных целурозавров» (используется в Твиттере для обозначения хорошо известных таксонов с нестабильным положением в основании целурозаврии)
- Тираннораптора («воры-тираны»)
- † Tyrannosauroidea (в основном крупные примитивные целурозавры)
- † Proceratosauridae (тираннозавроиды с гребнями на голове)
- † Tyrannosauridae ( тираннозавр и близкие родственники)
- Манирапториформы (птицеобразные динозавры)
- † Орнитомимозаврия (малоголовые, преимущественно беззубые, всеядные или, возможно, травоядные)
- † Ornithomimidae (динозавры, очень похожие на страуса)
- Манираптора (динозавры с перистыми перьями)

- † Alvarezsauroidea (маленькие охотники с редуцированными передними конечностями)
- † Alvarezsauridae (насекомоядные только с одним увеличенным пальцем)
- † Теризинозаврия (высокие тероподы с длинной шеей; всеядные и травоядные)
- † Therizinosauroidea (более крупные теризинозавры)
- † Therizinosauridae (ленивцы-травоядные, часто с увеличенными когтями)
- † Овирапторозаврия (всеядные клювистые динозавры)
- † Caudipteridae (птицеобразные, базальные овирапторозавры)
- † Caenagnathoidea (казуароподобные овирапторозавры)
- † Caenagnathidae (беззубые овирапторозавры, известные из Северной Америки и Азии)
- † Oviraptoridae (характеризуются двумя костными выступами в задней части рта; исключительно для Азии)
- Паравесы (авиаланы и их ближайшие родственники)
- † Scansoriopterygidae (небольшие древесные тероподы с перепончатыми крыльями)
- † Дейнонихозаврия (когтистые динозавры; не могут образовывать естественную группу)
- † Archaeopterygidae (маленькие крылатые тероподы или примитивные птицы)
- † Troodontidae (всеядные; увеличенные полости мозга)
- † Dromaeosauridae («хищники»)
- † Микрораптория (характеризуется большими крыльями на руках и ногах; возможно, была способна летать с помощью двигателя)
- † Эудромеозаврия (охотники с сильно увеличенными серповидными когтями)
- † Unenlagiidae (рыбоядные; могут быть дромеозавридами)
- † Halszkaraptorinae (утиные; потенциально полуводные)
- † Unenlagiinae (длиннорылые)
- Avialae (современные птицы и вымершие родственники)
Хронология основных групп
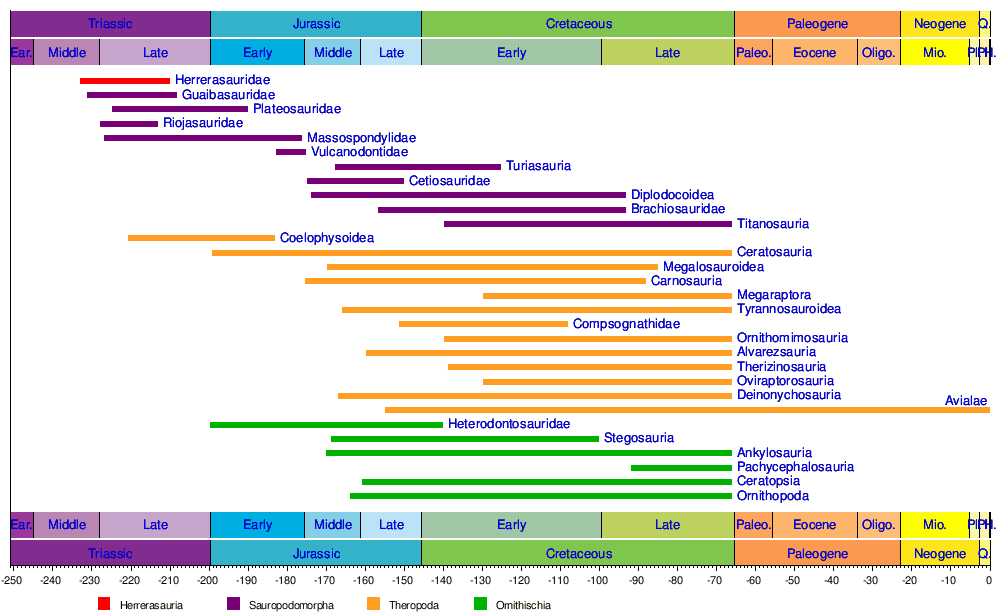
Хронология основных групп динозавров по Хольцу (2007) .
Палеобиология
Знания о динозаврах получены из множества ископаемых и неископаемых материалов, включая окаменелые кости, фекалии , следы , гастролиты , перья , отпечатки кожи, внутренних органов и других мягких тканей . [90] [94] Многие области исследований способствуют нашему пониманию динозавров, включая физику (особенно биомеханику ), химию , биологию и науки о Земле которых является палеонтология ). ( поддисциплиной [136] [137] Две темы, представляющие особый интерес и изучение, — это размер и поведение динозавров. [138]
Размер

Имеющиеся данные свидетельствуют о том, что средний размер динозавров менялся в триасовом, раннем юрском, позднеюрском и меловом периодах. [111] Хищные динозавры-тероподы, которые занимали большую часть ниш наземных хищников в мезозое, чаще всего попадают в категорию от 100 до 1 000 кг (от 220 до 2 200 фунтов) при сортировке по предполагаемому весу на категории на основе порядка величины , тогда как современные хищные хищные млекопитающие пик в категории от 10 до 100 кг (от 22 до 220 фунтов). [139] Масса . тела мезозойских динозавров составляет от 1 до 10 метрических тонн (от 1,1 до 11,0 коротких тонн) [140] Это резко контрастирует со средним размером кайнозойских млекопитающих, который, по оценкам Национального музея естественной истории , составляет от 2 до 5 кг (от 4,4 до 11,0 фунтов). [141]
Зауроподы были самыми крупными и тяжелыми динозаврами. На протяжении большей части эпохи динозавров самые маленькие зауроподы были крупнее, чем все остальное в их среде обитания, а самые крупные были на порядок массивнее, чем все остальное, что с тех пор ступало по Земле. Гигантские доисторические млекопитающие, такие как Paraceratherium (крупнейшее наземное млекопитающее из когда-либо существовавших), были затмеваемы гигантскими зауроподами, и только современные киты приближаются или превосходят их по размеру. [142] Есть несколько предполагаемых преимуществ большого размера зауроподов, включая защиту от хищников, снижение энергопотребления и долголетие, но, возможно, самым важным преимуществом было диетическое питание. Крупные животные более эффективно переваривают пищу, чем мелкие, поскольку пища проводит больше времени в их пищеварительной системе. Это также позволяет им питаться пищей с более низкой питательной ценностью, чем у более мелких животных. Останки зауроподов в основном находят в скальных образованиях, которые считаются сухими или сезонно сухими, и возможность поедать большое количество растительности с низким содержанием питательных веществ была бы выгодна в таких средах. [143]
Самый большой и самый маленький
Ученые, вероятно, никогда не будут уверены в том, самые большие и самые маленькие динозавры какие когда-либо существовали . Это связано с тем, что лишь небольшой процент животных когда-либо был окаменел, и большинство из них остаются погребенными в земле. Лишь немногие обнаруженные экземпляры нептичьих динозавров представляют собой полные скелеты, а отпечатки кожи и других мягких тканей редки. Восстановление полного скелета путем сравнения размера и морфологии костей с костями аналогичных, более известных видов — неточное искусство, а реконструкция мышц и других органов живого животного — это в лучшем случае процесс обоснованных догадок. [144]

Самый высокий и тяжелый динозавр, известный по хорошим скелетам, — Giraffatitan brancai (ранее классифицировавшийся как разновидность брахиозавра ). Его останки были обнаружены в Танзании между 1907 и 1912 годами. Кости нескольких особей одинакового размера были включены в скелет, который сейчас установлен и выставлен в Музее природы в Берлине ; [145] это крепление имеет высоту 12 метров (39 футов) и длину от 21,8 до 22,5 метров (от 72 до 74 футов), [146] [147] и принадлежал бы животному весом от 30 000 до 60 000 килограммов ( от 70 000 до 130 000 фунтов). Самый длинный полный динозавр — это диплодок длиной 27 метров (89 футов) , который был обнаружен в Вайоминге в США и выставлен в Питтсбурге в Музее естественной истории Карнеги в 1907 году. [148] Самый длинный динозавр, известный по достоверному ископаемому материалу, — это Патаготитан : скелет в Американском музее естественной истории в Нью-Йорке имеет длину 37 метров (121 фут). В Муниципальном музее Кармен Фунес на площади Уинкул в Аргентине хранится реконструированный скелет аргентинозавра длиной 39,7 метра (130 футов). [149]


Существовали и более крупные динозавры, но знания о них полностью основаны на небольшом количестве фрагментарных окаменелостей. Большинство крупнейших травоядных экземпляров были обнаружены в 1970-х годах или позже, и включают массивного аргентинозавра , который, возможно, весил от 80 000 до 100 000 килограммов (от 88 до 110 коротких тонн) и достигал длины от 30 до 40 метров (от 98 до 110 коротких тонн). 131 фут); длиной 33,5 метра (110 футов). Одними из самых длинных были Diplodocus Hallorum [143] (ранее сейсмозавр длиной от 33 до 34 метров (от 108 до 112 футов) ), суперзавр , [150] длиной 37 метров (121 фут) и Патаготитан ; и самый высокий, 18-метровый (59 футов) Зауропосейдон , который мог достигать окна шестого этажа. Было несколько динозавров, которых считали самыми тяжелыми и длинными. Самым известным из них является Amphicoelias fragillimus , известный только по ныне утраченной частичной нервной дуге позвонка , описанной в 1878 году. Экстраполируя изображение этой кости, можно сказать, что животное могло иметь длину 58 метров (190 футов) и весить 122 400 кг ( 269 800). фунт). [143] Однако недавние исследования позволили амфикоэлиям перейти от длинных, изящных диплодоцидов к более коротким, но гораздо более коренастым реббахизавридам. Этот зауропод, переименованный теперь в Мараапунизавра , достигает 40 метров (130 футов) в длину и весит до 120 000 кг ( 260 000 фунтов). [151] [152] Еще одним претендентом на это звание является Bruhathkayosaurus , спорный таксон, существование которого недавно было подтверждено после того, как были обнаружены архивные фотографии. [153] Брухаткайозавр был титанозавром и, скорее всего, весил бы больше, чем даже маррапунизавр . 000–370 000 фунтов кг ) Недавние оценки размеров, сделанные в 2023 году, показали, что этот зауропод достигает длины до 44 м (144 футов) и колоссального веса в пределах 110 000–170 000 , если эти 240 верхние ( оценки верны. Брухаткайозавр мог бы соперничать с синим китом и колоссом Perucetus как одно из крупнейших животных, когда-либо существовавших. [154]
Самым крупным плотоядным динозавром был спинозавр , достигающий длины от 12,6 до 18 метров (от 41 до 59 футов) и веса от 7 до 20,9 метрических тонн (от 7,7 до 23,0 коротких тонн). [155] [156] Другими крупными хищными тероподами были гиганотозавр , кархародонтозавр и тираннозавр . [156] Теризинозавр и дейнохейрус были одними из самых высоких теропод. Самым крупным птицетазовым динозавром, вероятно, был гадрозаврид Shantungosaurus giganteus , длина которого составляла 16,6 метра (54 фута). [157] Самые крупные особи могли весить до 16 метрических тонн (18 коротких тонн). [158]

Самый маленький из известных динозавров — колибри-пчелка . [159] длиной всего 5 сантиметров (2,0 дюйма) и массой около 1,8 г (0,063 унции). [160] Самые маленькие из известных неавиальных динозавров были размером с голубя и были тероподами, наиболее близкими родственниками птиц. [161] Например, Anchiornis huxleyi в настоящее время является самым маленьким нептичьим динозавром, описанным по взрослому экземпляру, его предполагаемый вес составляет 110 г (3,9 унции). [162] и общая длина скелета 34 сантиметра (1,12 фута). [161] [162] Самыми маленькими травоядными нептичьими динозаврами были Microceratus и Wannanosaurus , длиной около 60 сантиметров (2,0 фута) каждый. [163] [164]
Поведение

Многие современные птицы очень социальны и часто живут стаями. Существует общее мнение, что некоторые модели поведения, характерные для птиц, а также для крокодилов (ближайших ныне живущих родственников птиц), также были распространены среди вымерших групп динозавров. Интерпретации поведения ископаемых видов обычно основаны на позе скелетов и их среде обитания , компьютерном моделировании их биомеханики и сравнении с современными животными в аналогичных экологических нишах. [136]
Первым потенциальным доказательством того, что стадное или стадное поведение широко распространено среди многих групп динозавров, помимо птиц, было открытие в 1878 году 31 игуанодона , птицетазовых животных, которые, как тогда считалось, погибли вместе в Берниссаре , Бельгия , после того, как упали в глубокую глубину. затопил провал и утонул. [165] Впоследствии были обнаружены и другие места массовой гибели людей. Это, наряду с наличием множества следов, позволяет предположить, что стадное поведение было обычным явлением для многих ранних видов динозавров. Следы сотен или даже тысяч травоядных указывают на то, что утконосые (гадрозавриды) могли перемещаться большими стадами, как американский бизон или африканский спрингбок . Следы зауроподов документально подтверждают, что эти животные путешествовали группами, состоящими из нескольких разных видов, по крайней мере, в Оксфордшире , Англия. [166] хотя нет никаких доказательств существования конкретной стадной структуры. [167] Объединение в стада могло возникнуть для защиты, миграции или защиты молодняка. Имеются данные о том, что многие виды медленнорастущих динозавров, в том числе различные тероподы, зауроподы, анкилозавры, орнитоподы и цератопсы, образовывали скопления неполовозрелых особей. Одним из примеров является участок во Внутренней Монголии , где были обнаружены останки более 20 синонитомимов в возрасте от одного до семи лет. Эта сборка интерпретируется как социальная группа, застрявшая в грязи. [168] Интерпретация динозавров как стадных животных также распространилась на изображение плотоядных теропод как стадных охотников, работающих вместе, чтобы поймать крупную добычу. [169] [170] Однако такой образ жизни необычен среди современных птиц, крокодилов и других рептилий, и тафономические данные, свидетельствующие о стайной охоте, похожей на млекопитающих, у таких теропод, как дейноних и аллозавр, также могут быть интерпретированы как результаты фатальных споров между кормящимися животными, как видно. у многих современных диапсидных хищников. [171]

Гребни и оборки некоторых динозавров, таких как маргиноцефалы , тероподы и ламбеозаврины , возможно, были слишком хрупкими, чтобы их можно было использовать для активной защиты, и поэтому они, вероятно, использовались для сексуальных или агрессивных проявлений, хотя о спаривании динозавров и территориализме мало что известно . Ранения головы от укусов позволяют предположить, что тероподы, как минимум, вели активные агрессивные противостояния. [172]
С поведенческой точки зрения, одна из самых ценных окаменелостей динозавров была обнаружена в пустыне Гоби в 1971 году. Среди них были велоцираптор, напавший на протоцератопса , [173] предоставляя доказательства того, что динозавры действительно нападали друг на друга. [174] Дополнительным свидетельством нападения на живую добычу является частично заживший хвост эдмонтозавра , динозавра-гадрозаврида; хвост поврежден таким образом, что видно, что животное было укушено тираннозавром, но выжило. [174] Каннибализм среди некоторых видов динозавров был подтвержден следами зубов, найденными на Мадагаскаре в 2003 году у теропода Majungasaurus . [175]
Сравнение склеральных колец динозавров и современных птиц и рептилий было использовано для определения режима повседневной активности динозавров. Хотя было высказано предположение, что большинство динозавров были активны в течение дня, эти сравнения показали, что мелкие хищные динозавры, такие как дромеозавриды, юравенатор и мегапнозавр , вероятно, вели ночной образ жизни . Крупные и средние травоядные и всеядные динозавры, такие как цератопсы, зауроподоморфы, гадрозавриды, орнитомимозавры, возможно, вели катемерный образ жизни и были активны в течение коротких промежутков времени в течение дня, хотя небольшой птицетазовый агилизавр предполагалось, что вел дневной образ жизни . [176]
Судя по ископаемым свидетельствам динозавров, таких как Oryctodromeus , некоторые виды птицетазовых, по-видимому, вели частично ископаемый (роющий) образ жизни. [177] Многие современные птицы ведут древесный образ жизни (лазат по деревьям), и это справедливо и для многих мезозойских птиц, особенно энантиорнитинов. [178] Хотя некоторые ранние птицеподобные виды, возможно, уже были древесными (включая дромеозаврид), например микрораптор. [179] ) большинство нептичьих динозавров, похоже, полагались на наземное передвижение. Хорошее понимание того, как динозавры передвигались по земле, является ключом к моделям поведения динозавров; наука биомеханика, основанная Робертом Макнейлом Александром , дала значительное понимание в этой области. Например, исследования сил, действующих на структуру скелета динозавров со стороны мышц и силы тяжести, позволили выяснить, насколько быстро динозавры могли бегать. [136] могут ли диплодоциды создавать звуковые удары , щелкая хвостом, как кнут , [180] и могли ли зауроподы плавать. [181]
Коммуникация
Известно, что современные птицы общаются с помощью визуальных и слуховых сигналов, а широкое разнообразие структур визуального отображения среди групп ископаемых динозавров, таких как рога, оборки, гребни, паруса и перья, позволяет предположить, что визуальная коммуникация всегда была важна в биологии динозавров. [182] Реконструкция цвета оперения Anchiornis позволяет предположить важность цвета в визуальной коммуникации у нептичьих динозавров. [183] Вокализация у нептичьих динозавров менее очевидна. У птиц гортань не играет никакой роли в производстве звука. Вместо этого они издают звуки с помощью нового органа, называемого сиринксом , расположенного дальше по трахее. [184] Самые ранние остатки сиринкса были обнаружены у экземпляра утиноподобного Vegavis iaai, датируемого 69–66 миллионами лет назад, и маловероятно, что этот орган существовал у нептичьих динозавров. [185]

Палеонтолог Фил Сентер предположил, что, поскольку у нептичьих динозавров не было сиринкса, а их ближайшие ближайшие родственники, крокодилы, использовали гортань, они не могли издавать звуки, поскольку общий предок был немым. Он утверждает, что они в основном связаны с визуальными проявлениями и, возможно, неголосовыми акустическими звуками, такими как шипение, скрежетание челюстями или хлопки в ладоши, плеск и взмах крыльев (возможно у крылатых динозавров-манирапторов). [182] Другие исследователи возражают, что вокализации также существуют у черепах, ближайших родственников архозавров, предполагая, что эта черта является предковой для их происхождения. Кроме того, на голосовое общение динозавров указывает развитие развитого слуха почти во всех основных группах. Следовательно, сиринкс, возможно, дополнил, а затем заменил гортань как голосовой орган, а не в «периоде молчания» в эволюции птиц. [186]
В 2023 году окаменевшая гортань была описана у экземпляра анкилозавридного пинакозавра . Структура состояла из перстневидных и черпаловидных хрящей , аналогичных таковым у нептичьих рептилий. Однако подвижный перстнечерпаловидный сустав и длинные черпаловидные хрящи позволяли бы контролировать воздушный поток, как у птиц, и, таким образом, могли издавать птичьи вокалы. Кроме того, хрящи были окостеневшими , а это означает, что окостенение гортани является особенностью некоторых нептичьих динозавров. [187] Исследование 2016 года пришло к выводу, что некоторые динозавры могли издавать звуки с закрытым ртом, такие как воркование, уханье и гул. Они встречаются как у рептилий, так и у птиц и включают раздувание пищевода или трахеальных мешков. Такие вокализации неоднократно развивались независимо у современных архозавров после увеличения размера тела. [188] Было высказано предположение, что гребни некоторых гадрозаврид и носовые камеры анкилозаврид функционировали в акустическом резонансе . [189] [190]
Репродуктивная биология

Все динозавры откладывали околоплодные яйца . Яйца динозавров обычно откладывались в гнезде. Большинство видов создают довольно сложные гнезда, которые могут представлять собой чашки, купола, тарелки, царапины на грядках, насыпи или норы. [191] Некоторые виды современных птиц не имеют гнезд; гнездящаяся на скалах, обыкновенная кайра, откладывает яйца на голые камни, а императорские пингвины- самцы держат яйца между телом и ногами. Примитивные птицы и многие нептичьи динозавры часто откладывают яйца в общие гнезда, причем яйца в основном насиживают самцы. В то время как современные птицы имеют только один функциональный яйцевод и откладывают одно яйцо за раз, у более примитивных птиц и динозавров было два яйцевода, как у крокодилов. Некоторые нептичьи динозавры, такие как троодон , демонстрировали итеративную кладку, при которой взрослая особь могла откладывать пару яиц каждые один или два дня, а затем обеспечивала одновременное вылупление, откладывая высиживание до тех пор, пока все яйца не были отложены. [192]
При откладке яиц у самок вырастает особый тип кости между твердой внешней костью и костным мозгом конечностей. Эта мозговая кость, богатая кальцием , используется для изготовления яичной скорлупы. Открытие особенностей скелета тираннозавра предоставило доказательства наличия медуллярной кости у вымерших динозавров и впервые позволило палеонтологам установить пол ископаемого экземпляра динозавра. Дальнейшие исследования обнаружили медуллярную кость у карнозавра Allosaurus и орнитопод Tenontosaurus . Поскольку линия динозавров, включающая аллозавра и тираннозавра, на очень ранних этапах эволюции динозавров отошла от линии, которая привела к тенонтозаврам , это позволяет предположить, что производство мозговой ткани является общей характеристикой всех динозавров. [193]

Еще одна широко распространенная черта среди современных птиц (но о ископаемых группах и современных многоногих см . Ниже ) — это родительская забота о птенцах после вылупления. Джеком Хорнером Открытие в 1978 году места гнездования майазауры («хорошей ящерицы-матери») в Монтане продемонстрировало, что родительская забота среди орнитопод продолжается еще долго после рождения. [194] Образец овирапторида Citipati osmolskae был обнаружен в насиживающем состоянии, напоминающем курицу, в 1993 году. [195] это может указывать на то, что они начали использовать изолирующий слой перьев, чтобы сохранять яйца теплыми. [196] Эмбрион базального зауроподоморфа Massospondylus был обнаружен без зубов, что указывает на то, что для кормления молодых динозавров требовалась некоторая родительская забота. [197] Следы также подтвердили родительское поведение среди орнитопод с острова Скай на северо-западе Шотландии . [198]
Тем не менее, существует множество свидетельств скороспелости или сверхпрекоциальности среди многих видов динозавров, особенно теропод. Например, было неоднократно продемонстрировано, что птицы, не являющиеся орнитуроморфами, имели медленные темпы роста, поведение, подобное мегаподам , закапывающим яйца, и способность летать вскоре после рождения. [199] [200] [201] [202] И у тираннозавра , и у троодона были молодые особи с явной сверхскороспелостью, которые, вероятно, занимали другие экологические ниши, чем взрослые особи. [192] Для зауроподов был сделан вывод о сверхпрекоциальности. [203]
Генитальные структуры вряд ли окаменеют, поскольку у них нет чешуи, которая могла бы обеспечить сохранение за счет пигментации или остаточных солей фосфата кальция. динозавра клоакального отверстия был описан наиболее сохранившийся образец внешней части В 2021 году у Psittacosaurus , демонстрирующий боковые вздутия, похожие на крокодиловые мускусные железы, используемые в социальных проявлениях представителями обоих полов и пигментированными областями, что также может отражать сигнальную функцию. Однако сам по себе этот экземпляр не дает достаточно информации, чтобы определить, обладал ли этот динозавр функциями передачи половых сигналов; оно только подтверждает возможность. Клоакальная визуальная сигнализация может возникать как у самцов, так и у самок ныне живущих птиц, поэтому маловероятно, что определение пола вымерших динозавров окажется полезным. [204]
Физиология
Поскольку и современные крокодилы, и птицы имеют четырехкамерное сердце (хотя и видоизмененное у крокодилов), вполне вероятно, что эта черта присуща всем архозаврам, включая всех динозавров. [205] Хотя все современные птицы обладают высоким метаболизмом и являются эндотермическими («теплокровными»), с 1960-х годов продолжаются энергичные дебаты о том, насколько далеко в линии динозавров распространилась эта черта. Различные исследователи считают, что динозавры были эндотермными, экзотермическими («хладнокровными») или чем-то средним между ними. [206] Среди исследователей появляется консенсус в том, что, хотя разные линии динозавров имели разный метаболизм, у большинства из них скорость метаболизма была выше, чем у других рептилий, но ниже, чем у ныне живущих птиц и млекопитающих. [207] называют это мезотермией . некоторые [208] Данные, полученные от крокодилов и их вымерших родственников, позволяют предположить, что такой повышенный метаболизм мог развиться у самых ранних архозавров, которые были общими предками динозавров и крокодилов. [209] [210]

После того, как были обнаружены нептичьи динозавры, палеонтологи впервые предположили, что они были экзотермическими. Это использовалось, чтобы подразумевать, что древние динозавры были относительно медленными и медлительными организмами, хотя многие современные рептилии быстры и легконоги, несмотря на то, что для регулирования температуры своего тела полагались на внешние источники тепла. Идея об экзотермических динозаврах оставалась преобладающей точкой зрения до тех пор, пока Роберт Т. Баккер , один из первых сторонников эндотермии динозавров, не опубликовал влиятельную статью по этой теме в 1968 году. Баккер специально использовал анатомические и экологические данные, чтобы доказать, что зауроподы, которые до сих пор изображались как распростертые водные животные с волочащимися по земле хвостами, они были эндотермными животными, ведущими энергичный наземный образ жизни. В 1972 году Баккер расширил свои аргументы, основанные на энергетических потребностях и соотношении хищник-жертва. Это был один из основополагающих результатов, приведших к возрождению динозавров. [63] [64] [60] [211]
Одним из величайших вкладов в современное понимание физиологии динозавров стала палеогистология — исследование микроскопической структуры тканей динозавров. [212] [213] Начиная с 1960-х годов Арманд де Риклес предположил, что наличие фиброламеллярной кости — костной ткани с неправильной волокнистой текстурой, заполненной кровеносными сосудами — указывает на постоянно быстрый рост и, следовательно, на эндотермию. Фиброламеллярная кость была распространена как у динозавров, так и у птерозавров. [214] [215] хотя и не повсеместно. [216] [217] Это привело к проведению значительного объема работы по реконструкции кривых роста и моделированию эволюции темпов роста различных линий динозавров. [218] что в целом позволяет предположить, что динозавры росли быстрее, чем нынешние рептилии. [213] Другие свидетельства, свидетельствующие об эндотермии, включают наличие перьев и других типов покровов тела у многих линий (см. § Перья ); более стабильное соотношение изотопа кислорода-18 в костной ткани по сравнению с эктотермными животными, особенно при изменении широты и, следовательно, температуры воздуха, что предполагает стабильные внутренние температуры. [219] [220] (хотя эти соотношения могут быть изменены во время окаменения [221] ); и открытие полярных динозавров , которые жили в Австралии, Антарктиде и на Аляске, когда в этих местах был прохладный умеренный климат. [222] [223] [224] [225]

У ящерообразных динозавров более высокий метаболизм был поддержан эволюцией птичьей дыхательной системы, характеризующейся обширной системой воздушных мешков , которые расширяли легкие и проникали во многие кости скелета, делая их полыми. [226] Такие дыхательные системы, которые, возможно, появились у самых ранних ящерообразных, [227] обеспечивало бы их большим количеством кислорода по сравнению с млекопитающими такого же размера, а также имело бы больший дыхательный объем в состоянии покоя и требовало бы более низкой частоты дыхания, что позволило бы им поддерживать более высокий уровень активности. [142] Быстрый поток воздуха также мог бы стать эффективным механизмом охлаждения, который в сочетании с более низкой скоростью метаболизма [228] предотвратил бы перегрев крупных зауроподов. Эти черты, возможно, позволили зауроподам быстро вырасти до гигантских размеров. [229] [230] Зауроподы, возможно, также выиграли от своего размера — их небольшое соотношение площади поверхности к объему означало, что они могли легче регулировать терморегуляцию — явление, называемое гигантотермией . [142] [231]
Как и другие рептилии, динозавры в первую очередь являются урикотеликами , то есть их почки извлекают азотистые отходы из кровотока и выделяют их в виде мочевой кислоты вместо мочевины или аммиака через мочеточники в кишечник. Это помогло бы им экономить воду. [207] У большинства живых видов мочевая кислота выделяется вместе с калом в виде полутвердых отходов. [232] [233] Однако, по крайней мере, некоторые современные птицы (например, колибри ) могут быть факультативно аммонотелическими , выделяя большую часть азотистых отходов в виде аммиака. [234] Этот материал, как и продукты кишечника, выходит из клоаки . [235] [236] Кроме того, многие виды срыгивают гранулы , [237] а ископаемые гранулы известны еще в юрском периоде из Анхиорниса . [238]
Размер и форму мозга можно частично реконструировать по окружающим костям. В 1896 году Марш рассчитал соотношение между массой мозга и массой тела семи видов динозавров, показав, что мозг динозавров был пропорционально меньше, чем у современных крокодилов, и что мозг стегозавра был меньше, чем у любого живущего наземного позвоночного. Это способствовало распространению в обществе представления о динозаврах как о медлительных и чрезвычайно глупых существах. Гарри Джерисон в 1973 году показал, что при больших размерах тела ожидается пропорционально меньший мозг, и что размер мозга динозавров был не меньше ожидаемого по сравнению с нынешними рептилиями. [239] Более поздние исследования показали, что относительный размер мозга постепенно увеличивался в ходе эволюции теропод, причем самый высокий интеллект, сравнимый с интеллектом современных птиц, был рассчитан у троодонтида Троодона . [240]
Происхождение птиц
Возможность того, что динозавры были предками птиц, была впервые предложена в 1868 году Томасом Генри Хаксли . [241] После работы Герхарда Хайльмана в начале 20-го века от теории птиц как потомков динозавров отказались в пользу идеи о том, что они являются потомками обобщенных текодонтов , причем ключевым доказательством является предполагаемое отсутствие ключиц у динозавров. [242] Однако, как показали более поздние открытия, ключицы (или один сросшийся поперечный рычаг , образовавшийся из отдельных ключиц) на самом деле не отсутствовали; [14] они были обнаружены еще в 1924 году у Овираптора , но ошибочно идентифицированы как межключица . [243] В 1970-х годах Остром возродил теорию динозавров и птиц. [244] который получил импульс в ближайшие десятилетия с появлением кладистического анализа, [245] и резкое увеличение количества открытий мелких теропод и ранних птиц. [32] Особого внимания заслуживают окаменелости биоты Джехол, где были обнаружены разнообразные тероподы и ранние птицы, часто с перьями того или иного типа. [70] [14] Птицы имеют более сотни общих анатомических особенностей с динозаврами-тероподами, которые в настоящее время считаются их ближайшими древними родственниками. [246] Они наиболее тесно связаны с целурозаврами-манирапторами. [14] Меньшая часть ученых, в первую очередь Алан Федучча и Ларри Мартин , предложили другие пути эволюции, включая пересмотренные версии предположения Хейльмана о базальных архозаврах. [247] или что тероподы-манирапторы являются предками птиц, но сами не являются динозаврами, а лишь сходятся с динозаврами. [248]
Перья

Перья — одна из наиболее узнаваемых характеристик современных птиц, а также черта, которая была свойственна нескольким нептичьим динозаврам. Судя по нынешнему распределению ископаемых свидетельств, кажется, что перья были наследственной чертой динозавров, хотя у некоторых видов она могла быть избирательно утеряна. [249] Прямые ископаемые свидетельства наличия перьев или пероподобных структур были обнаружены у множества видов во многих группах динозавров, не являющихся птицами. [70] как среди ящеротазовых, так и среди птицетазовых. Простые, разветвленные, перьевые структуры известны у гетеродонтозаврид , примитивных неорнитиший , [250] и тероподы, [251] и примитивные цератопсы. Доказательства существования настоящих лопастных перьев, похожих на маховые перья современных птиц, были обнаружены только у подгруппы теропод Maniraptora, в которую входят овирапторозавры, троодонтиды, дромеозавриды и птицы. [14] [252] Пероподобные структуры, известные как пикноволокна, также были обнаружены у птерозавров. [253]
Однако исследователи не пришли к единому мнению относительно того, имеют ли эти структуры общее происхождение между линиями (т. е. гомологичны ли они ). [254] [255] или же они были результатом широко распространенных экспериментов с кожными покровами среди орнитодиран. [256] Если это так, то нити, возможно, были обычным явлением в линии орнитодиров и развились до появления самих динозавров. [249] Исследования генетики американских аллигаторов показали, что щитки крокодилов действительно обладают кератинами перьев во время эмбрионального развития, но эти кератины не экспрессируются животными до вылупления. [257] Описание пернатых динозавров в целом не обошлось без противоречий; возможно, самыми ярыми критиками были Алан Федучча и Теагартен Лингем-Солиар, которые предположили, что некоторые предполагаемые перьевые окаменелости являются результатом разложения коллагеновых волокон, лежащих в основе кожи динозавров. [258] [259] [260] и что динозавры-манирапторы с лопастными перьями на самом деле были не динозаврами, а конвергентными динозаврами. [248] [259] Однако их взгляды по большей части не были приняты другими исследователями до такой степени, что научная природа предложений Федуччиа была поставлена под сомнение. [261]
Археоптерикс был первой найденной окаменелостью, которая выявила потенциальную связь между динозаврами и птицами. Его считают переходным ископаемым , поскольку он демонстрирует черты обеих групп. Обнаруженное всего через два года после Чарльза Дарвина основополагающей работы «Происхождение видов» (1859 г.), его открытие стимулировало зарождающуюся дискуссию между сторонниками эволюционной биологии и креационизмом . Эта ранняя птица настолько похожа на динозавра, что, не имея четких следов перьев на окружающей скале, по крайней мере, один экземпляр был принят за маленького теропода Compsognathus . [262] С 1990-х годов было обнаружено еще несколько пернатых динозавров, что еще больше доказывает тесную связь между динозаврами и современными птицами. Многие из этих экземпляров были обнаружены в лагерштеттене биоты Йехол. [255] Если перьевые структуры действительно широко присутствовали среди нептичьих динозавров, отсутствие обильных ископаемых свидетельств их существования может быть связано с тем, что такие тонкие особенности, как кожа и перья, редко сохраняются в результате окаменелости и, следовательно, часто отсутствуют в летописи окаменелостей. [263]
Скелет
Поскольку перья часто ассоциируются с птицами, пернатых динозавров часто рекламируют как недостающее звено между птицами и динозаврами. Однако множественные особенности скелета, также общие для этих двух групп, представляют собой еще одно важное доказательство для палеонтологов. Области скелета, имеющие важное сходство, включают шею, лобок, запястье (полулунное запястье ), руку и грудной пояс , вилку (поперечный рычаг) и грудную кость . Сравнение скелетов птиц и динозавров посредством кладистического анализа подтверждает наличие связи. [264]
Мягкая анатомия

Согласно исследованию 2005 года, проведенному Патриком М. О'Коннором, у крупных плотоядных динозавров была сложная система воздушных мешков, аналогичная той, что есть у современных птиц. Легкие динозавров-теропод (хищных животных, которые ходили на двух ногах и имели птичьи ступни), вероятно, закачивали воздух в полые мешочки в их скелетах, как и в случае с птицами. «То, что когда-то формально считалось уникальным для птиц, в той или иной форме присутствовало у предков птиц», - сказал О'Коннор. [265] [266] В 2008 году ученые описали Aerosteon riocoloradensis , скелет которого является самым убедительным на сегодняшний день доказательством существования динозавра с птичьей системой дыхания. КТ-сканирование ископаемых костей Аэростеона выявило доказательства существования воздушных мешков в полости тела животного. [226] [267]
Поведенческие доказательства
Окаменелости троодонтов Mei и Sinornithoides демонстрируют, что некоторые динозавры спали, подвернув головы под руки. [268] Такое поведение, которое, возможно, помогало сохранять голову в тепле, характерно и для современных птиц. Несколько экземпляров дейнонихозавров и овирапторозавров также были обнаружены сохранившимися на вершинах своих гнезд, вероятно, высиживающими птичьи яйца. [269] Соотношение между объемом яиц и массой тела взрослых особей этих динозавров позволяет предположить, что яйца в основном высиживал самец, а молодняк был очень скороспелым, как и многие современные наземные птицы. [270]
Известно, что некоторые динозавры использовали желудочные камни, как современные птицы. Животные проглатывают эти камни, чтобы улучшить пищеварение и расщепить пищу и твердые волокна, когда они попадают в желудок. Камни желудка, обнаруженные в сочетании с окаменелостями, называются гастролитами. [271]
Вымирание основных групп
Все нептичьи динозавры и большинство линий птиц. [272] вымерли в результате массового вымирания , названного событием мел-палеогенового (K-Pg) вымирания , в конце мелового периода. Над границей мела и палеогена , возраст которой датируется 66,038 ± 0,025 миллиона лет назад, [273] окаменелости нептичьих динозавров внезапно исчезают; отсутствие окаменелостей динозавров исторически использовалось для отнесения пород к следующему кайнозою. Природа события, вызвавшего это массовое вымирание, тщательно изучалась с 1970-х годов, что привело к развитию двух механизмов, которые, как полагают, сыграли важную роль: событие внеземного воздействия на полуострове Юкатан и наводнение базальтового вулканизма в Индии. . Однако конкретные механизмы вымирания и степень его воздействия на динозавров все еще остаются областями продолжающихся исследований. [274] динозаврами вымерли многие другие группы животных: птерозавры, морские рептилии, такие как мозазавры и плезиозавры, несколько групп млекопитающих, аммониты ( наутилусоподобные Наряду с моллюски ), рудисты ( рифообразующие двустворчатые моллюски ) и различные группы морского планктона. [275] [276] В целом примерно 47% родов и 76% видов на Земле вымерли во время вымирания K-Pg. [277] Относительно большой размер большинства динозавров и низкое разнообразие видов динозавров с мелким телом в конце мелового периода, возможно, способствовали их исчезновению; [278] Вымирание линий птиц, которые не выжили, также могло быть вызвано зависимостью от лесной среды обитания или отсутствием приспособлений к поеданию семян для выживания. [279] [280]
Разнообразие до вымирания
Незадолго до вымирания K-Pg число нептичьих видов динозавров, существовавших во всем мире, оценивалось от 628 до 1078. [281] Остается неясным, происходило ли постепенное сокращение разнообразия динозавров до вымирания K-Pg или динозавры действительно процветали до вымирания. скальные образования маастрихтской Было обнаружено, что эпохи, которая непосредственно предшествовала вымиранию, имели меньшее разнообразие, чем предыдущая кампанская эпоха, что привело к преобладающему мнению о долгосрочном снижении разнообразия. [275] [276] [282] Однако эти сравнения не учитывали ни различный потенциал сохранности между горными породами, ни различную степень разведки и раскопок. [274] В 1984 году Дейл Рассел провел анализ, объясняющий эти предубеждения, и не обнаружил никаких свидетельств снижения; [283] другой анализ, проведенный Дэвидом Фастовским и его коллегами в 2004 году, даже показал, что разнообразие динозавров постоянно увеличивалось вплоть до их исчезновения. [284] но этот анализ был опровергнут. [285] С тех пор различные подходы, основанные на статистике и математических моделях, по-разному поддерживали либо внезапное исчезновение [274] [281] [286] или постепенное снижение. [287] [288] Тенденции разнообразия в конце мелового периода могли различаться в зависимости от линии динозавров: было высказано предположение, что зауроподы не находились в упадке, в то время как птицетазовые и тероподы находились в упадке. [289] [290]
Ударное событие


Гипотеза о столкновении с болидом , впервые привлеченная к широкому вниманию в 1980 году Уолтером Альваресом , Луисом Альваресом и его коллегами, приписывает событие вымирания K-Pg удару болида (внеземного снаряда). [291] Альварес и его коллеги предположили, что внезапное повышение уровня иридия , зафиксированное по всему миру в горных отложениях на границе мела и палеогена, было прямым свидетельством этого воздействия. [292] Ударный кварц , свидетельствующий о сильной ударной волне, исходящей от удара, также был обнаружен по всему миру. [293] Фактическое место удара оставалось неуловимым до тех пор, пока не был обнаружен кратер шириной 180 км (110 миль) на полуострове Юкатан на юго-востоке Мексики , который был опубликован в статье 1991 года Алана Хильдебранда и его коллег. [294] Теперь большая часть доказательств свидетельствует о том, что болид Ширина от 5 до 15 километров (от 3 до мили ) ударила по 9 + 1 ⁄ полуострову Юкатан 66 миллионов лет назад, образовав этот кратер. [295] и создание «механизма уничтожения», который вызвал событие исчезновения. [296] [297] [298]
В течение нескольких часов удар Чиксулуб вызвал бы немедленные последствия, такие как землетрясения, [299] цунами, [300] и глобальная огненная буря, которая, вероятно, привела к гибели бездомных животных и вызвала лесные пожары. [301] [302] Однако это также имело бы долгосрочные последствия для окружающей среды. В течение нескольких дней сульфатные аэрозоли, выброшенные из скал на месте удара, могли способствовать возникновению кислотных дождей и закислению океана . [303] [304] Считается, что в последующие месяцы и годы аэрозоли сажи распространились по всему миру; они бы охладили поверхность Земли, отражая тепловое излучение , и значительно замедлили бы фотосинтез, блокируя солнечный свет, тем самым создав ударную зиму . [274] [305] [306] (Эту роль приписывали сульфатным аэрозолям, пока эксперименты не доказали обратное. [304] ) Прекращение фотосинтеза привело бы к распаду пищевых сетей, зависящих от листовых растений, в которые входили все динозавры, за исключением зерноядных птиц. [280]
Деканские ловушки
Во время вымирания K-Pg базальты Декана в Индии активно извергались. Извержения можно разделить на три фазы вокруг границы K-Pg: две до границы и одну после. Вторая фаза, которая произошла очень близко к границе, должна была вытеснить от 70 до 80% объема этих извержений в виде прерывистых импульсов, произошедших с интервалом около 100 000 лет. [307] [308] парниковые газы, такие как углекислый газ и диоксид серы . В результате этой вулканической активности были бы выделены [309] [310] что приводит к изменению климата из-за колебаний температуры примерно на 3 ° C (5,4 ° F), но, возможно, до 7 ° C (13 ° F). [311] Как и в случае с Чиксулубом, извержения могли также привести к выбросу сульфатных аэрозолей, которые могли вызвать кислотные дожди и глобальное похолодание. [312] Однако из-за большой погрешности датировки извержений роль Деканских траппов в вымирании K-Pg остается неясной. [273] [274] [313]
До 2000 года аргументы о том, что извержения Деканских траппов - в отличие от воздействия Чиксулуб - вызвали вымирание, обычно были связаны с мнением о том, что вымирание было постепенным. До открытия кратера Чиксулуб Деканские ловушки использовались для объяснения глобального слоя иридия; [309] [314] даже после открытия кратера все еще считалось, что его воздействие оказало лишь региональное, а не глобальное влияние на событие вымирания. [315] В ответ Луис Альварес отверг вулканическую активность как объяснение слоя иридия и вымирания в целом. [316] Однако с тех пор большинство исследователей заняли более умеренную позицию, которая определяет воздействие Чиксулуб как основную причину вымирания, а также признает, что ловушки Декана также могли сыграть свою роль. Сам Уолтер Альварес признал, что Деканские ловушки и другие экологические факторы, возможно, способствовали вымиранию в дополнение к воздействию Чиксулуба. [317] По некоторым оценкам, начало второй фазы извержений Деканских траппов произошло через 50 000 лет после удара Чиксулуб. [318] В сочетании с математическим моделированием сейсмических волн , которые могли возникнуть в результате удара, это привело к предположению, что удар Чиксулуб мог спровоцировать эти извержения за счет увеличения проницаемости мантийного шлейфа, лежащего под Деканскими траппами. [319] [320]
Остается неясным, были ли Деканские ловушки основной причиной вымирания, наряду с воздействием Чиксулуб. Сторонники считают, что климатическое воздействие выброса диоксида серы было на одном уровне с воздействием Чиксулуб, а также отмечают роль наводнения базальтового вулканизма в других массовых вымираниях, таких как пермско-триасовое вымирание . [321] [322] Они считают, что воздействие Чиксулуб усугубило продолжающееся изменение климата, вызванное извержениями. [323] Между тем, недоброжелатели отмечают внезапный характер вымирания и то, что другие импульсы активности Деканских ловушек сопоставимой величины, по-видимому, не вызвали вымирания. Они также утверждают, что причины различных массовых вымираний следует оценивать отдельно. [324] В 2020 году Альфио Кьяренца и его коллеги предположили, что ловушки Декана, возможно, даже имели противоположный эффект: они предположили, что долгосрочное потепление, вызванное выбросами углекислого газа, могло смягчить зиму, вызванную воздействием Чиксулуб. [298]
Возможные выжившие в палеоцене
Останки нептичьих динозавров время от времени находили выше границы K-Pg. В 2000 году Спенсер Лукас и его коллеги сообщили об открытии единственной правой бедренной кости гадрозавра в бассейне Сан-Хуан в Нью-Мексико и описали это как свидетельство существования динозавров палеоцена. Каменная толща, в которой была обнаружена кость, датируется эпохой раннего палеоцена , примерно 64,8 миллиона лет назад. [325] Если бы кость не была повторно отложена в результате выветривания, это стало бы доказательством того, что некоторые популяции динозавров выжили как минимум полмиллиона лет назад в кайнозойскую эпоху. [326] Другие свидетельства включают присутствие останков динозавров в формации Хелл-Крик на высоте до 1,3 м (4,3 фута) над границей мела и палеогена, что соответствует 40 000 лет прошедшего времени. Это использовалось для подтверждения мнения о том, что вымирание K-Pg было постепенным. [327] Однако многие другие исследователи считают, что эти предполагаемые палеоценовые динозавры были переработаны , то есть вымыты из своих первоначальных мест, а затем перезахоронены в более молодых отложениях. [328] [329] [330] Оценки возраста также были сочтены ненадежными. [331]
Культурные изображения

По человеческим меркам динозавры были существами фантастической внешности и зачастую огромных размеров. Таким образом, они захватили воображение людей и стали неотъемлемой частью человеческой культуры. Вхождение слова «динозавр» в обиход отражает культурную значимость животных: в английском языке «динозавр» обычно используется для описания всего, что непрактично велико, устарело или находится на грани исчезновения. [332]
Общественный энтузиазм по поводу динозавров впервые возник в викторианской Англии, где в 1854 году, спустя три десятилетия после первых научных описаний останков динозавров, зверинец из реалистичных скульптур динозавров был открыт в лондонском парке Кристал Пэлас . Динозавры Хрустального дворца оказались настолько популярными, что вскоре возник сильный рынок более мелких копий. В последующие десятилетия выставки динозавров открылись в парках и музеях по всему миру, гарантируя, что последующие поколения будут знакомиться с животными в захватывающей и захватывающей форме. [333] Непреходящая популярность динозавров, в свою очередь, привела к значительному государственному финансированию науки о динозаврах и часто стимулировала новые открытия. В Соединенных Штатах, например, конкуренция между музеями за внимание общественности привела непосредственно к Костяным войнам 1880-х и 1890-х годов, во время которых пара враждующих палеонтологов внесла огромный научный вклад. [334]
Популярная озабоченность динозаврами обеспечила их появление в литературе , кино и других средствах массовой информации . Начиная с 1852 года с мимолетного упоминания в « Чарльза Диккенса » Холодном доме , [335] динозавры фигурировали в большом количестве художественных произведений. Роман Жюля Верна 1864 года « Путешествие к центру Земли» , книга сэра Артура Конан Дойля 1912 года «Затерянный мир» , анимационный фильм 1914 года « Динозавр Герти » (с участием первого анимированного динозавра), культовый фильм 1933 года «Кинг-Конг» , фильм 1954 года. «Годзилла» и его многочисленные продолжения, бестселлер 1990 года « Парк Юрского периода» Майкла Крайтона и его экранизация 1993 года — лишь несколько ярких примеров появления динозавров в художественной литературе. Авторы научно-популярных работ о динозаврах, в том числе некоторые видные палеонтологи, часто пытались использовать животных как способ познакомить читателей с наукой в целом. Динозавры повсеместно используются в рекламе ; Многие компании упоминали динозавров в печатной или телевизионной рекламе либо для того, чтобы продавать свою продукцию, либо для того, чтобы охарактеризовать своих конкурентов как медлительных, тупых или устаревших. [336] [337]
См. также
- Диета и кормление динозавров
- Эволюционная история жизни
- Списки стратиграфических единиц, содержащих динозавров
- Список родов динозавров
- Список родов птиц
- Список птиц
- Список динозавров с неофициальным названием
- Список фильмов с участием динозавров
Примечания
- ^ Динозавры (включая птиц) относятся к естественной группе Reptilia . Их биология не совсем соответствует устаревшему классу Reptilia систематики Линнея , состоящему из хладнокровных амниот без меха и перьев. Поскольку таксономия Линнея была сформулирована для современных животных до изучения эволюции и палеонтологии, она не учитывает вымерших животных с промежуточными признаками между традиционными классами.
Ссылки
- ^ Мэтью Г. Барон; Меган Э. Уильямс (2018). «Переоценка загадочного динозаврообразного Caseosaurus crosbyensis из позднего триаса Техаса, США, и его значение для ранней эволюции динозавров». Acta Palaeontologica Polonica. 63. два : 10.4202/прил.00372.2017 .
- ^ Андреа Кау (2018). «Сборка плана тела птиц: процесс продолжительностью 160 миллионов лет» (PDF) . Бюллетень Итальянского палеонтологического общества. 57(1):1–25. два : 10.4435/BSPI.2018.01 .
- ^ Фериголо, Хорхе; Лангер, Макс К. (2007). «Динозоформ позднего триаса из южной Бразилии и происхождение птицетазовой предзубчатой кости». Историческая биология . 19 (1): 23–33. Бибкод : 2007HBio...19...23F . дои : 10.1080/08912960600845767 . ISSN 0891-2963 . S2CID 85819339 .
- ^ Лангер, Макс К.; Фериголо, Хорхе (2013). «Позднетриасовый динозавроморф Sacisaurus agudoensis (формация Катуррита; Риу-Гранди-ду-Сул, Бразилия): анатомия и сходство» . Геологическое общество, Лондон, специальные публикации . 379 (1): 353–392. Бибкод : 2013GSLSP.379..353L . дои : 10.1144/SP379.16 . ISSN 0305-8719 . S2CID 131414332 .
- ^ Кабрейра, Сан-Франциско; Келлнер, AWA; Диаш-да-Сильва, С.; да Силва, ЛР; Бронзати, М.; де Алмейда Марсола, JC; Мюллер, RT; де Соуза Биттенкур, Ж.; Батиста, Би Джей; Раугуст, Т.; Каррильо, Р.; Бродт, А.; Лангер, MC (2016). «Уникальное собрание динозавров позднего триаса раскрывает анатомию и диету предков динозавров» . Современная биология . 26 (22): 3090–3095. Бибкод : 2016CBio...26.3090C . дои : 10.1016/j.cub.2016.09.040 . ПМИД 27839975 .
- ^ Мюллер, Родриго Темп; Гарсия, Маурисио Силва (26 августа 2020 г.). «Парафилетические 'Silesauridae' как альтернативная гипотеза первоначальной радиации птицетазовых динозавров» . Письма по биологии . 16 (8): 20200417. doi : 10.1098/rsbl.2020.0417 . ПМК 7480155 . ПМИД 32842895 .
- ↑ Перейти обратно: Перейти обратно: а б с «Рождение» динозавров» . Больше, чем Додо . 28 апреля 2017 г. Проверено 15 марта 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б с «Рождение динозавров: Ричард Оуэн и динозаврозаврия» . Библиотека наследия биоразнообразия . 16 октября 2015 года . Проверено 15 марта 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Бретт-Сурман, депутат Кнессета; Хольц, Томас Р.; Фарлоу, Джеймс О. (27 июня 2012 г.). Полный динозавр . Издательство Университета Индианы. п. 25. ISBN 978-0-253-00849-7 .
- ↑ Перейти обратно: Перейти обратно: а б с д Weishampel, Dodson & Osmólska 2004 , стр. 7–19, гл. 1: «Происхождение и родство динозавров» Майкла Дж. Бентона .
- ^ Ольшевский 2000.
- ↑ Перейти обратно: Перейти обратно: а б Лангер, Макс К.; Эскурра, Мартин Д .; Биттенкорт, Джонатас С.; Новас, Фернандо Э. (февраль 2010 г.). «Происхождение и ранняя эволюция динозавров». Биологические обзоры . 85 (1). Кембридж : Кембриджское философское общество : 65–66, 82. doi : 10.1111/j.1469-185x.2009.00094.x . hdl : 11336/103412 . ISSN 1464-7931 . ПМИД 19895605 . S2CID 34530296 .
- ^ «Использование дерева для классификации» . Понимание эволюции . Беркли : Калифорнийский университет . Архивировано из оригинала 31 августа 2019 года . Проверено 14 октября 2019 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Weishampel, Dodson & Osmólska 2004 , стр. 210–231, гл. 11: «Basal Avialae» Кевина Падиана .
- ^ Уэйд, Николас (22 марта 2017 г.). «Встряхивание генеалогического древа динозавров» . Нью-Йорк Таймс . Нью-Йорк . ISSN 0362-4331 . Архивировано из оригинала 7 апреля 2018 года . Проверено 30 октября 2019 г. «Версия этой статьи появится в печати 28 марта 2017 года на странице D6 нью-йоркского издания под заголовком: «Встряхивание генеалогического древа динозавров».
- ^ Барон, Мэтью Г.; Норман, Дэвид Б .; Барретт, Пол М. (2017). «Новая гипотеза взаимоотношений динозавров и ранней эволюции динозавров». Природа . 543 (7646). Лондон: Исследования природы : 501–506. Бибкод : 2017Natur.543..501B . дои : 10.1038/nature21700 . ISSN 0028-0836 . ПМИД 28332513 . S2CID 205254710 . «Этот файл содержит дополнительный текст и данные, дополнительные таблицы 1–3 и дополнительные ссылки».: Дополнительная информация [ постоянная мертвая ссылка ]
- ^ Перенасыщение 1997 , с. 40
- ^ Чамары, СП (30 сентября 2014 г.). «Динозавры, птерозавры и другие заавры – большие различия» . Форбс . Джерси-Сити, Нью-Джерси . ISSN 0015-6914 . Архивировано из оригинала 10 ноября 2014 года . Проверено 2 октября 2018 г.
- ^ Ламберт и The Diagram Group 1990 , стр. 288
- ^ Фарлоу и Бретт-Сурман 1997 , стр. 607–624 , гл. 39: «Основные группы нединозавровых позвоночных мезозойской эры», Майкл Моралес.
- ↑ Перейти обратно: Перейти обратно: а б Теннант, Джонатан П.; Кьяренца, Альфио Алессандро; Барон Мэтью (19 февраля 2018 г.). «Как наши знания о разнообразии динозавров в геологическом времени изменились в истории исследований?» . ПерДж . 6 : е4417. дои : 10.7717/peerj.4417 . ПМЦ 5822849 . ПМИД 29479504 .
- ^ Старрфельт, Йостейн; Лиоу, Ли Сян (2016). «Сколько видов динозавров существовало? Смещение окаменелостей и истинное богатство оценивались с использованием модели выборки Пуассона» . Философские труды Королевского общества B: Биологические науки . 371 (1691): 20150219. doi : 10.1098/rstb.2015.0219 . ПМЦ 4810813 . ПМИД 26977060 .
- ^ Ван, Стив С.; Додсон, Питер (2006). «Оценка разнообразия динозавров» . Учеб. Натл. акад. наук. США 103 (37). Вашингтон, округ Колумбия : Национальная академия наук : 13601–13605. Бибкод : 2006PNAS..10313601W . дои : 10.1073/pnas.0606028103 . ISSN 0027-8424 . ПМК 1564218 . ПМИД 16954187 .
- ^ Рассел, Дейл А. (1995). «Китай и затерянные миры эпохи динозавров». Историческая биология . 10 (1). Милтон-Парк , Оксфордшир : Тейлор и Фрэнсис : 3–12. Бибкод : 1995HBio...10....3R . дои : 10.1080/10292389509380510 . ISSN 0891-2963 .
- ^ Старрфельт, Йостейн; Лиоу, Ли Сян (2016). «Сколько видов динозавров существовало? Смещение окаменелостей и истинное богатство оценивались с использованием модели выборки Пуассона» . Философские труды Королевского общества Б. 371 (1691). Лондон: Королевское общество : 20150219. doi : 10.1098/rstb.2015.0219 . ISSN 0962-8436 . ПМЦ 4810813 . ПМИД 26977060 .
- ^ Блэк, Райли (23 марта 2016 г.). «Большинство видов динозавров до сих пор не открыты» . Национальные географические новости . Архивировано из оригинала 6 марта 2021 года . Проверено 6 июня 2021 г.
- ^ Гилл, Ф.; Донскер, Д.; Расмуссен, П. (2021). "Добро пожаловать" . Всемирный список птиц МОК 11.1 .
- ^ Маклауд, Норман; Роусон, Питер Ф.; Фори, Питер Л.; и др. (1997). «Мел-третичный биотический переход». Журнал Геологического общества . 154 (2). Лондон: Лондонское геологическое общество : 265–292. Бибкод : 1997JGSoc.154..265M . дои : 10.1144/gsjgs.154.2.0265 . ISSN 0016-7649 . S2CID 129654916 .
- ↑ Перейти обратно: Перейти обратно: а б Амио, Ромен; Бюффо, Эрик ; Лекюйер, Кристоф; и др. (2010). «Изотопы кислорода свидетельствуют о полуводном образе жизни спинозавридных теропод». Геология . 38 (2). Боулдер, Колорадо : Геологическое общество Америки : 139–142. Бибкод : 2010Geo....38..139A . дои : 10.1130/G30402.1 . ISSN 0091-7613 .
- ↑ Перейти обратно: Перейти обратно: а б с Брусатте 2012 , стр. 9–20, 21
- ^ Несбитт, Стерлинг Дж. (2011). «Ранняя эволюция архозавров: взаимоотношения и происхождение основных клад» . Бюллетень Американского музея естественной истории . 2011 (352). Нью-Йорк: Американский музей естественной истории : 1–292. дои : 10.1206/352.1 . hdl : 2246/6112 . ISSN 0003-0090 . S2CID 83493714 .
- ↑ Перейти обратно: Перейти обратно: а б Пол 2000 , стр. 140–168, гл. 3: «Классификация и эволюция групп динозавров» Томаса Р. Хольца-младшего.
- ^ Смит, Дэйв; и др. «Динозаврия: Морфология» . Беркли: Музей палеонтологии Калифорнийского университета . Проверено 16 октября 2019 г.
- ^ Лангер, Макс К.; Абдала, Фернандо; Рихтер, Марта; Бентон, Майкл Дж. (1999). «Динозавр-зауроподоморф из верхнего триаса (карнского периода) на юге Бразилии». Доклады Академии наук, серия IIA . 329 (7). Амстердам : Elsevier от имени Французской академии наук : 511–517. Бибкод : 1999CRASE.329..511L . дои : 10.1016/S1251-8050(00)80025-7 . ISSN 1251-8050 .
- ^ Несбитт, Стерлинг Дж.; Ирмис, Рэндалл Б.; Паркер, Уильям Г. (2007). «Критическая переоценка таксонов динозавров позднего триаса Северной Америки». Журнал систематической палеонтологии . 5 (2). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис от имени Музея естественной истории, Лондон : 209–243. Бибкод : 2007JSPal...5..209N . дои : 10.1017/S1477201907002040 . ISSN 1477-2019 . S2CID 28782207 .
- ^ Это было признано не позднее 1909 года: Селески, Мэтт (2005). «Доктор У. Дж. Холланд и расползающиеся зауроподы» . Волосатый музей естественной истории . Архивировано из оригинала 12 июня 2011 года . Проверено 18 октября 2019 г.
- Холланд, Уильям Дж. (май 1910 г.). «Обзор некоторых недавних критических замечаний по поводу реставраций динозавров-зауроподов, существующих в музеях Соединенных Штатов, с особым упором на реставрацию диплодока Карнеги в Музее Карнеги» . Американский натуралист . 44 (521). Американское общество натуралистов : 259–283. дои : 10.1086/279138 . ISSN 0003-0147 . S2CID 84424110 . Проверено 18 октября 2019 г.
- Аргументы и многие изображения также представлены в Desmond 1975 .
- ↑ Перейти обратно: Перейти обратно: а б Бентон 2005 г.
- ^ Коуэн 2005 , стр. 151–175, гл. 12: «Динозавры».
- ↑ Перейти обратно: Перейти обратно: а б Кубо, Тай; Бентон, Майкл Дж. (ноябрь 2007 г.). «Эволюция позы задних конечностей у архозавров: нагрузки на конечности у вымерших позвоночных» (PDF) . Палеонтология . 50 (6). Хобокен, Нью-Джерси : Уайли-Блэквелл : 1519–1529 гг. Бибкод : 2007Palgy..50.1519K . дои : 10.1111/j.1475-4983.2007.00723.x . ISSN 0031-0239 . S2CID 140698705 .
- ^ Донг 1992
- ^ «Кости динозавров «используются как лекарство» » . Новости Би-би-си . Лондон: Би-би-си. 6 июля 2007 г. Архивировано из оригинала 27 августа 2019 г. Проверено 4 ноября 2019 г.
- ^ Пол 2000 , стр. 10–44, гл. 1: «Краткая история палеонтологии динозавров» Майкла Дж. Бентона.
- ↑ Перейти обратно: Перейти обратно: а б Фарлоу и Бретт-Сурман 1997 , стр. 3–11 , гл. 1: «Самые ранние открытия» Уильяма А.С. Сарджанта .
- ^ Сюжет 1677 , стр. 131–139, илл. опп. п. 142, рис. 4
- ^ Сюжет 1677 , с. [1]
- ^ «Сюжет Роберта» (PDF) . Узнать больше . Оксфорд: Музей естественной истории Оксфордского университета . 2006. Архивировано из оригинала (PDF) 1 октября 2006 года . Проверено 14 ноября 2019 г.
- ^ Люйд 1699 , с. 67
- ^ Делар, Джастин Б.; Сарджант, Уильям А.С. (2002). «Самые ранние открытия динозавров: пересмотренные записи». Труды Ассоциации геологов . 113 (3). Амстердам: Elsevier от имени Ассоциации геологов : 185–197. Бибкод : 2002ПрГА..113..185Д . дои : 10.1016/S0016-7878(02)80022-0 . ISSN 0016-7878 .
- ^ Гюнтер 1968
- ^ Бакленд, Уильям (1824). «Объявление о мегалозавре или огромной ископаемой ящерице из Стоунсфилда» . Труды Лондонского геологического общества . 1 (2). Лондон: Лондонское геологическое общество: 390–396. дои : 10.1144/transgslb.1.2.390 . ISSN 2042-5295 . S2CID 129920045 . Архивировано (PDF) из оригинала 21 октября 2019 г. Проверено 5 ноября 2019 г.
- ^ Мантелл, Гидеон А. (1825). «Обратите внимание на игуанодона , недавно обнаруженную ископаемую рептилию, найденную в песчанике леса Тилгейт в Сассексе» . Философские труды Лондонского королевского общества . 115 . Лондон: Королевское общество: 179–186. Бибкод : 1825RSPT..115..179M . дои : 10.1098/rstl.1825.0010 . ISSN 0261-0523 . JSTOR 107739 .
- ^ Фарлоу и Бретт-Сурман 1997 , стр. 14 , гл. 2: «Европейские охотники на динозавров» Ханса-Дитера Сьюса.
- ↑ Перейти обратно: Перейти обратно: а б Оуэн 1842 , с. 103 : «Сочетание таких признаков... предполагается, будет считаться достаточным основанием для создания отдельного племени или подотряда ящерообразных рептилий, для которых я бы предложил название Dinosauria *. (*греч. δεινός) , страшно великий; σαύρος , ящерица...)
- ^ «Динозаврия» . Словарь Merriam-Webster.com . Проверено 10 ноября 2019 г.
- ^ Крейн, Джордж Р. (ред.). «Результаты поиска по заголовку греческого словаря» . Персей 4.0 . Медфорд и Сомервилл, Массачусетс : Университет Тафтса . Проверено 13 октября 2019 г. Лемма для « δεινός » из Генри Джорджа Лидделла , Роберта Скотта , Греко-английский лексикон (1940): «страшный, ужасный».
- ^ Фарлоу и Бретт-Сурман 1997 , стр. ix–xi , Предисловие, «Динозавры: земная превосходная степень» Джеймса О. Фарлоу и М.К. Бретта-Сурмана.
- ^ Рупке 1994
- ^ Прието-Маркес, Альберт; Вейшампель, Дэвид Б.; Хорнер, Джон Р. (март 2006 г.). «Динозавр Hadrosaurus foulkii из кампана восточного побережья Северной Америки, с переоценкой рода» (PDF) . Acta Palaeontologica Polonica . 51 (1). Варшава: Институт палеобиологии Польской академии наук: 77–98. ISSN 0567-7920 . Архивировано (PDF) из оригинала 22 июня 2019 г. Проверено 5 ноября 2019 г.
- ^ Холмс 1998 г.
- ↑ Перейти обратно: Перейти обратно: а б с Тейлор, член парламента (2010). «Исследование динозавров зауроподов: исторический обзор». Геологическое общество, Лондон, специальные публикации . 343 (1): 361–386. Бибкод : 2010GSLSP.343..361T . дои : 10.1144/SP343.22 . S2CID 910635 .
- ^ Нэйш, Д. (2009). Великие открытия динозавров . Лондон, Великобритания: A&C Black Publishers Ltd., стр. 89–93. ISBN 978-1-4081-1906-8 .
- ^ Арбур, В. (2018). «Результаты возрождения динозавров». Наука . 360 (6389): 611. Бибкод : 2018Sci...360..611A . дои : 10.1126/science.aat0451 . S2CID 46887409 .
- ↑ Перейти обратно: Перейти обратно: а б Баккер, RT (1968). «Превосходство динозавров». Открытие: журнал Музея естественной истории Пибоди . 3 (2): 11–22. ISSN 0012-3625 . OCLC 297237777 .
- ↑ Перейти обратно: Перейти обратно: а б Баккер, RT (1972). «Анатомические и экологические доказательства эндотермии у динозавров». Природа . 238 (5359): 81–85. Бибкод : 1972Natur.238...81B . дои : 10.1038/238081a0 . S2CID 4176132 .
- ^ Баккер 1986
- ↑ Перейти обратно: Перейти обратно: а б Бентон, MJ (2008). «Качество ископаемых и наименование динозавров» . Письма по биологии . 4 (6): 729–732. дои : 10.1098/rsbl.2008.0402 . ПМК 2614166 . ПМИД 18796391 .
- ↑ Перейти обратно: Перейти обратно: а б Кэшмор, Д.Д.; Мэннион, Полиция; Апчерч, П.; Батлер, Р.Дж. (2020). «Еще десять лет открытий: пересмотр качества летописи окаменелостей динозавров-зауроподоморфов» . Палеонтология . 63 (6): 951–978. Бибкод : 2020Palgy..63..951C . дои : 10.1111/пала.12496 . S2CID 219090716 .
- ^ Кэшмор, Д.Д.; Батлер, Р.Дж. (2019). «Скелетная полнота летописи окаменелостей нептичьих тероподных динозавров» . Палеонтология . 62 (6): 951–981. Бибкод : 2019Palgy..62..951C . дои : 10.1111/пала.12436 . S2CID 197571209 .
- ^ Хольц, Т.Р. младший; Бретт-Сурман, МК (1997). «Таксономия и систематика динозавров» . Полный динозавр . Блумингтон: Издательство Университета Индианы. стр. 209–223. ISBN 978-0-253-33349-0 .
- ↑ Перейти обратно: Перейти обратно: а б с Сен-Флер, Николас (8 декабря 2016 г.). «Эта штука с перьями, застрявшими в янтаре? Это был хвост динозавра» . Трилобиты. Нью-Йорк Таймс . Нью-Йорк. ISSN 0362-4331 . Архивировано из оригинала 31 августа 2017 года . Проверено 8 декабря 2016 г.
- ^ Локли, МГ; Райт, Дж.Л. (2000). «Чтение о динозаврах - аннотированная библиография книг». Журнал геонаучного образования . 48 (2): 167–178. Бибкод : 2000JGeEd..48..167L . дои : 10.5408/1089-9995-48.2.167 . S2CID 151426669 .
- ^ Ллойд, GT; Дэвис, Кентукки; Пизани, Д.; Тарвер, Дж. Э.; Рута, Р.; Сакамото, М.; Хон, DWE; Дженнингс, Р.; Бентон, MJ (2008). «Динозавры и меловая земная революция» . Труды Королевского общества Б. 275 (1650): 2483–2490. дои : 10.1098/rspb.2008.0715 . ПМК 2603200 . ПМИД 18647715 .
- ↑ Перейти обратно: Перейти обратно: а б Швейцер, МХ (2011). «Сохранение мягких тканей у наземных мезозойских позвоночных». Ежегодный обзор наук о Земле и планетах . 39 : 187–216. Бибкод : 2011AREPS..39..187S . doi : 10.1146/annurev-earth-040610-133502 .
- ↑ Перейти обратно: Перейти обратно: а б Хули, RW (1917). «II — На покровах Iguanodon bernissartensis , Boulenger, и Morosaurus becklesii , Mantell» . Геологический журнал . 4 (4): 148–150. Бибкод : 1917GeoM....4..148H . дои : 10.1017/s0016756800192386 . S2CID 129640665 .
- ^ Осборн, HF (1912). «Покровы игуанодонтного динозавра Траходона ». Мемуары Американского музея естественной истории . 1 : 33–54.
- ^ Белл, PR (2014). «Обзор отпечатков кожи гадрозавров». В Эберте, Д.; Эванс, Д. (ред.). Гадрозавры: материалы Международного симпозиума по гадрозаврам . Блумингтон: Издательство Принстонского университета. стр. 572–590.
- ^ Элиасон, CM; Хадсон, Л.; Уоттс, Т.; Гарза, Х.; Кларк, Дж.А. (2017). «Исключительная сохранность и летопись окаменелостей покровов четвероногих» . Труды Королевского общества Б. 284 (1862): 1–10. дои : 10.1098/rspb.2017.0556 . ПМЦ 5597822 . ПМИД 28878057 .
- ^ Бентон, MJ (1998). «Окаменелости динозавров с мягкими частями» (PDF) . Тенденции в экологии и эволюции . 13 (8): 303–304. дои : 10.1016/s0169-5347(98)01420-7 . ПМИД 21238317 .
- ^ Чжоу, З.-Х.; Ван, Ю. (2017). «Ассамбляжи позвоночных юрской биоты Янляо и раннемеловой биоты Джехоль: сравнения и значения». Палеомир . 26 (2): 241–252. дои : 10.1016/j.palwor.2017.01.002 .
- ^ Норелл, Массачусетс; Сюй, X. (2005). «Пернатые динозавры». Ежегодный обзор наук о Земле и планетах . 33 : 277–299. Бибкод : 2005AREPS..33..277N . doi : 10.1146/annurev.earth.33.092203.122511 .
- ↑ Перейти обратно: Перейти обратно: а б Рой, А.; Питтман, М.; Сайтта, ET; Кэй, Т.Г.; Сюй, X. (2020). «Последние достижения в реконструкции палеоцвета амниот и основы для будущих исследований» . Биологические обзоры . 95 (1): 22–50. дои : 10.1111/brv.12552 . ПМК 7004074 . ПМИД 31538399 .
- ^ Винтер, Дж. (2020). «Реконструкция палеоколора позвоночных» . Ежегодный обзор наук о Земле и планетах . 48 : 345–375. Бибкод : 2020AREPS..48..345В . doi : 10.1146/annurev-earth-073019-045641 . S2CID 219768255 .
- ^ Чжан, Ф.; Кернс, СЛ; Орр, ПиДжей; Бентон, MJ; Чжоу, З.; Джонсон, Д.; Сюй, Х.; Ван, X. (2010). «Ископаемые меланосомы и цвет меловых динозавров и птиц» (PDF) . Природа . 463 (7284): 1075–1078. Бибкод : 2010Natur.463.1075Z . дои : 10.1038/nature08740 . ПМИД 20107440 . S2CID 205219587 .
- ^ Смитвик, FM; Николлс, Р.; Катхилл, IC; Винтер, Дж. (2017). «Затенение и полосы у теропод-динозавра Sinosauropteryx свидетельствуют о гетерогенных средах обитания в биоте Джехола раннего мела» . Современная биология . 27 (21): 3337–3343.e2. Бибкод : 2017CBio...27E3337S . дои : 10.1016/j.cub.2017.09.032 . hdl : 1983/8ee95b15-5793-42ad-8e57-da6524635349 . ПМИД 29107548 .
- ^ Винтер, Дж.; Николлс, Р.; Лаутеншлагер, С.; Питтман, М.; Кэй, Т.Г.; Рэйфилд, Э.; Майр, Г.; Катхилл, IC (2016). «3D-камуфляж у птицетазового динозавра» . Современная биология . 26 (18): 2456–2462. Бибкод : 2016CBio...26.2456V . дои : 10.1016/j.cub.2016.06.065 . ПМК 5049543 . ПМИД 27641767 .
- ^ Линдгрен, Дж.; Мойер, А.; Швейцер, Миннесота; Шёвалл, П.; Увдал, П.; Нильссон, Делавэр; Хеймдал, Дж.; Энгдаль, А.; Грен, Дж.А.; Шульц, БП; Кир, BP (2015). «Интерпретация окраски на основе меланина во времени: критический обзор» . Труды Королевского общества Б. 282 (1813): 20150614. doi : 10.1098/rspb.2015.0614 . ПМЦ 4632609 . ПМИД 26290071 .
- ^ Швейцер, Миннесота; Линдгрен, Дж.; Мойер, А.Е. (2015). «Пересмотр меланосом и древней окраски: ответ на Vinther 2015 (DOI 10.1002/bies.201500018)». Биоэссе . 37 (11): 1174–1183. doi : 10.1002/bies.201500061 . ПМИД 26434749 . S2CID 45178498 .
- ^ Чжоу, З. (2014). «Биота Джехол, наземный лагерштетт раннего мела: новые открытия и последствия» . Национальный научный обзор . 1 (4): 543–559. дои : 10.1093/nsr/nwu055 .
- ^ О'Коннор, Дж. К.; Чжоу, З. (2019). «Эволюция пищеварительной системы современных птиц: выводы из паравианских окаменелостей из биоты Яньляо и Джехол». Палеонтология . 63 (1): 13–27. дои : 10.1111/пала.12453 . S2CID 210265348 .
- ↑ Перейти обратно: Перейти обратно: а б Даль Сассо, Криштиану ; Синьор, Марко (26 марта 1998 г.). «Исключительная сохранность мягких тканей динозавра-теропода из Италии» (PDF) . Природа . 392 (6674). Лондон: Исследования природы: 383–387. Бибкод : 1998Natur.392..383D . дои : 10.1038/32884 . ISSN 0028-0836 . S2CID 4325093 . Архивировано (PDF) из оригинала 20 сентября 2016 г.
- ^ Морелл, В. (1993). «ДНК динозавра: Охота и шумиха». Наука . 261 (5118): 160–162. Бибкод : 1993Sci...261..160M . дои : 10.1126/science.8327889 . ПМИД 8327889 .
- ^ Павлицкий, Р.; Корбель, А.; Кубяк, Х. (1996). «Клетки, коллагеновые фибриллы и сосуды в костях динозавров». Природа . 211 (5049): 655–657. дои : 10.1038/211655a0 . ПМИД 5968744 . S2CID 4181847 .
- ^ Павлицкий, Р.; Новогродская-Загурская, М. (1998). «Кровеносные сосуды и эритроциты, сохранившиеся в костях динозавров». Анналы анатомии – Anatomischer Anzeiger . 180 (1): 73–77. дои : 10.1016/S0940-9602(98)80140-4 . ПМИД 9488909 .
- ↑ Перейти обратно: Перейти обратно: а б Швейцер, Мэри Х .; Виттмайер, Дженнифер Л.; Хорнер, Джон Р .; Топорски, Ян К. (2005). «Сосуды мягких тканей и сохранение клеток у тираннозавра рекса ». Наука . 307 (5717). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1952–1955. Бибкод : 2005Sci...307.1952S . дои : 10.1126/science.1108397 . ISSN 0036-8075 . ПМИД 15790853 . S2CID 30456613 .
- ^ Андерсон, Луизиана (2023). «Химическая основа сохранения ископаемых клеток и мягких тканей позвоночных» . Обзоры наук о Земле . 240 : 104367. Бибкод : 2023ESRv..24004367A . doi : 10.1016/j.earscirev.2023.104367 . S2CID 257326012 .
- ↑ Перейти обратно: Перейти обратно: а б Швейцер, Миннесота; Чжэн, В.; Орган, CL; Авчи, Р.; Суо, З.; Фраймарк, LM; ЛеБлё, В.С.; Дункан, МБ; ван дер Хайден, MG; Невё, Ж.М.; Лейн, Вашингтон; Коттрелл, Дж. С.; Хорнер, младший; Кэнтли, ЖК; Каллури, Р.; Асара, Дж. М. (2009). «Биомолекулярная характеристика и белковые последовательности кампанского гадрозавра B. canadensis ». Наука . 324 (5927): 626–631. Бибкод : 2009Sci...324..626S . дои : 10.1126/science.1165069 . ПМИД 19407199 . S2CID 5358680 .
- ^ Орган, CL; Швейцер, Миннесота; Чжэн, В.; Фраймарк, LM; Кэнтли, ЖК; Асара, Дж. М. (2008). «Молекулярная филогенетика мастодонта и тираннозавра рекса ». Наука . 320 (5875): 499. Бибкод : 2008Sci...320..499O . дои : 10.1126/science.1154284 . ПМИД 18436782 . S2CID 24971064 .
- ^ Швейцер, Миннесота; Чжэн, В.; Клеланд, ТП; Берн, М. (2013). «Молекулярный анализ остеоцитов динозавров подтверждает наличие эндогенных молекул». Кость . 52 (1). Амстердам: Эльзевир: 414–423. дои : 10.1016/j.bone.2012.10.010 . ISSN 8756-3282 . ПМИД 23085295 .
- ^ Байёль, AM; Чжэн, В.; Хорнер, младший; Холл, Британская Колумбия; Холлидей, СМ; Швейцер, МГ (2020). «Свидетельства наличия белков, хромосом и химических маркеров ДНК в исключительно сохранившемся хряще динозавра» . Национальный научный обзор . 7 (4): 815–822. дои : 10.1093/nsr/nwz206 . ПМЦ 8289162 . ПМИД 34692099 .
- ^ Бертаццо, С.; Мейдмент, ЮАР ; Каллепитис, К.; и др. (2015). «Волокна и клеточные структуры сохранились в экземплярах динозавров возрастом 75 миллионов лет» . Природные коммуникации . 6 : 7352. Бибкод : 2015NatCo...6.7352B . дои : 10.1038/ncomms8352 . ISSN 2041-1723 . ПМЦ 4468865 . ПМИД 26056764 .
- ^ Кэй, Т.Г.; Гоглер, Г.; Савлович, З. (2008). «Мягкие ткани динозавров, интерпретируемые как бактериальные биопленки» . ПЛОС ОДИН . 3 (7): e2808. Бибкод : 2008PLoSO...3.2808K . дои : 10.1371/journal.pone.0002808 . ПМЦ 2483347 . ПМИД 18665236 .
- ^ Петерсон, Дж. Э.; Ленчевский, МЭ; Шерер, Р.П. (2010). «Влияние микробных биопленок на сохранность первичных мягких тканей у ископаемых и современных архозавров» . ПЛОС ОДИН . 5 (10): е13334. Бибкод : 2010PLoSO...513334P . дои : 10.1371/journal.pone.0013334 . ISSN 1932-6203 . ПМЦ 2953520 . ПМИД 20967227 .
- ^ Бакли, М.; Уорвуд, С.; ван Донген, Б.; Китченер, AC; Мэннинг, Польша (2017). «Химера ископаемого белка; трудности в различении последовательностей пептидов динозавров от современного перекрестного загрязнения» . Труды Королевского общества Б. 284 (1855). дои : 10.1098/rspb.2017.0544 . ПМЦ 5454271 . ПМИД 28566488 .
- ^ Камп, Ли Р .; Павлов, Александр; Артур, Майкл А. (2005). «Массовый выброс сероводорода на поверхность океана и в атмосферу в периоды океанической аноксии». Геология . 33 (5). Боулдер, Колорадо: Геологическое общество Америки: 397–400. Бибкод : 2005Geo....33..397K . дои : 10.1130/G21295.1 . ISSN 0091-7613 . S2CID 34821866 .
- ^ Таннер, Лоуренс Х.; Лукас, Спенсер Г .; Чепмен, Мэри Г. (март 2004 г.). «Оценка данных и причин вымираний в позднем триасе» (PDF) . Обзоры наук о Земле . 65 (1–2). Амстердам: Эльзевир: 103–139. Бибкод : 2004ESRv...65..103T . дои : 10.1016/S0012-8252(03)00082-5 . ISSN 0012-8252 . Архивировано из оригинала (PDF) 25 октября 2007 г. Проверено 22 октября 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б Гриффин, Коннектикут; Винд, Б.М.; Муньиква, Д.; Бродерик, Ти Джей; Зондо, М.; Толан, С.; Лангер, MC; Несбитт, С.Дж.; Тарувинга, HR (2022). «Самые старые динозавры Африки демонстрируют раннее подавление распространения динозавров». Природа . 609 (7926): 313–319. Бибкод : 2022Natur.609..313G . дои : 10.1038/s41586-022-05133-x . ISSN 0028-0836 . ПМИД 36045297 . S2CID 251977824 .
- ^ Дезохо, Дж.Б.; Фиорелли, Ле; Эскурра, доктор медицины; Мартинелли, АГ; Рамезани Дж.; Да Роза, ААС; Белен фон Бачко, М.; Химена Тротейн, М.; Монтефельтро, ФК; Эзпелета, М.; Лангер, MC (2020). «Формация Ишигуаласто позднего триаса в Серро-Лас-Лахас (Ла-Риоха, Аргентина): ископаемые четвероногие, хроностратиграфия высокого разрешения и корреляции фауны» . Научные отчеты . 10 (1): 12782. Бибкод : 2020NatSR..1012782D . дои : 10.1038/s41598-020-67854-1 . ПМЦ 7391656 . ПМИД 32728077 .
- ^ Алькобер, Оскар А.; Мартинес, Рикардо Н. (2010). «Новый герреразаврид (Dinosauria, Saurischia) из формации Ишигуаласто верхнего триаса на северо-западе Аргентины» . ЗооКлючи (63). София : Издательство Pensoft : 55–81. Бибкод : 2010ЗооК...63...55А . дои : 10.3897/zookeys.63.550 . ISSN 1313-2989 . ПМК 3088398 . ПМИД 21594020 .
- ↑ Перейти обратно: Перейти обратно: а б с Новас, FE; Агнолин, Флорида; Эскурра, доктор медицины; Мюллер, RT; Мартинелли, А.; Лангер, М. (2021). «Обзор окаменелостей ранних динозавров из Южной Америки и его филогенетические последствия». Журнал южноамериканских наук о Земле . 110 : 103341. Бибкод : 2021JSAES.11003341N . дои : 10.1016/j.jsames.2021.103341 . ISSN 0895-9811 .
- ^ Несбитт, Стерлинг Дж; Сьюс, Ханс-Дитер (2021). «Остеология раннего дивергентного динозавра Daemonosaurus chauliodus (Archosauria: Dinosauria) из карьера Целофиза (триасовый период: ретийский период) в Нью-Мексико и его связи с другими ранними динозаврами» . Зоологический журнал Линнеевского общества . 191 (1): 150–179. doi : 10.1093/zoolinnean/zlaa080 .
- ↑ Перейти обратно: Перейти обратно: а б Серено, Пол К. (1999). «Эволюция динозавров» . Наука . 284 (5423). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки : 2137–2147. дои : 10.1126/science.284.5423.2137 . ISSN 0036-8075 . ПМИД 10381873 . Архивировано (PDF) из оригинала 5 января 2018 г. Проверено 8 ноября 2019 г.
- ^ Серено, Пол С.; Форстер, Кэтрин А .; Роджерс, Раймонд Р .; Монетта, Альфредо М. (1993). «Примитивный скелет динозавра из Аргентины и ранняя эволюция динозавров». Природа . 361 (6407). Лондон: Исследования природы: 64–66. Бибкод : 1993Natur.361...64S . дои : 10.1038/361064a0 . ISSN 0028-0836 . S2CID 4270484 .
- ↑ Перейти обратно: Перейти обратно: а б Лангер, Макс К.; Рамезани, Джахандар; Да Роза, Átila AS (май 2018 г.). «U-Pb возрастные ограничения роста динозавров из южной Бразилии». Исследования Гондваны . 57 . Амстердам: Эльзевир: 133–140. Бибкод : 2018GondR..57..133L . дои : 10.1016/j.gr.2018.01.005 . ISSN 1342-937X .
- ^ Новас, FE; Эскурра, доктор медицины; Чаттерджи, С.; Катти, ТС (2011). «Новые виды динозавров из формаций верхнего триаса Верхнего Малери и Нижнего Дхармарама в центральной Индии». Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 101 (3–4): 333–349. Бибкод : 2010EETR.101..333N . дои : 10.1017/S1755691011020093 . S2CID 128620874 .
- ^ Марсикано, Калифорния; Ирмис, РБ; Манкузо, AC; Мундил, Р.; Чемале, Ф. (2016). «Точная временная калибровка происхождения динозавров» . Труды Национальной академии наук . 113 (3): 509–513. Бибкод : 2016PNAS..113..509M . дои : 10.1073/pnas.1512541112 . ПМЦ 4725541 . ПМИД 26644579 .
- ^ Несбитт, Стерлинг Дж.; Барретт, Пол М.; Вернинг, Сара; и др. (2012). «Самый старый динозавр? Динозавроформ среднего триаса из Танзании» . Письма по биологии . 9 (1). Лондон: Королевское общество: 20120949. doi : 10.1098/rsbl.2012.0949 . ISSN 1744-9561 . ПМЦ 3565515 . ПМИД 23221875 .
- ^ Марсикано, Калифорния; Ирмис, РБ; Манкузо, AC; Мундил, Р.; Чемале, Ф. (2015). «Точная временная калибровка происхождения динозавров» . Труды Национальной академии наук . 113 (3): 509–513. Бибкод : 2016PNAS..113..509M . дои : 10.1073/pnas.1512541112 . ISSN 0027-8424 . ПМЦ 4725541 . ПМИД 26644579 .
- ^ Брусатте, Стивен Л .; Бентон, Майкл Дж.; Рута, Марчелло ; Ллойд, Грэм Т. (2008). «Превосходство, конкуренция и оппортунизм в эволюционном излучении динозавров» (PDF) . Наука . 321 (5895). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1485–1488. Бибкод : 2008Sci...321.1485B . дои : 10.1126/science.1161833 . hdl : 20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b . ISSN 0036-8075 . ПМИД 18787166 . S2CID 13393888 . Архивировано (PDF) из оригинала 19 июля 2018 г. Проверено 22 октября 2019 г.
- ^ Таннер, Шпильманн и Лукас 2013 , стр. 562–566 , «Первый норианский (ревуэльтский) ринхозавр: формация Булл-Каньон, Нью-Мексико, США» Джастина А. Спилмана, Спенсера Г. Лукаса и Адриана П. Ханта.
- ^ Сулей, Томаш; Недзведский, Гжегож (2019). «Синапсид позднего триаса размером с слона с прямыми конечностями» . Наука . 363 (6422). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 78–80. Бибкод : 2019Sci...363...78S . дои : 10.1126/science.aal4853 . ISSN 0036-8075 . ПМИД 30467179 . S2CID 53716186 .
- ^ «Ископаемые следы в Альпах помогают объяснить эволюцию динозавров» . Наука и технологии. Экономист . Лондон. 19 апреля 2018 г. ISSN 0013-0613 . Проверено 24 мая 2018 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Weishampel, Dodson & Osmólska 2004 , стр. 627–642, гл. 27: «Мезозойская биогеография динозавров» Томаса Р. Хольца-младшего, Ральфа Э. Чепмена и Мэтью К. Ламанна .
- ↑ Перейти обратно: Перейти обратно: а б с д и Weishampel, Dodson & Osmólska 2004 , стр. 614–626, гл. 26: «Палеоэкология динозавров» Дэвида Э. Фастовского и Джошуа Б. Смита.
- ^ Серено, Пол С.; Уилсон, Джеффри А .; Уитмер, Лоуренс М .; и др. (2007). Кемп, Том (ред.). «Структурные крайности мелового динозавра» . ПЛОС ОДИН . 2 (11). Сан-Франциско, Калифорния : PLOS : e1230. Бибкод : 2007PLoSO...2.1230S . дои : 10.1371/journal.pone.0001230 . ISSN 1932-6203 . ПМК 2077925 . ПМИД 18030355 .
- ^ Прасад, Вандана; Стрёмберг, Кэролайн А.Е.; Алимохаммадиан, Хабиб; и др. (2005). «Копролиты динозавров и ранняя эволюция трав и травоядных». Наука . 310 (5751). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1170–1180. Бибкод : 2005Sci...310.1177P . дои : 10.1126/science.1118806 . ISSN 0036-8075 . ПМИД 16293759 . S2CID 1816461 .
- ^ Weishampel, Dodson & Osmólska 2004 , стр. 672–684, гл. 30: «Вымирание динозавров» Дж. Дэвида Арчибальда и Дэвида Э. Фастовского.
- ^ Дайк и Кайзер 2011 , гл. 14: «Эволюция птиц через границу K – Pg и базальная диверсификация неорнитина», Бент Э.К. Линдоу. два : 10.1002/9781119990475.ch14
- ^ Кракрафт, Джоэл (1968). «Обзор Bathornithidae (Aves, Gruiformes) с замечаниями о взаимоотношениях подотряда Cariamae» (PDF) . Американский музей послушников (2326 г.). Нью-Йорк: Американский музей естественной истории: 1–46. hdl : 2246/2536 . ISSN 0003-0082 . Проверено 22 октября 2019 г.
- ^ Альваренга, Геркулано ; Джонс, Вашингтон, Запад; Риндеркнехт, Андрес (май 2010 г.). «Самая молодая находка птиц-форусрацидов (Aves, Phorusrhacidae) из позднего плейстоцена Уругвая» . Новогодний ежегодник геологии и палеонтологии . 256 (2). Штутгарт : Э. Швейцербарт : 229–234. дои : 10.1127/0077-7749/2010/0052 . ISSN 0077-7749 . Проверено 22 октября 2019 г.
- ^ Майр 2009 г.
- ^ Пол 1988 , стр. 248–250.
- ^ Weishampel, Dodson & Osmólska 2004 , стр. 151–164, гл. 7: «Therizinosauroidea» Джеймса М. Кларка, Терезы Марьянской и Ринчена Барсболда .
- ^ Weishampel, Dodson & Osmólska 2004 , стр. 196–210, гл. 10: «Dromaeosauridae» Питера Дж. Маковицки и Марка А. Норелла .
- ^ Тейлор, Майкл П .; Ведель, Мэтью Дж. (2013). «Почему у зауроподов были длинные шеи и почему у жирафов шеи короткие» . ПерДж . 1 . Корте-Мадера, Калифорния ; Лондон: e36. дои : 10.7717/peerj.36 . ISSN 2167-8359 . ПМЦ 3628838 . ПМИД 23638372 .
- ^ Джастин Твит. «Классификационные диаграммы» . Экваториальная Миннесота . Проверено 6 сентября 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б с Александр, Р. Макнил (2006). «Биомеханика динозавров» . Труды Королевского общества Б. 273 (1596). Лондон: Королевское общество: 1849–1855. дои : 10.1098/rspb.2006.3532 . ISSN 0962-8452 . ПМЦ 1634776 . ПМИД 16822743 .
- ^ Фарлоу, Джеймс О.; Додсон, Питер; Чинсами, Анусуя (ноябрь 1995 г.). «Биология динозавров». Ежегодный обзор экологии и систематики . 26 . Пало-Альто, Калифорния : Ежегодные обзоры : 445–471. doi : 10.1146/annurev.es.26.110195.002305 . ISSN 1545-2069 .
- ^ Вейшампель, Додсон и Осмольска, 2004 г.
- ^ Додсон и Джинджерич 1993 , стр. 167–199 , «О редкости крупных и свирепых животных: предположения о размерах тела, плотности населения и географических ареалах хищных млекопитающих и крупных плотоядных динозавров» Джеймса О. Фарлоу.
- ^ Печкис, Ян (1995). «Последствия оценок массы тела динозавров». Журнал палеонтологии позвоночных . 14 (4). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис от Общества палеонтологии позвоночных : 520–533. Бибкод : 1995JVPal..14..520P . дои : 10.1080/02724634.1995.10011575 . ISSN 0272-4634 . JSTOR 4523591 .
- ^ «Эволюция динозавров» . Отдел палеобиологии . Динозавры. Вашингтон, округ Колумбия: Национальный музей естественной истории . 2007. Архивировано из оригинала 11 ноября 2007 года . Проверено 21 ноября 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б с Сандер, П. Мартин; Кристиан, Андреас; Клаусс, Маркус; и др. (февраль 2011 г.). «Биология динозавров-зауроподов: эволюция гигантизма» . Биологические обзоры . 86 (1). Кембридж: Кембриджское философское общество: 117–155. дои : 10.1111/j.1469-185X.2010.00137.x . ISSN 1464-7931 . ПМК 3045712 . ПМИД 21251189 .
- ↑ Перейти обратно: Перейти обратно: а б с Foster & Lucas 2006 , стр. 131–138 , «Самый большой из больших: критическая переоценка мегазауропода Amphicoelias fragillimus Cope, 1878», Кеннет Карпентер .
- ^ Пол 2010
- ^ Кольбер 1971
- ^ Маццетта, Херардо В.; Кристиансенб, Пер; Фариньяа, Ричард А. (2004). «Гиганты и причуды: размер тела некоторых меловых динозавров южной и южной Америки» (PDF) . Историческая биология . 16 (2–4). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис: 71–83. Бибкод : 2004HBio...16...71M . CiteSeerX 10.1.1.694.1650 . дои : 10.1080/08912960410001715132 . ISSN 0891-2963 . S2CID 56028251 . Архивировано (PDF) из оригинала 25 февраля 2009 г.
- ^ Яненш, Вернер (1950). « Brachiosaurus brancai » Реконструкция скелета (PDF) . Палеонтографика . Приложение VII (1-я серия, часть 3, поставка 2). Перевод Герхарда Майера. Штутгарт: Э. Швейцербарт : 97–103. OCLC 45923346 . Архивировано (PDF) из оригинала 11 июля 2017 г. Проверено 24 октября 2019 г.
- ^ Лукас, Спенсер Г.; Херн, Мэтью С.; Хекет, Эндрю Б.; и др. (2004). Переоценка сейсмозавра , позднеюрского динозавра-зауропода из Нью-Мексико . Ежегодное собрание в Денвере, 2004 г. (7–10 ноября 2004 г.) . Том. 36. Боулдер, Колорадо: Геологическое общество Америки. п. 422. OCLC 62334058 . Статья № 181-4. Архивировано из оригинала 8 октября 2019 года . Проверено 25 октября 2019 г.
- ^ Селлерс, Уильям Ирвин; Маргеттс, Ли; Кориа, Родольфо Анибал ; Мэннинг, Филип Ларс (2013). Кэрриер, Дэвид (ред.). «Марш титанов: двигательные возможности динозавров-зауроподов» . ПЛОС ОДИН . 8 (10). Сан-Франциско, Калифорния: PLOS: e78733. Бибкод : 2013PLoSO...878733S . дои : 10.1371/journal.pone.0078733 . ISSN 1932-6203 . ПМЦ 3864407 . ПМИД 24348896 .
- ^ Лавлейс, Дэвид М.; Хартман, Скотт А.; Уол, Уильям Р. (октябрь – декабрь 2007 г.). «Морфология экземпляра суперзавра (Dinosauria, Sauropoda) из формации Моррисон в Вайоминге и переоценка филогении диплодоцидов» . Arquivos do Museu Nacional . 65 (4). Рио-де-Жанейро : Национальный музей Бразилии ; Федеральный университет Рио-де-Жанейро : 527–544. CiteSeerX 10.1.1.603.7472 . ISSN 0365-4508 . Проверено 26 октября 2019 г.
- ^ Карпентер, Кеннет (2018). «Maraapunisaurus fragillimus, NG (ранее Amphicoelias fragillimus), базальный реббахизаврид из формации Моррисон (верхняя юра) в Колорадо» . Геология Межгорного Запада . 5 : 227–244. дои : 10.31711/giw.v5i0.28 .
- ^ Пол, Грегори С. (2019). «Определение самого крупного известного наземного животного: критическое сравнение различных методов восстановления объема и массы вымерших животных» (PDF) . Анналы музея Карнеги . 85 (4): 335–358. дои : 10.2992/007.085.0403 . S2CID 210840060 .
- ^ Пал, Саураб; Айясами, Кришнан (27 июня 2022 г.). «Потерянный титан Кавери». Геология сегодня . 38 (3): 112–116. Бибкод : 2022GeolT..38..112P . дои : 10.1111/gto.12390 . ISSN 0266-6979 . S2CID 250056201 .
- ^ Пол, Грегори С.; Ларраменди, Азиер (11 апреля 2023 г.). «Оценка массы тела брухаткайозавра и других фрагментарных останков зауроподов позволяет предположить, что крупнейшие наземные животные были примерно такими же большими, как и величайшие киты». Летайя . 56 (2): 1–11. Бибкод : 2023Лета..56..2.5П . дои : 10.18261/let.56.2.5 . ISSN 0024-1164 . S2CID 259782734 .
- ^ Даль Сассо, Криштиану; Магануко, Симоне; Бюффето, Эрик; и др. (2005). «Новая информация о черепе загадочного теропода спинозавра с замечаниями о его размерах и сходстве» (PDF) . Журнал палеонтологии позвоночных . 25 (4). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис от Общества палеонтологии позвоночных: 888–896. doi : 10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2 . ISSN 0272-4634 . S2CID 85702490 . Архивировано из оригинала (PDF) 29 апреля 2011 года . Проверено 5 мая 2011 г.
- ↑ Перейти обратно: Перейти обратно: а б Терриен, Франсуа; Хендерсон, Дональд М. (2007). «Мой теропод больше вашего… или нет: оцениваем размер тела теропод по длине черепа». Журнал палеонтологии позвоночных . 27 (1). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис от Общества палеонтологии позвоночных: 108–115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . ISSN 0272-4634 . S2CID 86025320 .
- ^ Чжао, Сицзинь ; Ли, Дуньцзин; Хан, Банда; и др. (2007). «Zhuchengosaurus maximus из провинции Шаньдун». Acta Geoscientia Sinica . 28 (2). Пекин : Китайская академия геологических наук : 111–122. ISSN 1006-3021 .
- ^ Weishampel, Dodson & Osmólska 2004 , стр. 438–463, гл. 20: «Hadrosauridae» Джона Р. Хорнера, Дэвида Б. Вейшампеля и Кэтрин А. Форстер.
- ^ Норелл, Гаффни и Дингус 2000
- ^ «Колибри-пчелка ( Mellisuga helenae )» . Birds.com . Пейли Медиа. Архивировано из оригинала 3 апреля 2015 года . Проверено 27 октября 2019 г.
- ↑ Перейти обратно: Перейти обратно: а б Чжан, Фучэн; Чжоу, Чжунхэ ; Сюй, Син ; и др. (2008). «Причудливый юрский манирапторан из Китая с удлиненными лентообразными перьями». Природа . 455 (7216). Лондон: Исследования природы: 1105–1108. Бибкод : 2008Natur.455.1105Z . дои : 10.1038/nature07447 . ISSN 0028-0836 . ПМИД 18948955 . S2CID 4362560 .
- ↑ Перейти обратно: Перейти обратно: а б Сюй, Син; Чжао, Ци; Норелл, Марк; и др. (февраль 2008 г.). «Новая окаменелость пернатого динозавра-манираптора, которая заполняет морфологический пробел в птичьем происхождении» . Китайский научный бюллетень . 54 (3). Амстердам: Elsevier от имени науки в Китае Press: 430–435. Бибкод : 2009SciBu..54..430X . дои : 10.1007/s11434-009-0009-6 . ISSN 1001-6538 . S2CID 53445386 .
- ^ Хольц 2007
- ^ Батлер, Ричард Дж.; Чжао, Ци (февраль 2009 г.). «Маленькие птицетазовые динозавры Micropachycephalosaurus hongtuyanensis и Wannanosaurus yansiensis из позднего мела Китая». Меловые исследования . 30 (1). Амстердам: Эльзевир: 63–77. Бибкод : 2009CrRes..30...63B . дои : 10.1016/j.cretres.2008.03.002 . ISSN 0195-6671 .
- ^ Янс, Йохан; Дежакс, Жан; Понс, Дениз; и др. (январь – февраль 2005 г.). «Палеонтологическое и геодинамическое значение палинологического датирования отложений вельденской фации Берниссара (бассейн Монса, Бельгия)». Comptes Rendus Palevol (на французском языке). 4 (1–2). Амстердам: Elsevier от имени Французской академии наук: 135–150. Бибкод : 2005CRPal...4..135Y . дои : 10.1016/j.crpv.2004.12.003 . ISSN 1631-0683 .
- ^ Дэй, Джулия Дж.; Апчерч, Пол; Норман, Дэвид Б.; и др. (2002). «Следы, эволюция и поведение зауроподов» (PDF) . Наука . 296 (5573). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1659. doi : 10.1126/science.1070167 . ISSN 0036-8075 . ПМИД 12040187 . S2CID 36530770 .
- ^ Карри Роджерс и Уилсон 2005 , стр. 252–284, глава. 9: «Шаги к пониманию биологии зауроподов: важность следов зауроподов», Джоанна Л. Райт.
- ^ Варриккио, Дэвид Дж.; Серено, Пол С.; Чжао, Сицзинь; и др. (2008). «Стадо, застрявшее в грязи, является свидетельством отличительной социальности динозавров» (PDF) . Acta Palaeontologica Polonica . 53 (4). Варшава : Институт палеобиологии Польской академии наук : 567–578. дои : 10.4202/app.2008.0402 . ISSN 0567-7920 . S2CID 21736244 . Архивировано (PDF) из оригинала 30 марта 2019 г. Проверено 6 мая 2011 г.
- ^ Lessem & Glut 1993 , стр. 19–20 , «Аллозавр»
- ^ Максвелл, В. Десмонд; Остром, Джон Х. (1995). «Тафономия и палеобиологические последствия ассоциаций тенонтозавра и дейнониха ». Журнал палеонтологии позвоночных . 15 (4). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис от Общества палеонтологии позвоночных: 707–712. Бибкод : 1995JVPal..15..707M . дои : 10.1080/02724634.1995.10011256 . ISSN 0272-4634 .
- ^ Роуч, Брайан Т.; Бринкман, Дэниел Л. (апрель 2007 г.). «Переоценка совместной охоты стаей и стадности Deinonychus antirhopus и других нонавианских тероподных динозавров». Бюллетень Музея естественной истории Пибоди . 48 (1). Нью-Хейвен, Коннектикут : Музей естественной истории Пибоди : 103–138. doi : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . ISSN 0079-032X . S2CID 84175628 .
- ^ Танке, Даррен Х. (1998). «Кусание головы динозавров-теропод: палеопатологические данные» (PDF) . Гайя: Revista de Geociências (15). Лиссабон : Национальный музей естественной истории и науки : 167–184. дои : 10.7939/R34T6FJ1P . ISSN 0871-5424 . S2CID 90552600 . Архивировано из оригинала (PDF) 27 февраля 2008 г.
- ^ «Боевые динозавры» . Нью-Йорк: Американский музей естественной истории. Архивировано из оригинала 18 января 2012 года . Проверено 5 декабря 2007 г.
- ↑ Перейти обратно: Перейти обратно: а б Карпентер, Кеннет (1998). «Свидетельства хищнического поведения динозавров-теропод» (PDF) . Гайя: Revista de Geociências . 15 . Лиссабон: Национальный музей естественной истории и науки: 135–144. ISSN 0871-5424 . Архивировано (PDF) из оригинала 26 сентября 2013 г.
- ^ Роджерс, Раймонд Р.; Краузе, Дэвид В .; Карри Роджерс, Кристина (2007). «Каннибализм у мадагаскарского динозавра Majungatholus atopus ». Природа . 422 (6931). Лондон: Исследования природы: 515–518. Бибкод : 2003Natur.422..515R . дои : 10.1038/nature01532 . ISSN 0028-0836 . ПМИД 12673249 . S2CID 4389583 .
- ^ Шмитц, Ларс; Мотани, Рёске (2011). «Ночной образ жизни динозавров, выведенный на основании морфологии склерального кольца и орбиты». Наука . 332 (6030). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 705–708. Бибкод : 2011Sci...332..705S . дои : 10.1126/science.1200043 . ISSN 0036-8075 . ПМИД 21493820 . S2CID 33253407 .
- ^ Варриккио, Дэвид Дж.; Мартин, Энтони Дж .; Кацура, Ёсихиро (2007). «Первые следы и окаменелости тела роющего динозавра» . Труды Королевского общества Б. 274 (1616). Лондон: Королевское общество: 1361–1368. дои : 10.1098/rspb.2006.0443 . ISSN 0962-8452 . ПМК 2176205 . ПМИД 17374596 .
- ^ Чиаппе и Витмер, 2002 г.
- ^ Чаттерджи, Санкар ; Темплин, Р. Джек (2007). «Форма крыла биплана и летные характеристики пернатого динозавра Microraptor gui » (PDF) . Учеб. Натл. акад. наук. США . 104 (5). Вашингтон, округ Колумбия: Национальная академия наук: 1576–1580 гг. Бибкод : 2007PNAS..104.1576C . дои : 10.1073/pnas.0609975104 . ISSN 0027-8424 . ПМК 1780066 . ПМИД 17242354 . Архивировано (PDF) из оригинала 18 августа 2019 г. Проверено 29 октября 2019 г.
- ^ Гориели, Ален; Макмиллен, Тайлер (2002). «Форма трескающегося кнута». Письма о физических отзывах . 88 (24). Ридж, Нью-Йорк : Американское физическое общество : 244301. Бибкод : 2002PhRvL..88x4301G . doi : 10.1103/PhysRevLett.88.244301 . ISSN 0031-9007 . ПМИД 12059302 .
- ^ Хендерсон, Дональд М. (2003). «Влияние камней в желудке на плавучесть и равновесие плавающего крокодила: вычислительный анализ». Канадский журнал зоологии . 81 (8). Оттава : NRC Research Press : 1346–1357. дои : 10.1139/z03-122 . ISSN 0008-4301 .
- ↑ Перейти обратно: Перейти обратно: а б Сентер, Фил (2008). «Голоса прошлого: обзор звуков палеозойских и мезозойских животных» . Историческая биология . 20 (4). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис: 255–287. Бибкод : 2008HBio...20..255S . дои : 10.1080/08912960903033327 . ISSN 0891-2963 . S2CID 84473967 .
- ^ Ли, Цюаньго; Гао, Кэ-Цинь; Винтер, Якоб; и др. (2010). «Цвет оперения вымершего динозавра» (PDF) . Наука . 327 (5971). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1369–1372. Бибкод : 2010Sci...327.1369L . дои : 10.1126/science.1186290 . ISSN 0036-8075 . ПМИД 20133521 . S2CID 206525132 . Архивировано (PDF) из оригинала 30 марта 2019 г. Проверено 7 ноября 2019 г.
- ^ Риде, Т. (2019). «Эволюция сиринкса: акустическая теория» . ПЛОС ОДИН . 17 (2): e2006507. doi : 10.1371/journal.pbio.2006507 . ПМК 6366696 . ПМИД 30730882 .
- ^ Кларк, Джулия А .; Чаттерджи, Санкар; Чжихэн, Ли; и др. (2016). «Ископаемые свидетельства птичьего голосового органа мезозоя». Природа . 538 (7626). Лондон: Исследования природы: 502–505. Бибкод : 2016Natur.538..502C . дои : 10.1038/nature19852 . ISSN 0028-0836 . ПМИД 27732575 . S2CID 4389926 .
- ^ Кингсли, EP; и др. (2018). «Идентичность и новизна птичьего сиринкса» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (41): 10109–10217. Бибкод : 2018PNAS..11510209K . дои : 10.1073/pnas.1804586115 . ПМК 6187200 . ПМИД 30249637 .
- ^ Ёсида, Джунки; Кобаяши, Ёсицугу; Норелл, Марк А. (15 февраля 2023 г.). «Гортань анкилозавра дает представление о птичьих вокалах у нептичьих динозавров» . Коммуникационная биология . 6 (1): 152. дои : 10.1038/s42003-023-04513-x . ISSN 2399-3642 . ПМЦ 9932143 . ПМИД 36792659 .
- ^ Риде, Тобиас; Элиасон, Чад М.; Миллер, Эдвард Х.; и др. (2016). «Воркование, гул и уханье: эволюция вокального поведения птиц с закрытым ртом» . Эволюция . 70 (8). Хобокен, Нью-Джерси: Джон Уайли и сыновья от Общества изучения эволюции : 1734–1746. дои : 10.1111/evo.12988 . ISSN 0014-3820 . ПМИД 27345722 . S2CID 11986423 .
- ^ Вейшампель, Дэвид Б. (весна 1981 г.). «Акустический анализ вокализации динозавров-ламбеозавров (Reptilia: Ornithischia)» (PDF) . Палеобиология . 7 (2). Бетесда, Мэриленд : Палеонтологическое общество : 252–261. дои : 10.1017/S0094837300004036 . ISSN 0094-8373 . JSTOR 2400478 . S2CID 89109302 . Архивировано из оригинала (PDF) 6 октября 2014 г. Проверено 30 октября 2019 г.
- ^ Мияшита, Тецуто; Арбур, Виктория М .; Уитмер, Лоуренс М.; и др. (декабрь 2011 г.). «Внутренняя морфология черепа панцирного динозавра Euoplocephalus, подтвержденная рентгеновской компьютерной томографической реконструкцией» (PDF) . Журнал анатомии . 219 (6). Хобокен, Нью-Джерси: Джон Уайли и сыновья: 661–675. дои : 10.1111/j.1469-7580.2011.01427.x . ISSN 1469-7580 . ПМК 3237876 . ПМИД 21954840 . Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 30 октября 2019 г.
- ^ Ханселл 2000
- ↑ Перейти обратно: Перейти обратно: а б Варриккио, Дэвид Дж.; Хорнер, Джон Р.; Джексон, Фрэнки Д. (2002). «Эмбрионы и яйца мелового теропода динозавра Troodon formosus ». Журнал палеонтологии позвоночных . 22 (3). Милтон-Парк, Оксфордшир: Тейлор и Фрэнсис от Общества палеонтологии позвоночных: 564–576. doi : 10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2 . ISSN 0272-4634 . S2CID 85728452 .
- ^ Ли, Эндрю Х.; Вернинг, Сара (2008). «Половая зрелость растущих динозавров не соответствует моделям роста рептилий» . Учеб. Натл. акад. наук. США . 105 (2). Вашингтон, округ Колумбия: Национальная академия наук: 582–587. Бибкод : 2008PNAS..105..582L . дои : 10.1073/pnas.0708903105 . ISSN 0027-8424 . ПМК 2206579 . ПМИД 18195356 .
- ^ Хорнер, Джон Р.; Макела, Роберт (1979). «Гнездо молоди является свидетельством семейной структуры динозавров». Природа . 282 (5736). Лондон: Исследования природы: 296–298. Бибкод : 1979Natur.282..296H . дои : 10.1038/282296a0 . ISSN 0028-0836 . S2CID 4370793 .
- ^ «Открытие поведения динозавров: взгляд с 1960 года по настоящее время» . Британская энциклопедия . Чикаго, Иллинойс : Британская энциклопедия, Inc. Архивировано из оригинала 13 декабря 2013 года . Проверено 30 октября 2019 г.
- ^ Карри и др. 2004 , стр. 234–250 , гл. 11: «Поведение динозавров и происхождение маховых перьев» Томаса П. Хоппа и Марка Дж. Орсена.
- ^ Рейс, Роберт Р .; Скотт, Дайан; Сьюс, Ханс-Дитер ; и др. (2005). «Эмбрионы раннеюрского динозавра-прозауропода и их эволюционное значение» (PDF) . Наука . 309 (5735). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 761–764. Бибкод : 2005Sci...309..761R . дои : 10.1126/science.1114942 . ISSN 0036-8075 . ПМИД 16051793 . S2CID 37548361 . Архивировано (PDF) из оригинала 22 июля 2018 г.
- ^ Кларк, Нил Д.Л.; Бут, Пол; Бут, Клэр Л.; и др. (2004). «Следы динозавров из формации Дантулм (бат, юра) острова Скай» (PDF) . Шотландский геологический журнал . 40 (1). Лондон: Лондонское геологическое общество : 13–21. Бибкод : 2004ScJG...40...13C . дои : 10.1144/sjg40010013 . ISSN 0036-9276 . S2CID 128544813 . Архивировано (PDF) из оригинала 22 июля 2013 г. Проверено 12 декабря 2019 г.
- ^ Чжоу, Чжунхэ; Чжан, Фучэн (2004). «Преждевременный птичий эмбрион из нижнего мела Китая». Наука . 306 (5696). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 653. doi : 10.1126/science.1100000 . ISSN 0036-8075 . ПМИД 15499011 . S2CID 34504916 .
- ^ Нэйш, Даррен (15 мая 2012 г.). «Затонувшая гнездовая колония позднемеловых птиц» . Наука . 306 (5696). Scientific American : 653. doi : 10.1126/science.1100000 . ПМИД 15499011 . S2CID 34504916 . Архивировано из оригинала 25 сентября 2018 года . Проверено 16 ноября 2019 г.
- ^ Фернандес, Мариэла С.; Гарсия, Родольфо А.; Фиорелли, Лукас; и др. (2013). «Большое скопление птичьих яиц из позднего мела Патагонии (Аргентина) раскрывает новую стратегию гнездования мезозойских птиц» . ПЛОС ОДИН . 8 (4). Сан-Франциско, Калифорния: PLOS: e61030. Бибкод : 2013PLoSO...861030F . дои : 10.1371/journal.pone.0061030 . ISSN 1932-6203 . ПМК 3629076 . ПМИД 23613776 .
- ^ Диминг, Денис Чарльз; Майр, Джеральд (май 2018 г.). «Морфология таза предполагает, что ранние мезозойские птицы были слишком тяжелыми, чтобы высиживать яйца при контакте» (PDF) . Журнал эволюционной биологии . 31 (5). Хобокен, Нью-Джерси: Уайли-Блэквелл от имени Европейского общества эволюционной биологии : 701–709. дои : 10.1111/jeb.13256 . ISSN 1010-061X . ПМИД 29485191 . S2CID 3588317 . Архивировано (PDF) из оригинала 2 июня 2020 г.
- ^ Майерс, Тимоти С.; Фиорилло, Энтони Р. (2009). «Доказательства стадного поведения и возрастной сегрегации динозавров-зауроподов» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 274 (1–2). Амстердам: Эльзевир: 96–104. Бибкод : 2009PPP...274...96M . дои : 10.1016/j.palaeo.2009.01.002 . ISSN 0031-0182 . Архивировано (PDF) из оригинала 29 мая 2020 г.
- ^ Винтер, Якоб; Николлс, Роберт; Келли, Дайан А. (22 февраля 2021 г.). «Клоакальное отверстие у нептичьего динозавра» . Современная биология . 31 (4). Эльзевир: R1–R3. Бибкод : 2021CBio...31.R182V . дои : 10.1016/j.cub.2020.12.039 . ПМИД 33472049 . S2CID 231644183 .
- ^ Weishampel, Dodson & Osmólska 2004 , стр. 643–659, гл. 28: «Физиология нонавианских динозавров» Анусуя Чинсами и Виллем Дж. Хиллениус.
- ^ Понцер, Х.; Аллен, В.; Хатчинсон, младший (2009). «Биомеханика бега указывает на эндотермию у прямоногих динозавров» . ПЛОС ОДИН . 4 (11): е7783. Бибкод : 2009PLoSO...4.7783P . дои : 10.1371/journal.pone.0007783 . ISSN 1932-6203 . ПМЦ 2772121 . ПМИД 19911059 .
- ↑ Перейти обратно: Перейти обратно: а б Бенсон, RBJ (2018). «Макроэволюция динозавров и макроэкология» . Ежегодный обзор экологии, эволюции и систематики . 49 : 379–408. doi : 10.1146/annurev-ecolsys-110617-062231 . S2CID 92837486 .
- ^ Грейди, Дж. М.; Энквист, Би Джей; Деттвейлер-Робинсон, Э.; Райт, Северная Каролина; Смит, Ф.А. (2014). «Доказательства мезотермии у динозавров». Наука . 344 (6189): 1268–1272. Бибкод : 2014Sci...344.1268G . дои : 10.1126/science.1253143 . ПМИД 24926017 . S2CID 9806780 .
- ^ Лежандр, LJ; Генар, Ж.; Бота-Бринк, Дж.; Кубо, Дж. (2016). «Палеогистологические доказательства наследственной высокой скорости метаболизма у архозавров» . Систематическая биология . 65 (6): 989–996. doi : 10.1093/sysbio/syw033 . ПМИД 27073251 .
- ^ Сеймур, RS; Беннетт-Стампер, CL; Джонстон, Южная Дакота; Перевозчик, ДР; Григг, GC (2004). «Доказательства эндотермических предков крокодилов в основе эволюции архозавров» . Физиологическая и биохимическая зоология . 77 (6): 1051–1067. doi : 10.1093/sysbio/syw033 . ПМИД 27073251 .
- ^ Парсонс 2001 , стр. 22–48, «Ереси доктора Баккера».
- ^ Эриксон, генеральный директор (2014). «О росте динозавров». Ежегодный обзор наук о Земле и планетах . 42 (1): 675–697. Бибкод : 2014AREPS..42..675E . doi : 10.1146/annurev-earth-060313-054858 .
- ↑ Перейти обратно: Перейти обратно: а б Байёль, AM; О'Коннор, Дж.; Швейцер, МГ (2019). «Палеогистология динозавров: обзор, тенденции и новые направления исследований» . ПерДж . 7 : е7764. дои : 10.7717/peerj.7764 . ПМК 6768056 . ПМИД 31579624 .
- ^ Де Риклес, А. (1974). «Эволюция эндотермии: гистологические данные» (PDF) . Эволюционная теория . 1 (2): 51–80. Архивировано (PDF) из оригинала 17 апреля 2021 г.
- ^ Де Риклес, А. (1980). «Тканевые структуры костей динозавров, функциональное значение и возможное отношение к физиологии динозавров». В Томасе, РДК; Олсон, ЕС (ред.). Холодный взгляд на теплокровных динозавров . Нью-Йорк: Американская ассоциация содействия развитию науки. стр. 103–139.
- ^ Падиан, К.; Хорнер, младший; де Рикле, А. (2004). «Рост мелких динозавров и птерозавров: эволюция стратегий роста архозавров» (PDF) . Журнал палеонтологии позвоночных . 24 (3): 555–571. doi : 10.1671/0272-4634(2004)024[0555:GISDAP]2.0.CO;2 . S2CID 86019906 .
- ^ де Соуза, Джорджия; Бенто Соарес, М.; Соуза Брум, А.; Зуколотто, М.; Саяо, Дж. М.; Карлос Вайншютц, Л.; Келлнер, AWA (2020). «Остеогистология и динамика роста бразильского ноазавра Vespersaurus paranaensis Langer et al., 2019 (Theropoda: Abelisauroidea)» . ПерДж . 8 : е9771. дои : 10.7717/peerj.9771 . ПМЦ 7500327 . ПМИД 32983636 . S2CID 221906765 .
- ^ Примеры этой работы, проведенной на разных линиях динозавров, см.
- Эриксон, генеральный директор; Туманова, Т.А. (2000). «Кривая роста Psittacosaurus mongoliensis Osborn (Ceratopsia: Psittacosauridae), полученная на основе гистологии длинных костей». Зоологический журнал Линнеевского общества . 130 (4): 551–566. дои : 10.1111/j.1096-3642.2000.tb02201.x . S2CID 84241148 .
- Эриксон, Г.; Роджерс, К.; Йерби, С. (2001). «Модели роста динозавров и быстрые темпы роста птиц». Природа . 412 (429–433): 429–433. Бибкод : 2001Natur.412..429E . дои : 10.1038/35086558 . ПМИД 11473315 . S2CID 4319534 . (Ошибка: дои : 10.1038/nature16488 , PMID 26675731 , Часы втягивания )
- Эриксон, Г.; Маковицкий, П.; Карри, П.; Норелл, Массачусетс; Йерби, ЮАР; Брочу, Калифорния (2004). «Гигантизм и сравнительные параметры жизненного цикла динозавров-тираннозаврид» (PDF) . Природа . 430 (7001): 772–775. Бибкод : 2004Natur.430..772E . дои : 10.1038/nature02699 . ПМИД 15306807 . S2CID 4404887 . Архивировано (PDF) из оригинала 14 июля 2020 г. (Ошибка: дои : 10.1038/nature16487 , PMID 26675726 , Часы втягивания )
- Леман, ТМ; Вудворд, Х.Н. (2008). «Моделирование темпов роста динозавров-зауроподов» (PDF) . Палеобиология . 34 (2): 264–281. doi : 10.1666/0094-8373(2008)034[0264:MGRFSD]2.0.CO;2 . S2CID 84163725 .
- Хорнер, младший; де Риклс, А.; Падиан, К.; Шитц, Р.Д. (2009). «Сравнительная гистология длинных костей и рост динозавров-гипсилофодонтидов Orodromeus makelai , Dryosaurus altus и Tenontosaurustilettii (Ornithischia: Euornithopoda)». Журнал палеонтологии позвоночных . 29 (3): 734–747. Бибкод : 2009JVPal..29..734H . дои : 10.1671/039.029.0312 . S2CID 86277619 .
- Вудворд, Х.; Фридман Фаулер, Э.; Фарлоу, Дж.; Хорнер, Дж. (2015). « Майазавр , модельный организм для биологии вымерших популяций позвоночных: большая выборочная статистическая оценка динамики роста и выживаемости». Палеобиология . 41 (4): 503–527. Бибкод : 2015Pbio...41..503W . дои : 10.1017/pab.2015.19 . S2CID 85902880 .
- ^ Амиот, Р.; Лекюйер, К.; Бюффето, Э.; Эскаргель, Г.; Флюто, Ф.; Мартино, Ф. (2006). «Изотопы кислорода из биогенных апатитов предполагают широко распространенную эндотермию у меловых динозавров» (PDF) . Письма о Земле и планетологии . 246 (1–2): 41–54. Бибкод : 2006E&PSL.246...41A . дои : 10.1016/j.epsl.2006.04.018 .
- ^ Амиот, Р.; Ван, X.; Лекюйер, К.; Бюффето, Э.; Будад, Л.; Кэвин, Л.; Дин, З.; Флюто, Ф.; Келлнер, AWA; Тонг, Х.; Чжан, Ф. (2010). «Изотопный состав кислорода и углерода позвоночных среднего мела Северной Африки и Бразилии: экологическое и экологическое значение». Палеогеография, Палеоклиматология, Палеоэкология . 297 (2): 439–451. Бибкод : 2010PPP...297..439A . дои : 10.1016/j.palaeo.2010.08.027 .
- ^ Колодный Ю.; Луз, Б.; Сандер, М.; Клеменс, Вашингтон (1996). «Кости динозавров: окаменелости или псевдоморфозы? Ловушки физиологической реконструкции на основе апатитовых окаменелостей» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 126 (1–2): 161–171. Бибкод : 1996PPP...126..161K . дои : 10.1016/S0031-0182(96)00112-5 .
- ^ Пол, GS (1988). «Физиологические, миграционные, климатологические, геофизические, выживательные и эволюционные последствия меловых полярных динозавров». Журнал палеонтологии . 62 (4): 640–652. JSTOR 1305468 .
- ^ Клеменс, Вашингтон; Нелмс, Л.Г. (1993). «Палеоэкологические последствия фауны наземных позвоночных Аляски в последний меловой период на высоких палеоширотах». Геология . 21 (6): 503–506. Бибкод : 1993Geo....21..503C . doi : 10.1130/0091-7613(1993)021<0503:PIOATV>2.3.CO;2 .
- ^ Рич, TH; Викерс-Рич, П.; Ганглофф, Р.А. (2002). «Полярные динозавры». Наука . 295 (5557): 979–980. дои : 10.1126/science.1068920 . ПМИД 11834803 . S2CID 28065814 .
- ^ Баффето, Э. (2004). «Полярные динозавры и вопрос вымирания динозавров: краткий обзор» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 214 (3): 225–231. дои : 10.1016/j.palaeo.2004.02.050 . Архивировано (PDF) из оригинала 8 июня 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б Серено, Пол С.; Мартинес, Рикардо Н.; Уилсон, Джеффри А.; и др. (сентябрь 2008 г.). Кемп, Том (ред.). «Доказательства существования птичьих внутригрудных воздушных мешков у нового хищного динозавра из Аргентины» . ПЛОС ОДИН . 3 (9). Сан-Франциско, Калифорния: PLOS: e3303. Бибкод : 2008PLoSO...3.3303S . дои : 10.1371/journal.pone.0003303 . ISSN 1932-6203 . ПМЦ 2553519 . ПМИД 18825273 .
- ^ О'Коннор, премьер-министр (2009). «Эволюция строения тела архозавров: скелетные адаптации дыхательного аппарата на основе воздушного мешка у птиц и других архозавров». Журнал экспериментальной зоологии. Часть A: Экологическая генетика и физиология . 311 (8): 629–646. Бибкод : 2009JEZA..311..629O . дои : 10.1002/jez.548 . ПМИД 19492308 .
- ^ Игл, РА; Тюткен, Т.; Мартин, Т.С.; Трипати, АК; Фрике, ХК; Коннели, М.; Чифелли, РЛ; Эйлер, Дж. М. (2011). «Температура тела динозавра определяется по изотопному ( 13 С- 18 O) упорядоченность ископаемых биоминералов». Science . 333 (6041): 443–445. Bibcode : ...333..443E . doi : 10.1126/science.1206196 . PMID 21700837. 2011Sci S2CID 206534244 .
- ^ Ведель, MJ (2003). «Пневматичность позвонков, воздушные мешки и физиология динозавров-зауроподов» (PDF) . Палеобиология . 29 (2): 243–255. Бибкод : 2003Pbio...29..243W . дои : 10.1017/S0094837300018091 .
- ^ Перри, Сан-Франциско; Кристиан, А.; Брейер, Т.; Пайор, Н.; Кодд, младший (2009). «Последствия дыхательной системы птичьего типа для гигантизма у динозавров-зауроподов». Журнал экспериментальной зоологии. Часть A: Экологическая генетика и физиология . 311 (8): 600–610. Бибкод : 2009JEZA..311..600P . дои : 10.1002/jez.517 . ПМИД 19189317 .
- ^ Александр, РМ (1998). «Гиганты на все времена: самые большие животные и их проблемы» . Палеонтология . 41 : 1231–1245.
- ^ Царь, Э.; Мартинес дель Рио, К.; Ижаки И.; Арад, З. (2005). «Могут ли птицы быть аммонотелическими? Баланс и выделение азота у двух плодоядных» (PDF) . Журнал экспериментальной биологии . 208 (6): 1025–1034. дои : 10.1242/jeb.01495 . ISSN 0022-0949 . ПМИД 15767304 . S2CID 18540594 . Архивировано (PDF) из оригинала 17 октября 2019 г. Проверено 31 октября 2019 г.
- ^ Скадхауге, Э.; Эрльвангер, К.Х.; Рузива, SD; Данцер, В.; Эльбронд, В.С.; Чамунорва, JP (2003). «Обладает ли страусиный копродеум ( Struthio Camelus ) электрофизиологическими свойствами и микроструктурой других птиц?». Сравнительная биохимия и физиология. Часть A: Молекулярная и интегративная физиология . 134 (4): 749–755. дои : 10.1016/S1095-6433(03)00006-0 . ISSN 1095-6433 . ПМИД 12814783 .
- ^ Прест, MR; Беша, Калифорния (1997). «Выделение аммиака колибри». Природа . 386 (6625): 561–562. Бибкод : 1997Natur.386..561P . дои : 10.1038/386561a0 . ISSN 0028-0836 . S2CID 4372695 .
- ^ Мора, Дж.; Мартучелли, Дж.; Ортис Пинеда, Дж.; Соберон, Г. (1965). «Регуляция ферментов биосинтеза мочевины у позвоночных» . Биохимический журнал . 96 (1): 28–35. дои : 10.1042/bj0960028 . ISSN 0264-6021 . ПМК 1206904 . ПМИД 14343146 .
- ^ Паккард, GC (1966). «Влияние температуры и засушливости окружающей среды на способы размножения и выделения амниотных позвоночных». Американский натуралист . 100 (916): 667–682. дои : 10.1086/282459 . ISSN 0003-0147 . JSTOR 2459303 . S2CID 85424175 .
- ^ Балгуен, Т.Г. (1971). «Регургитация гранул у содержащихся в неволе ястребов-перепелятников ( Falco sparverius )» (PDF) . Кондор . 73 (3): 382–385. дои : 10.2307/1365774 . JSTOR 1365774 . Архивировано из оригинала (PDF) 4 апреля 2019 г. Проверено 30 октября 2019 г.
- ^ Сюй, Х.; Ли, Ф.; Ван, Ю.; Салливан, К.; Чжан, Ф.; Чжан, X.; Салливан, К.; Ван, X.; Чжэн, X. (2018). «Исключительные окаменелости динозавров указывают на раннее происхождение птичьего пищеварения» . Научные отчеты . 8 (1): 14217. Бибкод : 2018NatSR...814217Z . дои : 10.1038/s41598-018-32202-x . ISSN 2045-2322 . ПМК 6155034 . ПМИД 30242170 .
- ^ Рассел, Дейл А. (1997). "Интеллект". У Кевина Падиана; Филип Дж. Карри (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 370–372. ISBN 978-0-12-226810-6 .
- ^ Брусатте 2012 , с. 83
- ^ Хаксли, Томас Х. (1868). «О животных, занимающих почти промежуточное положение между птицами и пресмыкающимися» . Анналы и журнал естественной истории . 4 (2). Лондон: Тейлор и Фрэнсис: 66–75 . Проверено 31 октября 2019 г.
- ^ Хайльманн 1926 г.
- ^ Осборн, Генри Фэрфилд (1924). «Три новых теропода, зона протоцератопса , центральная Монголия» (PDF) . Американский музей послушников (144). Нью-Йорк: Американский музей естественной истории: 1–12. ISSN 0003-0082 . Архивировано (PDF) из оригинала 12 июня 2007 г.
- ^ Остром, Джон Х. (1973). «Происхождение птиц» . Природа . 242 (5393). Лондон: Nature Research: 136. Бибкод : 1973NPhS..242..136O . дои : 10.1038/242136a0 . ISSN 0028-0836 . S2CID 29873831 .
- ^ Падиан 1986 , стр. 1–55 , «Ящеричья монофилия и происхождение птиц» Жака Готье .
- ^ Майр, Джеральд; Пол, Буркхард; Питерс, Д. Стефан (2005). «Хорошо сохранившийся образец археоптерикса с чертами теропода» (PDF) . Наука . 310 (5753). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1483–1486. Бибкод : 2005Sci...310.1483M . дои : 10.1126/science.1120331 . ISSN 0036-8075 . ПМИД 16322455 . S2CID 28611454 .
- ^ Мартин, Ларри Д. (2006). «Базальное архозавровое происхождение птиц». Acta Zoologica Sinica . 50 (6): 977–990. ISSN 1674-5507 .
- ↑ Перейти обратно: Перейти обратно: а б Федучча, Алан (1 октября 2002 г.). «Птицы — динозавры: простой ответ на сложную проблему» . Аук . 119 (4). Вашингтон, округ Колумбия: Союз американских орнитологов : 1187–1201. doi : 10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2 . ISSN 0004-8038 . JSTOR 4090252 . S2CID 86096746 . Проверено 3 ноября 2019 г.
- ↑ Перейти обратно: Перейти обратно: а б Свитек, Брайан (2 июля 2012 г.). «Восстание пушистых динозавров» . Новости. Природа . Лондон: Исследования природы. дои : 10.1038/nature.2012.10933 . ISSN 0028-0836 . S2CID 123219913 . Проверено 1 января 2019 г.
- ^ Годфруа, П.; Синица, С.М.; Дуайи, Д.; Болоцкий Ю.Л.; Сизов А.В.; Макнамара, Мэн; Бентон, MJ; Спагна, П. (2014). «Юрский птицетазовый динозавр из Сибири с перьями и чешуей» (PDF) . Наука . 345 (6195): 451–455. Бибкод : 2014Sci...345..451G . дои : 10.1126/science.1253351 . hdl : 1983/a7ae6dfb-55bf-4ca4-bd8b-a5ea5f323103 . ПМИД 25061209 . S2CID 206556907 . Архивировано из оригинала (PDF) 9 февраля 2019 года . Проверено 27 июля 2016 г.
- ^ Сюй, Син; Норелл, Марк А.; Куанг, Сюэвэнь; и др. (2004). «Базальные тираннозавроиды из Китая и свидетельства наличия протоперьев у тираннозавроидов» (PDF) . Природа . 431 (7009). Лондон: Исследования природы: 680–684. Бибкод : 2004Natur.431..680X . дои : 10.1038/nature02855 . ISSN 0028-0836 . ПМИД 15470426 . S2CID 4381777 .
- ^ Гёлих, Урсула Б.; Кьяппе, Луис М. (2006). «Новый плотоядный динозавр с позднеюрского архипелага Зольнхофен» (PDF) . Природа . 440 (7082). Лондон: Исследования природы: 329–332. Бибкод : 2006Natur.440..329G . дои : 10.1038/nature04579 . ISSN 0028-0836 . ПМИД 16541071 . S2CID 4427002 . Архивировано из оригинала (PDF) 26 апреля 2019 г. Проверено 1 ноября 2019 г.
- ^ Келлнер, Александр В.А .; Ван, Сяолинь; Тишлингер, Гельмут; и др. (2010). «Мягкие ткани Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) и строение мембраны крыла птерозавра» . Труды Королевского общества Б. 277 (1679). Лондон: Королевское общество: 321–329. дои : 10.1098/rspb.2009.0846 . ISSN 0962-8452 . ПМЦ 2842671 . ПМИД 19656798 .
- ^ Майр, Г.; Питтман, М.; Сайтта, Э.; Кэй, Т.Г.; Винтер, Дж. (2016). «Строение и гомология пситтакозавра щетинок хвоста » . Палеонтология . 59 (6): 793–802. Бибкод : 2016Palgy..59..793M . дои : 10.1111/пала.12257 . hdl : 1983/029c668f-08b9-45f6-a0c5-30ce9256e593 . S2CID 89156313 .
- ↑ Перейти обратно: Перейти обратно: а б Бентон, MJ; Дуайи, Д.; Цзян, Б.; Макнамара, М. (2019). «Раннее происхождение перьев». Тенденции в экологии и эволюции . 34 (9): 856–869. дои : 10.1016/j.tree.2019.04.018 . hdl : 10468/8068 . ПМИД 31164250 . S2CID 174811556 .
- ^ Барретт, премьер-министр; Эванс, округ Колумбия; Кампионе, Невада (2015). «Эволюция эпидермальных структур динозавров» . Письма по биологии . 11 (6): 20150229. doi : 10.1098/rsbl.2015.0229 . ПМЦ 4528472 . ПМИД 26041865 .
- ^ Алибарди, Лоренцо; Кнапп, Лорен В.; Сойер, Роджер Х. (2006). «Локализация бета-кератина в развивающейся чешуе и перьях аллигатора в связи с развитием и эволюцией перьев». Журнал субмикроскопической цитологии и патологии . 38 (2–3). Сиена : Nuova Immagine Editrice: 175–192. ISSN 1122-9497 . ПМИД 17784647 .
- ^ Лингхэм-Солиар, Теагартен (декабрь 2003 г.). «Динозавровое происхождение перьев: перспективы коллагеновых волокон дельфинов (китообразных)». Naturwissenschaften . 90 (12). Берлин: Springer Science+Business Media : 563–567. Бибкод : 2003NW.....90..563L . дои : 10.1007/s00114-003-0483-7 . ISSN 0028-1042 . ПМИД 14676953 . S2CID 43677545 .
- ↑ Перейти обратно: Перейти обратно: а б Федучча, Алан; Лингем-Солиар, Теагартен; Хинчлифф, Дж. Ричард (ноябрь 2005 г.). «Существуют ли пернатые динозавры? Проверка гипотезы на неонтологических и палеонтологических данных». Журнал морфологии . 266 (2). Хобокен, Нью-Джерси: Джон Уайли и сыновья: 125–166. дои : 10.1002/jmor.10382 . ISSN 0362-2525 . ПМИД 16217748 . S2CID 15079072 .
- ^ Лингем-Солиар, Теагартен; Федучча, Алан; Ван, Сяолинь (2007). «Новый китайский образец указывает на то, что «протоперья» раннемелового тероподного динозавра Sinosauropteryx представляют собой деградировавшие коллагеновые волокна» . Труды Королевского общества Б. 274 (1620). Лондон: Королевское общество: 1823–1829. дои : 10.1098/rspb.2007.0352 . ISSN 0962-8452 . ПМК 2270928 . ПМИД 17521978 .
- ^ Прум, Ричард О. (2003). «Является ли современная критика происхождения теропод наукой? Опровержение Feduccia 2002» . Аук . 120 (2). Вашингтон, округ Колумбия: Союз американских орнитологов: 550–561. doi : 10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2 . ISSN 0004-8038 . JSTOR 4090212 .
- ^ Велнхофер, Питер (1988). «Новый экземпляр археоптерикса ». Наука . 240 (4860). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1790–1792. Бибкод : 1988Sci...240.1790W . дои : 10.1126/science.240.4860.1790 . ISSN 0036-8075 . JSTOR 1701652 . ПМИД 17842432 . S2CID 32015255 .
- —— (1988). «Новый образец археоптерикса ». Археоптерикс . 6 :1–30.
- ^ Швейцер, Мэри Х.; Ватт, Дж.А.; Авчи, Р.; и др. (1999). «Специфическая иммунологическая реактивность бета-кератина в перьевых структурах меловых альваресзаврид, Shuvuuia Deserti ». Журнал экспериментальной зоологии, часть B. 285 (2). Хобокен, Нью-Джерси: Уайли-Блэквелл: 146–157. Бибкод : 1999JEZ...285..146S . doi : 10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A . ISSN 1552-5007 . ПМИД 10440726 .
- ^ « Археоптерикс : ранняя пташка» . Беркли: Музей палеонтологии Калифорнийского университета . Проверено 30 октября 2019 г.
- ^ О'Коннор, Патрик М.; Классенс, Леон ПАМ (2005). «Базовая конструкция птичьих легких и проточная вентиляция у нептичьих динозавров-теропод». Природа . 436 (7048). Лондон: Исследования природы: 253–256. Бибкод : 2005Natur.436..253O . дои : 10.1038/nature03716 . ISSN 0028-0836 . ПМИД 16015329 . S2CID 4390587 .
- ^ Гибсон, Андреа (13 июля 2005 г.). «Исследование: у хищных динозавров была птичья легочная система» . Исследовательские коммуникации . Афины, Огайо : Университет Огайо . Проверено 18 ноября 2019 г. [ постоянная мертвая ссылка ]
- ^ «Плотоядный динозавр из Аргентины имел птичью систему дыхания» . Новости Мичиганского университета . Анн-Арбор, Мичиган : Офис вице-президента по коммуникациям; Регенты Мичиганского университета . 2 октября 2008 года . Проверено 2 ноября 2019 г.
- ^ Сюй, Син; Норелл, Марк А. (2004). «Новый троодонтидный динозавр из Китая с птичьей позой сна» (PDF) . Природа . 431 (7010). Лондон: Исследования природы: 838–841. Бибкод : 2004Natur.431..838X . дои : 10.1038/nature02898 . ISSN 0028-0836 . ПМИД 15483610 . S2CID 4362745 .
- ^ Норелл, Марк А.; Кларк, Джеймс М.; Кьяппе, Луис М.; и др. (1995). «Гнездящийся динозавр». Природа . 378 (6559). Лондон: Исследования природы: 774–776. Бибкод : 1995Natur.378..774N . дои : 10.1038/378774a0 . ISSN 0028-0836 . S2CID 4245228 .
- ^ Варриккио, Дэвид Дж.; Мур, Джейсон Р.; Эриксон, Грегори М .; и др. (2008). «Птичья отцовская забота имела происхождение от динозавров» . Наука . 322 (5909). Вашингтон, округ Колумбия: Американская ассоциация содействия развитию науки: 1826–1828 гг. Бибкод : 2008Sci...322.1826V . дои : 10.1126/science.1163245 . ISSN 0036-8075 . ПМИД 19095938 . S2CID 8718747 .
- ^ Крылья, Оливер (2007). «Обзор функции гастролитов с последствиями для ископаемых позвоночных и пересмотренная классификация» (PDF) . Палеонтологическая Полоника . 52 (1). Варшава: Институт палеобиологии Польской академии наук : 1–16. ISSN 0567-7920 . Архивировано (PDF) из оригинала 17 декабря 2008 г. Проверено 2 ноября 2019 г.
- ^ Лонгрич, Северная Каролина; Токарык Т.; Филд, диджей (2011). «Массовое вымирание птиц на рубеже мела и палеогена (K – Pg)» . Труды Национальной академии наук . 108 (37): 15253–15257. Бибкод : 2011PNAS..10815253L . дои : 10.1073/pnas.1110395108 . ПМК 3174646 . ПМИД 21914849 .
- ↑ Перейти обратно: Перейти обратно: а б Ренне, PR; Дейно, Алабама; Хильген, Ф.Дж.; Койпер, К.Ф.; Марк, Д.Ф.; Митчелл, У.С.; Морган, Ле; Мундил, Р.; Смит, Дж. (2013). «Временные шкалы критических событий на границе мела и палеогена». Наука . 339 (6120): 684–687. Бибкод : 2013Sci...339..684R . дои : 10.1126/science.1230492 . ПМИД 23393261 . S2CID 6112274 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Брусатте, СЛ; Батлер, Р.Дж.; Барретт, премьер-министр; Каррано, Монтана; Эванс, округ Колумбия; Ллойд, GT; Мэннион, Полиция; Норелл, Массачусетс; Пеппе, диджей; Апчерч, П.; Уильямсон, Т.Э. (2015). «Вымирание динозавров» . Биологические обзоры . 90 (2): 628–642. дои : 10.1111/brv.12128 . hdl : 20.500.11820/176e5907-26ec-4959-867f-0f2e52335f88 . ПМИД 25065505 . S2CID 115134484 .
- ↑ Перейти обратно: Перейти обратно: а б Маклауд, Н.; Роусон, ПФ; Фори, Польша; и др. (1997). «Мел-третичный биотический переход». Журнал Геологического общества . 154 (2): 265–292. Бибкод : 1997JGSoc.154..265M . дои : 10.1144/gsjgs.154.2.0265 . ISSN 0016-7649 . S2CID 129654916 .
- ↑ Перейти обратно: Перейти обратно: а б Арчибальд, доктор медицинских наук; Клеменс, Вашингтон (1982). «Позднемеловое вымирание». Американский учёный . 70 (4): 377–385. Бибкод : 1982AmSci..70..377A . JSTOR 27851545 .
- ^ Яблонски, Д. (1991). «Вымирания: палеонтологическая перспектива». Наука . 253 (5021): 754–757. Бибкод : 1991Sci...253..754J . дои : 10.1126/science.253.5021.754 . ПМИД 17835491 .
- ^ Лонгрич, Северная Каролина; Бхуллар, Б.-А.С.; Готье, Ж.А. (2012). «Массовое вымирание ящериц и змей на рубеже мела и палеогена» . Труды Национальной академии наук . 109 (52): 21396–21401. Бибкод : 2012PNAS..10921396L . дои : 10.1073/pnas.1211526110 . ISSN 0027-8424 . ПМЦ 3535637 . ПМИД 23236177 .
- ^ Филд, диджей; Берковичи, А.; Берв, Дж.С.; Данн, Р.; Фастовский, Д.Э.; Лисон, ТР; Вайда, В.; Готье, JA (2018). «Ранняя эволюция современных птиц, структурированная глобальным вымиранием лесов в конце мелового периода» . Современная биология . 28 (11): 1825–1831. Бибкод : 2018CBio...28E1825F . дои : 10.1016/j.cub.2018.04.062 . ПМИД 29804807 . S2CID 44075214 .
- ↑ Перейти обратно: Перейти обратно: а б Ларсон, Д.В.; Браун, СМ; Эванс, округ Колумбия (2016). «Дентальное неравенство и экологическая стабильность птицеподобных динозавров до массового вымирания в конце мелового периода» . Современная биология . 26 (10): 1325–1333. Бибкод : 2016CBio...26.1325L . дои : 10.1016/j.cub.2016.03.039 . ПМИД 27112293 . S2CID 3937001 .
- ↑ Перейти обратно: Перейти обратно: а б Ле Лефф, Ж. (2012). «Палеобиогеография и биоразнообразие позднемаастрихтских динозавров: сколько видов динозавров вымерло на рубеже мела и третичного периода?». Бюллетень геологического общества Франции . 183 (6): 547–559. дои : 10.2113/gssgfbull.183.6.547 . ISSN 0037-9409 .
- ^ Карпентер, К. (1983). «Свидетельства, свидетельствующие о постепенном вымирании последних меловых динозавров». Naturwissenschaften . 70 (12): 611–612. Бибкод : 1983NW.....70..611C . дои : 10.1007/BF00377404 . S2CID 20078285 .
- ^ Рассел, Д.А. (1984). «Постепенное вымирание динозавров — факт или заблуждение?». Природа . 307 (5949): 360–361. Бибкод : 1984Natur.307..360R . дои : 10.1038/307360a0 . S2CID 4269426 .
- ^ Фастовский, Д.Э.; Хуанг, Ю.; Сюй, Дж.; Мартин-МакНотон, Дж.; Шиэн, премьер-министр; Вейшампель, Д.Б. (2004). «Форма мезозойского богатства динозавров» (PDF) . Геология . 32 (10): 877–880. Бибкод : 2004Geo....32..877F . дои : 10.1130/G20695.1 .
- ^ Салливан, РМ (2006). «Форма мезозойского богатства динозавров: переоценка» . В Лукасе, СГ; Салливан, Р.М. (ред.). Позднемеловые позвоночные Западного Внутреннего региона . Бюллетень Музея естественной истории и науки Нью-Мексико. Том. 35. С. 403–405.
- ^ Кьяренца, А.А.; Мэннион, Полиция; Лант, диджей; Фарнсворт, А.; Джонс, Луизиана; Келланд, С.Дж.; Эллисон, Пенсильвания (2019). «Моделирование экологической ниши не подтверждает климатическое сокращение разнообразия динозавров до массового вымирания в меловом и палеогеновом периодах» . Природные коммуникации . 10 (1): 1–14. Бибкод : 2019NatCo..10.1091C . дои : 10.1038/s41467-019-08997-2 . ПМК 6403247 . ПМИД 30842410 .
- ^ Ллойд, GT (2012). «Усовершенствованный подход к моделированию для оценки влияния отбора проб на кривые палеобиоразнообразия: новое подтверждение снижения богатства динозавров мелового периода» . Письма по биологии . 8 (1): 123–126. дои : 10.1098/rsbl.2011.0210 . ПМК 3259943 . ПМИД 21508029 . S2CID 1376734 .
- ^ Сакамото, М.; Бентон, MJ; Вендитти, К. (2016). «Динозавры приходят в упадок за десятки миллионов лет до их окончательного вымирания» . Труды Национальной академии наук . 113 (18): 5036–5040. Бибкод : 2016PNAS..113.5036S . дои : 10.1073/pnas.1521478113 . ПМЦ 4983840 . ПМИД 27092007 .
- ^ Барретт, премьер-министр; Макгоуэн, Эй Джей; Пейдж, В. (2009). «Разнообразие динозавров и рок-альбом» . Труды Королевского общества B: Биологические науки . 276 (1667): 2667–2674. дои : 10.1098/rspb.2009.0352 . ПМК 2686664 . ПМИД 19403535 .
- ^ Апчерч, П.; Мэннион, Полиция; Бенсон, РБ; Батлер, Р.Дж.; Каррано, Монтана (2011). «Геологический и антропогенный контроль над отбором образцов наземных окаменелостей: тематическое исследование динозавров». Геологическое общество, Лондон, специальные публикации . 358 (1): 209–240. Бибкод : 2011ГСЛСП.358..209У . дои : 10.1144/SP358.14 . S2CID 130777837 .
- ^ Рэндалл 2015
- ^ Альварес, LW ; Альварес, В .; Асаро, Ф .; Мишель, Х.В. (1980). «Внеземная причина мел-третичного вымирания» (PDF) . Наука . 208 (4448): 1095–1108. Бибкод : 1980Sci...208.1095A . CiteSeerX 10.1.1.126.8496 . дои : 10.1126/science.208.4448.1095 . ISSN 0036-8075 . ПМИД 17783054 . S2CID 16017767 . Архивировано из оригинала (PDF) 8 июля 2010 года . Проверено 30 октября 2019 г.
- ^ Бохор, Б.Ф.; Модрески, П.Дж.; Фуд, Э.Э. (1987). «Потрясенный кварц в пограничных глинах мелового и третичного периодов: свидетельства глобального распространения» . Наука . 236 (4802): 705–709. Бибкод : 1987Sci...236..705B . дои : 10.1126/science.236.4802.705 . ПМИД 17748309 . S2CID 31383614 .
- ^ Хильдебранд, Арканзас; Пенфилд, GT; Кринг, Д.А.; Пилкингтон, М.; Камарго, ЗА; Якобсен, С.Б.; Бойнтон, Западная Вирджиния (1991). «Кратер Чиксулуб: возможный ударный кратер на границе мелового и третичного периодов на полуострове Юкатан, Мексика». Геология . 19 (9): 867–871. Бибкод : 1991Geo....19..867H . doi : 10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2 .
- ^ Поуп, КО ; Окампо, AC ; Кинсленд, GL; и др. (1996). «Поверхностное выражение кратера Чиксулуб». Геология . 24 (6). Боулдер, Колорадо: Геологическое общество Америки: 527–530. Бибкод : 1996Geo....24..527P . doi : 10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2 . ISSN 0091-7613 . ПМИД 11539331 .
- ^ Шульте, П.; Алегрет, Л.; Аренильяс, И.; Арз, Дж.А.; Бартон, ПиДжей; Баун, PR; Бралоуэр, Ти Джей; Кристесон, GL; Клейс, П.; Кокелл, CS; Коллинз, Дж.С.; Дойч, А.; Голдин, Ти Джей; Гото, К.; Грахалес-Нишимура, Дж. М.; Грив, Королевские ВВС; Гулик, СПС; Джонсон, КР; Кисслинг, В.; Кеберл, К.; Кринг, Д.А.; Маклауд, КГ; Мацуи, Т.; Мелош, Дж.; Монтанари, А.; Морган, СП; Нил, ЧР; Николс, диджей; Норрис, РД; Пьераццо, Э.; Равицца, Г.; Реболледо-Виейра, М.; Уве Реймолд, В.; Робин, Э.; Салдж, Т.; Шпейер, Р.П.; Сладкий, АР; Уррутиа-Фукугаучи, Дж.; Вайда, В.; Уэлен, Монтана; Виллумсен, PS (2010). «Удар астероида Чиксулуб и массовое вымирание на границе мела и палеогена» . Наука . 327 (5970): 1214–1218. Бибкод : 2010Sci...327.1214S . дои : 10.1126/science.1177265 . ПМИД 20203042 . S2CID 2659741 .
- ^ Кринг, Д.А. (2007). «Ударное событие Чиксулуб и его экологические последствия на границе мелового и третичного периодов». Палеогеография, Палеоклиматология, Палеоэкология . 255 (1–2): 4–21. Бибкод : 2007PPP...255....4K . дои : 10.1016/j.palaeo.2007.02.037 .
- ↑ Перейти обратно: Перейти обратно: а б Кьяренца, А.А.; Фарнсворт, А.; Мэннион, Полиция; Лант, диджей; Вальдес, П.Дж.; Морган, СП ; Эллисон, Пенсильвания (2020). «Вымирание динозавров в конце мелового периода было вызвано ударом астероида, а не вулканизмом» . Труды Национальной академии наук . 117 (29): 17084–17093. Бибкод : 2020PNAS..11717084C . дои : 10.1073/pnas.2006087117 . ПМЦ 7382232 . ПМИД 32601204 .
- ^ Иванов, Б.А. (2005). «Численное моделирование крупнейших земных метеоритных кратеров». Исследования Солнечной системы . 39 (5): 381–409. Бибкод : 2005SoSyR..39..381I . дои : 10.1007/s11208-005-0051-0 . S2CID 120305483 .
- ^ Мацуи, Т.; Имамура, Ф.; Таджика, Э.; Накано, Ю.; Фудзисава, Ю. (2002). «Генерация и распространение цунами в результате ударного события мел-третичного периода». Специальные статьи Геологического общества Америки . 356 : 69–78. дои : 10.1130/0-8137-2356-6.69 . ISBN 978-0-8137-2356-3 .
- ^ Робертсон, Д.С.; Маккенна, MC ; Тун, Одесса ; и др. (2004). «Выживание в первые часы кайнозоя» (PDF) . Бюллетень Геологического общества Америки . 116 (5–6): 760–768. Бибкод : 2004GSAB..116..760R . дои : 10.1130/B25402.1 . ISSN 0016-7606 . Архивировано из оригинала (PDF) 18 сентября 2012 года . Проверено 15 июня 2011 г.
- ^ Робертсон, Д.С.; Льюис, В.М.; Шиэн, премьер-министр; Тун, О.Б. (2013). «Вымирание K-Pg: переоценка гипотезы теплового пожара» . Журнал геофизических исследований: Биогеонауки . 118 (1): 329–336. Бибкод : 2013JGRG..118..329R . дои : 10.1002/jgrg.20018 . S2CID 17015462 .
- ^ Поуп, КО; Бейнс, К.Х.; Окампо, AC; Иванов, Б.А. (1997). «Энергия, нестабильное производство и климатические последствия воздействия Чиксулуб мелового / третичного периода» . Журнал геофизических исследований: Планеты . 102 (Е9): 21645–21664. Бибкод : 1997JGR...10221645P . дои : 10.1029/97JE01743 . ПМИД 11541145 . S2CID 8447773 .
- ↑ Перейти обратно: Перейти обратно: а б Оно, С.; Кадоно, Т.; Куросава, К.; Хамура, Т.; Сакайя, Т.; Сигэмори, К.; Хиронака, Ю.; Сано, Т.; Ватари, Т.; Отани, К.; Мацуи, Т.; Сугита, С. (2014). «Производство паров, богатых сульфатами, во время воздействия Чиксулуб и последствия для закисления океана». Природа Геонауки . 7 (4): 279–282. Бибкод : 2014NatGe...7..279O . дои : 10.1038/ngeo2095 .
- ^ Кайхо, К.; Осима, Н.; Адачи, К.; Адачи, Ю.; Мизуками, Т.; Фудзибаяси, М.; Сайто, Р. (2016). «Глобальное изменение климата, вызванное сажей на границе K-Pg, как причина массового вымирания» . Научные отчеты . 6 (1): 1–13. Бибкод : 2016НатСР...628427К . дои : 10.1038/srep28427 . ПМЦ 4944614 . ПМИД 27414998 .
- ^ Лайонс, СЛ; Карп, АТ; Бралоуэр, Ти Джей; Грайс, К.; Шефер, Б.; Гулик, ИП; Морган, СП ; Фриман, КХ (2020). «Органические вещества из кратера Чиксулуб усугубили ударную зиму K – Pg» . Труды Национальной академии наук . 117 (41): 25327–25334. Бибкод : 2020PNAS..11725327L . дои : 10.1073/pnas.2004596117 . ПМЦ 7568312 . ПМИД 32989138 .
- ^ Шене, Алабама; Куртильо, В.; Флюто, Ф.; Жерар, М.; Кидельёр, X.; Хадри, СФР; Суббарао, КВ; Тордарсон, Т. (2009). «Определение быстрых извержений Декана на границе мелового и третичного периодов с использованием палеомагнитных вековых вариаций: 2. Ограничения, связанные с анализом восьми новых разрезов и синтезом составного разреза толщиной 3500 м» (PDF) . Журнал геофизических исследований: Solid Earth . 114 (Б6): В06103. Бибкод : 2009JGRB..114.6103C . дои : 10.1029/2008JB005644 . S2CID 140541003 .
- ^ Шене, Б.; Эдди, член парламента; Сампертон, КМ; Келлер, CB; Келлер, Г.; Адатте, Т.; Хадри, Сан-Франциско (2019). «U-Pb ограничения на импульсное извержение Деканских траппов во время массового вымирания в конце мелового периода» . Наука . 363 (6429): 862–866. Бибкод : 2019Sci...363..862S . дои : 10.1126/science.aau2422 . ОСТИ 1497969 . ПМИД 30792300 . S2CID 67876950 .
- ↑ Перейти обратно: Перейти обратно: а б Маклин, DM (1985). «Дегазация мантии ловушек Декана в условиях конечного вымирания морской среды в меловой период». Меловые исследования . 6 (3): 235–259. Бибкод : 1985CrRes...6..235M . дои : 10.1016/0195-6671(85)90048-5 .
- ^ Селф, С.; Виддоусон, М.; Тордарсон, Т.; Джей, А.Е. (2006). «Неустойчивые потоки во время паводковых извержений базальтов и потенциальное воздействие на глобальную окружающую среду: взгляд на Декан». Письма о Земле и планетологии . 248 (1–2): 518–532. Бибкод : 2006E&PSL.248..518S . дои : 10.1016/j.epsl.2006.05.041 .
- ^ Тобин, Т.С.; Битц, СМ; Арчер, Д. (2017). «Моделирование климатических последствий выбросов углекислого газа в результате извержений вулканов Деканских ловушек на границе мела и палеогена». Палеогеография, Палеоклиматология, Палеоэкология . 478 : 139–148. Бибкод : 2017PPP...478..139T . дои : 10.1016/j.palaeo.2016.05.028 .
- ^ Шмидт, А.; Скеффингтон, РА; Тордарсон, Т.; Селф, С.; Форстер, премьер-министр; Рэп, А.; Риджвелл, А.; Фаулер, Д.; Уилсон, М.; Манн, ГВ; Виналл, ПБ; Карслав, Канзас (2016). «Выборочный экологический стресс из-за серы, выбрасываемой в результате континентальных паводковых извержений базальтов» (PDF) . Природа Геонауки . 9 (1): 77–82. Бибкод : 2016NatGe...9...77S . дои : 10.1038/ngeo2588 . S2CID 59518452 . Архивировано (PDF) из оригинала 22 сентября 2017 г.
- ^ Хоффман Дж.; Феро, Ж.; Куртильо, В. (2000). " 40 с/ 39 Датирование отдельных минералов и целых пород из лавовой груды Западных Гат: дополнительные ограничения на продолжительность и возраст ловушек Декана». Earth and Planetary Science Letters . 180 (1–2): 13–27. Бибкод : 2000E&PSL.180. ..13H .дои - : 10.1016/S0012-821X(00)00159 X ISSN 0012-821X .
- ^ Сахни, А. (1988). «Пограничные события мелового и третичного периодов: массовые вымирания, обогащение иридием и деканский вулканизм». Современная наука . 57 (10): 513–519. JSTOR 24090754 .
- ^ Глэсби, врач общей практики; Кунцендорф, Х. (1996). «Множественные факторы происхождения границы мелового и третичного периодов: роль экологического стресса и вулканизма Деканской ловушки». Геологическое Рундшау . 85 (2): 191–210. Бибкод : 1996IJEaS..85..191G . дои : 10.1007/BF02422228 . ПМИД 11543126 . S2CID 19155384 .
- ^ Альварес, ЛВ (1987). Массовые вымирания, вызванные ударами крупных болидов (Отчет). Лаборатория Лоуренса Беркли . п. 39. ЛБЛ-22786 . Проверено 27 января 2021 г.
- ^ Альварес 1997 , стр. 130–146 , гл. 7: «Мир после Чиксулуба».
- ^ Ренне, PR; Растяжение, CJ; Ричардс, Массачусетс; Селф, С.; Вандерклюйсен, Л.; Панде, К. (2015). «Сдвиг состояния деканского вулканизма на границе мела и палеогена, возможно, вызванный ударом» . Наука . 350 (6256): 76–78. Бибкод : 2015Sci...350...76R . doi : 10.1126/science.aac7549 . ПМИД 26430116 . S2CID 30612906 .
- ^ Ричардс, Массачусетс; Альварес, В.; Селф, С.; Карлстрем, Л.; Ренне, PR; Манга, М.; Растяжение, CJ; Смит, Дж.; Вандерклюйсен, Л.; Гибсон, SA (2015). «Вызов крупнейших извержений Декана в результате удара Чиксулуб» . Бюллетень Геологического общества Америки . 127 (11–12): 1507–1520. Бибкод : 2015GSAB..127.1507R . дои : 10.1130/B31167.1 . S2CID 3463018 .
- ^ Хазинс В.; Шувалов, В. (2019). «Воздействие Чиксулуб как триггер одной из фаз деканского вулканизма: порог плотности сейсмической энергии». Кочарян Г.; Ляхов А. (ред.). Триггерные эффекты в геосистемах . Спрингерские труды по наукам о Земле и окружающей среде. Чам: Спрингер. стр. 523–530. дои : 10.1007/978-3-030-31970-0_55 . ISBN 978-3-030-31969-4 . S2CID 210277965 .
- ^ Арчибальд, доктор медицинских наук; Клеменс, Вашингтон; Падиан, К.; Роу, Т.; Маклауд, Н.; Барретт, премьер-министр; Гейл, А.; Холройд, П.; Сьюс, Х.-Д.; Аренс, Северная Каролина; Хорнер, младший; Уилсон, врач общей практики; Гудвин, МБ; Брошу, Калифорния; Лофгрен, Д.Л.; Херлберт, Ш.; Хартман, Дж. Х.; Эберт, Д.А.; Виналл, ПБ; Карри, Пи Джей; Вейль, А.; Прасад, GVR; Дингус, Л.; Куртильо, В.; Милнер, А.; Милнер, А.; Баджпай, С.; Уорд, диджей; Сахни, А. (2010). «Меловые вымирания: несколько причин». Наука . 328 (5981): 973, ответ автора 975–6. дои : 10.1126/science.328.5981.973-a . ПМИД 20489004 .
- ^ Куртильо, В.; Флюто, Ф. (2010). «Меловое вымирание: вулканическая гипотеза». Наука . 328 (5981): 973–974. дои : 10.1126/science.328.5981.973-b . ПМИД 20489003 .
- ^ Келлер, Г. (2014). «Декканский вулканизм, воздействие Чиксулуб и массовое вымирание в конце мелового периода: совпадение? Причина и следствие». Специальные статьи Геологического общества Америки . 505 : 57–89. дои : 10.1130/2014.2505(03) . ISBN 978-0-8137-2505-5 .
- ^ Шульте, П.; Алегрет, Л.; Аренильяс, И.; Арз, Дж.А.; Бартон, ПиДжей; Баун, PR; Бралоуэр, Ти Джей; Кристесон, GL; Клейс, П.; Кокелл, CS; Коллинз, Дж.С.; Дойч, А.; Голдин, Ти Джей; Гото, К.; Грахалес-Нишимура, Дж. М.; Грив, Королевские ВВС; Гулик, СПС; Джонсон, КР; Кисслинг, В.; Кеберл, К.; Кринг, Д.А.; Маклауд, КГ; Мацуи, Т.; Мелош, Дж.; Монтанари, А.; Морган, СП ; Нил, ЧР; Николс, диджей; Норрис, РД; Пьераццо, Э.; Равицца, Г.; Реболледо-Виейра, М.; Уве Реймолд, В.; Робин, Э.; Салдж, Т.; Шпейер, Р.П.; Сладкий, АР; Уррутиа-Фукугаучи, Дж.; Вайда, В.; Уэлен, Монтана; Виллумсен, PS (2010). «Ответ — меловое вымирание». Наука . 328 (5981): 975–976. дои : 10.1126/science.328.5981.975 .
- ^ Фассетт, Дж. Э.; Химан, LM; Симонетти, А. (2011). «Прямое U – Pb датирование костей динозавров мелового и палеоценового периода, бассейн Сан-Хуан, Нью-Мексико». Геология . 39 (2): 159–162. Бибкод : 2011Geo....39..159F . дои : 10.1130/G31466.1 . ISSN 0091-7613 .
- ^ Фассетт, Дж. Э.; Химан, LM; Симонетти, А. (2009). «Новые геохронологические и стратиграфические данные подтверждают палеоценовый возраст песчаника Охо Аламо, содержащего динозавров, и формации Анимас в бассейне Сан-Хуан, Нью-Мексико и Колорадо» . Электронная палеонтология . 12 (1):3А.
- ^ Слоан, RE; Ригби, Дж. К. младший; Ван Вален, Луизиана ; и др. (1986). «Постепенное вымирание динозавров и одновременная радиация копытных в формации Хелл-Крик». Наука . 232 (4750): 629–633. Бибкод : 1986Sci...232..629S . дои : 10.1126/science.232.4750.629 . ISSN 0036-8075 . ПМИД 17781415 . S2CID 31638639 .
- ^ Лукас, СГ; Салливан, РМ; Кэтэр, С.М.; Ясински, SE; Фаулер, Д.В.; Хекерт, AB; Шпильманн, Дж.А.; Хант, AP (2009). «Нет окончательных свидетельств существования динозавров палеоцена в бассейне Сан-Хуан». Электронная палеонтология . 12 (2):8А.
- ^ Ренне, PR; Гудвин, МБ (2012). «Прямое U-Pb датирование костей динозавров мелового и палеоценового периода, бассейн Сан-Хуан, Нью-Мексико: КОММЕНТАРИЙ» . Геология . 40 (4): е259. Бибкод : 2012Geo....40E.259R . дои : 10.1130/G32521C.1 .
- ^ Лофгрен, Д.Л.; Хоттон, CL; Рункель, AC (1990). «Переработка меловых динозавров в палеоценовый канал, отложения, верхняя часть формации Хелл-Крик, Монтана». Геология . 18 (9): 874–877. Бибкод : 1990Geo....18..874L . doi : 10.1130/0091-7613(1990)018<0874:ROCDIP>2.3.CO;2 .
- ^ Кениг, А.Е.; Лукас, СГ; Неймарк, Луизиана; Хекерт, AB; Салливан, РМ; Ясински, SE; Фаулер, Д.В. (2012). «Прямое U-Pb датирование костей динозавров мелового и палеоценового периода, бассейн Сан-Хуан, Нью-Мексико: КОММЕНТАРИЙ» . Геология . 40 (4): е262. Бибкод : 2012Geo....40E.262K . дои : 10.1130/G32154C.1 .
- ^ «Динозавр» . Словарь Merriam-Webster.com . Проверено 7 ноября 2019 г.
- ^ Сарджант 1995 , стр. 255–284, гл. 15: «Динозавры и диномания за 150 лет», Хью С. Торренс .
- ^ Карри и Падиан 1997 , стр. 347–350, «История открытий динозавров: первый золотой период», Брент Х. Брайтаупт.
- ^ Диккенс 1853 , с. 1 , гл. Я: « Лондон. Недавно закончился Михайловский срок, и лорд-канцлер сидит в Линкольнс-Инн-холле. Неумолимая ноябрьская погода. Столько грязи на улицах, как будто воды только что сошли с лица земли и больше не уйдут. было бы чудесно встретить мегалозавра длиной около сорока футов, ковыляющего, как слоновья ящерица, по холму Холборн » .
- ^ Фарлоу и Бретт-Сурман 1997 , стр. 675–697 , гл. 43: «Динозавры и средства массовой информации» Дональда Ф. Глата и депутата Бретта-Сурмана.
- ^ Ли, Ньютон; Мадей, Кристина (2012). «Ранняя анимация: приколы и ситуации». Диснеевские истории . стр. 17–24. дои : 10.1007/978-1-4614-2101-6_3 . ISBN 978-1-4614-2100-9 . S2CID 192335675 .
Библиография
- Альварес, Уолтер (1997). Ти-рекс и Кратер Судьбы . Принстон, Нью-Джерси : Издательство Принстонского университета . ISBN 978-0-691-01630-6 . LCCN 96049208 . OCLC 1007846558 . Проверено 4 ноября 2019 г.
- Баккер, Роберт Т. (1986). Ереси динозавров: новые теории, раскрывающие тайну динозавров и их исчезновения . Нью-Йорк : Уильям Морроу и компания . ISBN 978-0-688-04287-5 . LCCN 86012643 . OCLC 13699558 . Проверено 6 ноября 2019 г.
- Бентон, Майкл Дж. (2005). Палеонтология позвоночных (3-е изд.). Молден, Массачусетс : Blackwell Publishing . ISBN 978-0-632-05637-8 . LCCN 2003028152 . OCLC 53970617 . Проверено 30 октября 2019 г.
- Брусатте, Стивен Л. (2012). Бентон, Майкл Дж. (ред.). Палеобиология динозавров . Темы палеобиологии. Предисловие Майкла Дж. Бентона. Хобокен, Нью-Джерси : Уайли-Блэквелл . Бибкод : 2012dipa.book.....B . дои : 10.1002/9781118274071 . ISBN 978-0-470-65658-7 . LCCN 2011050466 . OCLC 781864955 .
- Кьяппе, Луис М .; Уитмер, Лоуренс М. , ред. (2002). Мезозойские птицы: над головами динозавров . Беркли: Издательство Калифорнийского университета. ISBN 978-0-520-20094-4 . LCCN 2001044600 . OCLC 901747962 .
- Кольбер, Эдвин Х. (1971) [Первоначально опубликовано, Нью-Йорк: EP Dutton , 1968; Лондон : Evans Brothers Ltd , 1969]. Люди и динозавры: поиски в полевых условиях и в лаборатории . Хармондсворт : Пингвин . ISBN 978-0-14-021288-4 . OCLC 16208760 . Проверено 31 октября 2019 г.
- Коуэн, Ричард (2005). История жизни (4-е изд.). Молден, Массачусетс: Blackwell Publishing. ISBN 978-1-4051-1756-2 . LCCN 2003027993 . OCLC 53970577 . Пятое издание книги доступно в Интернет-архиве . Проверено 19 октября 2019 г.
- Карри, Филип Дж .; Падиан, Кевин , ред. (1997). Энциклопедия динозавров . Сан-Диего, Калифорния : Academic Press . ISBN 978-0-12-226810-6 . LCCN 97023430 . OCLC 436848919 . Проверено 30 октября 2019 г.
- Карри, Филип Дж.; Коппельхус, Ева Б.; Шугар, Мартин А.; Райт, Джоанна Л., ред. (2004). Пернатые драконы: исследования перехода от динозавров к птицам . Жизнь прошлого. Блумингтон, Индиана : Издательство Университета Индианы . ISBN 978-0-253-34373-4 . LCCN 2003019035 . OCLC 52942941 .
- Карри Роджерс, Кристина А .; Уилсон, Джеффри А. , ред. (2005). Зауроподы: эволюция и палеобиология . Беркли : Издательство Калифорнийского университета . ISBN 978-0-520-24623-2 . LCCN 2005010624 . OCLC 879179542 .
- Десмонд, Адриан Дж . (1975). Горячие динозавры: революция в палеонтологии . Лондон: Блондинка и Бриггс. ISBN 978-0-8037-3755-6 . LCCN 76359907 . ОЛ 4933052М . Проверено 30 октября 2019 г.
- Диккенс, Чарльз (1853). Холодный дом . Лондон: Брэдбери и Эванс . Проверено 7 ноября 2019 г.
- Додсон, Питер ; Джинджерич, Филип Д. , ред. (1993). Функциональная морфология и эволюция . Американский научный журнал . Том. 293А. Нью-Хейвен, Коннектикут : Геологическая лаборатория Клайна, Йельский университет . ISSN 0002-9599 . ОСЛК 27781160 .
- Донг, Чжимин (1992). Динозавровая фауна Китая (английское изд.). Пекин ; Берлин ; Нью-Йорк: China Ocean Press ; Спрингер-Верлаг . ISBN 978-3-540-52084-9 . LCCN 92207835 . OCLC 26522845 .
- Дайк, Гарет; Кайзер, Гэри, ред. (2011). Живые динозавры: эволюционная история современных птиц . Чичестер ; Хобокен, Нью-Джерси: Уайли-Блэквелл. ISBN 978-0-470-65666-2 . LCCN 2010043277 . OCLC 729724640 .
- Фарлоу, Джеймс О.; Бретт-Сурман, депутат Кнессета, ред. (1997). Полный динозавр . Блумингтон, Индиана: Издательство Университета Индианы. ISBN 978-0-253-33349-0 . LCCN 97-23698 . OCLC 924985811 . Проверено 14 октября 2019 г.
- Фостер, Джон Р .; Лукас, Спенсер Г. , ред. (2006). «Палеонтология и геология верхнеюрской формации Моррисон» . Бюллетень Музея естественной истории и науки Нью-Мексико . Бюллетень Музея естественной истории и науки Нью-Мексико. 36 . Альбукерке, Нью-Мексико : Музей естественной истории и науки Нью-Мексико . ISSN 1524-4156 . OCLC 77520577 . Проверено 21 октября 2019 г.
- Перенасыщение, Дональд Ф. (1997). Динозавры: Энциклопедия . Предисловие Майкла К. Бретта-Сурмана. Джефферсон, Северная Каролина : McFarland & Company . ISBN 978-0-89950-917-4 . LCCN 95047668 . OCLC 33665881 .
- Гюнтер, Роберт Теодор , изд. (1968) [Впервые напечатано в Оксфорде в 1945 году]. Жизнь и письма Эдварда Лвида . Ранняя наука в Оксфорде. Том. XIV. Предисловие Альберта Эверарда Гюнтера (переиздание). Лондон: Доусонс из Пэлл-Мэлл. ISBN 978-0-7129-0292-2 . LCCN 22005926 . OCLC 43529321 . Проверено 4 ноября 2019 г.
- Ханселл, Майк (2000). Птичьи гнезда и строительное поведение . Иллюстрация пером и тушью Райта Оверхилла. Кембридж : Издательство Кембриджского университета. ISBN 978-0-521-46038-5 . LCCN 99087681 . OCLC 876286627 . Проверено 30 октября 2019 г.
- Хайльманн, Герхард (1926). Происхождение птиц . Лондон; Нью-Йорк: HF & G. Witherby ; Д. Эпплтон и компания . LCCN 27001127 . OCLC 606021642 .
- Холмс, Том (1998). Ископаемая вражда: соперничество первых американских охотников на динозавров . Парсиппани, Нью-Джерси : Джулиан Месснер . ISBN 978-0-382-39149-1 . LCCN 96013610 . OCLC 34472600 .
- Хольц, Томас Р. младший (2007). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов . Иллюстрировано Луисом В. Рей . Нью-Йорк: Рэндом Хаус . ISBN 978-0-375-82419-7 . LCCN 2006102491 . OCLC 77486015 . Проверено 22 октября 2019 г.
- Ламберт, Дэвид; Группа диаграмм (1990). Книга данных о динозаврах: полная, полностью иллюстрированная энциклопедия динозавров . Нью-Йорк: Книги Эйвона . ISBN 978-0-380-75896-8 . LCCN 89092487 . OCLC 21833417 . Проверено 14 октября 2019 г.
- Лессем, Дон ; Перенасыщение, Дональд Ф. (1993). Энциклопедия динозавров Общества динозавров . Иллюстрации Трейси Ли Форд; научные консультанты Питер Додсон и др. Нью-Йорк: Рэндом Хаус. ISBN 978-0-679-41770-5 . LCCN 94117716 . OCLC 30361459 . Проверено 30 октября 2019 г.
- Люйд, Эдвард (1699). Lithophylacii Britannici ichnographia [ Британские фигурные камни ]. Лондон: Ex Officina MC Архивировано из оригинала 14 августа 2020 г. Проверено 4 ноября 2019 г.
- Майр, Джеральд (2009). Палеогеновые ископаемые птицы . Берлин : Springer-Verlag. дои : 10.1007/978-3-540-89628-9 . ISBN 978-3-540-89627-2 . LCCN 2008940962 . OCLC 916182693 . S2CID 88941254 . Проверено 30 октября 2019 г.
- Норелл, Марк ; Гаффни, Юджин С .; Дингус, Лоуэлл (2000) [Первоначально опубликовано как «Открытие динозавров» в Американском музее естественной истории . Нью-Йорк: Кнопф , 1995]. Открытие динозавров: эволюция, вымирание и уроки предыстории (пересмотренное издание). Беркли: Издательство Калифорнийского университета. ISBN 978-0-520-22501-5 . LCCN 99053335 . OCLC 977125867 . Проверено 30 октября 2019 г.
- Ольшевский, Георгий (2000). Аннотированный контрольный список видов динозавров по континенту . Мезозойские извилины. Том. 3. Иллюстрировано Трейси Ли Форд. Сан-Диего, Калифорния : Публикации, требующие исследования. ISSN 0271-9428 . LCCN 00708700 . OCLC 44433611 .
- Оуэн, Ричард (1842). «Отчет о британских ископаемых рептилиях. Часть II» . Отчет одиннадцатого собрания Британской ассоциации содействия развитию науки; Состоялось в Плимуте в июле 1841 года . Лондон : Джон Мюррей . стр. 60 –204. ISBN 978-0-8201-1526-9 . LCCN 99030427 . OCLC 1015526268 . Проверено 13 октября 2019 г.
- Падиан, Кевин, изд. (1986). Происхождение птиц и эволюция полета . Мемуары Калифорнийской академии наук. Том. 8. Сан-Франциско, Калифорния: Калифорнийская академия наук . ISBN 978-0-940228-14-6 . OCLC 946083441 . ОЛ 9826926М .
- Парсонс, Кейт М. (2001). Рисование Левиафана: динозавры и научные войны . Жизнь в прошлом. Блумингтон, Индиана: Издательство Университета Индианы. ISBN 978-0-253-33937-9 . LCCN 2001016803 . OCLC 50174737 . Проверено 30 октября 2019 г.
- Пол, Грегори С. (1988). Хищные динозавры мира: полное иллюстрированное руководство . Нью-Йорк: Саймон и Шустер . ISBN 978-0-671-61946-6 . LCCN 88023052 . ОСЛК 859819093 . Проверено 30 октября 2019 г.
- Пол, Грегори С., изд. (2000). Научно-американская книга о динозаврах (1-е изд.). Нью-Йорк: Пресса Святого Мартина . ISBN 978-0-312-26226-6 . LCCN 2001269051 . OCLC 45256074 .
- Пол, Грегори С. (2010). Принстонский полевой справочник по динозаврам . Принстонские полевые гиды. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-13720-9 . LCCN 2010014916 . OCLC 907619291 .
- Сюжет, Роберт (1677). Естественная история Оксфордшира: очерк естественной истории Англии . Оксфорд; Лондон: С. Миллерс. LCCN 11004267 . OCLC 933062622 . Проверено 13 ноября 2019 г.
- Рэндалл, Лиза (2015). Темная материя и динозавры: поразительная взаимосвязанность Вселенной . Нью-Йорк: HarperCollins : Ecco. ISBN 978-0-06-232847-2 . LCCN 2016427646 . OCLC 962371431 .
- Рупке, Николаас А. (1994). Ричард Оуэн: викторианский натуралист . Нью-Хейвен: Издательство Йельского университета . ISBN 978-0-300-05820-8 . LCCN 93005739 . OCLC 844183804 . Проверено 5 ноября 2019 г.
- Сарджант, Уильям А.С. , изд. (1995). Окаменелости позвоночных и эволюция научных концепций: сочинения некоторых из его многочисленных друзей, посвященные Беверли Холстед . Современная геология, том. 18. Амстердам: Издательство Гордон и Брич. ISBN 978-2-88124-996-9 . ISSN 0026-7775 . LCCN 00500382 . OCLC 34672546 . «Перепечатка статей, опубликованных в специальном томе журнала «Современная геология» [т. 18 (памятный том Холстеда), 1993 г.], с пятью дополнительными статьями. - Преф.»
- Таннер, Лоуренс Х.; Спилманн, Джастин А.; Лукас, Спенсер Г., ред. (2013). «Триасовая система: новые достижения в стратиграфии и палеонтологии» . Бюллетень Музея естественной истории и науки Нью-Мексико . Бюллетень Музея естественной истории и науки Нью-Мексико. 61 . Альбукерке, Нью-Мексико: Музей естественной истории и науки Нью-Мексико. ISSN 1524-4156 . OCLC 852432407 . Проверено 21 октября 2019 г.
- Вейшампель, Дэвид Б .; Додсон, Питер; Осмольска, Гальшка , ред. (2004). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. ISBN 978-0-520-25408-4 . LCCN 2004049804 . OCLC 154697781 .
Дальнейшее чтение
- Университет Саутгемптона (29 сентября 2021 г.). «Два новых вида крупных хищных динозавров с крокодилоподобными черепами обнаружены на острове Уайт» . СайТехДейли .
- Чжоу, Чжунхэ (октябрь 2004 г.). «Происхождение и ранняя эволюция птиц: открытия, споры и перспективы на основе ископаемых свидетельств» (PDF) . Naturwissenschaften . 91 (10). Берлин: Springer Science+Business Media: 455–471. Бибкод : 2004NW.....91..455Z . дои : 10.1007/s00114-004-0570-4 . ISSN 0028-1042 . ПМИД 15365634 . S2CID 3329625 . Архивировано из оригинала (PDF) 21 июля 2011 года . Проверено 6 ноября 2019 г.
- Пол, Грегори С. (2002). Воздушные динозавры: эволюция и потеря полета у динозавров и птиц . Балтимор ; Лондон: Издательство Университета Джонса Хопкинса . ISBN 978-0-8018-6763-7 . LCCN 2001000242 . OCLC 1088130487 . .
- Стюарт, Табори и Чанг (1997). Огромная книга динозавров . Нью-Йорк: Стюарт, Табори и Чанг . ISBN 978-1-55670-596-0 . LCCN 97000398 . OCLC 1037269801 .
- Штернберг, Чарльз Мортрам (1966) [Оригинальное издание опубликовано Э. Клотье, типографом короля, 1946]. Канадские динозавры . Геологическая серия. Том. 54 (2-е изд.). Оттава : Национальный музей Канады . LCCN gs46000214 . OCLC 1032865683 .



Списки динозавров по континентам |
|---|
